Association of KIR Genes with Middle East Respiratory Syndrome Coronavirus Infection in South Koreans

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.1.1. MERS Patients
2.1.2. Control Group
2.2. Genomic DNA Preparation
2.3. Multiplex PCR-SSP for KIR Genotyping and Gel Analysis of PCR Products
2.4. Statistical Analysis
3. Results
3.1. Frequencies of KIR Gene Clusters in Patients with MERS-CoV Infection and Controls
3.2. The Association between KIR Genes and Their Ligands in MERS Patients versus Control Groups
3.3. Association of KIR Haplotype and Genotype with MERS Risk
4. Discussion
4.1. KIR Genes
4.2. HLA Class I Ligands
4.3. MICA Molecules
4.4. Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, G.; Hu, Y.; Wang, Q.; Qi, J.; Gao, F.; Li, Y.; Zhang, Y.; Zhang, W.; Yuan, Y.; Bao, J.; et al. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 2013, 500, 227–231. [Google Scholar] [CrossRef]
- Pallesen, J.; Wang, N.; Corbett, K.S.; Wrapp, D.; Kirchdoerfer, R.N.; Turner, H.L.; Cottrell, C.A.; Becker, M.M.; Wang, L.; Shi, W.; et al. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA 2017, 114, E7348–E7357. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Sikkema, R.S.; Farag, E.; Islam, M.; Atta, M.; Reusken, C.; Al-Hajri, M.M.; Koopmans, M.P.G. Global status of Middle East respiratory syndrome coronavirus in dromedary camels: A systematic review. Epidemiol. Infect. 2019, 147, e84. [Google Scholar] [CrossRef] [PubMed]
- Cowling, B.J.; Park, M.; Fang, V.J.; Wu, P.; Leung, G.M.; Wu, J.T. Preliminary epidemiological assessment of MERS-CoV outbreak in South Korea, May to June 2015. Euro Surveill. Eur. Commun. Dis. Bull. 2015, 20, 7–13. [Google Scholar] [CrossRef]
- Ki, M. 2015 MERS outbreak in Korea: Hospital-to-hospital transmission. Epidemiol. Health 2015, 37, e2015033. [Google Scholar] [CrossRef]
- Kim, K.H.; Tandi, T.E.; Choi, J.W.; Moon, J.M.; Kim, M.S. Middle East respiratory syndrome coronavirus (MERS-CoV) outbreak in South Korea, 2015: Epidemiology, characteristics and public health implications. J. Hosp. Infect. 2017, 95, 207–213. [Google Scholar] [CrossRef]
- Robertson, M.J.; Ritz, J. Biology and clinical relevance of human natural killer cells. Blood 1990, 76, 2421–2438. [Google Scholar] [CrossRef]
- Hanna, N. The role of natural killer cells in the control of tumor growth and metastasis. Biochim. Biophys. Acta 1985, 780, 213–226. [Google Scholar] [CrossRef]
- Hu, W.; Wang, G.; Huang, D.; Sui, M.; Xu, Y. Cancer Immunotherapy Based on Natural Killer Cells: Current Progress and New Opportunities. Front. Immunol. 2019, 10, 1205. [Google Scholar] [CrossRef]
- Allison, M.; Mathews, J.; Gilliland, T.; Mathew, S.O. Natural Killer Cell-Mediated Immunotherapy for Leukemia. Cancers 2022, 14, 843. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Parham, P.; Guethlein, L.A. Genetics of Natural Killer Cells in Human Health, Disease, and Survival. Annu. Rev. Immunol. 2018, 36, 519–548. [Google Scholar] [CrossRef] [PubMed]
- Rajalingam, R. Human diversity of killer cell immunoglobulin-like receptors and disease. Korean J. Hematol. 2011, 46, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Pende, D.; Falco, M.; Vitale, M.; Cantoni, C.; Vitale, C.; Munari, E.; Bertaina, A.; Moretta, F.; Del Zotto, G.; Pietra, G.; et al. Killer Ig-Like Receptors (KIRs): Their Role in NK Cell Modulation and Developments Leading to Their Clinical Exploitation. Front. Immunol. 2019, 10, 1179. [Google Scholar] [CrossRef]
- Trowsdale, J.; Barten, R.; Haude, A.; Stewart, C.A.; Beck, S.; Wilson, M.J. The genomic context of natural killer receptor extended gene families. Immunol. Rev. 2001, 181, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Cooley, S.; Weisdorf, D.J.; Guethlein, L.A.; Klein, J.P.; Wang, T.; Le, C.T.; Marsh, S.G.; Geraghty, D.; Spellman, S.; Haagenson, M.D.; et al. Donor selection for natural killer cell receptor genes leads to superior survival after unrelated transplantation for acute myelogenous leukemia. Blood 2010, 116, 2411–2419. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.; Torkar, M.; Haude, A.; Milne, S.; Jones, T.; Sheer, D.; Beck, S.; Trowsdale, J. Plasticity in the organization and sequences of human KIR/ILT gene families. Proc. Natl. Acad. Sci. USA 2000, 97, 4778–4783. [Google Scholar] [CrossRef]
- Trowsdale, J. Genetic and functional relationships between MHC and NK receptor genes. Immunity 2001, 15, 363–374. [Google Scholar] [CrossRef]
- Dębska-Zielkowska, J.; Moszkowska, G.; Zieliński, M.; Zielińska, H.; Dukat-Mazurek, A.; Trzonkowski, P.; Stefańska, K. KIR Receptors as Key Regulators of NK Cells Activity in Health and Disease. Cells 2021, 10, 1777. [Google Scholar] [CrossRef]
- Maxwell, L.D.; Wallace, A.; Middleton, D.; Curran, M.D. A common KIR2DS4 deletion variant in the human that predicts a soluble KIR molecule analogous to the KIR1D molecule observed in the rhesus monkey. Tissue Antigens 2002, 60, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.; Williams, F.; Halfpenny, I.A. KIR genes. Transpl. Immunol. 2005, 14, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.; Gonzelez, F. The extensive polymorphism of KIR genes. Immunology 2010, 129, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Parham, P. MHC class I molecules and KIRs in human history, health and survival. Nat. Rev. Immunol. 2005, 5, 201–214. [Google Scholar] [CrossRef]
- Varbanova, V.; Naumova, E.; Mihaylova, A. Killer-cell immunoglobulin-like receptor genes and ligands and their role in hematologic malignancies. Cancer Immunol. Immunother. 2016, 65, 427–440. [Google Scholar] [CrossRef]
- D’Andrea, A.; Chang, C.; Franz-Bacon, K.; McClanahan, T.; Phillips, J.H.; Lanier, L.L. Molecular cloning of NKB1. A natural killer cell receptor for HLA-B allotypes. J. Immunol. 1995, 155, 2306–2310. [Google Scholar] [CrossRef]
- Saunders, P.M.; Pymm, P.; Pietra, G.; Hughes, V.A.; Hitchen, C.; O’Connor, G.M.; Loiacono, F.; Widjaja, J.; Price, D.A.; Falco, M.; et al. Killer cell immunoglobulin-like receptor 3DL1 polymorphism defines distinct hierarchies of HLA class I recognition. J. Exp. Med. 2016, 213, 791–807. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Mulrooney, T.J.; Le Luduec, J.B.; Barker, E.; Hsu, K.C. KIR3DL1 and HLA-B Density and Binding Calibrate NK Education and Response to HIV. J. Immunol. 2016, 196, 3398–3410. [Google Scholar] [CrossRef]
- Stern, M.; Ruggeri, L.; Capanni, M.; Mancusi, A.; Velardi, A. Human leukocyte antigens A23, A24, and A32 but not A25 are ligands for KIR3DL1. Blood 2008, 112, 708–710. [Google Scholar] [CrossRef]
- Saunders, P.M.; MacLachlan, B.J.; Widjaja, J.; Wong, S.C.; Oates, C.V.L.; Rossjohn, J.; Vivian, J.P.; Brooks, A.G. The Role of the HLA Class I α2 Helix in Determining Ligand Hierarchy for the Killer Cell Ig-like Receptor 3DL1. J. Immunol. 2021, 206, 849–860. [Google Scholar] [CrossRef]
- Hansasuta, P.; Dong, T.; Thananchai, H.; Weekes, M.; Willberg, C.; Aldemir, H.; Rowland-Jones, S.; Braud, V.M. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 2004, 34, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.A.; Laugier-Anfossi, F.; Vély, F.; Saulquin, X.; Riedmuller, J.; Tisserant, A.; Gauthier, L.; Romagné, F.; Ferracci, G.; Arosa, F.A.; et al. Recognition of peptide-MHC class I complexes by activating killer immunoglobulin-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 13224–13229. [Google Scholar] [CrossRef]
- Chewning, J.H.; Gudme, C.N.; Hsu, K.C.; Selvakumar, A.; Dupont, B. KIR2DS1-positive NK cells mediate alloresponse against the C2 HLA-KIR ligand group in vitro. J. Immunol. 2007, 179, 854–868. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, H.G.; Kärre, K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, J.; Li, S.; Sun, J.; Teng, Y.; Wu, M.; Li, J.; Li, Y.; Hu, N.; Wang, H.; et al. Epitope-Based Vaccine Target Screening against Highly Pathogenic MERS-CoV: An In Silico Approach Applied to Emerging Infectious Diseases. PLoS ONE 2015, 10, e0144475. [Google Scholar] [CrossRef] [PubMed]
- Badawi, M.M.; SalahEldin, M.A.; Suliman, M.M.; AbduRahim, S.A.; Mohammed, A.A.e.; SidAhmed, A.S.A.; Osman, M.M.; Salih, M.A. In Silico Prediction of a Novel Universal Multi-epitope Peptide Vaccine in the Whole Spike Glycoprotein of MERS CoV. Am. J. Microbiol. Res. 2016, 4, 101–121. [Google Scholar]
- Liu, W.J.; Zhao, M.; Liu, K.; Xu, K.; Wong, G.; Tan, W.; Gao, G.F. T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV. Antivir. Res. 2017, 137, 82–92. [Google Scholar] [CrossRef]
- Cong, Y.; Hart, B.J.; Gross, R.; Zhou, H.; Frieman, M.; Bollinger, L.; Wada, J.; Hensley, L.E.; Jahrling, P.B.; Dyall, J.; et al. MERS-CoV pathogenesis and antiviral efficacy of licensed drugs in human monocyte-derived antigen-presenting cells. PLoS ONE 2018, 13, e0194868. [Google Scholar] [CrossRef]
- Srivastava, S.; Kamthania, M.; Singh, S.; Saxena, A.K.; Sharma, N. Structural basis of development of multi-epitope vaccine against Middle East respiratory syndrome using in silico approach. Infect. Drug Resist. 2018, 11, 2377–2391. [Google Scholar] [CrossRef]
- Shokri, S.; Mahmoudvand, S.; Taherkhani, R.; Farshadpour, F. Modulation of the immune response by Middle East respiratory syndrome coronavirus. J. Cell. Physiol. 2019, 234, 2143–2151. [Google Scholar] [CrossRef]
- Tahir Ul Qamar, M.; Saleem, S.; Ashfaq, U.A.; Bari, A.; Anwar, F.; Alqahtani, S. Epitope-based peptide vaccine design and target site depiction against Middle East Respiratory Syndrome Coronavirus: An immune-informatics study. J. Transl. Med. 2019, 17, 362. [Google Scholar] [CrossRef] [PubMed]
- Nallusamy, S.; Mannu, J.; Ravikumar, C.; Angamuthu, K.; Nathan, B.; Nachimuthu, K.; Ramasamy, G.; Muthurajan, R.; Subbarayalu, M.; Neelakandan, K. Exploring Phytochemicals of Traditional Medicinal Plants Exhibiting Inhibitory Activity Against Main Protease, Spike Glycoprotein, RNA-dependent RNA Polymerase and Non-Structural Proteins of SARS-CoV-2 Through Virtual Screening. Front. Pharmacol. 2021, 12, 667704. [Google Scholar] [CrossRef] [PubMed]
- Mahallawi, W.H.; Khabour, O.F.; Zhang, Q.; Makhdoum, H.M.; Suliman, B.A. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine 2018, 104, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Market, M.; Angka, L.; Martel, A.B.; Bastin, D.; Olanubi, O.; Tennakoon, G.; Boucher, D.M.; Ng, J.; Ardolino, M.; Auer, R.C. Flattening the COVID-19 Curve With Natural Killer Cell Based Immunotherapies. Front. Immunol. 2020, 11, 1512. [Google Scholar] [CrossRef] [PubMed]
- Soleimanian, S.; Yaghobi, R. Harnessing Memory NK Cell to Protect Against COVID-19. Front. Pharmacol. 2020, 11, 1309. [Google Scholar] [CrossRef] [PubMed]
- Baek, I.C.; Choi, E.J.; Shin, D.H.; Kim, H.J.; Choi, H.; Shin, H.S.; Lim, D.G.; Kim, T.G. Association of HLA class I and II genes with Middle East respiratory syndrome coronavirus infection in Koreans. Immun. Inflamm. Dis. 2022, 10, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Baek, I.C.; Park, S.; Kim, H.J.; Kim, T.G. Development of cost-effective and fast KIR genotyping by multiplex PCR-SSP. Hla 2023, 103, e15191. [Google Scholar] [CrossRef]
- Kim, H.J.; Choi, H.B.; Jang, J.P.; Baek, I.C.; Choi, E.J.; Park, M.; Kim, T.G.; Oh, S.T. HLA-Cw polypmorphism and killer cell immunoglobulin-like receptor (KIR) gene analysis in Korean colorectal cancer patients. Int. J. Surg. 2014, 12, 815–820. [Google Scholar] [CrossRef]
- Jang, J.P.; Cho, W.K.; Baek, I.C.; Choi, E.J.; Shin, D.H.; Suh, B.K.; Kim, T.G. Comprehensive analysis of cytokine gene polymorphisms defines the association of IL-12 gene with ophthalmopthy in Korean children with autoimmune thyroid disease. Mol. Cell. Endocrinol. 2016, 426, 43–49. [Google Scholar] [CrossRef]
- Cho, W.K.; Jang, J.P.; Choi, E.J.; Ahn, M.; Kim, S.H.; Cho, K.S.; Park, S.H.; Baek, I.C.; Jung, M.H.; Kim, T.G.; et al. Association of Polymorphisms in Toll-Like Receptors 4 and 9 with Autoimmune Thyroid Disease in Korean Pediatric Patients. Int. J. Endocrinol. 2017, 2017, 2304218. [Google Scholar] [CrossRef]
- Baek, I.C.; Shin, D.H.; Choi, E.J.; Kim, H.J.; Yoon, J.H.; Cho, B.S.; Kim, Y.J.; Lee, S.; Min, W.S.; Kim, H.J.; et al. Association of MICA and MICB polymorphisms with the susceptibility of leukemia in Korean patients. Blood Cancer J. 2018, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Baek, I.C.; Kim, H.J.; Choi, E.J.; Ahn, M.; Jung, M.H.; Suh, B.K.; Cho, W.K.; Kim, T.G. HLA alleles, especially amino-acid signatures of HLA-DPB1, might contribute to the molecular pathogenesis of early-onset autoimmune thyroid disease. PLoS ONE 2019, 14, e0216941. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Shin, H.R.; Lee, N.Y.; Kim, S.K.; Ahn, M.B.; Baek, I.C.; Kim, T.G.; Suh, B.K. GPR174 and ITM2A Gene Polymorphisms rs3827440 and rs5912838 on the X chromosome in Korean Children with Autoimmune Thyroid Disease. Genes 2020, 11, 858. [Google Scholar] [CrossRef] [PubMed]
- Min, G.J.; Kim, H.J.; Kim, T.G.; Hyun, Y.S.; Hyun, S.J.; Baek, I.C.; Yoon, S.Y.; Park, S.S.; Park, S.; Yoon, J.H.; et al. Specific donor HLA allotypes as predictors of cytomegalovirus disease risk in acute myeloid leukemia. Hla 2020, 96, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.R.; Cho, W.K.; Baek, I.C.; Lee, N.Y.; Lee, Y.J.; Kim, S.K.; Ahn, M.B.; Suh, B.K.; Kim, T.G. Polymorphisms of IRAK1 Gene on X Chromosome Is Associated with Hashimoto Thyroiditis in Korean Children. Endocrinology 2020, 161, bqaa088. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Baek, I.C.; Kim, S.E.; Kim, M.; Kim, T.G.; Suh, B.K. Association of ITM2A rs1751094 polymorphism on X chromosome in Korean pediatric patients with autoimmune thyroid disease. Immun. Inflamm. Dis. 2023, 11, e800. [Google Scholar] [CrossRef]
- Choi, S.S.; Choi, H.; Baek, I.C.; Park, S.A.; Park, J.S.; Kim, T.G.; Jeun, S.S.; Ahn, S. HLA polymorphisms and risk of glioblastoma in Koreans. PLoS ONE 2021, 16, e0260618. [Google Scholar] [CrossRef]
- Choi, H.; Baek, I.C.; Park, S.A.; Park, J.S.; Jeun, S.S.; Kim, T.G.; Ahn, S. Polymorphisms of Killer Ig-like Receptors and the Risk of Glioblastoma. J. Clin. Med. 2023, 12, 4780. [Google Scholar] [CrossRef]
- Cui, W.; Fan, Y.; Wu, W.; Zhang, F.; Wang, J.Y.; Ni, A.P. Expression of lymphocytes and lymphocyte subsets in patients with severe acute respiratory syndrome. Clin. Infect. Dis. 2003, 37, 857–859. [Google Scholar] [CrossRef]
- Zawawi, A.; Naser, A.Y.; Alwafi, H.; Minshawi, F. Profile of Circulatory Cytokines and Chemokines in Human Coronaviruses: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 666223. [Google Scholar] [CrossRef]
- Petrosillo, N.; Viceconte, G.; Ergonul, O.; Ippolito, G.; Petersen, E. COVID-19, SARS and MERS: Are they closely related? Clin. Microbiol. Infect. 2020, 26, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zheng, X.; Tong, Q.; Li, W.; Wang, B.; Sutter, K.; Trilling, M.; Lu, M.; Dittmer, U.; Yang, D. Overlapping and discrete aspects of the pathology and pathogenesis of the emerging human pathogenic coronaviruses SARS-CoV, MERS-CoV, and 2019-nCoV. J. Med. Virol. 2020, 92, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Wang, Q.; Zhang, D.; Ding, J.; Huang, Q.; Tang, Y.Q.; Wang, Q.; Miao, H. Lymphopenia predicts disease severity of COVID-19: A descriptive and predictive study. Signal Transduct. Target. Ther. 2020, 5, 33. [Google Scholar] [CrossRef]
- He, R.; Lu, Z.; Zhang, L.; Fan, T.; Xiong, R.; Shen, X.; Feng, H.; Meng, H.; Lin, W.; Jiang, W.; et al. The clinical course and its correlated immune status in COVID-19 pneumonia. J. Clin. Virol. 2020, 127, 104361. [Google Scholar] [CrossRef] [PubMed]
- Maruthamuthu, S.; Rajalingam, K.; Kaur, N.; Morvan, M.G.; Soto, J.; Lee, N.; Kong, D.; Hu, Z.; Reyes, K.; Ng, D.; et al. Individualized Constellation of Killer Cell Immunoglobulin-Like Receptors and Cognate HLA Class I Ligands that Controls Natural Killer Cell Antiviral Immunity Predisposes COVID-19. Front. Genet. 2022, 13, 845474. [Google Scholar] [CrossRef] [PubMed]
- Saresella, M.; Trabattoni, D.; Marventano, I.; Piancone, F.; La Rosa, F.; Caronni, A.; Lax, A.; Bianchi, L.; Banfi, P.; Navarro, J.; et al. NK Cell Subpopulations and Receptor Expression in Recovering SARS-CoV-2 Infection. Mol. Neurobiol. 2021, 58, 6111–6120. [Google Scholar] [CrossRef]
- Hu, S.; Shao, Z.; Ni, W.; Sun, P.; Qiao, J.; Wan, H.; Huang, Y.; Liu, X.; Zhai, H.; Xiao, M.; et al. The KIR2DL2/HLA-C1C1 Gene Pairing Is Associated With an Increased Risk of SARS-CoV-2 Infection. Front. Immunol. 2022, 13, 919110. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Y.; Kuang, B.H.; Yan, X.; Lei, J.; Lin, Y.X.; Tian, J.; Li, Y.; Xie, X.; Chen, T.; et al. HLA-Bw4 in association with KIR3DL1 favors natural killer cell-mediated protection against severe COVID-19. Emerg. Microbes Infect. 2023, 12, 2185467. [Google Scholar] [CrossRef]
- Ligotti, M.E.; Aiello, A.; Accardi, G.; Calabrò, A.; Ciaccio, M.; Colomba, C.; Di Bona, D.; Lo Sasso, B.; Pojero, F.; Tuttolomondo, A.; et al. Distribution of KIR Genes and Their HLA Ligands in Different Viral Infectious Diseases: Frequency Study in Sicilian Population. Int. J. Mol. Sci. 2022, 23, 15466. [Google Scholar] [CrossRef]
- Raulet, D.H. Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev. Immunol. 2003, 3, 781–790. [Google Scholar] [CrossRef]
- Beetz, S.; Marischen, L.; Kabelitz, D.; Wesch, D. Human gamma delta T cells: Candidates for the development of immunotherapeutic strategies. Immunol. Res. 2007, 37, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Busche, A.; Goldmann, T.; Naumann, U.; Steinle, A.; Brandau, S. Natural killer cell-mediated rejection of experimental human lung cancer by genetic overexpression of major histocompatibility complex class I chain-related gene A. Hum. Gene Ther. 2006, 17, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Germain, C.; Larbouret, C.; Cesson, V.; Donda, A.; Held, W.; Mach, J.P.; Pelegrin, A.; Robert, B. MHC class I-related chain A conjugated to antitumor antibodies can sensitize tumor cells to specific lysis by natural killer cells. Clin. Cancer Res. 2005, 11, 7516–7522. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.S.; Lee, Y.J.; Wu, C.W.; Liu, C.J.; Huang, J.W.; Lui, W.Y. The increase of MICA gene A9 allele associated with gastric cancer and less schirrous change. Br. J. Cancer 2004, 90, 1809–1813. [Google Scholar] [CrossRef]
- Salih, H.R.; Rammensee, H.G.; Steinle, A. Cutting edge: Down-regulation of MICA on human tumors by proteolytic shedding. J. Immunol. 2002, 169, 4098–4102. [Google Scholar] [CrossRef]
- Luo, Q.Z.; Lin, L.; Gong, Z.; Mei, B.; Xu, Y.J.; Huo, Z.; Yu, P. Positive association of major histocompatibility complex class I chain-related gene A polymorphism with leukemia susceptibility in the people of Han nationality of Southern China. Tissue Antigens 2011, 78, 178–184. [Google Scholar] [CrossRef]
- Gonzalez-Gay, M.A.; Rueda, B.; Vilchez, J.R.; Lopez-Nevot, M.A.; Robledo, G.; Ruiz, M.P.; Fernandez, O.; Garcia-Porrua, C.; Gonzalez-Escribano, M.F.; Martin, J. Contribution of MHC class I region to genetic susceptibility for giant cell arteritis. Rheumatology 2007, 46, 431–434. [Google Scholar] [CrossRef]
- Park, Y.; Lee, H.; Sanjeevi, C.B.; Eisenbarth, G.S. MICA polymorphism is associated with type 1 diabetes in the Korean population. Diabetes Care 2001, 24, 33–38. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Group (Clinical Severity) | Total | ||
|---|---|---|---|
| Mo/Mi | Severe | ||
| No. of patients (%) | 16 (50) | 16 (50) | 32 |
| Male/female (n) | 8/8 | 13/3 | 21/11 |
| Age (year), mean ± SD | 50 ± 21 | 58 ± 15 | 54 ± 19 |
| Underlying disease, n (%) | |||
| Diabetes | 1 (6.3) | 3 (18.8) | 4 (12.5) |
| Chronic renal disease | 1 (6.3) | 2 (12.5) | 3 (9.4) |
| Chronic cardiac disease | 1 (6.3) | 4 (25.0) | 5 (15.6) |
| Chronic pulmonary disease | 0 (0) | 1 (6.3) | 1 (3.1) |
| Hypertension | 2 (12.5) | 7 (43.8) | 9 (28.1) |
| Chronic liver disease | 1 (6.3) | 0 (0) | 1 (3.1) |
| Solid tumor | 2 (12.5) | 2 (12.5) | 4 (12.5) |
| Obesity | 0 (0) | 1 (6.3) | 1 (3.1) |
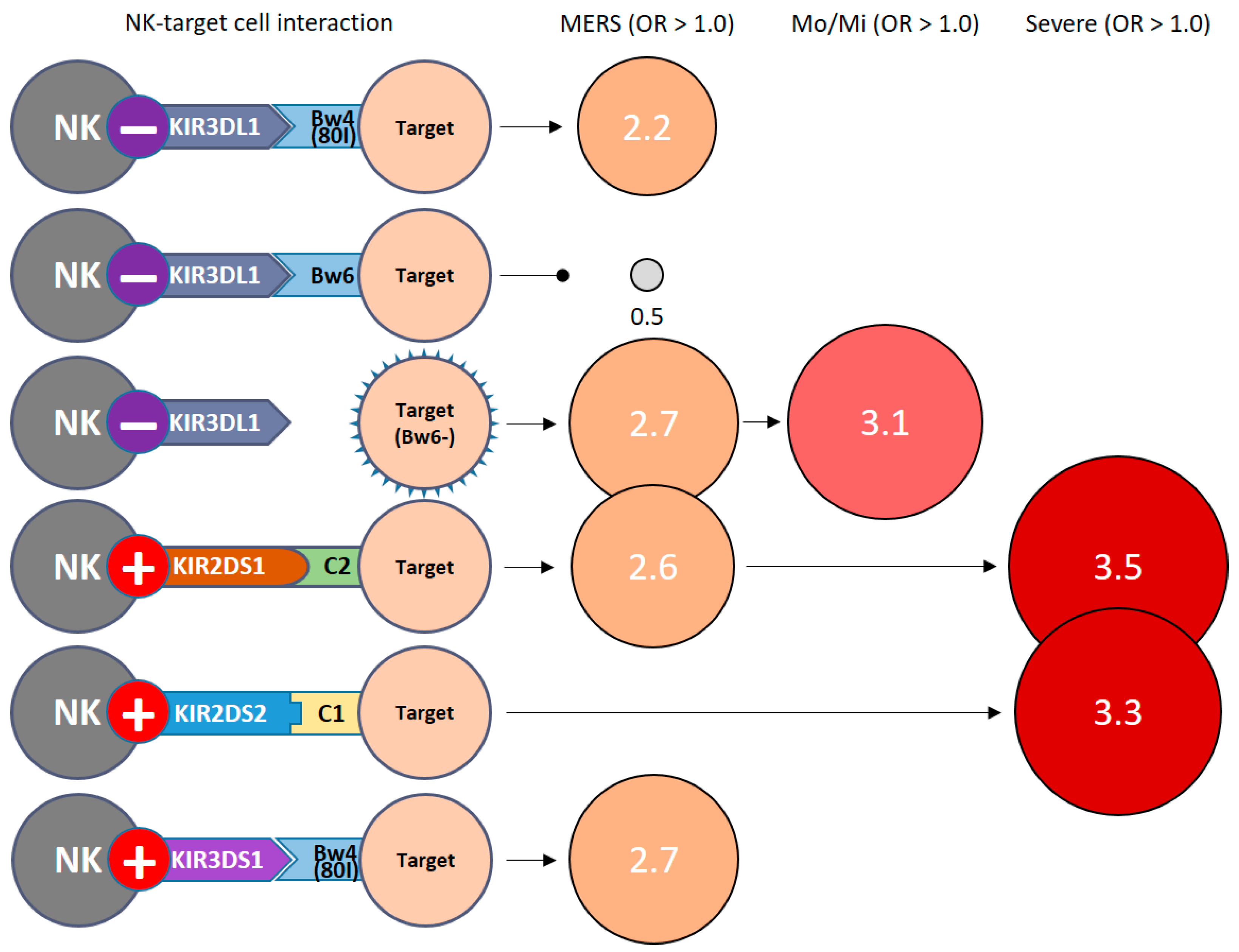
| Function | Gene Clusters | Controls | MERS Total | Mo/Mi Cases | Severe Cases | ||||
|---|---|---|---|---|---|---|---|---|---|
| n = 200 (%) | n = 32 (%) | n = 16 (%) | n = 16 (%) | ||||||
| Inhibitory | KIR2DL1 | 200 | (100.0) | 31 | (96.9) | 15 | (93.8) c | 16 | (100.0) |
| KIR2DL2 | 25 | (12.5) | 8 | (25.0) | 3 | (18.8) | 5 | (31.3) f | |
| KIR2DL3 | 199 | (99.5) | 31 | (96.9) | 15 | (93.8) | 16 | (100.0) | |
| KIR2DL4 | 200 | (100.0) | 32 | (100.0) | 16 | (100.0) | 16 | (100.0) | |
| KIR2DL5 | 74 | (37.0) | 17 | (53.1) | 9 | (56.3) | 8 | (50.0) | |
| KIR2DL5A | 69 | (34.5) | 16 | (50.0) | 9 | (56.3) | 7 | (43.8) | |
| KIR2DL5B | 14 | (7.0) | 2 | (6.3) | 1 | (6.3) | 1 | (6.3) | |
| KIR3DL1 | 183 | (91.5) | 30 | (93.8) | 15 | (93.8) | 15 | (93.8) | |
| KIR3DL2 | 200 | (100.0) | 32 | (100.0) | 16 | (100.0) | 16 | (100.0) | |
| KIR3DL3 | 200 | (100.0) | 32 | (100.0) | 16 | (100.0) | 16 | (100.0) | |
| Activating | KIR2DS1 | 72 | (36.0) | 16 | (50.0) | 9 | (56.3) | 7 | (43.8) |
| KIR2DS2 | 31 | (15.5) | 9 | (28.1) | 3 | (18.8) | 6 | (37.5) g | |
| KIR2DS3 | 37 | (18.5) | 9 | (28.1) | 4 | (25.0) | 5 | (31.3) | |
| KIR2DS4 | 183 | (91.5) | 31 | (96.9) | 15 | (93.8) | 16 | (100.0) | |
| KIR2DS4D | 88 | (44.0) | 21 | (65.6) a | 11 | (68.8) | 10 | (62.5) | |
| KIR2DS4F | 146 | (73.0) | 23 | (71.9) | 10 | (62.5) | 13 | (81.3) | |
| KIR2DS5 | 48 | (24.0) | 9 | (28.1) | 6 | (37.5) | 3 | (18.8) | |
| KIR3DS1 | 73 | (36.5) | 16 | (50.0) | 9 | (56.3) | 7 | (43.8) | |
| Pseudogene | KIR2DP1 | 200 | (100.0) | 31 | (96.9) | 15 | (93.8) d | 16 | (100.0) |
| KIR3DP1 | 200 | (100.0) | 32 | (100.0) | 16 | (100.0) | 16 | (100.0) | |
| KIR3DP1D | 200 | (100.0) | 31 | (96.9) | 15 | (93.8) e | 16 | (100.0) | |
| KIR3DP1F | 19 | (9.5) | 7 | (21.9) b | 2 | (12.5) | 5 | (31.3) h | |
| KIR Genes | Ligand Genes (HLA) | Controls | MERS Total | Mo/Mi Cases | Severe Cases | ||||
|---|---|---|---|---|---|---|---|---|---|
| n = 200 (%) | n = 32 (%) | n = 16 (%) | n = 16 (%) | ||||||
| KIR2DL1+ | C2+ | 61 | (30.5) | 12 | (37.5) | 6 | (37.5) | 6 | (37.5) |
| C2− | 139 | (69.5) | 19 | (59.4) | 9 | (56.3) | 10 | (62.5) | |
| KIR2DL2+ | C1+ | 25 | (12.5) | 8 | (25.0) | 3 | (18.8) | 5 | (31.3) |
| C1− | 0 | (0.0) | 0 | (0.0) | 0 | (0.0) | 0 | (0.0) | |
| KIR2DL3+ | C1+ | 192 | (96.0) | 31 | (96.9) | 15 | (93.8) | 16 | (100.0) |
| C1− | 7 | (3.5) | 0 | (0.0) | 0 | (0.0) | 0 | (0.0) | |
| KIR3DL1+ | Bw4+ | 133 | (66.5) | 25 | (78.1) | 11 | (68.8) | 14 | (87.5) |
| BW4− | 50 | (25.0) | 5 | (15.6) | 4 | (25.0) | 1 | (6.3) | |
| Bw4 (80I)+ | 68 | (34.0) | 17 | (53.1) a | 8 | (50.0) | 9 | (56.3) | |
| Bw4 (80I)− | 115 | (57.5) | 13 | (40.6) | 7 | (43.8) | 6 | (37.5) | |
| Bw4 (80T)+ | 33 | (16.5) | 5 | (15.6) | 1 | (6.3) | 4 | (25.0) | |
| Bw4 (80T)− | 150 | (75.0) | 25 | (78.1) | 14 | (87.5) | 11 | (68.8) | |
| Bw6+ | 143 | (71.5) | 17 | (53.1) b | 8 | (50.0) | 9 | (56.3) | |
| Bw6− | 40 | (20.0) | 13 | (40.6) c | 7 | (43.8) f | 6 | (37.5) | |
| KIR3DL2+ | A3+ or A11+ | 49 | (24.5) | 6 | (18.8) | 3 | (18.8) | 3 | (18.8) |
| A3− and A11− | 151 | (75.5) | 26 | (81.3) | 13 | (81.3) | 13 | (81.3) | |
| KIR2DS1+ | C2+ | 23 | (11.5) | 8 | (25.0) d | 3 | (18.8) | 5 | (31.3) g |
| C2− | 49 | (24.5) | 8 | (25.0) | 6 | (37.5) | 2 | (12.5) | |
| KIR2DS2+ | C1+ | 31 | (15.5) | 9 | (28.1) | 3 | (18.8) | 6 | (37.5) h |
| C1− | 0 | (0.0) | 0 | (0.0) | 0 | (0.0) | 0 | (0.0) | |
| KIR3DS1+ | Bw4+ | 56 | (28.0) | 13 | (40.6) | 7 | (43.8) | 6 | (37.5) |
| Bw4− | 17 | (8.5) | 3 | (9.4) | 2 | (12.5) | 1 | (6.3) | |
| Bw4 (80I)+ | 29 | (14.5) | 10 | (31.3) e | 5 | (31.3) | 5 | (31.3) | |
| Bw4 (80I)− | 44 | (22.0) | 6 | (18.8) | 4 | (25.0) | 2 | (12.5) | |
| Bw4 (80T)+ | 13 | (6.5) | 4 | (12.5) | 1 | (6.3) | 3 | (18.8) | |
| Bw4 (80T)− | 60 | (30.0) | 12 | (37.5) | 8 | (50.0) | 4 | (25.0) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, I.-C.; Choi, E.-J.; Kim, H.-J.; Choi, H.; Shin, H.-S.; Lim, D.-G.; Kim, T.-G. Association of KIR Genes with Middle East Respiratory Syndrome Coronavirus Infection in South Koreans. J. Clin. Med. 2024, 13, 258. https://doi.org/10.3390/jcm13010258
Baek I-C, Choi E-J, Kim H-J, Choi H, Shin H-S, Lim D-G, Kim T-G. Association of KIR Genes with Middle East Respiratory Syndrome Coronavirus Infection in South Koreans. Journal of Clinical Medicine. 2024; 13(1):258. https://doi.org/10.3390/jcm13010258
Chicago/Turabian StyleBaek, In-Cheol, Eun-Jeong Choi, Hyoung-Jae Kim, Haeyoun Choi, Hyoung-Shik Shin, Dong-Gyun Lim, and Tai-Gyu Kim. 2024. "Association of KIR Genes with Middle East Respiratory Syndrome Coronavirus Infection in South Koreans" Journal of Clinical Medicine 13, no. 1: 258. https://doi.org/10.3390/jcm13010258