
Asthma Phenotypes in the Era of Personalized Medicine



{kind=link}
{kind=link}
Abstract
:1. Introduction
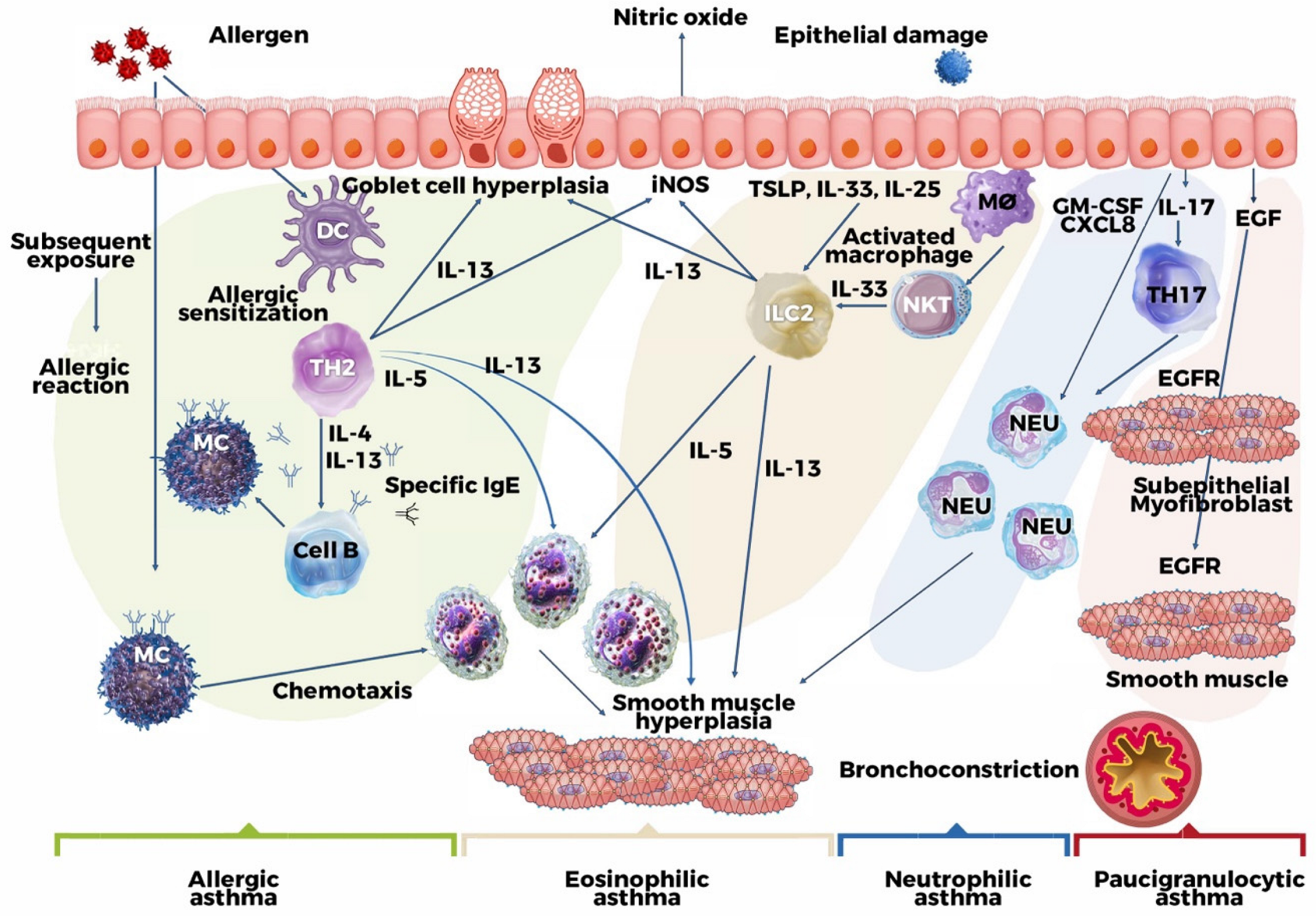
2. Pathogenetic Mechanism
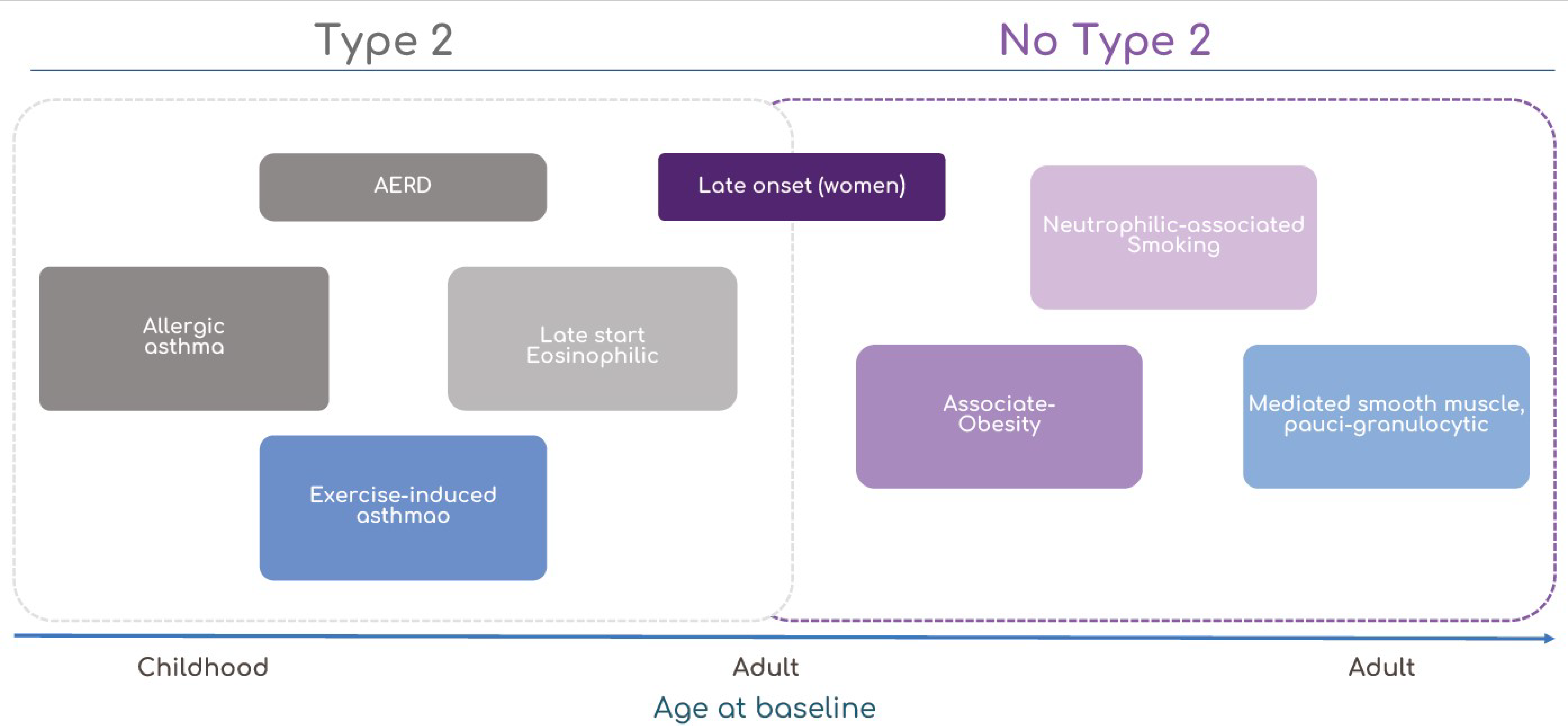
3. Clinical and Molecular Phenotypes of Asthma
3.1. Mild-Early-Onset Allergic Asthma
3.2. Late-Onset Eosinophilic Asthma
3.2.1. Non-Steroidal Anti-Inflammatory-Drug-Exacerbated Respiratory Disease
3.2.2. Exercise-Induced Bronchoconstriction
3.3. Non-Eosinophilic Neutrophilic Asthma Phenotypes
3.3.1. Obesity-Related Asthma
3.3.2. Perimenstrual Asthma
4. Therapeutic Targets in the Era of Personalized Medicine
4.1. Therapeutic Strategies for Anti-IgE
4.2. Therapeutic Strategies Involving Anti-IL5
4.3. Therapeutic Strategies for Anti-IL4-Receptor Alpha
4.4. Therapeutic Strategies against TSLP
5. Tools That Use Machine Learning to Improve Asthma Care in the Clinic
6. Gene and Cell Therapy
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hopkin, J.M. The diagnosis of asthma, a clinical syndrome. Thorax 2012, 67, 660–662. [Google Scholar] [CrossRef]
- Wenzel, S.E. Emergence of Biomolecular Pathways to Define Novel Asthma Phenotypes. Type-2 Immunity and Beyond. Am. J. Respir. Cell Mol. Biol. 2016, 55, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. 2017, 4, 158. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Agache, I.; Akdis, C.A. Precision medicine and phenotypes, endotypes, genotypes, regiotypes, and theratypes of allergic diseases. J. Clin. Investig. 2019, 129, 1493–1503. [Google Scholar] [CrossRef]
- Chang, H.S.; Lee, T.-H.; Jun, J.A.; Baek, A.R.; Park, J.-S.; Koo, S.-M.; Kim, Y.K.; Lee, H.S.; Park, C.-S. Neutrophilic inflammation in asthma: Mechanisms and therapeutic considerations. Expert Rev. Respir. Med. 2017, 11, 29–40. [Google Scholar] [CrossRef]
- Salter, B.M.; Aw, M.; Sehmi, R. The role of type 2 innate lymphoid cells in eosinophilic asthma. J. Leukoc. Biol. 2019, 106, 889–901. [Google Scholar] [CrossRef]
- Pavord, I.D.; Korn, S.; Howarth, P.; Bleecker, E.R.; Buhl, R.; Keene, O.N.; Ortega, H.; Chanez, P. Mepolizumab for severe eosinophilic asthma (DREAM): A multicentre, double-blind, placebo-controlled trial. Lancet 2012, 380, 651–659. [Google Scholar] [CrossRef]
- Ortega, H.G.; Yancey, S.W.; Mayer, B.; Gunsoy, N.B.; Keene, O.N.; Bleecker, E.R.; E Brightling, C.; Pavord, I.D. Severe eosinophilic asthma treated with mepolizumab stratified by baseline eosinophil thresholds: A secondary analysis of the DREAM and MENSA studies. Lancet Respir. Med. 2016, 4, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Bleecker, E.R.; FitzGerald, J.M.; Chanez, P.; Papi, A.; Weinstein, S.F.; Barker, P.; Sproule, S.; Gilmartin, G.; Aurivillius, M.; Werkström, V.; et al. Efficacy and safety of benralizumab for patients with severe asthma uncontrolled with high-dosage inhaled corticosteroids and long-acting beta(2)-agonists (SIROCCO): A randomised, multicentre, placebo-controlled phase 3 trial. Lancet 2016, 388, 2115–2127. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Corren, J.; Pavord, I.D.; Maspero, J.; Wenzel, S.; Rabe, K.F.; Busse, W.W.; Ford, L.; Sher, L.; Fitzgerald, J.M.; et al. Dupilumab Efficacy and Safety in Moderate-to-Severe Uncontrolled Asthma. N. Engl. J. Med. 2018, 378, 2486–2496. [Google Scholar] [CrossRef]
- Maspero, J.F.; Katelaris, C.H.; Busse, W.W.; Castro, M.; Corren, J.; Chipps, B.E.; Peters, A.T.; Pavord, I.D.; Ford, L.B.; Sher, L.; et al. Dupilumab Efficacy in Uncontrolled, Moderate-to-Severe Asthma with Self-Reported Chronic Rhinosinusitis. J. Allergy Clin. Immunol Pract. 2020, 8, 527–539.e9. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, M.E.; Ford, L.B.; Maspero, J.F.; Pavord, I.D.; Papi, A.; Bourdin, A.; Watz, H.; Castro, M.; Nenasheva, N.M.; Tohda, Y.; et al. Long-term safety and efficacy of dupilumab in patients with moderate-to-severe asthma (TRAVERSE): An open-label extension study. Lancet Respir. Med. 2022, 10, 11–25. [Google Scholar] [CrossRef]
- Jonckheere, A.C.; Bullens, D.M.A.; Seys, S.F. Innate lymphoid cells in asthma: Pathophysiological insights from murine models to human asthma phenotypes. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 53–60. [Google Scholar] [CrossRef]
- Godar, M.; Blanchetot, C.; de Haard, H.; Lambrecht, B.N.; Brusselle, G. Personalized medicine with biologics for severe type 2 asthma: Current status and future prospects. MAbs 2018, 10, 34–45. [Google Scholar] [CrossRef]
- Eberl, G.; Colonna, M.; Di Santo, J.P.; McKenzie, A.N. Innate lymphoid cells. Innate lymphoid cells: A new paradigm in immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef]
- Cortez, V.S.; Robinette, M.L.; Colonna, M. Innate lymphoid cells: New insights into function and development. Curr. Opin. Immunol. 2015, 32, 71–77. [Google Scholar] [CrossRef]
- Roan, F.; Obata-Ninomiya, K.; Ziegler, S.F. Epithelial cell-derived cytokines: More than just signaling the alarm. J. Clin. Investig. 2019, 129, 1441–1451. [Google Scholar] [CrossRef]
- Varricchi, G.; Pecoraro, A.; Marone, G.; Criscuolo, G.; Spadaro, G.; Genovese, A.; Marone, G. Thymic Stromal Lymphopoietin Isoforms, Inflammatory Disorders, and Cancer. Front. Immunol. 2018, 9, 1595. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Verma, M.; Michalec, L.; Liu, W.; Sripada, A.; Rollins, D.; Good, J.; Ito, Y.; Chu, H.; Gorska, M.M.; et al. Steroid resistance of airway type 2 innate lymphoid cells from patients with severe asthma: The role of thymic stromal lymphopoietin. J. Allergy Clin. Immunol. 2018, 141, 257–268.e6. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiri, K.; Fornasa, G.; Rescigno, M. Thymic Stromal Lymphopoietin: To Cut a Long Story Short. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Boita, M.; Garzaro, M.; Raimondo, L.; Riva, G.; Mazibrada, J.; Vizio, B.; Bellone, G.; Pecorari, G.; Bucca, C.; Rolla, G.; et al. The expression of TSLP receptor in chronic rhinosinusitis with and without nasal polyps. Int. J. Immunopathol. Pharmacol. 2011, 24, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Boita, M.; Heffler, E.; Omedè, P.; Bellocchia, M.; Bussolino, C.; Solidoro, P.; Giorgis, V.; Guerrera, F.; Riva, G.; Brussino, L.; et al. Basophil Membrane Expression of Epithelial Cytokine Receptors in Patients with Severe Asthma. Int. Arch. Allergy Immunol. 2018, 175, 171–176. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef]
- Peebles, R.S., Jr.; Aronica, M.A. Proinflammatory Pathways in the Pathogenesis of Asthma. Clin. Chest Med. 2019, 40, 29–50. [Google Scholar] [CrossRef]
- Yao, X.; Sun, Y.; Wang, W.; Sun, Y. Interleukin (IL)-25: Pleiotropic roles in asthma. Respirology 2016, 21, 638–647. [Google Scholar] [CrossRef]
- Kouzaki, H.; Tojima, I.; Kita, H.; Shimizu, T. Transcription of interleukin-25 and extracellular release of the protein is regulated by allergen proteases in airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2013, 49, 741–750. [Google Scholar] [CrossRef]
- Ray, A.; Das, J.; Wenzel, S.E. Determining asthma endotypes and outcomes: Complementing existing clinical practice with modern machine learning. Cell Rep. Med. 2022, 3, 100857. [Google Scholar] [CrossRef]
- Zhu, J. T helper 2 (Th2) cell differentiation, type 2 innate lymphoid cell (ILC2) development and regulation of interleukin-4 (IL-4) and IL-13 production. Cytokine 2015, 75, 14–24. [Google Scholar] [CrossRef]
- Tindemans, I.; Serafini, N.; Di Santo, J.P.; Hendriks, R.W. GATA-3 function in innate and adaptive immunity. Immunity 2014, 41, 191–206. [Google Scholar] [CrossRef]
- Varricchi, G.; Bagnasco, D.; Borriello, F.; Heffler, E.; Canonica, G.W. Interleukin-5 pathway inhibition in the treatment of eosinophilic respiratory disorders: Evidence and unmet needs. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 186–200. [Google Scholar] [CrossRef]
- Doran, E.; Cai, F.; Holweg, C.T.J.; Wong, K.; Brumm, J.; Arron, J.R. Interleukin-13 in Asthma and Other Eosinophilic Disorders. Front. Med. 2017, 4, 139. [Google Scholar] [CrossRef]
- Gandhi, N.A.; Pirozzi, G.; Graham, N.M.H. Commonality of the IL-4/IL-13 pathway in atopic diseases. Expert Rev. Clin. Immunol. 2017, 13, 425–437. [Google Scholar] [CrossRef]
- Maspero, J.; Adir, Y.; Al-Ahmad, M.; Celis-Preciado, C.A.; Colodenco, F.D.; Giavina-Bianchi, P.; Lababidi, H.; Ledanois, O.; Mahoub, B.; Perng, D.-W.; et al. Type 2 inflammation in asthma and other airway diseases. ERJ Open Res. 2022, 8. [Google Scholar] [CrossRef]
- Pelaia, C.; Pelaia, G.; Maglio, A.; Tinello, C.; Gallelli, L.; Lombardo, N.; Terracciano, R.; Vatrella, A. Pathobiology of Type 2 Inflammation in Asthma and Nasal Polyposis. J. Clin. Med. 2023, 12, 3371. [Google Scholar] [CrossRef]
- Walker, C.; Bode, E.; Boer, L.; Hansel, T.T.; Blaser, K.; Virchow, J.C., Jr. Allergic and nonallergic asthmatics have distinct patterns of T-cell activation and cytokine production in peripheral blood and bronchoalveolar lavage. Am. Rev. Respir. Dis. 1992, 146, 109–115. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, Y.; Jabeen, R.; Nguyen, E.T.; Wilkes, D.S.; Tepper, R.S.; Kaplan, M.H.; Zhou, B. Interleukin-9 is required for allergic airway inflammation mediated by the cytokine TSLP. Immunity 2013, 38, 360–372. [Google Scholar] [CrossRef]
- Ricciardolo, F.L.; Sorbello, V.; Folino, A.; Gallo, F.; Massaglia, G.M.; Favatà, G.; Conticello, S.; Vallese, D.; Gani, F.; Malerba, M.; et al. Identification of IL-17F/frequent exacerbator endotype in asthma. J. Allergy Clin. Immunol. 2017, 140, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.R.; Siegel, L.; Leith, A.; Mohn, D.; Escobar, S.; Wannberg, S.; Misura, K.; Rickel, E.; Rottman, J.B.; Comeau, M.R.; et al. IL-17RA Signaling in Airway Inflammation and Bronchial Hyperreactivity in Allergic Asthma. Am. J. Respir. Cell Mol. Biol. 2015, 53, 810–821. [Google Scholar] [CrossRef]
- Ota, K.; Kawaguchi, M.; Matsukura, S.; Kurokawa, M.; Kokubu, F.; Fujita, J.; Morishima, Y.; Huang, S.-K.; Ishii, Y.; Satoh, H.; et al. Potential involvement of IL-17F in asthma. J. Immunol. Res. 2014, 2014, 602846. [Google Scholar] [CrossRef]
- Nalbant, A.; Eskier, D. Genes associated with T helper 17 cell differentiation and function. Front. Biosci. 2016, 8, 427–435. [Google Scholar] [CrossRef]
- Wan, Y.I.; Shrine, N.R.G.; Artigas, M.S.; Wain, L.V.; Blakey, J.D.; Moffatt, M.F.; Bush, A.; Chung, K.F.; Cookson, W.O.C.M.; Strachan, D.P.; et al. Genome-wide association study to identify genetic determinants of severe asthma. Thorax 2012, 67, 762–768. [Google Scholar] [CrossRef]
- Shrine, N.; A Portelli, M.; John, C.; Artigas, M.S.; Bennett, N.; Hall, R.; Lewis, J.; Henry, A.P.; Billington, C.K.; Ahmad, A.; et al. Moderate-to-severe asthma in individuals of European ancestry: A genome-wide association study. Lancet Respir. Med. 2019, 7, 20–34. [Google Scholar] [CrossRef]
- Hekking, P.-P.; Loza, M.J.; Pavlidis, S.; de Meulder, B.; Lefaudeux, D.; Baribaud, F.; Auffray, C.; Wagener, A.H.; Brinkman, P.; Lutter, R.; et al. Pathway discovery using transcriptomic profiles in adult-onset severe asthma. J. Allergy Clin. Immunol. 2018, 141, 1280–1290. [Google Scholar] [CrossRef]
- Bigler, J.; Boedigheimer, M.; Schofield, J.P.R.; Skipp, P.J.; Corfield, J.; Rowe, A.; Sousa, A.R.; Timour, M.; Twehues, L.; Hu, X.; et al. A Severe Asthma Disease Signature from Gene Expression Profiling of Peripheral Blood from U-BIOPRED Cohorts. Am. J. Respir. Crit. Care Med. 2017, 195, 1311–1320. [Google Scholar] [CrossRef]
- Modena, B.D.; Bleecker, E.R.; Busse, W.W.; Erzurum, S.C.; Gaston, B.M.; Jarjour, N.N.; Meyers, D.A.; Milosevic, J.; Tedrow, J.R.; Wu, W.; et al. Gene Expression Correlated with Severe Asthma Characteristics Reveals Heterogeneous Mechanisms of Severe Disease. Am. J. Respir. Crit. Care Med. 2017, 195, 1449–1463. [Google Scholar] [CrossRef]
- Singhania, A.; Rupani, H.; Jayasekera, N.; Lumb, S.; Hales, P.; Gozzard, N.; Davies, D.E.; Woelk, C.H.; Howarth, P.H. Altered Epithelial Gene Expression in Peripheral Airways of Severe Asthma. PLoS ONE 2017, 12, e0168680. [Google Scholar] [CrossRef]
- Gautam, Y.; Johansson, E.; Mersha, T.B. Multi-Omics Profiling Approach to Asthma: An Evolving Paradigm. J. Pers. Med. 2022, 12, 66. [Google Scholar] [CrossRef]
- Tyler, S.R.; Bunyavanich, S. Leveraging-omics for asthma endotyping. J. Allergy Clin. Immunol. 2019, 144, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, L.; Chen, D.; Feng, M.; Lu, Y.; Chen, R.; Qiu, C.; Li, J. The application of proteomics in the diagnosis and treatment of bronchial asthma. Ann. Transl. Med. 2020, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Rhim, T. Application of proteomics in asthma research. Expert Rev. Proteom. 2011, 8, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Zhang, X.; Chen, X.; Brown, A.P.; Weirauch, M.T.; Guilbert, T.W.; Hershey, G.K.K.; Biagini, J.M.; Ji, H. Nasal DNA methylation differentiates severe from non-severe asthma in African-American children. Allergy 2021, 76, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mao, Z.D.; Shi, Y.J.; Qian, Y.; Liu, Z.G.; Yin, X.W.; Zhang, Q. Comprehensive analysis of miRNA-mRNA-lncRNA networks in severe asthma. Epigenomics 2019, 11, 115–131. [Google Scholar] [CrossRef]
- Moore, W.C.; Meyers, D.A.; Wenzel, S.E.; Teague, W.G.; Li, H.; Li, X.; D’Agostino, R., Jr.; Castro, M.; Curran-Everett, D.; Fitzpatrick, A.M.; et al. Identification of asthma phenotypes using cluster analysis in the Severe Asthma Research Program. Am. J. Respir. Crit. Care Med. 2010, 181, 315–323. [Google Scholar] [CrossRef]
- Haldar, P.; Pavord, I.D.; Shaw, D.E.; Berry, M.A.; Thomas, M.; Brightling, C.E.; Wardlaw, A.J.; Green, R.H. Cluster analysis and clinical asthma phenotypes. Am. J. Respir. Crit. Care Med. 2008, 178, 218–224. [Google Scholar] [CrossRef]
- Lefaudeux, D.; De Meulder, B.; Loza, M.J.; Peffer, N.; Rowe, A.; Baribaud, F.; Bansal, A.T.; Lutter, R.; Sousa, A.R.; Corfield, J.; et al. U-BIOPRED clinical adult asthma clusters linked to a subset of sputum omics. J. Allergy Clin. Immunol. 2017, 139, 1797–1807. [Google Scholar] [CrossRef]
- Yan, X.; Chu, J.-H.; Gomez, J.; Koenigs, M.; Holm, C.; He, X.; Perez, M.F.; Zhao, H.; Mane, S.; Martinez, F.D.; et al. Noninvasive analysis of the sputum transcriptome discriminates clinical phenotypes of asthma. Am. J. Respir. Crit. Care Med. 2015, 191, 1116–1125. [Google Scholar] [CrossRef]
- Kaur, R.; Chupp, G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J. Allergy Clin. Immunol. 2019, 144, 1–12. [Google Scholar] [CrossRef]
- Khalaf, K.; Paoletti, G.; Puggioni, F.; Racca, F.; De Luca, F.; Giorgis, V.; Canonica, G.W.; Heffler, E. Asthma from immune pathogenesis to precision medicine. Semin. Immunol. 2019, 46, 101294. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Taunk, S.T.; Cardet, J.C.; Ledford, D.K. Clinical implications of asthma endotypes and phenotypes. Allergy Asthma Proc. 2022, 43, 375–382. [Google Scholar] [CrossRef]
- Makrinioti, H.; Tiotiu, A.; Gonzalez-Barcala, F.J. Severe asthma patients’ and physicians’ perspectives of disease burden: Do they match? ERJ Open Res. 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, B.; Chatburn, E.; Bansal, A.T.; Fulton, O.; Hamerlijnck, D.; Coleman, C.; Eger, K.; Hyland, M.; Holmes, J.; Heaney, L.; et al. What bothers severe asthma patients most? A paired patient-clinician study across seven European countries. ERJ Open Res. 2023, 9. [Google Scholar] [CrossRef]
- Porsbjerg, C.; Menzies-Gow, A. Co-morbidities in severe asthma: Clinical impact and management. Respirology 2017, 22, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Bucca, C.; Culla, B.; Brussino, L.; Ricciardolo, F.L.; Cicolin, A.; Heffler, E.; Bugiani, M.; Rolla, G. Effect of iron supplementation in women with chronic cough and iron deficiency. Int. J. Clin. Pract. 2012, 66, 1095–1100. [Google Scholar] [CrossRef]
- Robinson, D.; Humbert, M.; Buhl, R.; Cruz, A.A.; Inoue, H.; Korom, S.; Hanania, N.A.; Nair, P. Revisiting Type 2-high and Type 2-low airway inflammation in asthma: Current knowledge and therapeutic implications. Clin. Exp. Allergy 2017, 47, 161–175. [Google Scholar] [CrossRef]
- Gao, J.; Wu, F.; Wu, S.; Yang, X. Inflammatory Subtypes in Classic Asthma and Cough Variant Asthma. J. Inflamm. Res. 2020, 13, 1167–1173. [Google Scholar] [CrossRef]
- Matsuoka, H.; Niimi, A.; Matsumoto, H.; Takemura, M.; Ueda, T.; Yamaguchi, M.; Jinnai, M.; Inoue, H.; Ito, I.; Chin, K.; et al. Inflammatory subtypes in cough-variant asthma: Association with maintenance doses of inhaled corticosteroids. Chest 2010, 138, 1418–1425. [Google Scholar] [CrossRef]
- Chen, M.; Shepard, K., 2nd; Yang, M.; Raut, P.; Pazwash, H.; Holweg, C.T.J.; Choo, E. Overlap of allergic, eosinophilic and type 2 inflammatory subtypes in moderate-to-severe asthma. Clin. Exp. Allergy 2021, 51, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Oppenheimer, J. Elucidating asthma phenotypes and endotypes: Progress towards personalized medicine. Ann. Allergy Asthma Immunol. 2016, 116, 394–401. [Google Scholar] [CrossRef] [PubMed]
- de Kleer, I.M.; Kool, M.; de Bruijn, M.J.; Willart, M.; van Moorleghem, J.; Schuijs, M.J.; Plantinga, M.; Beyaert, R.; Hams, E.; Fallon, P.G.; et al. Perinatal Activation of the Interleukin-33 Pathway Promotes Type 2 Immunity in the Developing Lung. Immunity 2016, 45, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Castro-Rodriguez, J.A.; Saglani, S.; Rodriguez-Martinez, C.E.; Oyarzun, M.A.; Fleming, L.; Bush, A. The relationship between inflammation and remodeling in childhood asthma: A systematic review. Pediatr. Pulmonol. 2018, 53, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.M. Severe asthma: Epidemiology, burden of illness, and heterogeneity. Allergy Asthma Proc. 2015, 36, 418–424. [Google Scholar] [CrossRef]
- Larenas-Linnemann, D.; Salas-Hernández, J.; Del Río-Navarro, B.E.; Luna-Pech, J.A.; Navarrete-Rodríguez, E.M.; Gochicoa, L.; Cano-Salas, M.d.C.; García-Ramírez, U.N.; López-Estrada, E.d.C.; Ortega-Martell, J.A.; et al. [MIA 2021, Comprehensive Asthma Management. Guidelines for Mexico]. Rev. Alerg. Mex. 2021, 68 (Suppl. S1), s1–s122. [Google Scholar] [CrossRef]
- Pakkasela, J.; Ilmarinen, P.; Honkamäki, J.; Tuomisto, L.E.; Andersén, H.; Piirilä, P.; Hisinger-Mölkänen, H.; Sovijärvi, A.; Backman, H.; Lundbäck, B.; et al. Age-specific incidence of allergic and non-allergic asthma. BMC Pulm. Med. 2020, 20, 9. [Google Scholar] [CrossRef]
- Bachert, C.; Marple, B.; Schlosser, R.J.; Hopkins, C.; Schleimer, R.P.; Lambrecht, B.N.; Bröker, B.M.; Laidlaw, T.; Song, W.-J. Adult chronic rhinosinusitis. Nat. Rev. Dis. Primers 2020, 6, 86. [Google Scholar] [CrossRef]
- Peters, M.C.; Ringel, L.; Dyjack, N.; Herrin, R.; Woodruff, P.G.; Rios, C.; O’connor, B.; Fahy, J.V.; Seibold, M.A. A Transcriptomic Method to Determine Airway Immune Dysfunction in T2-High and T2-Low Asthma. Am. J. Respir. Crit. Care Med. 2019, 199, 465–477. [Google Scholar] [CrossRef]
- Hirano, T.; Matsunaga, K. Late-onset asthma: Current perspectives. J. Asthma Allergy 2018, 11, 19–27. [Google Scholar] [CrossRef]
- Ozyigit, L.P.; Morita, H.; Akdis, M. Innate lymphocyte cells in asthma phenotypes. Clin. Transl. Allergy 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, M.L.; Agache, I.; Bavbek, S.; Bakirtas, A.; Blanca, M.; Bochenek, G.; Bonini, M.; Heffler, E.; Klimek, L.; Laidlaw, T.M.; et al. Diagnosis and management of NSAID-Exacerbated Respiratory Disease (N-ERD)-a EAACI position paper. Allergy 2019, 74, 28–39. [Google Scholar] [CrossRef]
- Woo, S.D.; Luu, Q.Q.; Park, H.S. NSAID-Exacerbated Respiratory Disease (NERD): From Pathogenesis to Improved Care. Front. Pharmacol. 2020, 11, 1147. [Google Scholar] [CrossRef] [PubMed]
- Bonini, M.; Silvers, W. Exercise-Induced Bronchoconstriction: Background, Prevalence, and Sport Considerations. Immunol. Allergy Clin. N. Am. 2018, 38, 205–214. [Google Scholar] [CrossRef]
- Malewska-Kaczmarek, K.; Podlecka, D.; Mankowski, T.; Jerzynska, J.; Stelmach, I. Exercise-Induced Bronchoconstriction in Children: A Comparison between Athletes and Non-Athletes. Healthcare 2023, 11, 1349. [Google Scholar] [CrossRef]
- Vollsaeter, M.; Stensrud, T.; Maat, R.; Halvorsen, T.; Roksund, O.D.; Sandnes, A.; Clemm, H. Exercise Related Respiratory Problems in the Young-Is It Exercise-Induced Bronchoconstriction or Laryngeal Obstruction? Front. Pediatr. 2021, 9, 800073. [Google Scholar] [CrossRef] [PubMed]
- Tikkakoski, A.P.; Tikkakoski, A.; Sipilä, K.; Kivistö, J.E.; Huhtala, H.; Kähönen, M.; Karjalainen, J.; Lehtimäki, L. Exercise-induced bronchoconstriction is associated with air humidity and particulate matter concentration in preschool children. Pediatr. Pulmonol. 2023, 58, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Klain, A.; Indolfi, C.; Dinardo, G.; Contieri, M.; Decimo, F.; Miraglia Del Giudice, M. Exercise-Induced Bronchoconstriction in Children. Front. Med. 2021, 8, 814976. [Google Scholar] [CrossRef]
- Tliba, O.; Panettieri, R.A., Jr. Paucigranulocytic asthma: Uncoupling of airway obstruction from inflammation. J. Allergy Clin. Immunol. 2019, 143, 1287–1294. [Google Scholar] [CrossRef]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef]
- Seys, S.F.; Lokwani, R.; Simpson, J.L.; Bullens, D.M.A. New insights in neutrophilic asthma. Curr. Opin. Pulm. Med. 2019, 25, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Strzelak, A.; Ratajczak, A.; Adamiec, A.; Feleszko, W. Tobacco Smoke Induces and Alters Immune Responses in the Lung Triggering Inflammation, Allergy, Asthma and Other Lung Diseases: A Mechanistic Review. Int. J. Environ. Res. Public Health 2018, 15, 1033. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Uribe, V.; Martinez-Tenopala, R.; Baro-Alvarez, P.d.; Mojica-Gonzalez, Z. Frequency of ADIPOQ 276 and ADIPOQ 45 Polymorphisms in Obese and Eutrophic Adolescents with and without Asthma and their Relationship with Serum Adiponectin Levels. Med. Res. Arch. 2022, 10. [Google Scholar] [CrossRef]
- Hanania, N.A.; King, M.J.; Braman, S.S.; Saltoun, C.; Wise, R.A.; Enright, P.; Falsey, A.R.; Mathur, S.K.; Ramsdell, J.W.; Rogers, L.; et al. Asthma in the elderly: Current understanding and future research needs—A report of a National Institute on Aging (NIA) workshop. J. Allergy Clin. Immunol. 2011, 128 (Suppl. S3), S4–S24. [Google Scholar] [CrossRef]
- Ford, M.L.; Ruwanpathirana, A.; Lewis, B.W.; Britt, R.D., Jr. Aging-Related Mechanisms Contribute to Corticosteroid Insensitivity in Elderly Asthma. Int. J. Mol. Sci. 2023, 24, 6347. [Google Scholar] [CrossRef] [PubMed]
- Khosa, J.K.; Louie, S.; Moreno, P.L.; Abramov, D.; Rogstad, D.K.; Alismail, A.; Matus, M.J.; Tan, L.D. Asthma Care in the Elderly: Practical Guidance and Challenges for Clinical Management—A Framework of 5 “Ps”. J. Asthma Allergy 2023, 16, 33–43. [Google Scholar] [CrossRef]
- Jartti, T.; Saarikoski, L.; Jartti, L.; Lisinen, I.; Jula, A.; Huupponen, R.; Viikari, J.; Raitakari, O.T. Obesity, adipokines and asthma. Allergy 2009, 64, 770–777. [Google Scholar] [CrossRef]
- Yuksel, H.; Sogut, A.; Yilmaz, O.; Onur, E.; Dinc, G. Role of adipokines and hormones of obesity in childhood asthma. Allergy Asthma Immunol. Res. 2012, 4, 98–103. [Google Scholar] [CrossRef]
- Leija-Martínez, J.J.; Giacoman-Martínez, A.; Del-Río-Navarro, B.E.; Sanchéz-Muñoz, F.; Hernández-Diazcouder, A.; Muñoz-Hernández, O.; Romero-Nava, R.; Villafaña, S.; Marchat, L.A.; Hong, E.; et al. Promoter methylation status of RORC, IL17A, and TNFA in peripheral blood leukocytes in adolescents with obesity-related asthma. Heliyon 2022, 8, e12316. [Google Scholar] [CrossRef]
- Vieira, C.P.; de Oliveira, L.P.; Da Silva, M.B.; Majolli Andre, D.; Tavares, E.B.G.; Pimentel, E.R.; Antunes, E. Role of metalloproteinases and TNF-alpha in obesity-associated asthma in mice. Life Sci. 2020, 259, 118191. [Google Scholar] [CrossRef]
- Leija-Martínez, J.J.; Del-Río-Navarro, B.E.; Sanchéz-Muñoz, F.; Muñoz-Hernández, O.; Hong, E.; Giacoman-Martínez, A.; Romero-Nava, R.; Patricio-Román, K.L.; Hall-Mondragon, M.S.; Espinosa-Velazquez, D.; et al. Associations of TNFA, IL17A, and RORC mRNA expression levels in peripheral blood leukocytes with obesity-related asthma in adolescents. Clin. Immunol. 2021, 229, 108715. [Google Scholar] [CrossRef] [PubMed]
- Yon, C.; Thompson, D.A.; Jude, J.A.; Panettieri, R.A.; Jr Rastogi, D. Crosstalk between CD4(+) T Cells and Airway Smooth Muscle in Pediatric Obesity-related Asthma. Am. J. Respir Crit. Care Med. 2023, 207, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Aguilar, N.E.; Del Rio-Navarro, B.E.; Navarro-Olivos, E.; Garcia-Ortiz, H.; Orozco, L.; Jimenez-Morales, S. SPINK5 and ADRB2 haplotypes are risk factors for asthma in Mexican pediatric patients. J. Asthma 2015, 52, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhu, Z.; Xiao, Q.; Li, J.; Hong, X.; Wang, X.; Hasegawa, K.; Camargo, C.A.; Liang, L. Obesity-related biomarkers underlie a shared genetic architecture between childhood body mass index and childhood asthma. Commun. Biol. 2022, 5, 1098. [Google Scholar] [CrossRef] [PubMed]
- Everaere, L.; Ait Yahia, S.; Bouté, M.; Audousset, C.; Chenivesse, C.; Tsicopoulos, A. Innate lymphoid cells at the interface between obesity and asthma. Immunology 2018, 153, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.E.; Peters, U. The effect of obesity on lung function. Expert Rev. Respir. Med. 2018, 12, 755–767. [Google Scholar] [CrossRef]
- Spathopoulos, D.; Paraskakis, E.; Trypsianis, G.; Tsalkidis, A.; Arvanitidou, V.; Emporiadou, M.; Bouros, D.; Chatzimichael, A. The effect of obesity on pulmonary lung function of school aged children in Greece. Pediatr. Pulmonol. 2009, 44, 273–280. [Google Scholar] [CrossRef]
- Mahadev, S.; Salome, C.M.; Berend, N.; King, G.G. The effect of low lung volume on airway function in obesity. Respir. Physiol. Neurobiol. 2013, 188, 192–199. [Google Scholar] [CrossRef]
- Castro-Rodríguez, J.A.; Holberg, C.J.; Morgan, W.J.; Wright, A.L.; Martinez, F.D. Increased incidence of asthmalike symptoms in girls who become overweight or obese during the school years. Am. J. Respir. Crit. Care Med. 2001, 163, 1344–1349. [Google Scholar] [CrossRef]
- Guerra, S.; Wright, A.L.; Morgan, W.J.; Sherrill, D.L.; Holberg, C.J.; Martinez, F.D. Persistence of asthma symptoms during adolescence: Role of obesity and age at the onset of puberty. Am. J. Respir. Crit. Care Med. 2004, 170, 78–85. [Google Scholar] [CrossRef]
- Castro-Rodriguez, J.A. A new childhood asthma phenotype: Obese with early menarche. Paediatr. Respir. Rev. 2016, 18, 85–89. [Google Scholar] [CrossRef]
- Starr, S.; Wysocki, M.; DeLeon, J.D.; Silverstein, G.; Arcoleo, K.; Rastogi, D.; Feldman, J.M. Obesity-related pediatric asthma: Relationships between pulmonary function and clinical outcomes. J. Asthma 2023, 60, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Bhatawadekar, S.A.; Peters, U.; Walsh, R.R.; Daphtary, N.; MacLean, E.S.; Mori, V.; Hodgdon, K.; Kinsey, C.M.; Kaminsky, D.A.; Bates, J.H.; et al. Central airway collapse is related to obesity independent of asthma phenotype. Respirology 2021, 26, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Angel, J.; Kaviany, P.; Rastogi, D.; Forno, E. Obesity-related asthma in children and adolescents. Lancet Child Adolesc. Health 2022, 6, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Hudler, A.C.; Diaz, I.R.R.; Sharma, S.; Holguin, F. Gaps and Future Directions in Clinical Research on Obesity-Related Asthma. Pulm. Ther. 2023, 9, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Agache, I.; Eguiluz-Gracia, I.; Cojanu, C.; Laculiceanu, A.; del Giacco, S.; Zemelka-Wiacek, M.; Kosowska, A.; Akdis, C.A.; Jutel, M. Advances and highlights in asthma in 2021. Allergy 2021, 76, 3390–3407. [Google Scholar] [CrossRef]
- Becker, A.B.; Abrams, E.M. Asthma guidelines: The Global Initiative for Asthma in relation to national guidelines. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 99–103. [Google Scholar] [CrossRef]
- Murphy, V.E.; Gibson, P.G. Premenstrual asthma: Prevalence, cycle-to-cycle variability and relationship to oral contraceptive use and menstrual symptoms. J. Asthma 2008, 45, 696–704. [Google Scholar] [CrossRef]
- Brenner, B.E.; Holmes, T.M.; Mazal, B.; Camargo, C.A., Jr. Relation between phase of the menstrual cycle and asthma presentations in the emergency department. Thorax 2005, 60, 806–809. [Google Scholar] [CrossRef]
- Graziottin, A.; Serafini, A. Perimenstrual asthma: From pathophysiology to treatment strategies. Multidiscip. Respir. Med. 2016, 11, 30. [Google Scholar] [CrossRef]
- Douin-Echinard, V.; Calippe, B.; Billon-Galès, A.; Fontaine, C.; Lenfant, F.; Trémollières, F.; Bayard, F.; Guéry, J.C.; Arnal, J.F.; Gourdy, P. Estradiol administration controls eosinophilia through estrogen receptor-alpha activation during acute peritoneal inflammation. J. Leukoc. Biol. 2011, 90, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Michael, E.; Bundy, D.A.; Grenfell, B.T. Re-assessing the global prevalence and distribution of lymphatic filariasis. Parasitology. 1996, 112 Pt 4, 409–428. [Google Scholar] [CrossRef] [PubMed]
- Whitacre, C.C. Sex differences in autoimmune disease. Nat. Immunol. 2001, 2, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Huygen, K.; Palfliet, K. Strain variation in interferon gamma production of BCG-sensitized mice challenged with PPD II. Importance of one major autosomal locus and additional sexual influences. Cell. Immunol. 1984, 85, 75–81. [Google Scholar] [CrossRef]
- Beato, M.; Herrlich, P.; Schütz, G. Steroid hormone receptors: Many actors in search of a plot. Cell 1995, 83, 851–857. [Google Scholar] [CrossRef]
- Phiel, K.L.; Henderson, R.A.; Adelman, S.J.; Elloso, M.M. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol. Lett. 2005, 97, 107–113. [Google Scholar] [CrossRef]
- Graziottin, A. The shorter, the better: A review of the evidence for a shorter contraceptive hormone-free interval. Eur. J. Contracept. Reprod. Health Care 2016, 21, 93–105. [Google Scholar] [CrossRef]
- Berbic, M.; Ng, C.H.; Fraser, I.S. Inflammation and endometrial bleeding. Climacteric 2014, 17 (Suppl. S2), 47–53. [Google Scholar] [CrossRef]
- Menzies, F.M.; Shepherd, M.C.; Nibbs, R.J.; Nelson, S.M. The role of mast cells and their mediators in reproduction, pregnancy and labour. Hum. Reprod. Update 2011, 17, 383–396. [Google Scholar] [CrossRef]
- Muñoz-Cruz, S.; Mendoza-Rodríguez, Y.; Nava-Castro, K.E.; Yepez-Mulia, L.; Morales-Montor, J. Gender-related effects of sex steroids on histamine release and FcεRI expression in rat peritoneal mast cells. J. Immunol. Res. 2015, 2015, 351829. [Google Scholar] [CrossRef]
- Padilla, L.; Reinicke, K.; Montesino, H.; Villena, F.; Asencio, H.; Cruz, M.; Rudolph, M.I. Histamine content and mast cells distribution in mouse uterus: The effect of sexual hormones, gestation and labor. Cell. Mol. Biol. 1990, 36, 93–100. [Google Scholar]
- Vliagoftis, H.; Dimitriadou, V.; Theoharides, T.C. Progesterone triggers selective mast cell secretion of 5-hydroxytryptamine. Int. Arch. Allergy Appl. Immunol. 1990, 93, 113–119. [Google Scholar] [CrossRef]
- Kim, M.S.; Chae, H.J.; Shin, T.Y.; Kim, H.M.; Kim, H.R. Estrogen regulates cytokine release in human mast cells. Immunopharmacol. Immunotoxicol. 2001, 23, 495–504. [Google Scholar] [CrossRef]
- Osman, M. Therapeutic implications of sex differences in asthma and atopy. Arch. Dis. Child. 2003, 88, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.J.; McKerr, G.; Dong, Z.; Higgins, C.A.; Carson, J.; Yang, Z.Q.; Hannigan, B.M. Expression of oestrogen and progesterone receptors by mast cells alone, but not lymphocytes, macrophages or other immune cells in human upper airways. Thorax 2001, 56, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Chandler, M.H.; Schuldheisz, S.; Phillips, B.A.; Muse, K.N. Premenstrual asthma: The effect of estrogen on symptoms, pulmonary function, and beta 2-receptors. Pharmacotherapy 1997, 17, 224–234. [Google Scholar] [PubMed]
- Salam, M.T.; Wenten, M.; Gilliland, F.D. Endogenous and exogenous sex steroid hormones and asthma and wheeze in young women. J. Allergy Clin. Immunol. 2006, 117, 1001–1007. [Google Scholar] [CrossRef]
- Tan, K.S.; McFarlane, L.C.; Lipworth, B.J. Modulation of airway reactivity and peak flow variability in asthmatics receiving the oral contraceptive pill. Am. J. Respir. Crit. Care Med. 1997, 155, 1273–1277. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Asthma. 2023. Available online: www.ginasthma.com (accessed on 30 April 2023).
- Fukunaga, K. [Asthma Prevention and Management Guidelines 2021]. Arerugi 2023, 72, 214–219. [Google Scholar] [CrossRef]
- Makoni, M. Guidelines might help reduce the burden of asthma in African children. Lancet Respir. Med. 2022, 10, e83–e84. [Google Scholar] [CrossRef]
- GEMA 5.1. Guía Española para el Manejo del Asma 2023. 6 May 2023. Available online: www.gemasma.com (accessed on 30 April 2023).
- Cloutier, M.M.; Baptist, A.P.; Blake, K.V.; Brooks, E.G.; Bryant-Stephens, T.; DiMango, E.; Dixon, A.E.; Elward, K.S.; Hartert, T.; Krishnan, J.A.; et al. 2020 Focused Updates to the Asthma Management Guidelines: A Report from the National Asthma Education and Prevention Program Coordinating Committee Expert Panel Working Group. J. Allergy Clin. Immunol. 2020, 146, 1217–1270. [Google Scholar] [CrossRef]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef]
- Canonica, G.W.; Ferrando, M.; Baiardini, I.; Puggioni, F.; Racca, F.; Passalacqua, G.; Heffler, E. Asthma: Personalized and precision medicine. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 51–58. [Google Scholar] [CrossRef]
- Pelaia, C.; Calabrese, C.; Terracciano, R.; de Blasio, F.; Vatrella, A.; Pelaia, G. Omalizumab, the first available antibody for biological treatment of severe asthma: More than a decade of real-life effectiveness. Ther. Adv. Respir. Dis. 2018, 12, 1753466618810192. [Google Scholar] [CrossRef]
- Genentech USA INPC. Xolair® (Omalizumab) Prescribing Information: 2023. 2023. Available online: https://www.xolairhcp.com (accessed on 23 May 2023).
- Riccio, A.M.; Dal Negro, R.W.; Micheletto, C.; De Ferrari, L.; Folli, C.; Chiappori, A.; Canonica, G.W. Omalizumab modulates bronchial reticular basement membrane thickness and eosinophil infiltration in severe persistent allergic asthma patients. Int. J. Immunopathol. Pharmacol. 2012, 25, 475–484. [Google Scholar] [CrossRef]
- Hochhaus, G.; Brookman, L.; Fox, H.; Johnson, C.; Matthews, J.; Ren, S.; Deniz, Y. Pharmacodynamics of omalizumab: Implications for optimised dosing strategies and clinical efficacy in the treatment of allergic asthma. Curr. Med. Res. Opin. 2003, 19, 491–498. [Google Scholar] [CrossRef]
- Casale, T.B.; Luskin, A.T.; Busse, W.; Zeiger, R.S.; Trzaskoma, B.; Yang, M.; Griffin, N.M.; Chipps, B.E. Omalizumab Effectiveness by Biomarker Status in Patients with Asthma: Evidence From PROSPERO, A Prospective Real-World Study. J. Allergy Clin. Immunol. Pract. 2019, 7, 156–164.e1. [Google Scholar] [CrossRef] [PubMed]
- Canonica, G.W.; Rottoli, P.; Bucca, C.; Zappa, M.C.; Michetti, G.; Macciocchi, B.; Caruso, C.; Santus, P.; Bartezaghi, M.; Rigoni, L. Improvement of patient-reported outcomes in severe allergic asthma by omalizumab treatment: The real life observational PROXIMA study. World Allergy Organ. J. 2018, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, M.; Yang, W.H.; Hebert, J.; de Takacsy, F.; Stril, J.L. The real world effect of omalizumab add on therapy for patients with moderate to severe allergic asthma: The ASTERIX Observational study. PLoS ONE 2017, 12, e0183869. [Google Scholar] [CrossRef] [PubMed]
- Barnes, N.; Menzies-Gow, A.; Mansur, A.H.; Spencer, D.; Percival, F.; Radwan, A.; Niven, R. Effectiveness of omalizumab in severe allergic asthma: A retrospective UK real-world study. J. Asthma 2013, 50, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Humbert, M.; Gibson, P.G.; Kostikas, K.; Jaumont, X.; Pfister, P.; Nissen, F. Real-World Effectiveness of Omalizumab in Severe Allergic Asthma: A Meta-Analysis of Observational Studies. J. Allergy Clin. Immunol. Pract. 2021, 9, 2702–2714. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, K.M.; MacDonald, K.; Abraham, I.; Alhossan, A.; Lee, C.S. ‘Real-world’ effectiveness of omalizumab in adults with severe allergic asthma: A meta-analysis. Expert Rev. Clin. Immunol. 2021, 17, 73–83. [Google Scholar] [CrossRef]
- Torres-Duque, C.A.; Ocampo-Gómez, J.; Castillo, M.M.; Cano-Rosales, D.; Giraldo-Montoya, Á.; Rodríguez, F.; Palacios-Ortega, I.; Durán-Silva, M.; Reynales, H.; García, E.; et al. Real-world effectiveness of omalizumab for severe allergic asthma treatment in Colombia. BMC Pulm. Med. 2022, 22, 447. [Google Scholar] [CrossRef] [PubMed]
- Braunstahl, G.J.; Chen, C.W.; Maykut, R.; Georgiou, P.; Peachey, G.; Bruce, J. The eXpeRience registry: The ‘real-world’ effectiveness of omalizumab in allergic asthma. Respir. Med. 2013, 107, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Kirchnerova, O.R.; Valena, T.; Novosad, J.; Terl, M.; Czech eXpeRience Study, G. Real-world effectiveness and safety of omalizumab in patients with uncontrolled severe allergic asthma from the Czech Republic. Postepy Dermatol. Alergol. 2019, 36, 34–43. [Google Scholar] [CrossRef]
- Su, N.; Zhi, L.; Liu, F.; Wang, Y.; Zhang, Q.; Liu, X.; Wang, X.; Hao, G.; Zhang, X.; Hu, Q.; et al. Real-World Safety and Effectiveness of Omalizumab in Moderate to Severe Allergic Asthma Patients in China: A Post-Authorization Study. J. Asthma Allergy 2023, 16, 625–636. [Google Scholar] [CrossRef]
- Emma, R.; Morjaria, J.B.; Fuochi, V.; Polosa, R.; Caruso, M. Mepolizumab in the management of severe eosinophilic asthma in adults: Current evidence and practical experience. Ther. Adv. Respir. Dis. 2018, 12, 1753466618808490. [Google Scholar] [CrossRef]
- Chupp, G.L.; Bradford, E.S.; Albers, F.C.; Bratton, D.J.; Wang-Jairaj, J.; Nelsen, L.M.; Trevor, J.L.; Magnan, A.; Brinke, A.T. Efficacy of mepolizumab add-on therapy on health-related quality of life and markers of asthma control in severe eosinophilic asthma (MUSCA): A randomised, double-blind, placebo-controlled, parallel-group, multicentre, phase 3b trial. Lancet Respir. Med. 2017, 5, 390–400. [Google Scholar] [CrossRef]
- Bel, E.H.; Wenzel, S.E.; Thompson, P.J.; Prazma, C.M.; Keene, O.N.; Yancey, S.W.; Ortega, H.G.; Pavord, I.D. Oral glucocorticoid-sparing effect of mepolizumab in eosinophilic asthma. N. Engl. J. Med. 2014, 371, 1189–1197. [Google Scholar] [CrossRef]
- Canonica, G.W.; Colombo, G.L.; Bruno, G.M.; Di Matteo, S.; Martinotti, C.; Blasi, F.; Bucca, C.; Crimi, N.; Paggiaro, P.; Pelaia, G.; et al. Shadow cost of oral corticosteroids-related adverse events: A pharmacoeconomic evaluation applied to real-life data from the Severe Asthma Network in Italy (SANI) registry. World Allergy Organ. J. 2019, 12, 100007. [Google Scholar] [CrossRef]
- Heffler, E.; Bagnasco, D.; Canonica, G.W. Strategies to reduce corticosteroid-related adverse events in asthma. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 61–67. [Google Scholar] [CrossRef]
- Deeks, E.D. Mepolizumab: A Review in Eosinophilic Asthma. BioDrugs 2016, 30, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Sahota, J.; Robinson, D.S. Update on new biologics for intractable eosinophilic asthma: Impact of reslizumab. Drug Des. Dev. Ther. 2018, 12, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Zangrilli, J.E.; Wechsler, M.E.; Bateman, E.D.; Brusselle, G.G.; Bardin, P.; Murphy, K.; Maspero, J.F.; O’Brien, C.; Korn, S. Reslizumab for inadequately controlled asthma with elevated blood eosinophil counts: Results from two multicentre, parallel, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet Respir. Med. 2015, 3, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Calabrese, C.; Vatrella, A.; Busceti, M.T.; Garofalo, E.; Lombardo, N.; Terracciano, R.; Pelaia, G. Benralizumab: From the Basic Mechanism of Action to the Potential Use in the Biological Therapy of Severe Eosinophilic Asthma. Biomed. Res. Int. 2018, 2018, 4839230. [Google Scholar] [CrossRef] [PubMed]
- Ghazi, A.; Trikha, A.; Calhoun, W.J. Benralizumab--a humanized mAb to IL-5Ralpha with enhanced antibody-dependent cell-mediated cytotoxicity—A novel approach for the treatment of asthma. Expert Opin. Biol. Ther. 2012, 12, 113–118. [Google Scholar] [CrossRef]
- Menzella, F.; Latorre, M.; Ruggiero, P.; Bagnasco, D.; Heffler, E. Reduction of oral corticosteroids in patients with severe eosinophilic asthma treated with Benralizumab: Could it represent a marker of treatment efficacy? Expert Opin Biol Ther. 2019, 19, 601–606. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, J.; Chen, Y. Efficacy and safety of treatment with benralizumab for eosinophilic asthma. Int Immunopharmacol. 2022, 111, 109131. [Google Scholar] [CrossRef]
- Bagnasco, D.; Ferrando, M.; Varricchi, G.; Passalacqua, G.; Canonica, G.W. A Critical Evaluation of Anti-IL-13 and Anti-IL-4 Strategies in Severe Asthma. Int. Arch. Allergy Immunol. 2016, 170, 122–131. [Google Scholar] [CrossRef]
- Balboul, S.; Kahn, J.; Tracy, A.; Peacock, A.; Cline, A. The Application of Dupilumab to Pediatric Patients Aged 6-11yrs with Moderate-to-Severe Atopic Dermatitis Whose Disease is Not Adequately Controlled: The Clinical Data so Far. Drug Des. Devel. Ther. 2023, 17, 1323–1327. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Yosipovitch, G.; Simpson, E.L.; Kim, B.S.; Wu, J.J.; Eckert, L.; Guillemin, I.; Chen, Z.; Ardeleanu, M.; Bansal, A.; et al. Dupilumab treatment results in early and sustained improvements in itch in adolescents and adults with moderate to severe atopic dermatitis: Analysis of the randomized phase 3 studies SOLO 1 and SOLO 2, AD ADOL, and CHRONOS. J. Am. Acad. Dermatol. 2020, 82, 1328–1336. [Google Scholar] [CrossRef]
- Senner, S.; Seegraber, M.; Frey, S.; Kendziora, B.; Eicher, L.; Wollenberg, A. Dupilumab for the treatment of adolescents with atopic dermatitis. Expert Rev. Clin. Immunol. 2020, 16, 641–650. [Google Scholar] [CrossRef]
- Ferrante, G.; Tenero, L.; Piazza, M.; Piacentini, G. Severe pediatric asthma therapy: Dupilumab. Front. Pediatr. 2022, 10, 963610. [Google Scholar] [CrossRef]
- Rabe, K.F.; Nair, P.; Brusselle, G.; Maspero, J.F.; Castro, M.; Sher, L.; Zhu, H.; Hamilton, J.D.; Swanson, B.N.; Khan, A.; et al. Efficacy and Safety of Dupilumab in Glucocorticoid-Dependent Severe Asthma. N. Engl. J. Med. 2018, 378, 2475–2485. [Google Scholar] [CrossRef]
- Albrecht, T.; Sailer, M.M.; Capitani, F.; van Schaik, C.; Lowenheim, H.; Becker, S. Real-world evidence for the effectiveness and safety of dupilumab in patients with CRSwNP after 1 year of therapy. World Allergy Organ. J. 2023, 16, 100780. [Google Scholar] [CrossRef]
- De Corso, E.; Pasquini, E.; Trimarchi, M.; La Mantia, I.; Pagella, F.; Ottaviano, G.; Garzaro, M.; Pipolo, C.; Torretta, S.; Seccia, V.; et al. Dupilumab in the treatment of severe uncontrolled chronic rhinosinusitis with nasal polyps (CRSwNP): A multicentric observational Phase IV real-life study (DUPIREAL). Allergy 2023. [Google Scholar] [CrossRef]
- Greuter, T.; Schoepfer, A.M. Dupilumab in Patients with Eosinophilic Esophagitis. N. Engl. J. Med. 2023, 388, 955–956. [Google Scholar] [CrossRef]
- Dellon, E.S.; Rothenberg, M.E.; Collins, M.H.; Hirano, I.; Chehade, M.; Bredenoord, A.J.; Lucendo, A.J.; Spergel, J.M.; Aceves, S.; Sun, X.; et al. Dupilumab in Adults and Adolescents with Eosinophilic Esophagitis. N. Engl. J. Med. 2022, 387, 2317–2330. [Google Scholar] [CrossRef]
- Maspero, J.F.; Cardona, G.; Schonffeldt, P.; Tolcachier, A.; González-Diaz, S.N.; Yañez, A.; Galvao, C.E.; Msihid, J.; Gall, R.; Siddiqui, S.; et al. Dupilumab efficacy and safety in Latin American patients with uncontrolled, moderate-to-severe asthma: Phase 3 LIBERTY ASTHMA QUEST study. J. Asthma 2023, 60, 981–990. [Google Scholar] [CrossRef]
- Hopkins, C.; Buchheit, K.M.; Heffler, E.; A Cohen, N.; Olze, H.; Khan, A.H.; Msihid, J.; Siddiqui, S.; Nash, S.; A Jacob-Nara, J.; et al. Improvement in Health-Related Quality of Life with Dupilumab in Patients with Moderate-to-Severe Asthma with Comorbid Chronic Rhinosinusitis with/without Nasal Polyps: An Analysis of the QUEST Study. J. Asthma Allergy 2022, 15, 767–773. [Google Scholar] [CrossRef]
- Busse, W.W.; Pavord, I.D.; Siddiqui, S.; Khan, A.H.; Praestgaard, A.; Nash, S.; Jacob-Nara, J.A.; Rowe, P.J.; Deniz, Y. Dupilumab Improves Outcomes in Patients with Chronic Rhinosinusitis with Nasal Polyps and Coexisting Asthma Irrespective of Baseline Asthma Characteristics. J. Asthma Allergy 2023, 16, 411–419. [Google Scholar] [CrossRef]
- Dinardo, G.; Indolfi, C.; Klain, A.; Decimo, F.; Miraglia Del Giudice, M. Treatment of severe asthma: Fast action of dupilumab in the pediatric setting. Minerva Pediatr. 2023, 75, 312–313. [Google Scholar] [CrossRef]
- Papadopoulos, N.G.; Szefler, S.J.; Bacharier, L.B.; Maspero, J.F.; Domingo, C.; Fiocchi, A.; Lee, J.K.; Daizadeh, N.; Lederer, D.J.; Hardin, M.; et al. Assessment of dupilumab in children with moderate-to-severe type 2 asthma with or without evidence of allergic asthma. Allergy 2023, 78, 2157–2167. [Google Scholar] [CrossRef]
- Yang, D.Y.; Li, L.; Lu, T.; Jing, W.W.; Liu, X.; Li, X.L. Efficacy and safety of dupilumab in pediatric patients with moderate to severe atopic dermatitis: A real-world study. Arch. Dermatol. Res. 2023, 315, 467–472. [Google Scholar] [CrossRef]
- Simpson, E.L.; Paller, A.S.; Siegfried, E.C.; Thaçi, D.; Wollenberg, A.; Cork, M.J.; Marcoux, D.; Huang, R.; Chen, Z.; Rossi, A.B.; et al. Dupilumab Demonstrates Rapid and Consistent Improvement in Extent and Signs of Atopic Dermatitis Across All Anatomical Regions in Pediatric Patients 6 Years of Age and Older. Dermatol. Ther. 2021, 11, 1643–1656. [Google Scholar] [CrossRef]
- Corren, J.; Menzies-Gow, A.; Chupp, G.; Israel, E.; Korn, S.; Cook, B.; Ambrose, C.S.; Hellqvist, Å.; Roseti, S.L.; Molfino, N.A.; et al. Efficacy of Tezepelumab in Severe, Uncontrolled Asthma: Pooled Analysis of PATHWAY and NAVIGATOR Studies. Am. J. Respir. Crit. Care Med. 2023, 208, 13–24. [Google Scholar] [CrossRef]
- Roy, P.; Rafa, Z.I.; Haque, S.N.; Tasha, T.; Arko, S.B.; Agrawal, H.; Razu, I.; Parisapogu, A.; Maisha, S.; Siddique, M.A.; et al. The Impact of Tezepelumab in Uncontrolled Severe Asthma: A Systematic Review of Randomized Controlled Trials. Cureus 2022, 14, e32156. [Google Scholar] [CrossRef]
- Corren, J.; Parnes, J.R.; Wang, L.; Mo, M.; Roseti, S.L.; Griffiths, J.M.; van der Merwe, R. Tezepelumab in Adults with Uncontrolled Asthma. N. Engl. J. Med. 2017, 377, 936–946. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Colice, G.; Griffiths, J.M.; Almqvist, G.; Skärby, T.; Piechowiak, T.; Kaur, P.; Bowen, K.; Hellqvist, Å.; Mo, M.; et al. SOURCE: A phase 3, multicentre, randomized, double-blind, placebo-controlled, parallel group trial to evaluate the efficacy and safety of tezepelumab in reducing oral corticosteroid use in adults with oral corticosteroid dependent asthma. Respir. Res. 2020, 21, 264. [Google Scholar] [CrossRef]
- Menzies-Gow, A.; Bourdin, A.; Chupp, G.; Israel, E.; Hellqvist, Å.; Hunter, G.; Roseti, S.L.; Ambrose, C.S.; Llanos, J.-P.; Cook, B.; et al. Effect of tezepelumab on healthcare utilization in patients with severe, uncontrolled asthma: The NAVIGATOR study. Ann. Allergy Asthma Immunol. 2023. [Google Scholar] [CrossRef]
- Chagas, G.C.L.; Xavier, D.; Gomes, L.; Ferri-Guerra, J.; Oquet, R.E.H. Effects of Tezepelumab on Quality of Life of Patients with Moderate-to-Severe, Uncontrolled Asthma: Systematic Review and Meta-Analysis. Curr. Allergy Asthma Rep. 2023. [Google Scholar] [CrossRef] [PubMed]
- Lommatzsch, M.; Suhling, H.; Korn, S.; Bergmann, K.C.; Schreiber, J.; Bahmer, T.; Rabe, K.F.; Buhl, R.; Virchow, J.C.; Milger, K. Safety of combining biologics in severe asthma: Asthma-related and unrelated combinations. Allergy 2022, 77, 2839–2843. [Google Scholar] [CrossRef] [PubMed]
- Rappoport, N.; Shamir, R. Multi-omic and multi-view clustering algorithms: Review and cancer benchmark. Nucleic Acids Res. 2018, 46, 10546–10562. [Google Scholar] [CrossRef]
- Dugourd, A.; Kuppe, C.; Sciacovelli, M.; Gjerga, E.; Gabor, A.; Emdal, K.B.; Vieira, V.; Bekker-Jensen, D.B.; Kranz, J.; Bindels, E.M.J.; et al. Causal integration of multi-omics data with prior knowledge to generate mechanistic hypotheses. Mol. Syst. Biol. 2021, 17, e9730. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, M.I.; Neerincx, A.H.; Vijverberg, S.J.; Kraneveld, A.D.; Maitland-van der Zee, A.H. Omics for the future in asthma. Semin. Immunopathol. 2020, 42, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Bérubé, J.C.; Bossé, Y. Future clinical implications emerging from recent genome-wide expression studies in asthma. Expert Rev. Clin. Immunol. 2014, 10, 985–1004. [Google Scholar] [CrossRef]
- Bossé, Y. Genome-wide expression quantitative trait loci analysis in asthma. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 487–494. [Google Scholar] [CrossRef]
- Lopes-Pacheco, M.; Rocco, P.R.M. Functional enhancement strategies to potentiate the therapeutic properties of mesenchymal stromal cells for respiratory diseases. Front. Pharmacol. 2023, 14, 1067422. [Google Scholar] [CrossRef]
- Zhou, T.; Yuan, Z.; Weng, J.; Pei, D.; Du, X.; He, C.; Lai, P. Challenges and advances in clinical applications of mesenchymal stromal cells. J. Hematol. Oncol. 2021, 14, 24. [Google Scholar] [CrossRef]
- Silva, L.H.A.; Antunes, M.A.; Dos Santos, C.C.; Weiss, D.J.; Cruz, F.F.; Rocco, P.R.M. Strategies to improve the therapeutic effects of mesenchymal stromal cells in respiratory diseases. Stem Cell Res. Ther. 2018, 9, 45. [Google Scholar] [CrossRef]
- Abreu, S.C.; Lopes-Pacheco, M.; Weiss, D.J.; Rocco, P.R.M. Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Lung Diseases: Current Status and Perspectives. Front. Cell Dev. Biol. 2021, 9, 600711. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Kang, K.S. Functional enhancement strategies for immunomodulation of mesenchymal stem cells and their therapeutic application. Stem Cell Res. Ther. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Granton, J.; Langleben, D.; Kutryk, M.B.; Camack, N.; Galipeau, J.; Courtman, D.W.; Stewart, D.J. Endothelial NO-Synthase Gene-Enhanced Progenitor Cell Therapy for Pulmonary Arterial Hypertension: The PHACeT Trial. Circ. Res. 2015, 117, 645–654. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Uribe, V.; Romero-Tapia, S.J.; Castro-Rodriguez, J.A. Asthma Phenotypes in the Era of Personalized Medicine. J. Clin. Med. 2023, 12, 6207. https://doi.org/10.3390/jcm12196207
Gonzalez-Uribe V, Romero-Tapia SJ, Castro-Rodriguez JA. Asthma Phenotypes in the Era of Personalized Medicine. Journal of Clinical Medicine. 2023; 12(19):6207. https://doi.org/10.3390/jcm12196207
Chicago/Turabian StyleGonzalez-Uribe, Victor, Sergio J. Romero-Tapia, and Jose A. Castro-Rodriguez. 2023. "Asthma Phenotypes in the Era of Personalized Medicine" Journal of Clinical Medicine 12, no. 19: 6207. https://doi.org/10.3390/jcm12196207
APA StyleGonzalez-Uribe, V., Romero-Tapia, S. J., & Castro-Rodriguez, J. A. (2023). Asthma Phenotypes in the Era of Personalized Medicine. Journal of Clinical Medicine, 12(19), 6207. https://doi.org/10.3390/jcm12196207