Abstract
Pancreatic ductal adenocarcinoma (PDAC) is one of humans’ most common and fatal neoplasms. Nowadays, a number of PDAC studies are being conducted in two different fields: non-coding RNA (especially microRNA and long non-coding RNA) and microbiota. It has been recently discovered that not only does miRNA affect particular bacteria in the gut microbiome that can promote carcinogenesis in the pancreas, but the microbiome also has a visible impact on the miRNA. This suggests that it is possible to use the combined impact of the microbiome and noncoding RNA to suppress the development of PDAC. Nevertheless, insufficient research has focused on bounding both approaches to the diagnosis, treatment, and prevention of pancreatic ductal adenocarcinoma. In this article, we summarize the recent literature on the molecular basis of carcinogenesis in the pancreas, the two-sided impact of particular types of non-coding RNA and the pancreatic cancer microbiome, and possible medical implications of the discovered phenomenon.
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of the most common and fatal neoplasms in humans [1]. It is estimated that PDAC will be the second leading cause of cancer death in the United States of America by 2030 [2]. The high mortality of PDAC is due to its late detection caused by a lack of sufficient and specific biomarkers and resistance to treatment [3]. The overall survival rate of PDAC is 9% [4,5]. The vast majority of patients are not suitable for surgical treatment and undergo chemotherapy and palliative therapy instead.
According to Wang et al., the human microbiota consists of microbes colonizing different sites of the human body as well as its ecosystem [6]. The term microbiome refers to the genetic information encoded by microbiota and its host. Microbiota consists of bacteria, fungi, viruses, and archaea. It has been discovered that the pancreas has its own microbiota, which is more abundant in a cancerous pancreas compared to a healthy pancreas [7,8]. The microbiota of a cancerous pancreas consists mostly of Proteobacteria, Bacteroides, and Firmicutes and is different from the one in a healthy pancreas [9]. Recent studies have shown that the microbiome of the pancreas and other organs contributes to shaping the course and progression of PDAC [1]. The pancreatic microbiome is partially predestined to treatment response and impacts the tumor environment [10].
MicroRNA (miRNA) and long non-coding RNA (lncRNA) are subtypes of non-coding RNA, which are functional RNA molecules without protein-coding abilities. miRNA consists of approximately 22 nucleotides, processed from larger hairpin precursors. As Mahesh et al. has stated, approximately 60% of all protein-coding genes are regulated by miRNAs with multiple targets per miRNA type [5]. LncRNA are longer than 200 nucleotides [11]. A significant impact of non-coding RNA on PDAC progression but also in repression via different pathways has already been discovered [12]. Both miRNA and lncRNA affect the carcinogenesis of PDAC by binding to different genes and one another, and thus modulate the production of amino acids of different metabolic pathways. They regulate numerous actions of carcinogenesis such as proliferation, cell differentiation, cell apoptosis, migration, and the creation of metastases [13,14].
The modern work of Wang et al. suggests a bi-directional influence of miRNA and bacteria in the gut microbiome on one another, which can promote carcinogenesis in the pancreas, but the microbiome also has a visible impact on the miRNA [7]. This implies that it is possible to use a combined impact of the microbiome and noncoding RNA to suppress the development of PDAC.
In this article, we summarize the recent literature on the molecular basis of carcinogenesis in the pancreas, the impact of particular types of non-coding RNA and the pancreatic cancer microbiome, and possible medical implications of the discovered phenomenon.
2. Evolution of Pancreatic Adenocarcinoma—What Occurs in Cells before Cancer?
Several risk factors predispose to the development of PDAC, including smoking, obesity, diabetes, chronic pancreatitis, and bacterial or viral infections (Helicobacter pylori, Hepatitis B Virus, Hepatitis C Virus) [3]. Another important factor is the acquisition of many genetic mutations, such as KRAS (85–100% of pancreatic cancer cases) and mutations of suppressor genes such as p16 (CDKN2A), TP53, SMAD, and mutations of damage repair genes like hMLH1 [6]. The accumulation of genetic mutations over time combined with exposure to environmental risk factors or familial predisposition promotes cellular damage and the development of precancerous lesions of PDAC. The most important risk factors are shown in Figure 1.
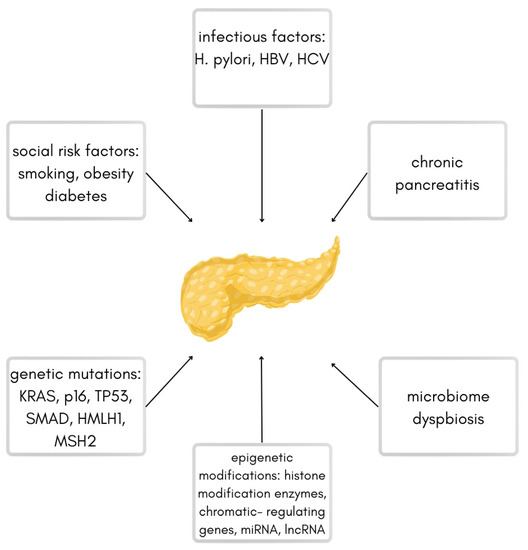
Figure 1.
Scheme of different risk factors for pancreatic ductal adenocarcinoma.
The precursor pre-cancer state of PDAC is pancreatic intraepithelial neoplasia (PanIN), whilst states predisposing to cancer are intraductal papillary mucinous neoplasms (IPMNs) and mucinous cystic neoplasms (MCNs).
The graph above shows the most common risk factors leading to pancreatic ductal adenocarcinoma (based on the work by Ryan et al.) [3].
3. Genetic Alterations in PanIN, MCN, and IPMNs
The early benign lesions preceding PDAC are pancreatic intraepithelial neoplasia (PanIN), intraductal papillary mucinous neoplasm (IPMN), and mucinous cystic neoplasm (MCN). PanINs are microscopic changes, and MCN and IPMN are macroscopic lesions visible in imaging diagnostics [15].
The PanINs can be classified into three grades, PanIN-I, -II, and –III, based on their cytology, architecture, and nuclear characteristics, with grade I being the least advanced and grade III being referred to as “carcinoma in situ” [16]. One of the earliest genetic changes in PanIN is loss of telomeric integrity, which accounts for a significant majority of these type of lesions [17]. A mutation present in both mucinous lesions is Kirsten Rat Sarcoma (KRAS), detected in 90% of invasive PanINs [18] and the vast majority of IPMN [15,19,20]. The KRAS mutation causes impaired GTPase activity of KRAS2 products, which results in a constitutively active protein that is involved in the signaling transduction. Another frequent mutation in PanIN is tumor suppressor gene CDK2/INK4A, causing loss of p16, which inhibits the G1-S cell transition. The mechanism of these genetic alterations is due to various intragenic mutations, deletions, and promoter methylation [21,22]. In grades 2 and 3 of PanIN, mutations of TP53 (causing impaired ability to respond to stress in physiological processes) [15,23] and tumor suppressor gene BRCA2 [24], as well as the mutation of MADH4/SMAD/DPC4 (responsible for creating tumor suppressor protein Dpc4, causing insufficient gene expression regulation) [25,26], are present. Additionally, it has been reported that in PanIN, as well as in invasive adenocarcinomas, carcinogenetic proteins created by mutated genes are overexpressed. Some of these are components of epidermal growth factors, Notch [27], and Hedgehog [28] signaling pathways.
Intraductal papillary mucinous neoplasm (IPMN) is a lesion that produces mucin and arises from major or secondary pancreatic ducts [29]. The most common mutations in this lesion are KRAS, GNAS, and RNF43 [30]. The MADH4/SMAD/DPC4 mutation also appears in IPMN, yet only in the most advanced lesions [15]. Another common characteristic of IPMN is the expression of apomucins such as MUC1, MUC2, and MUC3 [31]. Other genetic alterations found in IPMN are the STK11/LKB1 mutation (perceived in one-third of cases, which results in the impaired production of serine/threonine protein kinase, also present in patients with Peutz–Jeghers Syndrome [32]). The mutation of GNAS occurs in around 4% of IPMNs. It is a secondary mutation activated by the mutated KRAS gene and affects the subnormal Gαs protein by activating the adenylyl cyclase cascade of G-protein-coupled receptors, therefore responding to hormones and other extracellular signals) [3,15,33]. An amount of 15% of IPMN has the RNF43 mutation, impairing the antagonist of Wnt signaling [3].
Mucinous Cystic Neoplasms (MCNs) are mucin-producing neoplasms with distinctive ovarian-type stroma, present in the epithelium of the body and tail of the pancreas. The female gender predisposes to this neoplasm as over 90% of patients suffering from it are middle-aged women [34]. The MCNs can be classified based on histological architecture into three stages: mild MCN—mucinous cystadenomas, MCN with moderate dysplasia (MCN borderline), and MCN with carcinoma in situ [34]. Analogically to PanIN, one of the earliest molecular changes in MCN development is the KRAS mutation, present in sparse lesions of the mild stage, yet in up to all high-grade dysplasias [35]. Moreover, the advanced lesions are characterized by the inactivation of TP53 and MADH4/MAD4/DPC4 and mutation of RNF43 [16,36].
Table 1 compares the most common mutations in three early benign lesions proceeding to PDAC (Chr. stands for chromosome). It is based on the work of Singh et al. 2007 [15].

Table 1.
Genetic mutation in PanIN (pancreatic intraepithelial neoplasia), MCN (mucinous cystic neoplasm), and IPMN (intraductal papillary mucinous neoplasm).
4. Genetic Alterations in PDAC and Their Consequences
PDAC is characterized by severe metabolic stress caused by hypovascularization, leading to extreme hypoxia and limitations in nutrition.
All of the factors causing metabolic stress and therefore limitations in nutrition create conditions for genetic mutations in cancer cells. As the gene mutations accumulate, the precancerous lesions may turn into pancreatic adenocarcinoma. Genetic modifications in PDAC may be divided into epigenetic regulatory gene mutations, histone modification enzyme mutations, chromatic regulating gene and cell proliferation gene mutations, and epigenetic differentiation such as histone-modifying genes [6].
The most common and one of the most researched mutations is the KRAS mutation. It is present in over 90% of PDAC according to the CGD (CITIZEN-GENERATED DATA) Data Portal by the American National Institutes of Health. This mutation plays a significant role in cancer initiation and maintenance. Within the RAS pathway, it can activate the effector signaling cascades and transcription factors that are involved in cell proliferation, transformation, and metastasis. Activation of the RAS pathway promotes pro-inflammatory signaling by activating NF-kappa beta, which, in PDAC, is considered to moderate many aspects of cancer development and progression [37]. Tumors with KRAS have constitutively high levels of autophagy [38], which can lead to both the detoxification of the cell from damaged particles and the provision of metabolites for biosynthesis and energy production [3,6]. It has been discovered in both mouse models and clinical trials that the progression of PDAC is inhibited by genetic inhibition of autophagic processes or by chloroquine (as it inhibits lysosomal acidification) [38]. KRAS also alters the expression of enzymes involved in glucose utilization. PDAC demands a high level of glycolysis, instead of oxidative phosphorylation [39]. Mutant KRAS promotes mitochondrial translocation of phosphoglycerate kinase-1. That results in the production of phosphorylated pyruvate dehydrogenase kinase-1 and restricted oxidative phosphorylation (OXPHOS) [40]. The KRAS mutation is involved in most of the metabolic reprogramming pathways that occur in pancreatic cancer. Therefore, it is crucial to incorporate KRAS mutation silencing or repairing as a therapeutic target.
In addition to the KRAS mutation, PDAC is abundant in hundreds of different mutations and the ones characterized below are the most common. The SMAD family member 4 (SMAD4) mutation is caused by homozygous deletion or mutation. It lowers the amount of SMAD-4-dependent inhibition of transforming growth factor-beta (TGF-beta) and reduces the induction of TGF-beta signaling, which promotes pro-tumorigenic responses [41]. The cyclin-dependent kinase inhibitor 2A (CDKN2A), loss of p16, appears in around half of PDAC cases [6]. It may lead to the loss of regulation of the cyclin-dependent kinase (CDK) 4 and 6 cell cycle checkpoints, which cause the dysregulation of the cell cycle and subsequent carcinogenesis [42]. The inactivation of TP53 is detected in 50 to 74% of PDAC cases according to Wang et al. [6]. Combined with the KRAS mutation, it can promote metabolic changes before malignant transformations.
Other mutations found in less than 7% of PDAC cases according to the Harmonized Cancer Data Portal created by the National Institutes of Health are MUC16 (6,7%), ring-finger protein 47 (RFN43) (5,95%), GLI3 (5,59%), TGFBR (4,32%), and GNAS (3,91%). Another type of mutation detected in 10 to 25% of PDAC lesions is loss-of-function mutations encoding components of the SWI/SNF nucleosome remodeling complex [43].
The stem cell state is crucial in the development of PDAC. Cancer stem cells (CSCs) are cells of any state and purpose. Due to their plasticity, they can transform into the stem state at any time and renew infinitely [44]. This phenomenon explains cancer development and survival. The CSCs were first identified by the expression of CD44, CD24, and epithelial-specific antigen (ESA) [45]. It is known that CD133+/CXCR4+ cells (cell with both CD133 and CXCR4 antigens present, both being cancer stem cell markers) are responsible for metastasis development [46]. Other studies suggest that ALDH+ cells have CSC properties in pancreatic cancer [47]. ALDH+ cells are more tumorigenic than CD133+. Additionally, ALDH++/CD44+/CD24− cells are also highly tumorigenic [48].
Amidst all researched genes, FoxQ1 seemed to be vital for the aggressive biological properties of these cells. It knocks down EpCAM (epithelial cell adhesion molecule), which is overexpressed in malignant tumors [49]. Moreover, FOXQ1 restrains the Snail gene, which regulates the epithelial-to-mesenchymal transition of cells in cancer-stem-cell-like cells [50,51]. Taken together, it seems crucial to incorporate CSC-targeted treatment in the pancreatic treatment protocol [44]. A promising method to improve chemotherapy could be the double inhibition of the sonic hedgehog pathway and mTOR pathway as they significantly eradicate pancreatic CSCs and result in a long survival rate in both mice and human xenograft models [52,53].
Considering the general trend of focusing on high-throughput studies, more and more research using omics technologies emerge in the pre-cancer lesions and PDAC. As it can give a deeper understating of processes underlying the progression and development of PDAC [54,55,56], most of the omics studies focus on finding a perfectly sensitive biomarker of PDAC [57]. A novel work of Kobayashi et al. summed and compared the proposed biomarkers for PDAC, including germline mutations, miRNA, metabolites, and sugars [57]. Other works focus on finding the best way to differentiate and group subtypes of PDAC based on their histology and gene expression [58]. Yet, it is quintessential to realize that the majority of genomic analysis is performed on resected lesions, which is available only in around 15% of PDAC cases. [3]. Omics research sheds new light on the complexity of pancreatic cancer and its origin. Therefore, it is crucial to conduct omics research on the non-resective cases of pancreatic adenocarcinoma such as analyzing biopsy material.
5. Molecular Aspects of PDAC Metastases
Cancer metastases are formed by a detachment of cancer cells from the primary mass and transport via blood or lymphatic vessels. The majority of pancreatic metastases are localized in the liver (79–94%), peritoneum (41–56%), abdominal lymph nodes (41%), and the lungs (45–48%) [59,60,61,62]. According to Oldfield et al., all patients with pancreatic cancer have cells with metastatic potential in their primary lesion at the time of diagnosis [63]. The main difference between primary and metastatic tumor cells is the ability to proliferate, which is variable in the metastases as it is greatly influenced by the tumor environment [63].
Against popular belief that metastases occur only in the late [64,65] stage of cancer, in pancreatic cancer, they can appear even before the large mass formation of the primary lesion. It is mostly plausible that the step-by-step accumulation of different mutations and epigenetic alterations is the impulse for this process [66]. The so-far-discovered driving forces for metastases in pancreatic cancer are numerous non-coding RNAs [67,68], transcription factors (like Kruppel-like factor 4, KLF4 [69]), growth factors (like vascular endothelial growth factor, VEGF [70]), and oxygen conditions. MicroRNA is associated with a high risk of lymph node metastasis in patients with pancreatic cancer [71].
An experimentally reduced metastatic behavior of tumor has been observed by reducing the transcription of RUNX3 factor, which is overexpressed in KRAS and TP53 mutant mice (KPC mice). The floxed SMAD4 gene leads to rendering the mutational status of the TP53 allele in the above-described mice. Since the SMAD gene regulates the RUNX3 levels, the reduction in metastasis can be achieved in this way [72].
A possibly relevant way to understand and manage pancreatic cancer metastases is to explore the epithelial-to-mesenchymal (ETM) transition in PDAC. The EMT-transcription factor Zeb1 is responsible for enabling the colonization and plasticity of the phenotype of tumor cells and promotes PDAC metastasis [73]. Reducing Zeb1 suppresses the colonization of cancer and the phenotypic/metabolic plasticity of cancerous cells [74]. Therefore, high Zeb1 levels in PDAC correlate with poor diagnosis [74].
Another crucial aspect of PDAC metastasis is altered genes. Based on the organoid culture system, Roe et al. discovered that the metastatic process goes hand in hand with recurrent alterations in enhancer activation. Therefore, it adds to the likelihood of the transcription of genes responsible for PDAC activity and its aggressiveness [75]. Enhancers are DNA sequences that increase the transcription of particular genes [76]. The main driver of enhancer activation in PDAC metastasis is FOXA1. It makes the cells more invasive and more predestined to metastasize in vivo, and it makes the anchorage less dependent on in vitro growth [75].
Table 2 presents the most common genetic mutations in the particular stages of carcinogenesis in the pancreas causing PDAC and metastases, as well as characteristic mutations present in the pre-malignant lesions observed prior to PDAC.
Table 2.
Map of genetic alterations associated with various premalignant lesions and PDAC.
6. miRNA and lncRNA Mechanisms of Intervention in PDAC Progression
Both microRNA and long non-coding RNA belong to the family of non-coding RNA. They are transcripts without the function of coding proteins. They can be divided by size (short and long ncRNA), function (housekeeping RNA and regulatory RNA), direction of transcription (sense/antisense, bidirectional), as well as location (intronic and intergenic). Non-coding RNA may interact with a wide range of other molecules such as messenger RNA (mRNA) [11].
MicroRNA is an approximately 22-codon-long ribonucleic acid molecule created from larger hairpin precursors [70]. It is estimated that around 60% of all protein-coding genes are regulated by miRNAs, whereas one miRNA targets hundreds of genes [5].
Filipowicz et al. stated that miRNAs interact with the corresponding mRNA by base pairing, yet it pairs imperfectly. The miRNAs interact with mRNA mostly in the post-translational phases. The possible mechanisms of the miRNA-mediated post-translational impact on mRNA are repressing deacetylation (before decapping and degradation), repressing cap recognition or 60 s joining, and proteolysis of nascent peptide or blocking elongation by slowing it or dropping-off [70]. MicroRNA seems to have different functions in the cancer microenvironment. Some of these regulate the epigenetic processes by direct modulation of the enzymes taking part in methylation-mediated silencing and chromatic modeling or by epigenetically regulating themselves. MicroRNA also regulates RNA via exosomes, microvesicles, and protein complexes in a paracrine manner to influence the tumor microenvironment. MicroRNA also promotes the release of mediators in both pro- and anticarcinogenesis directions [77]. MicroRNA is classically perceived as either an oncogenic or tumor suppressor. Tumor suppressor means targeting oncogenic particles and molecules; therefore, the normal states combined with its function dictate the end phenotype [77].
Long non-coding RNAs (lncRNAs) are non-coding transcripts shorter or equal to over 200 nucleotides. They take a significant part in many molecular mechanisms like transcriptional and post-transcriptional regulation, processing different types of ncRNAs. LncRNA is responsible for the inactivation of X-chromosome and takes part in cell differentiation, immune responses, apoptosis, and T cells activation. In cancer, the role of lncRNA is complex and includes sustaining proliferative signaling, equivocating growth suppressors, enhancing replicative immortality, creating metastases, enhancing angiogenesis, resisting cell death, and reprogramming the energy metabolic paths [11]. LncRNA can be divided into prooncogenic and tumor-suppressing. Some examples of the prooncogenic long noncoding RNA are PCGEM1, HOTAIR, MALAT1, and CDK2B-ASI. Representative examples of the second group are GAS5, TERRA, and CCND1 [11].
In the table below (Table 3), we summarize the miRNA and lncRNA that bind to particular genes and influence their input in the progression and inhibition of pancreatic adenocarcinoma.
Table 3.
microRNA and long non-coding RNA and their genetic targets in PDAC.
Table 3 presents some of the researched miRNA and lncRNA that play a significant role in the PDAC, as well as its up- or down-regulation status and its pro- or anticarcinogenic characteristics.
From the above-stated theses, it is known that both miRNA and lncRNA interfere with mRNA, causing tumor enhancement or attenuation. It seems that more research is needed in the field of miRNA than lncRNA in PDAC and associated cancerous processes. Yet, we do not know all mechanisms behind those changes and ways to use them in order to stop cancer progression.
7. Cross-Talk between Chosen ncRNA Types and Microbiome in PDAC
The human microbiota is both a collection of microbes colonizing different sites of the human body and its ecosystem [153]. The intestinal tract microbiota is a combination of over 1014 microorganisms [153], whilst the microbiome is the genetic information encoded by microbiota (bacteria, fungi, viruses, and archaea) and its host [153].
The microbiome of a healthy pancreas differentiates from the microbiome in a cancerous pancreas (later referred to as “cancer microbiome”) with their composition and ratios of different species of bacteria and viruses [9]. Moreover, the “healthy” microbiota consists mostly of bacteria belonging to Firmicutes, Bacteroidetes, Actinetobacteria, and Proteobacteria [1], whereas the microbiota of a cancerous pancreas consists mostly of Proteobacteria, Bacteroides, and Firmicutes according to Pushalkar et al. [9]. It is known that mRNA can enter bacteria via endocytosis [154].
Figure 2 presents the schematic composition of pancreatic microbiota in eubiosis and a microbiome in the state of carcinogenesis. The most common species of bacteria from the most to the least numerous are listed. Additionally, it is important to mention that the number of viruses increases in the cancer microbiome. It is based on the works of Daniluk et al. 2021 and Gesualdo et al. 2020 [1,155].
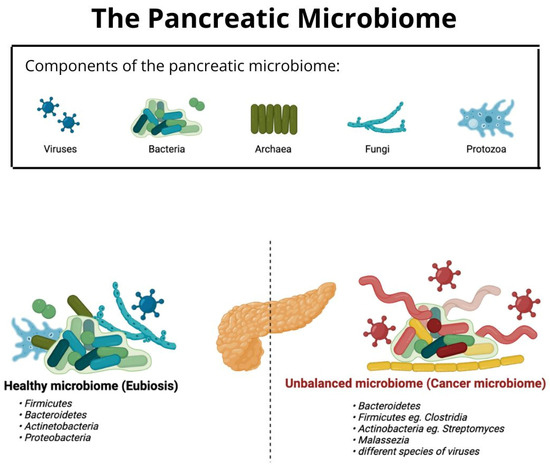
Figure 2.
Comparison of the composition of bacterial microbiota of a healthy and cancerous pancreas.
Riquelme et al. stated that the immune microenvironment and the immune responses are created by the crosstalk between a tumor and its microbiome [156]. Moreover, tumor-associated specific microbiota is expected to directly regulate cancer initiation, progression, and responses to chemo- or immune-therapies [157]. Microbiota can enhance tumorigenesis through epigenetic effects, regulation of miRNA expression, inflammation induction, DNA damage, and differential expression of driver genes [158].
The two strongest theories about its mechanism for altering cancer are that gut microbes release metabolites that affect cancer cells [7], and that the microbes suppress the immune system by creating inflammation [1]. Daniluk et al. found that the Gram-negative bacteria component, namely lipopolysaccharide, enhances the oncogenic signaling by promoting pancreatic inflammation [1]. It is possible through activation of Toll-like receptors, and increasing the tumor-promoting myeloid-derived suppressor cells (MDSC) as well as M2 macrophages [9]. Work by Yuan et al. sheds light on the miRNA regulating the microbiome; they discovered that the profusion of Akkermansia correlates with miRNA differentially expressed in samples with colorectal cancer and intestinal tissue around cancer [159].
The report of Wang et al. showed that miRNA and particular bacteria in the gut microbiome co-influence each other, which can be the cause of carcinogenesis in the pancreas [7]. Additionally, it suggested the possibility of using of the combined impact of the microbiome and noncoding RNA to suppress the development of PDAC.
Increased amounts of Herbaspirillum and Catenibacterium are correlated with NRAS and PT53 mutations. Moreover, a decreased amount of Barnesiella is associated with mutated RAS genes [160].
Fusobacterium nucleatum bacteria in colorectal cancer may be influenced by an elevated glycan production associated with miRNA [159]. This shows a potential mechanism of miRNAs to modify the microbial composition by regulating glucose metabolism [7]. Marin-Muller et al. found that MiR-198 acted as a crucial tumor suppressor that modulates the molecular composition of an essential interactome in pancreatic cancer [161]. Additionally, they explored a reciprocal regulatory loop between mesothelin and miR-198, which partially explains the MSLN (mesothelin)-gene-mediated pathogenesis [161]. Gironella et al. proved that in mouse xenograft models of pancreatic cancer (TP53INP1), a protein involved in the proapoptotic response upon p53 activation is down-regulated by miR-155 [120].
Shirazi et al. stated that H. pylori and P. gingivalis play a significant role in altering the cell cycle, damaging DNA, influencing the expression of miRNA, and affecting epigenetics in pancreatic cancer [162].
On the other hand, Guo et al. stated that host genetics generally influence microbiota in pancreatic cancer.
Pushalkar et al. shed new light on the influence of non-coding RNA on the microbiota by stating that the mutated KRAS gene may influence the diversity and composition of pancreatic and gut microbiota [9].
So far, the most researched area of the relationship between microbiota and miRNA in neoplasm is colorectal cancer [163]. Malmuthuge et al. suggested that microbiota-associated oncogenic miRNAs could be used as a way to treat cancers both locally and systemically by targeting microbial interventions [164]. Numerous researchers have informed about an urgent need for further research into the relationship between microbiota and non-coding RNA in pancreatic cancer as it may revolutionize the way we perceive oncogene regulation and creation of metastases [165,166].
Moreover, Sammallahti et at. stated that bacterial metabolites are expected to influence both the tumorigenic processes and cancer mutations themselves [158].
Additionally, the possible genetic toxicity and further interference with DNA are another possible mechanism that is used by microbiota to moderate DNA and epigenetic factors accelerating and influencing carcinogenesis. For example, reactive nitrogen species and other toxins are mediated by microbiota [167,168].
The above-mentioned studies suggest that the influence of microbiota on cancer mutations as well as epigenetics is more complex than we have thought.
Many studies have been conducted on the relationship between the microbiota and various cancers, but data on the effect of the microbiome on miRNAs in pancreatic cancer are still lacking. Sammallahti et al. suggested that stool represents a perfect material for researching microbiota and oncogenomic factors such as non-coding RNA, especially for seeking novel biomarkers of pancreatic ductal adenocarcinoma [158]. That opens a new field for further research.
8. Conclusions and Knowledge Gaps
The interactions between miRNA, lncRNA, and microbiota are a net of complex yet undefeated connections influencing carcinogenesis. Elinav et al. perceived a desperate need for studies of the microbiome both across the cancer continuum and across cancer types on a greater scale [8]. Therefore, further research in this field can provide us with early-stage diagnostics for pancreatic cancer, alternative targeted therapy options, and possibly a path to pancreatic cancer prevention. As the non-coding RNA field is being widely explored, the aspect of the cross-talk between miRNA and lncRNA and the pancreatic microbiome is still an open field for much-needed research.
Listed below are some of the key aspects of this area that require urgent research. Currently, we do not understand the mechanistic details of the function of miRNAs in repressing protein synthesis. The role of the other components from the microbiota (such as viruses, protozoa, and fungi) in the progression of PDAC and carcinogenesis remains unexplored [8].
Many studies were conducted in order to understand the influences of miRNA on the microbiome and vice versa on the colorectal cancer models, but not yet on pancreatic cancer.
Facing the complexity of the subject, many researchers may be discouraged to conduct experiments in this bidirectional approach, yet many studies suggest that this can lead to disenchanting the severity of PDAC. The co-influence of the microbiome and non-coding RNA creates opportunities to find new points for low-invasive targeted therapy, a gene-blocking approach to treating PDAC, and new biomarkers for diagnostic and treatment evaluation.
A promising application of the interaction of miRNAs and the microbiome would be to administer them orally, paving the way for future miRNA treatment in inflammation or cancer. As novel studies suggest, the implementation of the usage of the co-influence of ncRNA and the microbiome is not only possible but also easy to distribute. Therefore, it is tempting to say that the future will bring a safe and definite cure to one of the deadliest cancers—pancreatic cancer.
Author Contributions
Conceptualization, W.M.I. and J.D.; supervision, J.N; writing—original draft, W.M.I.; writing—review and editing, W.M.I., J.D. and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Daniluk, J.; Daniluk, U.; Rogalski, P.; Dabrowski, A.; Swidnicka-Siergiejko, A. Microbiome—Friend or Foe of Pancreatic Cancer? J. Clin. Med. 2021, 10, 5624. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic Adenocarcinoma. N. Engl. J. Med. 2014, 371, 1039–1049. [Google Scholar] [CrossRef]
- Watson, M.D.; Miller-Ocuin, J.L.; Driedger, M.R.; Beckman, M.J.; McKillop, I.H.; Baker, E.H.; Martinie, J.B.; Vrochides, D.; Iannitti, D.A.; Ocuin, L.M. Factors Associated with Treatment and Survival of Early Stage Pancreatic Cancer in the Era of Modern Chemotherapy: An Analysis of the National Cancer Database. J. Pancreat. Cancer 2020, 6, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, G.; Biswas, R. MicroRNA-155: A Master Regulator of Inflammation. J. Interf. Cytokine Res. 2019, 39, 321–330. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Yang, F.; Zhu, L.; Zhu, X.-Q.; Wang, Z.-F.; Wu, X.-L.; Zhou, C.-H.; Yan, J.-Y.; Hu, B.-Y.; et al. The molecular biology of pancreatic adenocarcinoma: Translational challenges and clinical perspectives. Signal Transduct. Target. Ther. 2021, 6, 249. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, H.; Dong, G.; Xu, L.; Jiang, F.; Mao, Q. Bi-direction effects between microbiome and MiRNAs in carcinogenesis. J. Cancer Res. Clin. Oncol. 2021, 147, 1299–1305. [Google Scholar] [CrossRef]
- Elinav, E.; Garrett, W.S.; Trinchieri, G.; Wargo, J. The cancer microbiome. Nat. Rev. Cancer 2019, 19, 371–376. [Google Scholar] [CrossRef]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Li, J.J.; Zhu, M.; Kashyap, P.C.; Chia, N.; Tran, N.H.; McWilliams, R.R.; Bekaii-Saab, T.S.; Ma, W.W. The role of microbiome in pancreatic cancer. Cancer Metastasis Rev. 2021, 40, 777–789. [Google Scholar] [CrossRef]
- Kunej, T.; Obsteter, J.; Pogacar, Z.; Horvat, S.; Calin, G.A. The decalog of long non-coding RNA involvement in cancer diagnosis and monitoring. Crit. Rev. Clin. Lab. Sci. 2014, 51, 344–357. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Huang, S. Role of non-coding RNA in pancreatic cancer (Review). Oncol. Lett. 2019, 18, 3963–3973. [Google Scholar] [CrossRef]
- Yonemori, K.; Kurahara, H.; Maemura, K.; Natsugoe, S. MicroRNA in pancreatic cancer. J. Hum. Genet. 2016, 62, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhi, X.; Gao, Y.; Ta, N.; Jiang, H.; Zheng, J. LncRNAs in pancreatic cancer. Oncotarget 2016, 7, 57379–57390. [Google Scholar] [CrossRef]
- Singh, M.; Maitra, A. Precursor Lesions of Pancreatic Cancer: Molecular Pathology and Clinical Implications. Pancreatology 2007, 7, 9–19. [Google Scholar] [CrossRef]
- Distler, M.; Aust, D.; Weitz, J.; Pilarsky, C.; Grützmann, R. Precursor lesions for sporadic pancreatic cancer: PanIN, IPMN, and MCN. BioMed Res. Int. 2014, 2014, 474905. [Google Scholar] [CrossRef]
- Meeker, A.K.; De Marzo, A.M. Recent advances in telomere biology: Implications for human cancer. Curr. Opin. Oncol. 2004, 16, 32–38. [Google Scholar] [CrossRef]
- Moskaluk, C.; Hruban, R.; Kern, S. P16 and k-ras gene mutations in the intraductal precursors of human pancreatic adenocarci-noma. Cancer Res. 1997, 57, 2140–2143. [Google Scholar] [PubMed]
- Kaino, M.; Kondoh, S.; Okita, S.; Hatano, S.; Shiraishi, K.; Kaino, S.; Okita, K. Detection of K-ras and p53 gene mutations in pancreatic juice for the diagnosis of intraductal papillary mucinous tumors. Pancreas 1999, 18, 294–299. [Google Scholar] [CrossRef]
- Yoshizawa, K.; Nagai, H.; Sakurai, S.; Hironaka, M.; Morinaga, S.; Saitoh, K.; Fukayama, M. Clonality and K- ras mutation analyses of epithelia in intraductal papillary mucinous tumor and mucinous cystic tumor of the pancreas. Virchows Arch. 2002, 441, 437–443. [Google Scholar] [CrossRef]
- Caldas, C.; Hahn, S.A.; da Costa, L.T.; Redston, M.S.; Schutte, M.; Seymour, A.B.; Weinstein, C.L.; Hruban, R.H.; Yeo, C.J.; Kern, S.E. Frequent somatic mutations and homozygous deletions of the p16 (MTS1) gene in pancreatic adenocarcinoma. Nat. Genet. 1994, 8, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Schutte, M.; Hruban, R.H.; Geradts, J.; Maynard, R.; Hilgers, W.; Rabindran, S.K.; Moskaluk, C.A.; Hahn, S.; Schwarte-Waldhoff, I.; Schmiegel, W.; et al. Abrogation of the Rb/p16 tumor-suppressive pathway in virtually all pancreatic carcinomas. Cancer Res. 1997, 15, 126–3130. [Google Scholar]
- Olivier, M.; Hollstein, M.; Hainaut, P. TP53 Mutations in Human Cancers: Origins, Consequences, and Clinical Use. Cold Spring Harb. Perspect. Biol. 2009, 2, a001008. [Google Scholar] [CrossRef] [PubMed]
- Goggins, M.; Hruban, R.H.; Kern, S.E. BRCA2 is inactivated late in the development of pancreatic intraepithelial neoplasia: Evidence and implications. Am. J. Pathol. 2000, 156, 1767–1771. [Google Scholar] [CrossRef]
- Loss of Expression of Dpc4 in Pancreatic Intraepithelial Neoplasia: Evidence That DPC4 Inactivation Occurs Late in Neoplastic Progression. Available online: https://pubmed.ncbi.nlm.nih.gov/10766191/ (accessed on 8 September 2022).
- Montgomery, E.; Goggins, M.; Zhou, S.; Argani, P.; Wilentz, R.E.; Kaushal, M.; Booker, S.; Romans, K.; Bhargava, P.; Hruban, R.H.; et al. Nuclear localization of Dpc4 (Madh4, Smad4) in colorectal carcinomas and relation to mismatch repair/transforming growth factor-β receptor defects. Am. J. Pathol. 2001, 158, 537–542. [Google Scholar] [CrossRef]
- Rooman, I.; De Medts, N.; Baeyens, L.; Lardon, J.; De Breuck, S.; Heimberg, H.; Bouwens, L. Expression of the Notch Signaling Pathway and Effect on Exocrine Cell Proliferation in Adult Rat Pancreas. Am. J. Pathol. 2006, 169, 1206–1214. [Google Scholar] [CrossRef]
- Sheng, T.; Li, C.; Zhang, X.; Chi, S.; He, N.; Chen, K.; McCormick, F.; Gatalica, Z.; Xie, J. Activation of the hedgehog pathway in advanced prostate cancer. Mol. Cancer 2004, 3, 29. [Google Scholar] [CrossRef]
- Al-Refaie, W.; Choi, E.; Tseng, J.; Tamm, E.; Lee, J.; Evans, D.; Pisters, P. Intraductal Papillary Mucinous Neoplasms of the Pancreas. Med. Princ. Pract. 2006, 15, 245–252. [Google Scholar] [CrossRef]
- Fischer, C.G.; Wood, L.D. From somatic mutation to early detection: Insights from molecular characterization of pancreatic cancer precursor lesions. J. Pathol. 2018, 246, 395–404. [Google Scholar] [CrossRef]
- Adsay, N.V.; Merati, K.; Andea, A.; Sarkar, F.; Hruban, R.H.; Wilentz, R.E.; Goggins, M.; Iocobuzio-Donahue, C.; Longnecker, D.S.; Klimstra, D.S. The Dichotomy in the Preinvasive Neoplasia to Invasive Carcinoma Sequence in the Pancreas: Differential Expression of MUC1 and MUC2 Supports the Existence of Two Separate Pathways of Carcinogenesis. Mod. Pathol. 2002, 15, 1087–1095. [Google Scholar] [CrossRef]
- Launonen, V. Mutations in the human LKB1/STK11 gene. Hum. Mutat. 2005, 26, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Hollstein, P.E.; Shaw, R.J. GNAS shifts metabolism in pancreatic cancer. Nature 2018, 20, 740–741. [Google Scholar] [CrossRef]
- Naveed; Naveed, S.; Qari, H.; Banday, T.; Altaf, A.; Para, M. Mucinous Cystic Neoplasms of Pancreas. Gastroenterol. Res. 2014, 7, 44–50. [Google Scholar] [CrossRef]
- Fujikura, K.; Akita, M.; Abe-Suzuki, S.; Itoh, T.; Zen, Y. Mucinous cystic neoplasms of the liver and pancreas: Relationship between KRAS driver mutations and disease progression. Histopathology 2017, 71, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Matthaei, H.; Schulick, R.D.; Hruban, R.H.; Maitra, A. Cystic precursors to invasive pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 141–150. [Google Scholar] [CrossRef]
- Prabhu, L.; Mundade, R.; Korc, M.; Loehrer, P.J.; Lu, T. Critical role of NF-κB in pancreatic cancer. Oncotarget 2014, 5, 10969. [Google Scholar] [CrossRef]
- Yang, S.; Wang, X.; Contino, G.; Liesa, M.; Sahin, E.; Ying, H.; Bause, A.; Li, Y.; Stommel, J.M.; Dell’Antonio, G.; et al. Pancreatic cancers require autophagy for tumor growth. Genes Dev. 2011, 25, 717. [Google Scholar] [CrossRef]
- Ying, H.; Kimmelman, A.C.; Lyssiotis, C.A.; Hua, S.; Chu, G.C.; Fletcher-Sananikone, E.; Locasale, J.W.; Son, J.; Zhang, H.; Coloff, J.L.; et al. Oncogenic Kras maintains pancreatic tumors through regulation of anabolic glucose metabolism. Cell 2012, 149, 656–670. [Google Scholar] [CrossRef]
- Li, X.; Jiang, Y.; Meisenhelder, J.; Yang, W.; Hawke, D.H.; Zheng, Y.; Xia, Y.; Aldape, K.; He, J.; Hunter, T.; et al. Mitochondria-Translocated PGK1 Functions as a Protein Kinase to Coordinate Glycolysis and the TCA Cycle in Tumorigenesis. Mol. Cell 2016, 61, 705–719. [Google Scholar] [CrossRef]
- Christenson, E.S.; Jaffee, E.; Azad, N.S. Current and emerging therapies for patients with advanced pancreatic ductal adenocarcinoma: A bright future. Lancet Oncol. 2020, 21, e135–e145. [Google Scholar] [CrossRef]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Biankin, A.V.; Waddell, N.; Kassahn, K.S.; Gingras, M.-C.; Muthuswamy, L.B.; Johns, A.L.; Miller, D.K.; Wilson, P.J.; Patch, A.-M.; Wu, J.; et al. Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature 2012, 491, 399–405. [Google Scholar] [CrossRef]
- Hermann, P.C.; Sainz, B. Pancreatic cancer stem cells: A state or an entity? Semin. Cancer Biol. 2018, 53, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Heidt, D.G.; Dalerba, P.; Burant, C.F.; Zhang, L.; Adsay, V.; Wicha, M.; Clarke, M.F.; Simeone, D.M. Identification of pancreatic cancer stem cells. Cancer Res 2007, 67, 1030–1037. [Google Scholar] [CrossRef]
- Hutcheson, J.; Balaji, U.; Porembka, M.R.; Wachsmann, M.B.; McCue, P.A.; Knudsen, E.S.; Witkiewicz, A.K. Immunologic and metabolic features of pancreatic ductal adenocarcinoma define prognostic subtypes of disease. Clin. Cancer Res. 2016, 22, 3606–3617. [Google Scholar] [CrossRef]
- Hermann, P.C.; Huber, S.L.; Herrler, T.; Aicher, A.; Ellwart, J.W.; Guba, M.; Bruns, C.J.; Heeschen, C. Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer. Cell Stem Cell 2007, 1, 313–323. [Google Scholar] [CrossRef]
- Kim, M.P.; Fleming, J.B.; Wang, H.; Abbruzzese, J.L.; Choi, W.; Kopetz, S.; McConkey, D.J.; Evans, D.B.; Gallick, G.E. ALDH activity selectively defines an enhanced tumor-initiating cell population relative to cd133 expression in human pancreatic adenocarcinoma. PLoS ONE 2011, 6, e20636. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F.; Struck, L.; Tachezy, M.; Vashist, Y.; Wicklein, D.; Schumacher, U.; Izbicki, J.R.; Bockhorn, M. Serum EpCAM Expression in Pancreatic Cancer. Anticancer Res. 2014, 34, 4741–4746. [Google Scholar]
- Liu, M.; Hancock, S.E.; Sultani, G.; Wilkins, B.; Ding, E.; Osborne, B.; Quek, L.-E.; Turner, N. Snail-Overexpression Induces Epithelial-mesenchymal Transition and Metabolic Reprogramming in Human Pancreatic Ductal Adenocarcinoma and Non-tumorigenic Ductal Cells. J. Clin. Med. 2019, 8, 822. [Google Scholar] [CrossRef]
- Pelosi, E.; Castelli, G.; Testa, U. Pancreatic Cancer: Molecular Characterization, Clonal Evolution and Cancer Stem Cells. Biomedicines 2017, 5, 65. [Google Scholar] [CrossRef]
- Mueller, M.; Hermann, P.C.; Witthauer, J.; Rubio–Viqueira, B.; Leicht, S.F.; Huber, S.; Ellwart, J.W.; Mustafa, M.; Bartenstein, P.; D’Haese, J.G.; et al. Combined targeted treatment to eliminate tumorigenic cancer stem cells in human pancreatic cancer. Gastroenterology 2009, 137, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Hermann, P.C.; Trabulo, S.M.; Sainz, B., Jr.; Balic, A.; Garcia, E.; Hahn, S.A.; Vandana, M.; Sahoo, S.K.; Tunici, P.; Bakker, A.; et al. Multimodal Treatment Eliminates Cancer Stem Cells and Leads to Long-Term Survival in Primary Human Pancreatic Cancer Tissue Xenografts. PLoS ONE 2013, 8, e66371. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.; Fong, C.; Luthra, A.; Smith, S.A.; DiNatale, R.G.; Nandakumar, S.; Walch, H.; Chatila, W.K.; Madupuri, R.; Kundra, R.; et al. Genomic characterization of metastatic patterns from prospective clinical sequencing of 25,000 patients. Cell 2022, 185, 563–575.e11. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Huang, C.; Cui Zhou, D.; Hu, Y.; Lih, T.M.; Savage, S.R.; Krug, K.; Clark, D.J.; Schnaubelt, M.; Chen, L.; et al. Proteogenomic characterization of pancreatic ductal adenocarcinoma. Cell 2021, 184, 5031–5052.e26. [Google Scholar] [CrossRef]
- Fraunhoffer, N.A.; Abuelafia, A.M.; Bigonnet, M.; Gayet, O.; Roques, J.; Nicolle, R.; Lomberk, G.; Urrutia, R.; Dusetti, N.; Iovanna, J. Multi-omics data integration and modeling unravels new mechanisms for pancreatic cancer and improves prognostic prediction. NPJ Precis. Oncol. 2022, 6, 57. [Google Scholar] [CrossRef]
- Kobayashi, T.; Honda, K. Trends in biomarker discoveries for the early detection and risk stratification of pancreatic cancer using omics studies. Expert Rev. Mol. Diagn. 2019, 19, 651–654. [Google Scholar] [CrossRef]
- Connor, A.A.; Gallinger, S. Pancreatic cancer evolution and heterogeneity: Integrating omics and clinical data. Nat. Rev. Cancer 2021, 22, 131–142. [Google Scholar] [CrossRef]
- Iacobuzio-Donahue, C.A.; Fu, B.; Yachida, S.; Luo, M.; Abe, H.; Henderson, C.M.; Vilardell, F.; Wang, Z.; Keller, J.W.; Banerjee, P.; et al. DPC4 Gene Status of the Primary Carcinoma Correlates With Patterns of Failure in Patients With Pancreatic Cancer. J. Clin. Oncol. 2009, 27, 1806–1813. [Google Scholar] [CrossRef]
- Yachida, S.; White, C.M.; Naito, Y.; Zhong, Y.; Brosnan, J.A.; Macgregor-Das, A.M.; Morgan, R.A.; Saunders, T.; Laheru, D.A.; Herman, J.M.; et al. Clinical significance of the genetic landscape of pancreatic cancer and implications for identification of potential long-term survivors. Clin. Cancer Res. 2012, 18, 6339–6347. [Google Scholar] [CrossRef]
- Haeno, H.; Gonen, M.; Davis, M.B.; Herman, J.M.; Iacobuzio-Donahue, C.A.; Michor, F. Computational modeling of pancreatic cancer reveals kinetics of metastasis suggesting optimum treatment strategies. Cell 2012, 148, 362–375. [Google Scholar] [CrossRef]
- Le Large, T.; Bijlsma, M.; Kazemier, G.; van Laarhoven, H.; Giovannetti, E.; Jimenez, C. Key biological processes driving metastatic spread of pancreatic cancer as identified by multi-omics studies. Semin. Cancer Biol. 2017, 44, 153–169. [Google Scholar] [CrossRef]
- Oldfield, L.E.; Connor, A.A.; Gallinger, S. Molecular Events in the Natural History of Pancreatic Cancer. Trends Cancer 2017, 3, 336–346. [Google Scholar] [CrossRef]
- Hegde, S.; Krisnawan, V.E.; Herzog, B.H.; Zuo, C.; Breden, M.A.; Knolhoff, B.L.; Hogg, G.D.; Tang, J.P.; Baer, J.M.; Mpoy, C.; et al. Dendritic cell paucity leads to dysfunctional immune surveillance in pancreatic cancer. Cancer Cell 2020, 37, 289–307.e9. [Google Scholar] [CrossRef]
- Chartier, C.; Raval, J.; Axelrod, F.; Bond, C.; Cain, J.; Dee-Hoskins, C.; Ma, S.; Fischer, M.M.; Shah, J.; Wei, J.; et al. Therapeutic targeting of tumor-derived r-spondin attenuates b-catenin signaling and tumorigenesis in multiple cancer types. Cancer Res. 2016, 76, 713–723. [Google Scholar] [CrossRef]
- Stratton, M.R. Exploring the genomes of cancer cells: Progress and promise. Science 2011, 331, 1553–1558. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Nakano, K.; Obchoei, S.; Setoguchi, K.; Matsumoto, M.; Yamamoto, T.; Obika, S.; Shimada, K.; Hiraoka, N. Small Nucleolar Noncoding RNA SNORA23, Up-Regulated in Human Pancreatic Ductal Adenocarcinoma, Regulates Expression of Spectrin Repeat-Containing Nuclear Envelope 2 to Promote Growth and Metastasis of Xenograft Tumors in Mice. Gastroenterology 2017, 153, 292–306.e2. [Google Scholar] [CrossRef]
- Guo, X.; Zhou, Q.; Su, D.; Luo, Y.; Fu, Z.; Huang, L.; Li, Z.; Jiang, D.; Kong, Y.; Li, Z.; et al. Circular RNA circBFAR promotes the progression of pancreatic ductal adenocarcinoma via the miR-34b-5p/MET/Akt axis. Mol. Cancer 2020, 19, 83. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Cui, J.; Quan, M.; Xie, D.; Jia, Z.; Wei, D.; Wang, L.; Gao, Y.; Ma, Q.; Xie, K. The novel KLF4/MSI2 signaling pathway regulates growth and metastasis of pancreatic cancer. Clin. Cancer Res. 2017, 23, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Nishiwada, S.; Sho, M.; Banwait, J.K.; Yamamura, K.; Akahori, T.; Nakamura, K.; Baba, H.; Goel, A. A MicroRNA Signature Identifies Pancreatic Ductal Adenocarcinoma Patients at Risk for Lymph Node Metastases. Gastroenterology 2020, 159, 562–574. [Google Scholar] [CrossRef]
- Whittle, M.C.; Izeradjene, K.; Rani, P.G.; Feng, L.; Carlson, M.A.; DelGiorno, K.E.; Wood, L.D.; Goggins, M.; Hruban, R.H.; Chang, A.E.; et al. RUNX3 Controls a Metastatic Switch in Pancreatic Ductal Adenocarcinoma. Cell 2015, 161, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Yu, J.; Suenaga, M.; Fesharakizadeh, S.; Cho, C.; Macgregor-Das, A.; Siddiqui, A.; Witmer, P.D.; Tamura, K.; Song, T.J.; et al. Deleterious Germline Mutations in Patients With Apparently Sporadic Pancreatic Adenocarcinoma. J. Clin. Oncol. 2017, 35, 3382–3390. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.-T.; Yang, M.-H. Revisiting epithelial-mesenchymal transition in cancer metastasis: The connection between epithelial plasticity and stemness. Mol. Oncol. 2017, 11, 792–804. [Google Scholar] [CrossRef]
- Roe, J.-S.; Hwang, C.-I.; Somerville, T.D.; Milazzo, J.P.; Lee, E.J.; Da Silva, B.; Maiorino, L.; Tiriac, H.; Young, C.M.; Miyabayashi, K.; et al. Enhancer Reprogramming Promotes Pancreatic Cancer Metastasis. Cell 2017, 170, 875–888.e20. [Google Scholar] [CrossRef] [PubMed]
- Pennacchio, L.A.; Bickmore, W.; Dean, A.; Nobrega, M.A.; Bejerano, G. Enhancers: Five essential questions. Nat. Rev. Genet. 2013, 14, 288–295. [Google Scholar] [CrossRef]
- Rusek, A.M.; Abba, M.; Eljaszewicz, A.; Moniuszko, M.; Niklinski, J.; Allgayer, H. MicroRNA modulators of epigenetic regulation, the tumor microenvironment and the immune system in lung cancer. Mol. Cancer 2015, 14, 34. [Google Scholar] [CrossRef]
- Zhai, L.; Wu, R.; Han, W.; Zhang, Y.; Zhu, D. miR-127 enhances myogenic cell differentiation by targeting S1PR3. Cell Death Dis. 2017, 8, e2707. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, L.; Ma, R.; Gong, H.; Xu, P.; Wang, C. MicroRNA-127 is aberrantly downregulated and acted as a functional tumor suppressor in human pancreatic cancer. Tumor Biol. 2016, 37, 14249–14257. [Google Scholar] [CrossRef]
- Dong, J.; Zhao, Y.-P.; Zhou, L.; Zhang, T.-P.; Chen, G. Bcl-2 upregulation induced by miR-21 via a direct interaction is associated with apoptosis and chemoresistance in MIA PaCa-2 pancreatic cancer cells. Arch. Med. Res. 2011, 42, 8–14. [Google Scholar] [CrossRef]
- Song, W.; Li, Q.; Wang, L.; Wang, L. Modulation of FoxO1 expression by miR-21 to promote growth of pancreatic ductal adenocarcinoma. Cell. Physiol. Biochem. 2015, 35, 184–190. [Google Scholar] [CrossRef]
- Nagao, Y.; Hisaoka, M.; Matsuyama, A.; Kanemitsu, S.; Hamada, T.; Fukuyama, T.; Nakano, R.; Uchiyama, A.; Kawamoto, M.; Yamaguchi, K.; et al. Association of microRNA-21 expression with its targets, PDCD4 and TIMP3, in pancreatic ductal adenocarcinoma. Mod. Pathol. 2012, 25, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Mark, J.; Park, J.K.; Lee, E.J.; Esau, C.; Schmittgen, T.D. Antisense inhibition of microRNA-21 or -221 arrests cell cycle, induces apoptosis, and sensitizesthe effects of gemcitabine in pancreatic adenocarcinoma. Pancreas 2009, 39, 190–199. [Google Scholar]
- Sarkar, S.; Dubaybo, H.; Ali, S.; Goncalves, P.; Kollepara, S.L.; Sethi, S.; APhilip, P.; Li, Y. Down-regulation of miR-221 inhibits proliferation of pancreatic cancer cells through up-regulation of PTEN, p27kip1, p57kip2, and PUMA. Am. J. Cancer Res. 2013, 3, 465–477. [Google Scholar] [PubMed]
- Xu, Q.; Li, P.; Chen, X.; Zong, L.; Jiang, Z.; Nan, L.; Lei, J.; Duan, W.; Zhang, D.; Li, X.; et al. miR-221/222 induces pancreatic cancer progression through the regulation of matrix metalloproteinases. Oncotarget 2015, 6, 14153–14164. [Google Scholar] [CrossRef]
- Su, A.; He, S.; Tian, B.; Hu, W.; Zhang, Z. MicroRNA-221 mediates the effects of PDGF-BB on migration, proliferation, and the epithelial-mesenchymal transition in pancreatic cancer cells. PLoS ONE 2013, 8, e71309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Yang, Y.; Liu, J.; Song, Y.; Cao, Y.; Chen, X.; Yang, W.; Wang, F.; Gao, J.; et al. MicroRNA-222 Controls Human Pancreatic Cancer Cell Line Capan-2 Proliferation by P57 Targeting. J. Cancer 2015, 6, 1230–1235. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ohuchida, K.; Mizumoto, K.; Lin, C.; Yamaguchi, H.; Ohtsuka, T.; Sato, N.; Toma, H.; Nakamura, M.; Nagai, E.; Hashizume, M.; et al. MicroRNA-10a is overexpressed in human pancreatic cancer and involved in its invasiveness partially via suppression of the HOXA1 gene. Ann. Surg. Oncol. 2012, 19, 2394–2402. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, D.; Qi, H.; Gao, Q. Thioredoxin-interacting protein is a favored target of miR-125b, promoting metastasis and progression of pancreatic cancer via the HIF1α pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22782. [Google Scholar] [CrossRef]
- Long, L.-M.; Zhan, J.-K.; Wang, H.-Q.; Li, S.; Chen, Y.-Y.; Liu, Y.-S. The Clinical Significance of miR-34a in Pancreatic Ductal Carcinoma and Associated Molecular and Cellular Mechanisms. Pathobiology 2016, 84, 38–48. [Google Scholar] [CrossRef]
- Xie, F.; Li, C.; Zhang, X.; Peng, W.; Wen, T. MiR-143-3p suppresses tumorigenesis in pancreatic ductal adenocarcinoma by targeting KRAS. Biomed. Pharmacother. 2019, 119, 109424. [Google Scholar] [CrossRef]
- Kent, O.A.; Chivukula, R.R.; Mullendore, M.; Wentzel, E.A.; Feldmann, G.; Lee, K.H.; Liu, S.; Leach, S.D.; Maitra, A.; Mendell, J.T. Repression of the miR-143/145 cluster by oncogenic Ras initiates a tumor-promoting feed-forward pathway. Genes Dev. 2010, 24, 2754–2759. [Google Scholar] [CrossRef]
- Gao, S.; Wang, P.; Hua, Y.; Xi, H.; Meng, Z.; Liu, T.; Chen, Z.; Liu, L. ROR functions as a ceRNA to regulate Nanog expression by sponging miR-145 and predicts poor prognosis in pancreatic cancer. Oncotarget 2016, 7, 1608–1618. [Google Scholar] [CrossRef]
- Khan, S.; Ebeling, M.C.; Zaman, M.S.; Sikander, M.; Yallapu, M.M.; Chauhan, N.; Yacoubian, A.M.; Behrman, S.W.; Zafar, N.; Kumar, D.; et al. MicroRNA-145 targets MUC13 and suppresses growth and invasion of pancreatic cancer. Oncotarget 2014, 5, 7599–7609. [Google Scholar] [CrossRef]
- Deng, S.; Zhu, S.; Wang, B.; Li, X.; Liu, Y.; Qin, Q.; Gong, Q.; Niu, Y.; Xiang, C.; Chen, J.; et al. Chronic pancreatitis and pancreatic cancer demonstrate active epithelial–mesenchymal transition profile, regulated by miR-217-SIRT1 pathway. Cancer Lett. 2014, 355, 184–191. [Google Scholar] [CrossRef]
- Zhao, W.-G.; Yu, S.-N.; Lu, Z.-H.; Ma, Y.-H.; Gu, Y.-M.; Chen, J. The miR-217 microRNA functions as a potential tumor suppressor in pancreatic ductal adenocarcinoma by targeting KRAS. Carcinog. 2010, 31, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wang, B.; Liu, Y.; Zhang, J.-G.; Deng, S.-C.; Qin, Q.; Tian, K.; Li, X.; Zhu, S.; Niu, Y.; et al. miRNA-141, downregulated in pancreatic cancer, inhibits cell proliferation and invasion by directly targeting MAP4K4. Mol. Cancer Ther. 2013, 12, 2569–2580. [Google Scholar] [CrossRef]
- Zhu, Z.-M.; Xu, Y.-F.; Su, Q.-J.; Du, J.-D.; Tan, X.-L.; Tu, Y.-L.; Tan, J.-W.; Jiao, H.-B. Prognostic significance of microRNA-141 expression and its tumor suppressor function in human pancreatic ductal adenocarcinoma. Mol. Cell. Biochem. 2013, 388, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Q.; Xu, D.; Wang, Q.; An, Y.; DU, Q.; Zhang, J.; Zhu, Y.; Miao, Y. hsa-miR-141 downregulates TM4SF1 to inhibit pancreatic cancer cell invasion and migration. Int. J. Oncol. 2013, 44, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Q.; Fang, Y.; Deng, X.; Chen, H.; Jin, J.; Lu, X.; Peng, C.; Li, H.; Shen, B. The Interplay Between miR-148a and DNMT1 Might be Exploited for Pancreatic Cancer Therapy. Cancer Investig. 2015, 33, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, M.; Zang, W.; Chen, X.; Wang, Y.; Li, P.; Du, Y.; Zhao, G.; Li, L. MiR-148a regulates the growth and apoptosis in pancreatic cancer by targeting CCKBR and Bcl-2. Tumor Biol. 2013, 35, 837–844. [Google Scholar] [CrossRef]
- Liffers, S.-T.; Munding, J.B.; Vogt, M.; Kuhlmann, J.D.; Verdoodt, B.; Nambiar, S.; Maghnouj, A.; Mirmohammadsadegh, A.; Hahn, S.A.; Tannapfel, A. MicroRNA-148a is down-regulated in human pancreatic ductal adenocarcinomas and regulates cell survival by targeting CDC25B. Lab. Investig. 2011, 91, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Song, S.; He, S.; Zhu, X.; Zhang, Y.; Yi, B.; Zhang, B.; Qin, G.; Li, D. MicroRNA-375 targets PDK1 in pancreatic carcinoma and suppresses cell growth through the Akt signaling pathway. Int. J. Mol. Med. 2014, 33, 950–956. [Google Scholar] [CrossRef]
- Sun, W.; Lu, Y.; Hu, J.; Li, S.; Deng, S.; Li, M. MiR-29c inhibits cell growth, invasion, and migration of pancreatic cancer by targeting ITGB1. OncoTargets Ther. 2015, 9, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Li, J.; Chen, Z.; Li, X.; Zheng, S.; Yi, D.; Zhong, A.; Chen, J. miR-29c suppresses pancreatic cancer liver metastasis in an orthotopic implantation model in nude mice and affects survival in pancreatic cancer patients. Carcinog. 2015, 36, 676–684. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, J.-G.; Shi, Y.; Qin, Q.; Liu, Y.; Wang, B.; Tian, K.; Deng, S.-C.; Li, X.; Zhu, S.; et al. MiR-130b is a prognostic marker and inhibits cell proliferation and invasion in pancreatic cancer through targeting STAT3. PLoS ONE 2013, 8, e73803. [Google Scholar] [CrossRef]
- Radhakrishnan, P.; Mohr, A.M.; Grandgenett, P.M.; Steele, M.M.; Batra, S.K.; Hollingsworth, M.A. MicroRNA-200c modulates the expression of MUC4 and MUC16 by directly targeting their coding sequences in human pancreatic cancer. PLoS ONE 2013, 8, e73356. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ohuchida, K.; Mizumoto, K.; Sato, N.; Kayashima, T.; Fujita, H.; Nakata, K.; Tanaka, M. MicroRNA, hsa-miR-200c, is an independent prognostic factor in pancreatic cancer and its upregulation inhibits pancreatic cancer invasion but increases cell proliferation. Mol. Cancer 2010, 9, 169. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Tang, M. MicroRNA-216a inhibits pancreatic cancer by directly targeting Janus kinase 2. Oncol. Rep. 2014, 32, 2824–2830. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, H.; Lin, S.; Ba, M.; Cui, S. MicroRNA-216a enhances the radiosensitivity of pancreatic cancer cells by inhibiting beclin-1-mediated autophagy. Oncol. Rep. 2015, 34, 1557–1564. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Chen, H.; Li, H.-Y.; Chen, R.; He, L.; Yang, J.-L.; Xiao, L.-L.; Chen, J.-L. Long non-coding RNA small nucleolar RNA host gene 6 aggravates pancreatic cancer through upregulation of far upstream element binding protein 1 by sponging microRNA-26a-5p. Chin. Med. J. 2020, 133, 1211–1220. [Google Scholar] [CrossRef]
- Deng, J.; He, M.; Chen, L.; Chen, C.; Zheng, J.; Cai, Z. The Loss of miR-26a-Mediated Post-Transcriptional Regulation of Cyclin E2 in Pancreatic Cancer Cell Proliferation and Decreased Patient Survival. PLoS ONE 2013, 8, e76450. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhang, J.-G.; Liu, Y.; Qin, Q.; Wang, B.; Tian, K.; Liu, L.; Li, X.; Niu, Y.; Deng, S.-C.; et al. miR-148b functions as a tumor suppressor in pancreatic cancer by targeting AMPKα1. Mol. Cancer Ther. 2013, 12, 83–93. [Google Scholar] [CrossRef]
- Gao, L.; Yang, Y.; Xu, H.; Liu, R.; Li, D.; Hong, H.; Qin, M.; Wang, Y. miR-335 functions as a tumor suppressor in pancreatic cancer by targeting OCT4. Tumor Biol. 2014, 35, 8309–8318. [Google Scholar] [CrossRef]
- Hamada, S.; Masamune, A.; Miura, S.; Satoh, K.; Shimosegawa, T. MiR-365 induces gemcitabine resistance in pancreatic cancer cells by targeting the adaptor protein SHC1 and pro-apoptotic regulator BAX. Cell. Signal. 2014, 26, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhu, C.-F.; Ma, M.-Z.; Chen, G.; Song, M.; Zeng, Z.-L.; Lu, W.-H.; Yang, J.; Wen, S.; Chiao, P.J.; et al. Micro-RNA-155 is induced by K-Ras oncogenic signal and promotes ROS stress in pancreatic cancer. Oncotarget 2015, 6, 21148–21158. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, J.; Wang, J.; Amos, C.; Killary, A.M.; Sen, S.; Wei, C.; Frazier, M.L. Putative tumor suppressor gene SEL1L was downregulated by aberrantly upregulated hsa-mir-155 in human pancreatic ductal adenocarcinoma. Mol. Carcinog. 2013, 53, 711–721. [Google Scholar] [CrossRef]
- Liu, W.-J.; Zhao, Y.-P.; Zhang, T.-P.; Zhou, L.; Cui, Q.-C.; Zhou, W.-X.; You, L.; Chen, G.; Shu, H. MLH1 as a direct target of MiR-155 and a potential predictor of favorable prognosis in pancreatic cancer. J. Gastrointest. Surg. 2013, 17, 1399–1405. [Google Scholar] [CrossRef]
- Huang, C.; Li, H.; Wu, W.; Jiang, T.; Qiu, Z. Regulation of miR-155 affects pancreatic cancer cell invasiveness and migration by modulating the STAT3 signaling pathway through SOCS1. Oncol. Rep. 2013, 30, 1223–1230. [Google Scholar] [CrossRef]
- Gironella, M.; Seux, M.; Xie, M.-J.; Cano, C.; Tomasini, R.; Gommeaux, J.; Garcia, S.; Nowak, J.; Yeung, M.L.; Jeang, K.-T.; et al. Tumor protein 53-induced nuclear protein 1 expression is repressed by miR-155, and its restoration inhibits pancreatic tumor development. Proc. Natl. Acad. Sci. USA 2007, 104, 16170–16175. [Google Scholar] [CrossRef]
- Liu, N.; Sun, Y.-Y.; Zhang, X.-W.; Chen, S.; Wang, Y.; Zhang, Z.-X.; Song, S.-W.; Qiu, G.-B.; Fu, W.-N. Oncogenic miR-23a in Pancreatic Ductal Adenocarcinogenesis Via Inhibiting APAF1. Dig. Dis. Sci. 2015, 60, 2000–2008. [Google Scholar] [CrossRef]
- Listing, H.; Mardin, W.A.; Wohlfromm, S.; Mees, S.T.; Haier, J. MiR-23a/-24-induced gene silencing results in mesothelial cell integration of pancreatic cancer. Br. J. Cancer 2014, 112, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ou, Y.; Wu, K.; Chen, Y.; Sun, W. miR-143 inhibits the metastasis of pancreatic cancer and an associated signaling pathway. Tumor Biol. 2012, 33, 1863–1870. [Google Scholar] [CrossRef]
- Li, Y.; VandenBoom, T.; Wang, Z.; Kong, D.; Ali, S.; Philip, P.A.; Sarkar, F.H. Abstract 5703: Up-regulation of miR-146a contributes to the inhibition of invasion of pancreatic cancer cells. Cancer Res 2010, 70 (Suppl. 8), 5703. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.K.; Bhardwaj, A.; Singh, S.; Arora, S.; Bin Wang, B.; Grizzle, W.E.; Singh, A.P. MicroRNA-150 directly targets MUC4 and suppresses growth and malignant behavior of pancreatic cancer cells. Carcinogenesis 2011, 32, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, D.; Wang, Q.; Zheng, D.; Jiang, X.; Xu, L. LPS induced miR-181a promotes pancreatic cancer cell migration via targeting PTEN and MAP2K4. Dig. Dis. Sci. 2014, 59, 1452–1460. [Google Scholar] [CrossRef]
- Zhang, P.; Guo, Z.; Hu, R.; He, X.; Jiao, X.; Zhu, X. Interaction between microRNA-181a and TNFAIP1 regulates pancreatic cancer proliferation and migration. Tumor Biol. 2015, 36, 9693–9701. [Google Scholar] [CrossRef]
- Zhang, X.J.; Ye, H.; Zeng, C.W.; He, B.; Zhang, H.; Chen, Y.Q. Dysregulation of miR-15a and miR-214 in human pancreatic cancer. J. Hematol. Oncol. 2010, 3, 46. [Google Scholar] [CrossRef]
- Zhang, W.-L.; Zhang, J.-H.; Wu, X.-Z.; Yan, T.; Lv, W. miR-15b promotes epithelial-mesenchymal transition by inhibiting SMURF2 in pancreatic cancer. Int. J. Oncol. 2015, 47, 1043–1053. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, J.; Zhang, L.; Zhu, Z.; Fan, J.; Chen, L.; Zhuang, L.; Luo, J.; Chen, H.; Liu, L.; et al. MicroRNA 23b regulates autophagy associated with radioresistance of pancreatic cancer cells. Gastroenterology 2013, 145, 1133–1143.e12. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, H.; Wang, X.; Zhou, L.; Li, H.; Deng, T.; Qu, Y.; Duan, J.; Bai, M.; Ge, S.; et al. The miR-24-Bim pathway promotes tumor growth and angiogenesis in pancreatic carcinoma. Oncotarget 2015, 6, 43831–43842. [Google Scholar] [CrossRef]
- He, G.; Zhang, L.; Li, Q.; Yang, L. miR-92a/DUSP10/JNK signalling axis promotes human pancreatic cancer cells proliferation. Biomed. Pharmacother. 2014, 68, 25–30. [Google Scholar] [CrossRef]
- Cai, B.; An, Y.; Lv, N.; Chen, J.; Tu, M.; Sun, J.; Wu, P.; Wei, J.; Jiang, K.; Miao, Y. miRNA-181b increases the sensitivity of pancreatic ductal adenocarcinoma cells to gemcitabine in vitro and in nude mice by targeting BCL-2. Oncol. Rep. 2013, 29, 1769–1776. [Google Scholar] [CrossRef]
- Takiuchi, D.; Eguchi, H.; Nagano, H.; Iwagami, Y.; Tomimaru, Y.; Wada, H.; Kawamoto, K.; Kobayashi, S.; Marubashi, S.; Tanemura, M.; et al. Involvement of microRNA-181b in the gemcitabine resistance of pancreatic cancer cells. Pancreatology 2013, 13, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Tang, J.; Zhuang, X.; Zhuang, Y.; Cheng, W.; Chen, W.; Yao, H.; Zhang, S. MiR-196a Promotes Pancreatic Cancer Progression by Targeting Nuclear Factor Kappa-B-Inhibitor Alpha. PLoS ONE 2014, 9, e87897. [Google Scholar] [CrossRef]
- Liu, M.B.; Du, Y.; Gao, J.; Liu, J.; Kong, X.; Gong, Y.B.; Li, Z.; Wu, H.B.; Chen, H.B. Aberrant expression miR-196a is associated with abnormal apoptosis, invasion, and proliferation of pancreatic cancer cells. Pancreas 2013, 42, 1169–1181. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, S.; Zhao, W.; Lu, Z.; Chen, J. miR-27a regulates the growth, colony formation and migration of pancreatic cancer cells by targeting Sprouty2. Cancer Lett. 2010, 298, 150–158. [Google Scholar] [CrossRef]
- Ma, J.; Fang, B.; Zeng, F.; Ma, C.; Pang, H.; Cheng, L.; Shi, Y.; Wang, H.; Yin, B.; Xia, J.; et al. Down-regulation of miR-223 reverses epithelial-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells. Oncotarget 2015, 6, 1740–1749. [Google Scholar] [CrossRef]
- Yong, S.; Yabin, Y.; Bing, Z.; Chuanrong, Z.; Dianhua, G.; Jianhuai, Z.; Weidong, Y.; Shuming, W.; Ling, L. Reciprocal regulation of DGCR5 and miR-320a affects the cellular malignant phenotype and 5-FU response in pancreatic ductal adenocarcinoma. Oncotarget 2017, 8, 90868–90878. [Google Scholar] [CrossRef]
- Laurila, E.M.; Sandström, S.; Rantanen, L.M.; Autio, R.; Kallioniemi, A. Both inhibition and enhanced expression of miR-31 lead to reduced migration and invasion of pancreatic cancer cells. Genes Chromosom. Cancer 2012, 51, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, B.; Shahbazi, R.; Khordadmehr, M. Dysregulation of key microRNAs in pancreatic cancer development. Biomed. Pharmacother. 2018, 109, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Nweke, E.E.; Brand, M. Downregulation of the let-7 family of microRNAs may promote insulin receptor/insulin-like growth factor signalling pathways in pancreatic ductal adenocarcinoma. Oncol. Lett. 2020, 20, 2613–2620. [Google Scholar] [CrossRef]
- Li, Z.; Li, X.; Yu, C.; Wang, M.; Peng, F.; Xiao, J.; Tian, R.; Jiang, J.; Sun, C. MicroRNA-100 regulates pancreatic cancer cells growth and sensitivity to chemotherapy through targeting FGFR3. Tumor Biol. 2014, 35, 11751–11759. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Zhong, X.; Jiang, X.; Leng, K.; Xu, Y.; Li, Z.; Huang, L.; Li, J.; Cui, Y. The role of long non-coding RNA AFAP1-AS1 in human malignant tumors. Pathol.-Res. Pract. 2018, 214, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, D.; Hua, R.; Zhang, J.; Liu, W.; Huo, Y.; Cheng, Y.; Hong, J.; Sun, Y. Long non-coding RNAs expressed in pancreatic ductal adenocarcinoma and lncRNA BC008363 an independent prognostic factor in PDAC. Pancreatology 2014, 14, 385–390. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, H.; Chen, C. Long non-coding RNA PCED1B-AS1 promotes pancreatic ductal adenocarcinoma progression by regulating the miR-411-3p/HIF-1α axis. Oncol. Rep. 2021, 46, 134. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.; Jiang, Y. Non-coding RNAs in Pancreatic Ductal Adenocarcinoma. Front. Oncol. 2020, 10, 309. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, L.; Shang, K.; Liu, F.; Che, J.; Li, H.; Cao, B. Long non-coding RNA H19, a novel therapeutic target for pancreatic cancer. Mol. Med. 2020, 26, 30. [Google Scholar] [CrossRef]
- Olivero, C.E.; Dimitrova, N. Identification and characterization of functional long noncoding RNAs in cancer. FASEB J. 2020, 34, 15630–15646. [Google Scholar] [CrossRef]
- Su, Y.; Gu, X.; Zheng, Q.; Zhu, L.; Lu, J.; Li, L. LncRNA PCGEM1 in Human Cancers: Functions, Mechanisms and Promising Clinical Utility. Front. Oncol. 2022, 12, 847745. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, L.; Mi, Y.; Tang, Y.; Reng, F.; Liu, B.; Zhang, Y.; Zheng, P. A ceRNA network and a potential regulatory axis in gastric cancer with different degrees of immune cell infiltration. Cancer Sci. 2020, 111, 4041–4050. [Google Scholar] [CrossRef]
- Bettin, N.; Oss Pegorar, C.; Cusanelli, E. The Emerging Roles of TERRA in Telomere Maintenance and Genome Stability. Cells 2019, 8, 246. [Google Scholar] [CrossRef]
- Feng, Q.; Chen, W.-D.; Wang, Y.-D. Gut Microbiota: An Integral Moderator in Health and Disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, X.; Cai, W.; Shi, R.; Yang, G.; Yuan, L. Host intestinal epithelium derived mirnas shape the microbiota and its implication in cardiovascular diseases. J. Am. Coll. Cardiol. 2017, 69, 1075. [Google Scholar] [CrossRef]
- Gesualdo, M.; Rizzi, F.; Bonetto, S.; Rizza, S.; Cravero, F.; Saracco, G.M.; De Angelis, C.G. Pancreatic Diseases and Microbiota: A Literature Review and Future Perspectives. J. Clin. Med. 2020, 9, 3535. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; Lucas, A.S.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef] [PubMed]
- Wong-Rolle, A.; Wei, H.K.; Zhao, C.; Jin, C. Unexpected guests in the tumor microenvironment: Microbiome in cancer. Protein Cell 2020, 12, 426–435. [Google Scholar] [CrossRef]
- Sammallahti, H.; Sarhadi, V.K.; Kokkola, A.; Ghanbari, R.; Rezasoltani, S.; Aghdaei, H.A.; Puolakkainen, P.; Knuutila, S. Oncogenomic Changes in Pancreatic Cancer and Their Detection in Stool. Biomolecules 2022, 12, 652. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Burns, M.B.; Subramanian, S.; Blekhman, R. Interaction between Host MicroRNAs and the Gut Microbiota in Colorectal Cancer. Msystems 2018, 3, e00205-17. [Google Scholar] [CrossRef]
- Sarhadi, V.; Lahti, L.; Saberi, F.; Youssef, O.; Kokkola, A.; Karla, T.; Tikkanen, M.; Rautelin, H.; Puolakkainen, P.; Salehi, R.; et al. Gut Microbiota and Host Gene Mutations in Colorectal Cancer Patients and Controls of Iranian and Finnish Origin. Anticancer Res. 2020, 40, 1325–1334. [Google Scholar] [CrossRef]
- Marin-Muller, C.; Li, D.; Bharadwaj, U.; Li, M.; Chen, C.; Hodges, S.E.; Fisher, W.E.; Mo, Q.; Hung, M.-C.; Yao, Q. A tumorigenic factor interactome connected through tumor suppressor MicroRNA-198 in human pancreatic cancer. Clin. Cancer Res. 2013, 19, 5901–5913. [Google Scholar] [CrossRef]
- Shirazi, M.S.R.; Al-Alo, K.Z.K.; Al-Yasiri, M.H.; Lateef, Z.M.; Ghasemian, A. Microbiome Dysbiosis and Predominant Bacterial Species as Human Cancer Biomarkers. J. Gastrointest. Cancer 2019, 51, 725–728. [Google Scholar] [CrossRef]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Gangemi, S. Interactions between the MicroRNAs and Microbiota in Cancer Development: Roles and Therapeutic Opportunities. Cancers 2020, 12, 805. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Noncoding RNAs: Regulatory Molecules of Host–Microbiome Crosstalk. Trends Microbiol. 2021, 29, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Tomkovich, S.; Gharaibeh, R.Z.; Dejea, C.M.; Pope, J.L.; Jiang, J.; Winglee, K.; Gauthier, J.; Newsome, R.C.; Yang, Y.; Fodor, A.A.; et al. Human Colon Mucosal Biofilms and Murine Host Communicate via Altered mRNA and microRNA Expression during Cancer. Msystems 2020, 5, e00451-19. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorak, K. Bile acids as endogenous etiologic agents in gastrointestinal cancer. World J. Gastroenterol. 2009, 15, 3329–3340. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).