1. Introduction
In female malignancies, breast cancer (BC) is considered to be the most frequently diagnosed cancer [
1]. For cancer patients, distal metastases and resistance to therapeutics are the leading causes of death [
2]. Biologically, distant cancer metastasis involves a variety of complex mechanisms, such as EMT, migration, invasion, angiogenesis, and adhesion [
3,
4]. Interpreting these mechanisms can be beneficial for discovering new therapeutic regimens and markers.
Lymphatic metastasis is the most common form of BC progression [
5]. In our previous research, the differentially expressed miRNAs, lncRNAs, and mRNAs in mouse hepatocarcinoma Hca-F (with high lymphatic metastasis potential) and Hca-P (with low lymphatic metastasis potential) cell lines were screened using a high-throughput RNA sequencing method. Among them, miR-330-3p, lnc045874, and EREG have shown significantly different expressions between the two cell lines. Murine lnc045874 is the most conservative homology of human NONHSAT021545 (lnc021545). Meanwhile, lnc021545 conserved same the binding site as miR-330-3p. TargetScan (
http://www.targetscan.org accessed on 1 March 2020) analysis suggested that miR-330-3p has two combining sites with the 3′-untranslated region (3′-UTR) of EREG and one combining site with lnc045874. We aimed to determine whether these three molecules could synergistically affect the progression of BC by modulating the lymphatic metastasis and evaluated the clinical feature of BC patients.
The microRNAs (miRNAs) are a type of small, non-coding RNA composed of 18–24 nucleotides. The miRNAs can negatively modulate genes’ expressions by binding their 3′-UTRs, which act as crucial effectors in the malignant behaviors of tumor cells [
6]. MiR-330-3p is located on chromosome 19q12.32 [
7] and is bound to the malignant behaviors of cancerous cells; It plays a moderator role in different cancers [
8]. Long non-coding RNAs (lncRNAs) are a type of RNA transcript with a length longer than 200 nucleotides without protein-coding ability [
9]. LncRNAs have been demonstrated to modulate the biological functions of mRNAs by combing miRNAs [
10]. Lnc021545 (NONCODE ID: NONHSAT021545.2) is located on human chromosome 11 with a length of 3149 bases; the exact role of lnc021545 in BC remains elusive. Epiregulin (EREG) is located on chromosome 4q13.3 and is a member of the epidermal growth factor family [
11]. EREG acts as a crucial effector in the modulation of angiogenesis [
12], inflammation [
13], drug resistance [
14], and metastasis [
15], whereas the effect of EREG in the origination and progression of BC remains unconfident.
The development of a disease is a complex process caused by the dysregulation of multiple genes’ functions rather than by the dysregulation of a single or an isolated gene’s functions [
16]. With the identification of a mass of cancer-related genes and their functions, the studies that coordinated function of multi-genes influence the prognosis and treatment of cancer are increasingly emerging. In the research of BC, the expression levels of 21 genes associated with the biological characteristics of BC are detected via RT-PCR, and the recurrence score (RS) is obtained using an algorithm based on previous clinical studies [
17]. The RS has been integrated into the National Comprehensive Cancer Network clinical guidelines for treatment and predictive prognosis of hormone-receptor-positive and HER-2-negative diseases [
18]. By studying the effects of lnc021545, miR-330-3p, and EREG in BC, we will explore whether these three molecules’ coordinated functions could affect BC’s prognosis and treatment.
As part of this study, we established a new lnc021545-miR-330-3p-EREG axis regulatory network in BC. We found that miR-330-3p was upregulated, while lnc021545 and EREG were downregulated in BC patients’ samples. Moreover, lnc021545 and EREG act as tumor suppressors in regulating the metastasis of BC cells, while miR-330-3p acts as a tumor promoter and showed opposite effects on BC cells. Furthermore, lnc021545/miR-330-3p affects the EMT process of BC cells by regulating EREG expression. Our study demonstrates lnc021545-miR-330-3p-EREG axis regulatory network in BC progression and provides a new clue to the treatment and prognosis of BC.
2. Materials and Methods
2.1. Tissue Samples and Cell Culture
Fifty pairs of BC tissues with their one-to-one corresponding adjacent non-tumor tissues were collected from patients who underwent surgical resection at the Second Affiliated Hospital of Dalian Medical University between 2015 and 2019. The study was approved by the Medical Ethics Committee of Dalian Medical University (approval number 2019-069). The informed consent forms were signed by all participants. After resection, the tissue specimens were stored at −80 °C until use. Human BC cell lines MCF-7 and T47D were obtained from the American Type Culture Collection. Both cell lines were cultured in RPMI 1640 (Gibco, New York, NY, USA) supplemented with 15% fetal bovine serum (FBS, TransGen, Beijing, China) and 100 U/mL penicillin/streptomycin (Gibco) at 37 °C with 5% CO2.
2.2. Bioinformatic Analysis
The data of miRNA-seq were downloaded from The Cancer Genome Atlas (TCGA,
https://tcgadata.nci.nih.gov/tcga accessed on 18 September 2022). We also queried the Gene Expression Omnibus (GEO,
https://www.ncbi.nlh.gov/geo accessed on 20 September 2022) database for miRNA and mRNA profiling by arrays. We analyzed the expression data of 1103 BC tissues and 104 non-tumor tissues from the TCGA-BRCA dataset by employing R package “edgeR” to identify and normalize the differentially expressed miRNAs. GSE22216 and GSE40267 were utilized for survival analysis according to miR-330-3p expression changes. To evaluate the prognostic value of EREG, survival analysis was performed using the web tool KMplot (
http://kmplot.com accessed on 26 September 2022). Gene set enrichment analysis (GSEA) was executed on the GSE3494 dataset to analyze the associated functional gene sets in BC pathogenesis.
2.3. Transient Transfection
Two small interfering RNAs of lnc021545 (si-lnc021545), three small interfering RNAs of EREG (si-EREG), siRNA negative control (si-NC), mimic or inhibitor of miR-330-3p, miRNA negative control (NC), PCDH-EF1-MCS-T2A-Puro-EREG (PCDH-EREG), and PCDH-EF1-MCS-T2A-Puro vector (PCDH) were used, and the sequences of si-RNAs for EREG and lnc021545 knockdown are shown in
Table S1.
In total, 3 × 105 MCF-7 and T47D cells were seeded per well and incubated at 37 °C with 5% CO2 for 12 h. To downregulate lnc021545 in cells, 3.5 µL si-lnc021545-1 and 3.5 µL si-lnc021545-2 or 7 µL si-NC with a consistency of 20 µM and the supplementary of 6 µL Lipofectamine™ 2000 (Lipo2000, Invitrogen, Waltham, MA, USA) were mixed to form a transfection mixture and kept still for 20 min at room temperature (RT). Then, 5 µL miR-330-3p mimic, mimic NC, miR-330-3p inhibitor or inhibitor NC with a consistency of 20 µM, and the supplementary of 5 µL Lipo2000 were mixed to form a transfection mixture to overexpress and downregulate miR-330-3p in cells. An amount of 1 µg PCDH-EREG or 1 µg PCDH was mixed with 50 µL serum-free medium, 5 µL Lipo2000 was mixed with 45 µL serum-free medium, and the above two mixtures were mixed in order to establish the cells of EREG over-expression and placed for 20 min at RT. 2.5 µL si-EREG-1, si-EREG-2, and si-EREG-3 or 7.5 µL si-NC with a consistency of 20 µM. A supplement of 6 µL Lipo2000 was mixed and placed for 20 min at RT as the transfection mixture to EREG downregulation. Each of the transfection mixtures were added into each group of MCF-7 or T47D cells. Then, the cells were cultured at 37 °C under 5% CO2 for 24 h or 48 h for further experiments.
2.4. Cell Proliferation Assay
The proliferation abilities of different group of MCF-7 and T47D cells were measured via MTT assay. Each group was seeded with a density of 3 × 103 cells/well in a 96-well plate (NEST, Wuxi, China). MTT reagent was added to each well in 200 μL increments at 24 h, 48 h, 72 h, 96 h, and 120 h. After 4 h, the MTT reagent was substituted with 150 μL dimethyl sulfoxide (Sigma, St. Louis, MO, USA). The absorbance of 492 nm was detected using a plate reader (Eppendorf, Hamburg, Germany) for the quantification of cell viability.
2.5. Cell Motility Assay
The motility ability of MCF-7 cells was measured by wound-healing experiments. In total, 6 × 105 of each group of MCF-7 cells were seeded into a 6-well plate (NEST) at 37 °C with 5% CO2 and cultured to 85–90% confluence. Then, a 200 μL sterilized tip drew vertical scratches on the monolayer cell surface. Detached and floated cells were washed with PBS, then cultured at 37 °C for 24 h. The widths of scratch wounds were obtained under a microscope (Olympus, Tokyo, Japan) at 100× magnification at 0 h, 24 h, and assessed using Image J software (version 2.3.0).
2.6. Cell Migration and Invasion Assay
The migration and invasion of cells were tested via transwell chamber assay. For migration assay, 1 × 104 transfected MCF-7 or T47D cells in 200 µL RPMI 1640 with serum-free media were seeded in the upper chamber of the transwell. For invasion assay, the chambers were layered in an extracellular matrix (Sigma). The upper chamber was assembled into the plate wells, and the bottom chamber was filled with 600 μL RPMI 1640 with 20% FBS and cultured for 48 h at 37 °C with 5% CO2. The migrative or invasive cells through the membrane were fixed for 30 min in absolute methanol and stained in 0.5% crystal violet stain solution (Solarbio, Beijing, China) for 1 h at RT, then imaged and counted with the microscope (Olympus) at 100× magnification.
2.7. Western Blotting (WB) Assay
RIPA lysis buffer was used to extract the total proteins of transfected BC cells. The quantitation of protein was evaluated using Bradford assay [
19]. After, the sample of protein (30 µg) was separated using 10% sodium dodecyl sulphate-polyacrylamide gels and transferred onto nitrocellulose membrane (PALL, Port Washington, NY, USA), followed by a blocking at RT with 5% skim milk for 2 h. Then, the membranes were incubated with the following primary antibodies: EREG (Abcam, Cambridge, MA, USA), E-cadherin (ProteinTech, Wuhan, China), N-cadherin (ProteinTech), vimentin (ProteinTech), Snail (ProteinTech), Slug (ProteinTech), and GAPDH (ProteinTech) overnight at 4 °C. After washing three times with TBST, the membranes were incubated in the secondary antibody (ProteinTech) at RT for 3 h and washed three times with TBST. The blots were developed by electrochemiluminescence (Advansta, San Jose, CA, USA) and imaged with the ChemiDoc MP imaging system (Bio-Rad, Hercules, CA, USA).
2.8. Luciferase Activity Assay
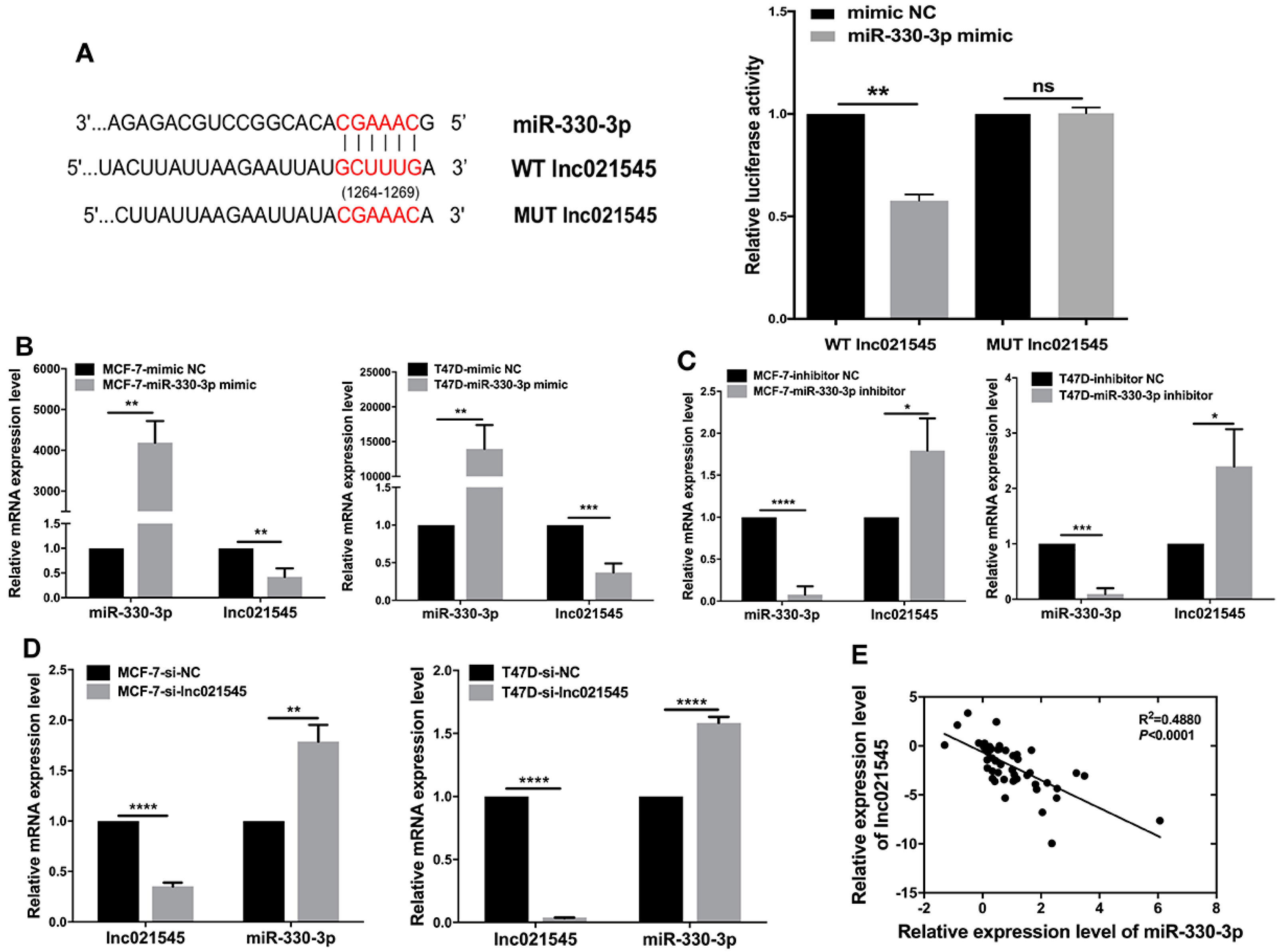
Wild-type (WT), mutant (MUT) lnc021545 3′-untranslated region (UTR), and EREG 3′-UTR were amplified and cloned into the luciferase reporter vector pmirGLO (Promega, Madison, WI, USA) to create pmirGLO lnc021545, pmirGLO EREG WT, and MUT plasmids. Then, 1 × 105 MCF-7 cells were co-transfected with 5 ng each of the corresponding plasmids and 20 pmol each of miR-330-3p mimics. A dual-luciferase reporter assay system (Promega) was used in this assay. Each well of luminometer plates was loaded with lysates and the firefly luciferase activity was detected through an EnSpire multifunctional microplate reader (PerkinElmer, Waltham, MA, USA). Each well of the luminometer plates was again loaded with 100 µL Stop&Glo reagent, and renilla luciferase activity was detected. The results were represented as normalized firefly luciferase activity/renilla luciferase activity values.
2.9. Quantitative Reverse Transcription PCR (qRT-PCR) Assay
Total RNA was obtained by Trizol
TM reagent (Invitrogen). Then, 1 µg of total RNA was reversely transcribed into high-quality cDNA with PrimeScript
TM 1st cDNA Synthesis (Takara, Kyoto, Japan). qRT-PCR was performed using the FastStart Universal SYBR Green Master (Roche, Basel, Switzerland) on a StepOne
TM Real-Time PCR System (ABI, Los Angeles, CA, USA). The relative expression levels of EREG and lnc021545 were quantified using β-actin as the internal reference, while the relative expression levels of miR-330-3p were analyzed against U6 reference. The relative expression levels of RNA were calculated using the 2
−ΔΔCT method. The primer sequences are listed in
Table S2.
2.10. Immunohistochemistry (IHC) Assay
The 2.5 μm paraffin slices were dewaxed by xylene, rehydrated with gradient ethanol, covered with 3% H
2O
2 for 10 min, washed with PBS three times, and incubated with goat anti-epiregulin polyclonal antibody (Invitrogen), rabbit anti estrogen receptor (ER) polyclonal antibody (Invitrogen), rabbit anti progesterone receptor (PR) monoclonal antibody (Invitrogen), or rabbit anti Ki-67 monoclonal antibody (Invitrogen) overnight at 4 °C. The slices were then nurtured with a biotin–streptavidin horseradish peroxidase detection system (ZSGB-BIO, Beijing, China) for 2 h and stained by DAB at RT. The slices were counterstained with hematoxylin, dehydrated with gradient ethanol, clarified with xylene, scanned with an Aperio GT450 (Leica, Wetzlar, Germany) at 200× magnification, and evaluated with an Aperio ImageScope (version 12.4.3.5008). The protein expression in the selected region was reflected by H-Score [
20].
2.11. Fluorescence In Situ Hybridization (FISH)
The 3 μm paraffin slices were heated for 30 min at 56 °C. The slices were deparaffinized for 10 min with xylene and tapped off xylene for 5 s in 100% ethanol, repeated twice. The slides were placed into gradient ethanol and deionized water, then into the boiled pre-treatment solution for 15 min. The slices were washed with 2 × SSC solution, incubated with proteinase K solution for 15 min, washed with 2 × SSC solution for 1 min, and dehydrated with each gradient ethanol for 3 min at RT. The specimen area was dropped to a 10 μL probe mix. The coverslip was sealed with rubber cement, and the slices were hybridized at 47 °C overnight. The slices were cleaned with 2 × SSC solution and dehydrated in each gradient ethanol for 3 min. Following air-drying, the slides were counterstained with 10 μL DAPI counterstain. After 15 min, the slides were scanned with fluorescence microscope (Olympus) at 100× magnification.
2.12. Amplification-Refractory Mutation System PCR (ARMS-PCR) Assay
A standard pathology methodology was used to select the tumor tissue with more than 30% tumor cells for DNA extraction. Fifty BC patients’ DNA extractions were carried out from FFPE tumor tissue according to the instructions of the DNA extraction kit (AmoyDx, Xiamen, China). The qualities of DNA extractions were assessed by NanoDrop (Thermo Fisher, Waltham, MA, USA). Then, 5 ng sample DNA was added into each PCR reaction mix tube of PIK3CA mutations (AmoyDx). The PIK3CA mutations were detected on a 7500 Real-Time PCR System (ABI). The PIK3CA mutations were analyzed using the “Results Interpretation” analysis module.
2.13. Statistical Analysis
SPSS 22.0 (IBM, Armonk, NY, USA) and GraphPad Prism 7 (GraphPad, La Jolla, CA, USA) were used for data statistical analyses. The differences and statistical significance between the two groups of data were assessed using Student’s t-test. The relation between any two molecules in BC tissues was assessed using the Pearson correlation coefficient. p value < 0.05 was regarded as significant.
4. Discussion
BC has become a significant health problem in modern society [
1]. In the past few years, the metastasis and poor prognosis of BC have presented a great challenge despite the progression of chemotherapy, endocrine therapy, and HER-2-targeted therapy [
21].
Lymphatic metastasis is the most important mode by which BC spreads, mainly including axillary lymphatic metastasis and parasternal lymphatic metastasis [
5]. The management of lymphatic metastasis has become an important link in the treatment of BC. Therefore, the research on lymphatic metastasis of BC has become an important part of the diagnosis and treatment of BC. In previous research, the high-throughput genetic sequencing assays using two murine hepatocarcinoma cell lines with different lymphatic metastasis potentials from our group (unpublished) prompted us to believe that miR-330-3p, lnc021545, and EREG might also form a regulation mechanism in human cancers. Through analyzing the TCGA-BRCA dataset, we noticed that miR-330-3p was overexpressed in BC tumorous tissues. Patients with low expression of miR-330-3p showed longer iDFS probabilities than those with high expression of miR-330-3p. We used KMplot to analyze the association between EREG expression and BC patients’ OS. We found that patients with high EREG expression showed better OS probabilities than the patients with low EREG expression. As a novel lncRNA, profile of lnc021545 in BC is unavailable. Herein, its role in BC was not analyzed. From the analysis of the database, it can be seen that miR-330-3p might be a tumor promoter and that EREG might be a tumor suppressor in BC. We herein investigated their mutual relationship and their correlation together with their function mechanism and potential clinical practice in BC metastasis.
MiRNAs play important roles in the progressions and treatments of multiple tumors [
22]. Earlier research has confirmed that miR-330-3p acted as a tumor promoter in multiple cancers. miR-330-3p acted as a tumor promoter in multiple cancers. It accelerated the migration and EMT of NSCLC cells via GGRIA3-TGFβ1 [
23]. In hepatocellular carcinoma, miR-330-3p enhanced cancer cells’ viability, migration, invasion, and apoptosis resistance by binding to BTG1 [
24]. MiR-330-3p targeted the CCBE1 and reduced its expression to promote the metastasis of BC [
25]. MiRNAs have multiple target genes and affect the progression of diseases by regulating the interaction of multiple genes, even in the same disease. For example, miR-200 activates the EMT by targeting HIPK1 or HDAC2 to promote the metastasis of BC [
26,
27]. Our study aims to demonstrate not only the role of miR-330-3p in BC metastasis but also its correlations with lnc021545 and EREG together with their axial regulation mechanism in BC metastasis.
LncRNAs are a type of new-style biomarkers and potential therapeutic targets in mediating BC progression and metastasis through regulating gene expression [
28,
29]. Currently, the research on lnc021545 in disease is limited. We found that lnc021545 knockdown promoted the metastatic abilities of BC cells rather than altering their proliferations. As a member of the EGR family, EREG can bind to each of EGFR and ErbB-4 through ligand-induced heterodimerization, which affects the progression of diseases through activating signaling pathways downstream of ErbB1/ErbB [
30]. EREG has largely been researched in the prognosis and drug resistance of BC [
31], but little is known about its action mechanism in BC progression. In BC cells, EREG also reduced the tendency of metastasis but had no effect on the proliferation. Therefore, lnc021545 and EREG deficiency promotes the progression of BC by increasing the metastatic behaviors of BC cells.
We discovered that the changes in the three molecules’ expression levels consistently conferred greater metastasis of BC cells rather than altering proliferation. We also found this phenomenon in BC tumorous samples; there was no correlation between any of the three molecules and the expression of Ki-67, which is a marker of cell proliferation and reflects the degree of proliferation in BC. Consequently, these data suggest that the three molecules regulate BC patients’ outcomes and treatment through mediating the metastasis of BC cells.
In recent studies, the association between miRNAs and lncRNA has been the most popular as lncRNAs can bind to miRNAs to antagonize miRNA-induced activity [
32]. We found that lnc021545 sponges miR-330-3p in BC cells. Bioinformatic analysis and dual-luciferase reporter assay demonstrated that lnc021545 targeted miR-330-3p to inhibit the latter’s expression. Our study also provided the relationship between lnc021545 and miR-330-3p in BC cells. Lnc021545 knockdown resulted in miR-330-3p upregulation, and the change in lnc021545 or miR-330-3p expression resulted in the corresponding change in miR-330-3p or lnc021545 expression in BC cells. MiR-330-3p and lnc021545 expression with a negative relationship was shown in the clinical data of 50 BC patients. By binding to each other to reversely regulate the expression of the other one, lnc021545 and miR-330-3p are functionalized in the metastasis of BC.
In order to prove that lnc021545, miR-330-3p, and EREG conform to this regulatory mode to modulate the metastasis of BC, we proved that lnc021545 could combine with miR-330-3p to affect BC’s metastasis in previous experiments, and we still need to prove that miR-330-3p and EREG could combine and interact with each other. Therefore, we proved that miR-330-3p could directly bound to 3′-UTR of EREG by using the online target gene prediction tool and the dual-luciferase reporters’ assay. Meanwhile, EREG expression levels was remarkably reversely regulated by the expression level change of miR-330-3p upregulated in BC cells. Further analysis confirmed that the mRNA and protein expression of EREG had a significant negative correlation with miR-330-3p in BC patients. We further showed that EREG mRNA and protein levels had an obviously positive association with lnc021545. Subsequently, EREG expression was remarkably suppressed in si-lnc021545 transfected cells. Our data demonstrate that lnc021545, miR-330-3p, and EREG are closely correlated in regulating the BC’s metastasis.
Cumulative evidence has pointed out that lncRNAs could modulate the biological activity of mRNA via hindering the miRNA from binding to its target mRNAs [
33]. Although we failed to overexpress lnc021545 in BC cells, its knockdown in BC cells still clearly led us to detect the increase of miR-330-3p expression and the decrease of EREG expression. The overexpression or knockdown of miR-330-3p level led to reduced or increased expressions of lnc021545 and EREG in BC cells. miR-330-3p was consistently inversely related to lnc021545 and EREG in BC tissues. EREG was positively related to lnc021545 in BC tissues. Meanwhile, we also confirmed that miR-330-3p had direct bindings between lnc021545 and EREG. In summary, lnc021545, miR-330-3p, and EREG form a negative feedback regulation pathway in promoting BC. Nevertheless, the functions and biological mechanism of the lnc021545-miR-330-3p-EREG axis in the metastasis of BC are unclear.
EMT has been demonstrated as an essential driver of carcinoma metastasis [
34,
35,
36]. Growing evidence has proved that miRNAs regulated the EMT by interacting with certain targeted mRNAs to impact carcinoma metastasis. miR-508-3p hindered the EMT by targeting ZEB1 to inhibit cell invasion in triple-negative breast cancer [
37]. Noyan et al. demonstrated that miR-770-5p targeted DNMT3A to suppress the EMT and the invasion of triple-negative breast cancer cells [
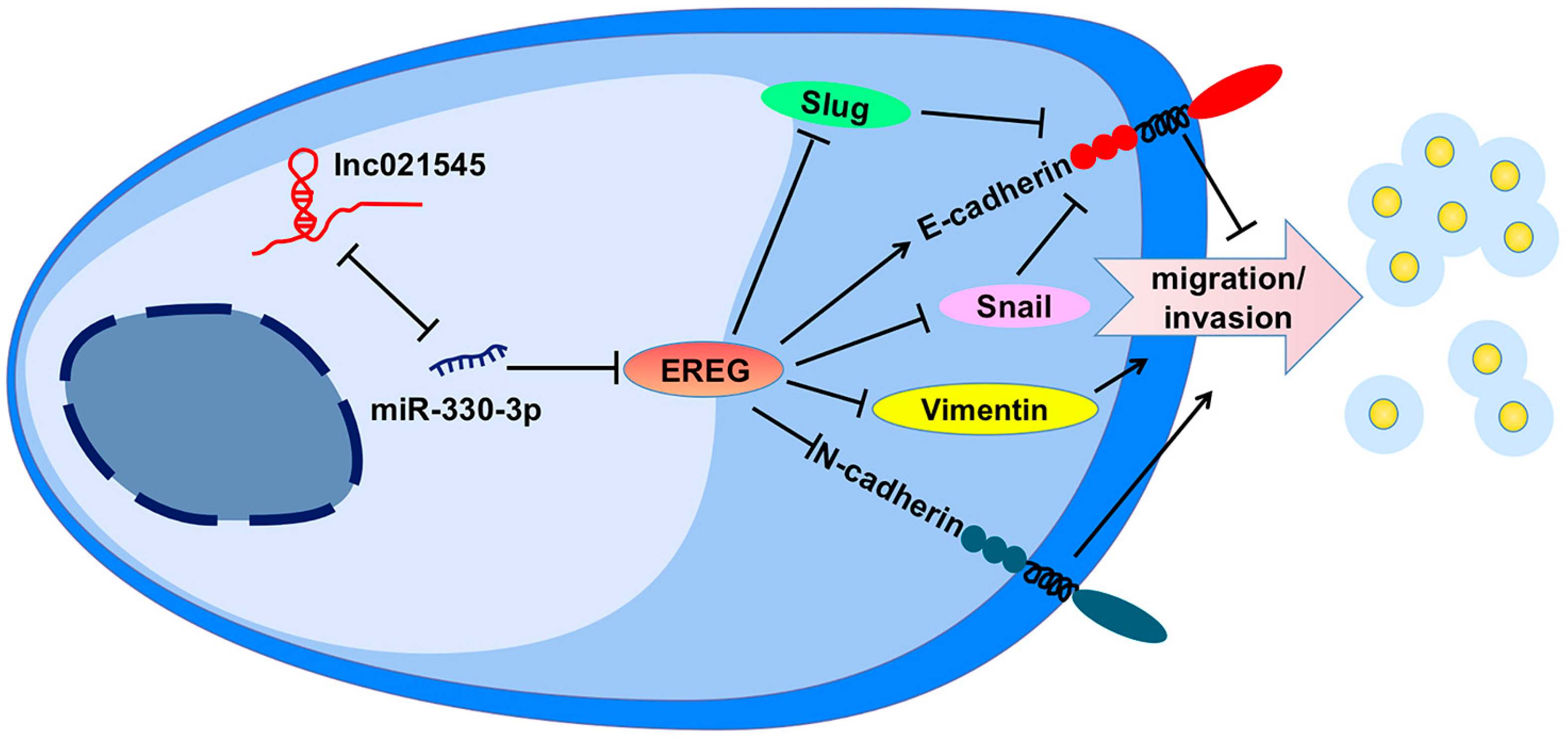
38]. In earlier research, we confirmed EREG was one of the downstream target genes of miR-330-3p. Specifically, we confirmed that miR-330-3p could reduce endogenous EREG protein expression to promote and restore the metastasis of BC cells. Their mechanisms for the changes in metastasis of BC were elusive. Single-gene GSEA database analysis indicated EREG had a negative correlation with EMT. We consistently detected that miR-330-3p overexpression increased the expressions of mesenchymal molecules vimentin and N-cadherin, repressors of the epithelial molecules Snail and Slug, but decreased the expression of the epithelial molecule E-cadherin in MCF-7 cells via directly targeting EREG. These data confirm that lnc021545 affects the metastasis of BC by inhibiting miR-330-3p regulation on EREG expression through EMT process (
Figure 9).
After we verified the mechanism by which the three molecules affect the metastasis of BC, we further observed the clinical effect of the three molecules on the therapeutic outcome and prognosis of BC patients. In the process of individualized treatment of BC, the expression of ER and PR, amplification of HER-2, and mutation of PIK3CA are some treatment-related indicators [
39,
40]. ER and PR expression levels correlate with endocrine therapy in BC and are the indexes used to develop therapeutic strategy and its efficacy [
39]. The detection of HER-2 amplification is very crucial for the prognosis and treatment of BC; the amplification of the HER-2 gene suggests that BC patients have a high degree of malignancy and poor prognosis, which is suitable for targeted therapy [
39]. Mutations of PIK3CA are one of the most common oncogenic mutations that occur in BC patients who may benefit from the treatments with PI3K inhibitors [
39].
In 50 BC patients, we observed that miR-330-3p expression was elevated in cancerous tissues and related to tumor size, TNM stage, lymph node metastasis, pathological stage, and HER-2 amplification while lnc021545 and EREG were decreased in BC tissues and exerted the opposite effects of miR-330-3p in the BC patient cohorts. We also established that high expression of miR-330-3p had a shorter iDFS and high expression of lnc021545 and EREG were conducive to lengthening BC patients’ iDFS. All of these suggest that miR-330-3p overexpression or lnc021545 or EREG downregulation contribute to the progression of BC and a poor prognosis while raising the chances for BC patients to benefit from trastuzumab therapy.
In summary, we found that miR-330-3p, lnc021545, and EREG had links to the prognosis and treatment of BC, but there were some differences among the three molecules. The process of tumor requires the cooperation of multiple genes [
16]. Therefore, we further analyzed the above results and tried to find an expressive mode to more accurately reflect the synergistic effect of the three molecules on the progression and treatment of BC. We established the expressive mode of the cooperative ratio according to the relationship and functions of the three molecules. Interestingly, we found that the patients with a low cooperative ratio had a more aggressive malignancy in BC patients, and the cooperative ratio was correlated with the size of the tumor, pathological TNM stage, lymphatic metastasis, pathological stage, and HER-2 amplification. It was concluded that the cooperative ratio had a good directivity for the malignant evaluation and clinical treatment of BC. More interestingly, the high cooperative ratio restrains the progression of BC and reduces the chance of trastuzumab therapy. Moreover, we also observed that a high cooperative ratio was conducive to lengthening BC patients’ iDFS, and the cooperative ratio was correlated with the months of iDFS. The results of these two parts showed that the cooperation ratio was closely tried with BC patients’ prognosis. Importantly, we found that the cooperative ratio of the three molecules was more comprehensive than each single molecule in the assessment of the patient’s malignant grade, recurrence, and death. Current work indicates that the roles and synergistic cooperation of lnc021545, miR-330-3p, and ERGE in regulating the malignant behaviors of BC cells as well as in evaluating the progression, therapeutic treatment, and prognosis of BC patients.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}