
Causal Roles of Ventral and Dorsal Neural Systems for Automatic and Control Self-Reference Processing: A Function Lesion Mapping Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.1.1. Patients
2.1.2. Healthy Controls
2.2. Neuropsychological Assessments
2.3. Principal Component Analysis (PCA)
2.4. Neuroimaging Assessment
2.4.1. Image Acquirement
2.4.2. Image Pre-Processing
2.4.3. Lesion Reconstruction
2.4.4. Voxel-Based Morphometry (VBM)
2.4.5. Track-Wise Lesion-Deficit Analysis
3. Results
3.1. Neuropsychological Profiles and the Self and Attentional Control Factors
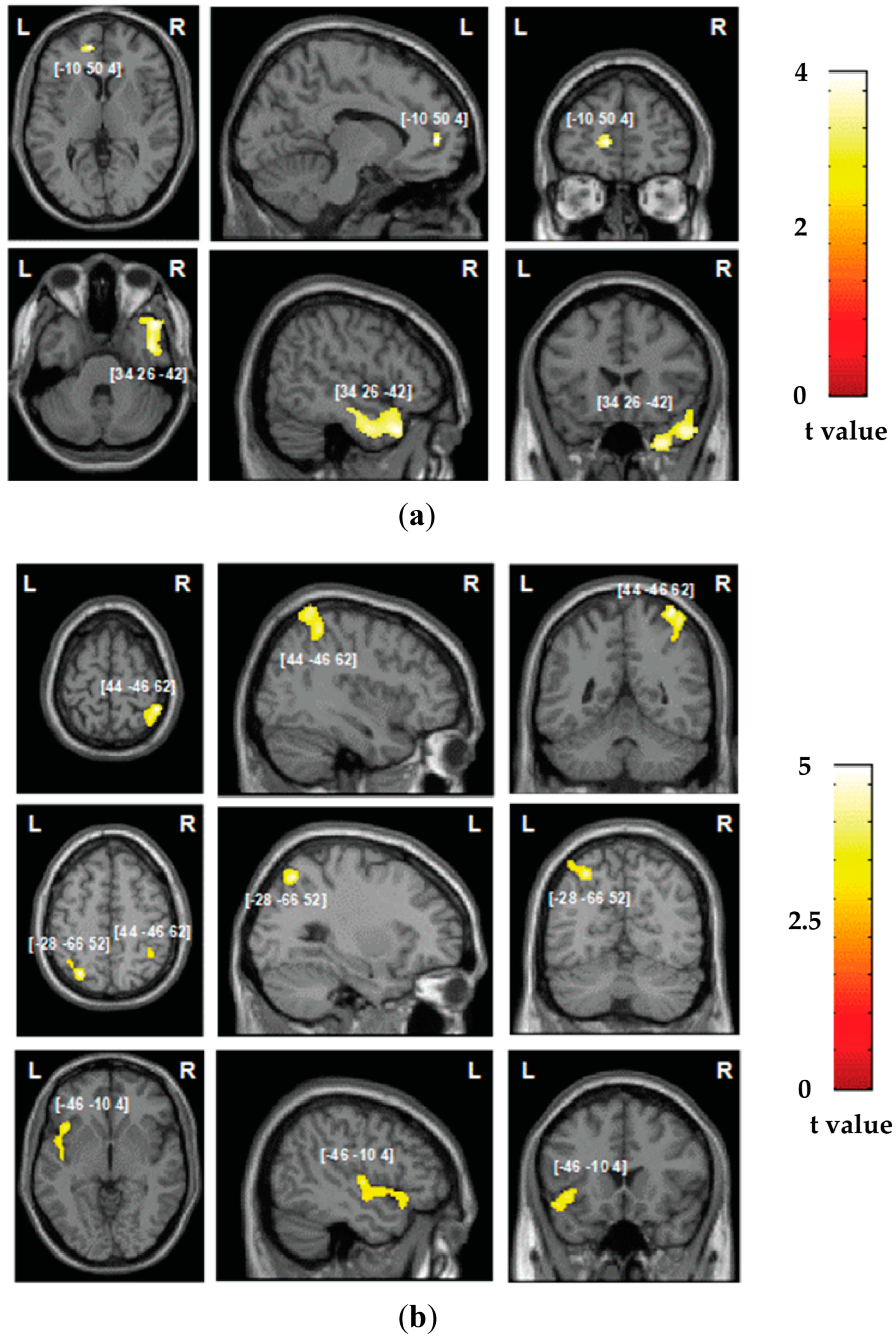
3.2. Neuroimaging Findings: Grey Matter Damage
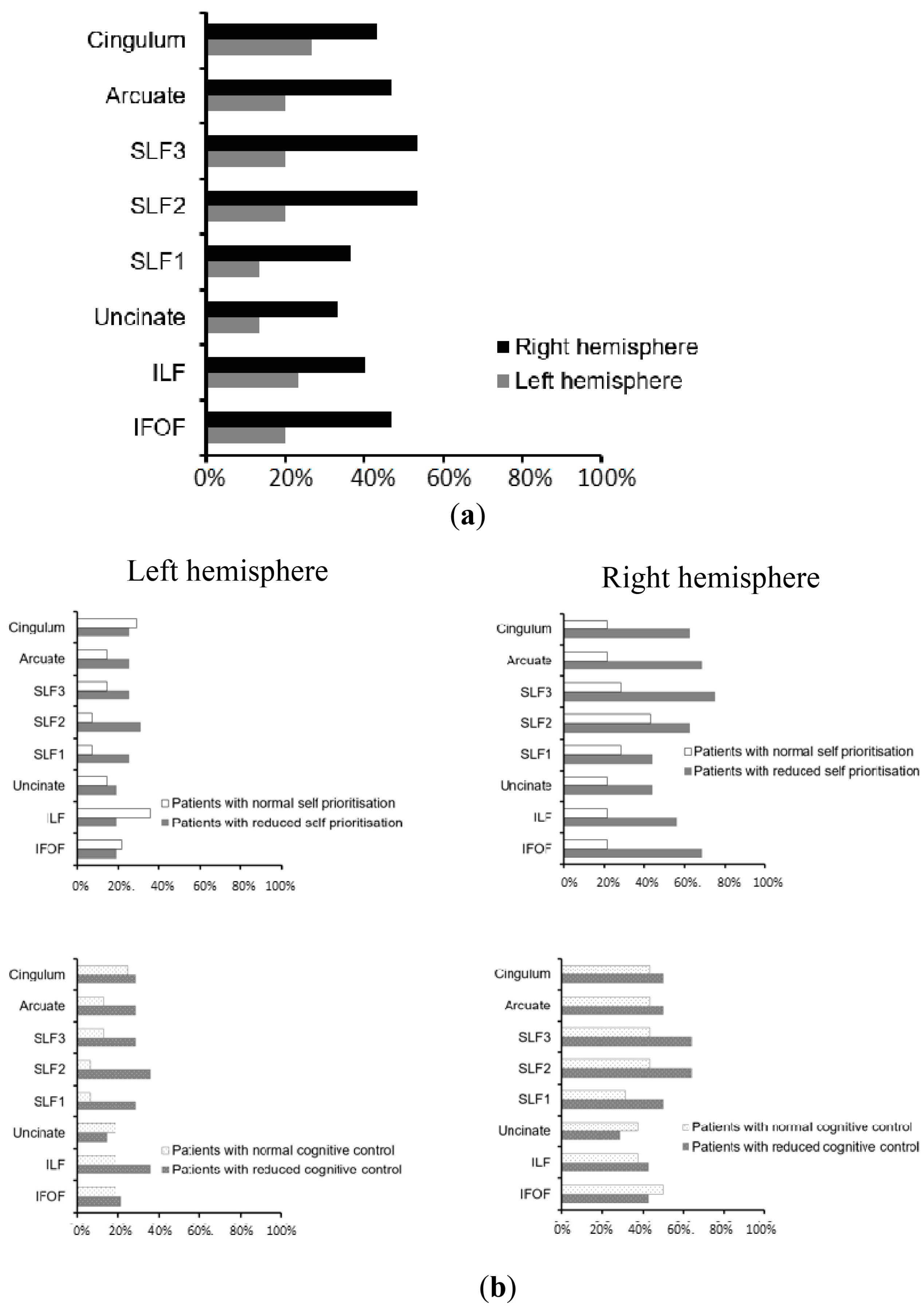
3.3. Neuroimaging Findings: White Matter Disconnections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGuire, W.J.; McGuire, C.V.; Cheever, J. The Self in Society: Effects of Social Contexts on the Sense of Self. Br. J. Soc. Psychol. 1986, 25, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Churchland, P.S. Self-Representation in Nervous Systems. Science 2002, 296, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, S. Philosophical Conceptions of the Self: Implications for Cognitive Science. Trends Cogn. Sci. 2000, 4, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, L. An Experimental Study on the Self-Reference Effect. Sci. China C Life Sci. 2002, 45, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Lichtenberg, J.D. The Development of the Sense of Self. J. Am. Psychoanal. Assoc. 1975, 23, 453–484. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.A.; Pleydell-Pearce, C.W. The Construction of Autobiographical Memories in the Self-Memory System. Psychol. Rev. 2000, 107, 261. [Google Scholar] [CrossRef]
- Sui, J.; Humphreys, G.W. The Ubiquitous Self: What the Properties of Self-bias Tell Us about the Self. Ann. N. Y. Acad. Sci. 2017, 1396, 222–235. [Google Scholar] [CrossRef]
- Desebrock, C.; Spence, C. The Self-Prioritization Effect: Self-Referential Processing in Movement Highlights Modulation at Multiple Stages. Atten. Percept. Psychophys. 2021, 83, 2656–2674. [Google Scholar] [CrossRef]
- Singh, D.; Karnick, H. Self-Prioritization Effect in Children and Adults. Front. Psychol. 2022, 13, 726230. [Google Scholar] [CrossRef]
- Palmero, L.B.; Martínez-Pérez, V.; Tortajada, M.; Campoy, G.; Fuentes, L.J. Testing the Modulation of Self-Related Automatic and Others-Related Controlled Processing by Chronotype and Time-of-Day. Conscious. Cogn. 2024, 118, 103633. [Google Scholar] [CrossRef]
- Apps, M.A.J.; Tsakiris, M. The Free-Energy Self: A Predictive Coding Account of Self-Recognition. Neurosci. Biobehav. Rev. 2014, 41, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, S.J.; Turk, D.J.; Macdonald, L.M.; Macrae, C.N. Yours or Mine? Ownership and Memory. Conscious. Cogn. 2008, 17, 312–318. [Google Scholar] [CrossRef]
- Golubickis, M.; Macrae, C.N. That’s Me in the Spotlight: Self-Relevance Modulates Attentional Breadth. Psychon. Bull. Rev. 2021, 28, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Knoblich, G. Self-Prioritization of Fully Unfamiliar Stimuli. Q. J. Exp. Psychol. 2019, 72, 2110–2120. [Google Scholar] [CrossRef]
- Woźniak, M.; Knoblich, G. Self-Prioritization Depends on Assumed Task-Relevance of Self-Association. Psychol. Res. 2022, 86, 1599–1614. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; He, X.; Humphreys, G.W. Perceptual Effects of Social Salience: Evidence from Self-Prioritization Effects on Perceptual Matching. J. Exp. Psychol. Hum. Percept. Perform. 2012, 38, 1105. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Humphreys, G.W. Aging Enhances Cognitive Biases to Friends but Not the Self. Psychon. Bull. Rev. 2017, 24, 2021–2030. [Google Scholar] [CrossRef]
- Sun, Y.; Fuentes, L.J.; Humphreys, G.W.; Sui, J. Try to See It My Way: Embodied Perspective Enhances Self and Friend-Biases in Perceptual Matching. Cognition 2016, 153, 108–117. [Google Scholar] [CrossRef]
- Sui, J.; Cao, B.; Song, Y.; Greenshaw, A.J. Individual Differences in Self-and Value-Based Reward Processing. Curr. Res. Behav. Sci. 2023, 4, 100095. [Google Scholar] [CrossRef]
- Żochowska, A.; Jakuszyk, P.; Nowicka, M.M.; Nowicka, A. The Self and a Close-Other: Differences between Processing of Faces and Newly Acquired Information. Cereb. Cortex 2023, 33, 2183–2199. [Google Scholar] [CrossRef]
- Schäfer, S.; Frings, C. Understanding Self-Prioritisation: The Prioritisation of Self-Relevant Stimuli and Its Relation to the Individual Self-Esteem. J. Cogn. Psychol. 2019, 31, 813–824. [Google Scholar] [CrossRef]
- Golubickis, M.; Persson, L.M.; Falbén, J.K.; Macrae, C.N. On Stopping Yourself: Self-Relevance Facilitates Response Inhibition. Atten. Percept. Psychophys. 2021, 83, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pérez, V.; Sandoval-Lentisco, A.; Tortajada, M.; Palmero, L.B.; Campoy, G.; Fuentes, L.J. Self-Prioritization Effect in the Attentional Blink Paradigm: Attention-Based or Familiarity-Based Effect? Conscious. Cogn. 2024, 117, 103607. [Google Scholar] [CrossRef] [PubMed]
- Conty, L.; George, N.; Hietanen, J.K. Watching Eyes Effects: When Others Meet the Self. Conscious. Cogn. 2016, 45, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Renninger, K.A.; Hidi, S.E. Interest Development, Self-Related Information Processing, and Practice. Theory Pract. 2022, 61, 23–34. [Google Scholar] [CrossRef]
- Tacikowski, P.; Weijs, M.L.; Ehrsson, H.H. Perception of Our Own Body Influences Self-Concept and Self-Incoherence Impairs Episodic Memory. Iscience 2020, 23, 101429. [Google Scholar] [CrossRef]
- Keil, J.; Barutchu, A.; Desebrock, C.; Spence, C. More of Me: Self-Prioritization of Numeric Stimuli. J. Exp. Psychol. Hum. Percept. Perform. 2023, 49, 1518–1533. [Google Scholar] [CrossRef]
- Roy, N.; Karnick, H.; Verma, A. Towards the Self and Away from the Others: Evidence for Self-Prioritization Observed in an Approach Avoidance Task. Front. Psychol. 2023, 14, 1041157. [Google Scholar] [CrossRef]
- Bogdanova, O.V.; Bogdanov, V.B.; Dureux, A.; Farne, A.; Hadj-Bouziane, F. The Peripersonal Space in a Social World. Cortex 2021, 142, 28–46. [Google Scholar] [CrossRef]
- Blanke, O.; Slater, M.; Serino, A. Behavioral, Neural, and Computational Principles of Bodily Self-Consciousness. Neuron 2015, 88, 145–166. [Google Scholar] [CrossRef]
- Serino, A.; Alsmith, A.; Costantini, M.; Mandrigin, A.; Tajadura-Jimenez, A.; Lopez, C. Bodily Ownership and Self-Location: Components of Bodily Self-Consciousness. Conscious. Cogn. 2013, 22, 1239–1252. [Google Scholar] [CrossRef] [PubMed]
- Symons, C.S.; Johnson, B.T. The Self-Reference Effect in Memory: A Meta-Analysis. Psychol. Bull. 1997, 121, 371. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; He, X.; Rotsthein, P.; Sui, J. Dynamically Orienting Your Own Face Facilitates the Automatic Attraction of Attention. Cogn. Neurosci. 2016, 7, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Sun, Y.; Peng, K.; Humphreys, G.W. The Automatic and the Expected Self: Separating Self- and Familiarity Biases Effects by Manipulating Stimulus Probability. Atten. Percept. Psychophys. 2014, 76, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G. How Does the ‘Rest-Self Overlap’ Mediate the Qualitative and Automatic Features of Self-Reference? Cogn. Neurosci. 2016, 7, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Herbert, C.; Pauli, P.; Herbert, B.M. Self-Reference Modulates the Processing of Emotional Stimuli in the Absence of Explicit Self-Referential Appraisal Instructions. Soc. Cogn. Affect. Neurosci. 2011, 6, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.P.; Macedo, J.; Oliveira, H.M.; Lages, A.; Hernández-Cabrera, J.; Pinheiro, A.P. Self-Reference Is a Fast-Acting Automatic Mechanism on Emotional Word Processing: Evidence from a Masked Priming Affective Categorisation Task. J. Cogn. Psychol. 2019, 31, 317–325. [Google Scholar] [CrossRef]
- Humphreys, G.W.; Sui, J. Attentional Control and the Self: The Self-Attention Network (SAN). Cogn. Neurosci. 2016, 7, 5–17. [Google Scholar] [CrossRef]
- Sui, J.; Gu, X. Self as Object: Emerging Trends in Self Research. Trends Neurosci. 2017, 40, 643–653. [Google Scholar] [CrossRef]
- Sui, J.; Rotshtein, P. Self-Prioritization and the Attentional Systems. Curr. Opin. Psychol. 2019, 29, 148–152. [Google Scholar] [CrossRef]
- Scalabrini, A.; Mucci, C.; Northoff, G. Is Our Self Related to Personality? A Neuropsychodynamic Model. Front. Hum. Neurosci. 2018, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, P.L.; Wittmann, M.K.; Apps, M.A.; Klein-Flügge, M.C.; Crockett, M.J.; Humphreys, G.W.; Rushworth, M.F. Neural Mechanisms for Learning Self and Other Ownership. Nat. Commun. 2018, 9, 4747. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Wang, M.; Northoff, G. Linking Bodily, Environmental and Mental States in the Self—A Three-Level Model Based on a Meta-Analysis. Neurosci. Biobehav. Rev. 2020, 115, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Soch, J.; Deserno, L.; Assmann, A.; Barman, A.; Walter, H.; Richardson-Klavehn, A.; Schott, B.H. Inhibition of Information Flow to the Default Mode Network during Self-Reference versus Reference to Others. Cereb. Cortex 2017, 27, 3930–3942. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, T.W.; Johnson, S.C. Self-Appraisal Decisions Evoke Dissociated Dorsal—Ventral aMPFC Networks. Neuroimage 2006, 30, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Yaoi, K.; Osaka, M.; Osaka, N. Neural Correlates of the Self-Reference Effect: Evidence from Evaluation and Recognition Processes. Front. Hum. Neurosci. 2015, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Haciahmet, C.C.; Golubickis, M.; Schäfer, S.; Frings, C.; Pastötter, B. The Oscillatory Fingerprints of Self-prioritization: Novel Markers in Spectral EEG for Self-relevant Processing. Psychophysiology 2023, 60, e14396. [Google Scholar] [CrossRef] [PubMed]
- Davey, C.G.; Pujol, J.; Harrison, B.J. Mapping the Self in the Brain’s Default Mode Network. NeuroImage 2016, 132, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Damme, K.S.; Pelletier-Baldelli, A.; Cowan, H.R.; Orr, J.M.; Mittal, V.A. Distinct and Opposite Profiles of Connectivity during Self-Reference Task and Rest in Youth at Clinical High Risk for Psychosis. Hum. Brain Mapp. 2019, 40, 3254–3264. [Google Scholar] [CrossRef]
- Gaubert, M.; Villain, N.; Landeau, B.; Mezenge, F.; Egret, S.; Perrotin, A.; Belliard, S.; de La Sayette, V.; Eustache, F.; Desgranges, B. Neural Correlates of Self-Reference Effect in Early Alzheimer’s Disease. J. Alzheimers Dis. 2017, 56, 717–731. [Google Scholar] [CrossRef]
- Sui, J.; Chechlacz, M.; Humphreys, G.W. Dividing the Self: Distinct Neural Substrates of Task-Based and Automatic Self-Prioritization after Brain Damage. Cognition 2012, 122, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Chechlacz, M.; Rotshtein, P.; Humphreys, G.W. Lesion-Symptom Mapping of Self-Prioritization in Explicit Face Categorization: Distinguishing Hypo-and Hyper-Self-Biases. Cereb. Cortex 2015, 25, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; He, X.; Yi, D.; Sui, J. Temporal Properties of Self-Prioritization. Entropy 2024, 26, 242. [Google Scholar] [CrossRef]
- Sui, J.; He, X.; Golubickis, M.; Svensson, S.L.; Macrae, C.N. Electrophysiological Correlates of Self-Prioritization. Conscious. Cogn. 2023, 108, 103475. [Google Scholar] [CrossRef]
- Frewen, P.; Schroeter, M.L.; Riva, G.; Cipresso, P.; Fairfield, B.; Padulo, C.; Kemp, A.H.; Palaniyappan, L.; Owolabi, M.; Kusi-Mensah, K. Neuroimaging the Consciousness of Self: Review, and Conceptual-Methodological Framework. Neurosci. Biobehav. Rev. 2020, 112, 164–212. [Google Scholar] [CrossRef]
- Northoff, G.; Heinzel, A.; De Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-Referential Processing in Our Brain—A Meta-Analysis of Imaging Studies on the Self. Neuroimage 2006, 31, 440–457. [Google Scholar] [CrossRef]
- Murray, R.J.; Schaer, M.; Debbané, M. Degrees of Separation: A Quantitative Neuroimaging Meta-Analysis Investigating Self-Specificity and Shared Neural Activation between Self- and Other-Reflection. Neurosci. Biobehav. Rev. 2012, 36, 1043–1059. [Google Scholar] [CrossRef]
- D’Argembeau, A.; Collette, F.; Van der Linden, M.; Laureys, S.; Del Fiore, G.; Degueldre, C.; Luxen, A.; Salmon, E. Self-Referential Reflective Activity and Its Relationship with Rest: A PET Study. Neuroimage 2005, 25, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Northoff, G. How Is Our Self Related to Midline Regions and the Default-Mode Network? NeuroImage 2011, 57, 1221–1233. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Moran, J.M.; Nieto-Castañón, A.; Triantafyllou, C.; Saxe, R.; Gabrieli, J.D. Associations and Dissociations between Default and Self-Reference Networks in the Human Brain. Neuroimage 2011, 55, 225–232. [Google Scholar] [CrossRef]
- Kolvoort, I.R.; Wainio-Theberge, S.; Wolff, A.; Northoff, G. Temporal Integration as “Common Currency” of Brain and Self-Scale-free Activity in Resting-state EEG Correlates with Temporal Delay Effects on Self-relatedness. Hum. Brain Mapp. 2020, 41, 4355–4374. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.; Di Giovanni, D.A.; Gómez-Pilar, J.; Nakao, T.; Huang, Z.; Longtin, A.; Northoff, G. The Temporal Signature of Self: Temporal Measures of Resting-state EEG Predict Self-consciousness. Hum. Brain Mapp. 2019, 40, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Rotshtein, P.; Humphreys, G.W. Coupling Social Attention to the Self Forms a Network for Personal Significance. Proc. Natl. Acad. Sci. USA 2013, 110, 7607–7612. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Bermpohl, F. Cortical Midline Structures and the Self. Trends Cogn. Sci. 2004, 8, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Chavez, R.S.; Heatherton, T.F.; Wagner, D.D. Neural Population Decoding Reveals the Intrinsic Positivity of the Self. Cereb. Cortex 2017, 27, 5222–5229. [Google Scholar] [CrossRef] [PubMed]
- Heleven, E.; Van Overwalle, F. The Neural Representation of the Self in Relation to Close Others Using fMRI Repetition Suppression. Soc. Neurosci. 2019, 14, 717–728. [Google Scholar] [CrossRef] [PubMed]
- D’Argembeau, A. On the Role of the Ventromedial Prefrontal Cortex in Self-Processing: The Valuation Hypothesis. Front. Hum. Neurosci. 2013, 7, 372. [Google Scholar] [CrossRef]
- Samson, D.; Apperly, I.A.; Chiavarino, C.; Humphreys, G.W. Left Temporoparietal Junction Is Necessary for Representing Someone Else’s Belief. Nat. Neurosci. 2004, 7, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.; Xiao, Y.; Xie, H.; Manwaring, S.S.; Farmer, C.; Thompson, L.; D’Souza, P.; Thurm, A.; Redcay, E. Posterior Superior Temporal Cortex Connectivity Is Related to Social Communication in Toddlers. Infant Behav. Dev. 2023, 71, 101831. [Google Scholar] [CrossRef]
- Martínez-Pérez, V.; Campoy, G.; Palmero, L.B.; Fuentes, L.J. Examining the Dorsolateral and Ventromedial Prefrontal Cortex Involvement in the Self-Attention Network: A Randomized, Sham-Controlled, Parallel Group, Double-Blind, and Multichannel HD-tDCS Study. Front. Neurosci. 2020, 14, 683. [Google Scholar] [CrossRef]
- Leszkowicz, E.; Maio, G.R.; Linden, D.E.J.; Ihssen, N. Neural Coding of Human Values Is Underpinned by Brain Areas Representing the Core Self in the Cortical Midline Region. Soc. Neurosci. 2021, 16, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Isoda, M. The Role of the Medial Prefrontal Cortex in Moderating Neural Representations of Self and Other in Primates. Annu. Rev. Neurosci. 2021, 44, 295–313. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Liu, M.; Mevorach, C.; Humphreys, G.W. The Salient Self: The Left Intraparietal Sulcus Responds to Social as Well as Perceptual-Salience after Self-Association. Cereb. Cortex 2015, 25, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Bi, T.; Chen, A.; Egner, T. Ventromedial Prefrontal Cortex Drives the Prioritization of Self-Associated Stimuli in Working Memory. J. Neurosci. 2021, 41, 2012–2023. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.; Frings, C. Searching for the Inner Self: Evidence against a Direct Dependence of the Self-Prioritization Effect on the Ventro-Medial Prefrontal Cortex. Exp. Brain Res. 2019, 237, 247–256. [Google Scholar] [CrossRef]
- Liang, Q.; Zhang, B.; Fu, S.; Sui, J.; Wang, F. The Roles of the LpSTS and DLPFC in Self-prioritization: A Transcranial Magnetic Stimulation Study. Hum. Brain Mapp. 2022, 43, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Soutschek, A.; Ruff, C.C.; Strombach, T.; Kalenscher, T.; Tobler, P.N. Brain Stimulation Reveals Crucial Role of Overcoming Self-Centeredness in Self-Control. Sci. Adv. 2016, 2, e1600992. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Enock, F.; Ralph, J.; Humphreys, G.W. Dissociating Hyper and Hypoself Biases to a Core Self-Representation. Cortex 2015, 70, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Abdi, H.; Williams, L.J. Principal Component Analysis. WIREs Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Bickerton, W.-L.; Riddoch, M.J.; Samson, D.; Balani, A.B.; Mistry, B.; Humphreys, G.W. Systematic Assessment of Apraxia and Functional Predictions from the Birmingham Cognitive Screen. J. Neurol. Neurosurg. Psychiatry 2012, 83, 513–521. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified Segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Thiebaut de Schotten, M.; Tomaiuolo, F.; Aiello, M.; Merola, S.; Silvetti, M.; Lecce, F.; Bartolomeo, P.; Doricchi, F. Damage to White Matter Pathways in Subacute and Chronic Spatial Neglect: A Group Study and 2 Single-Case Studies with Complete Virtual “in Vivo” Tractography Dissection. Cereb. Cortex 2014, 24, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Seghier, M.L.; Ramlackhansingh, A.; Crinion, J.; Leff, A.P.; Price, C.J. Lesion Identification Using Unified Segmentation-Normalisation Models and Fuzzy Clustering. NeuroImage 2008, 41, 1253–1266. [Google Scholar] [CrossRef]
- Humphreys, G.W. BCoS Brain Behaviour Analysis; Psychology Press: Hove, UK, 2012. [Google Scholar]
- Fabrigar, L.R.; Wegener, D.T.; MacCallum, R.C.; Strahan, E.J. Evaluating the Use of Exploratory Factor Analysis in Psychological Research. Psychol. Methods 1999, 4, 272–299. [Google Scholar] [CrossRef]
- Kim, J.-O.; Mueller, C.W. Factor Analysis: Statistical Methods and Practical Issues; SAGE: Beverly Hills, CA, USA, 1978; ISBN 978-0-8039-1166-6. [Google Scholar]
- Bartlett, M.S.; Fowler, R.H. Properties of Sufficiency and Statistical Tests. Proc. R. Soc. Lond. Ser.-Math. Phys. Sci. 1997, 160, 268–282. [Google Scholar] [CrossRef]
- Kaiser, H.F.; Rice, J. Little Jiffy, Mark Iv. Educ. Psychol. Meas. 1974, 34, 111–117. [Google Scholar] [CrossRef]
- Crinion, J.; Ashburner, J.; Leff, A.; Brett, M.; Price, C.; Friston, K. Spatial Normalization of Lesioned Brains: Performance Evaluation and Impact on fMRI Analyses. NeuroImage 2007, 37, 866–875. [Google Scholar] [CrossRef]
- Worsley, K.J. Developments in Random Field Theory. Hum. Brain Funct. 2003, 2, 881–886. [Google Scholar]
- Leff, A.P.; Schofield, T.M.; Crinion, J.T.; Seghier, M.L.; Grogan, A.; Green, D.W.; Price, C.J. The Left Superior Temporal Gyrus Is a Shared Substrate for Auditory Short-Term Memory and Speech Comprehension: Evidence from 210 Patients with Stroke. Brain 2009, 132, 3401–3410. [Google Scholar] [CrossRef]
- Chechlacz, M.; Terry, A.; Demeyere, N.; Douis, H.; Bickerton, W.-L.; Rotshtein, P.; Humphreys, G.W. Common and Distinct Neural Mechanisms of Visual and Tactile Extinction: A Large Scale VBM Study in Sub-Acute Stroke. NeuroImage Clin. 2013, 2, 291–302. [Google Scholar] [CrossRef]
- Chechlacz, M.; Rotshtein, P.; Bickerton, W.-L.; Hansen, P.C.; Deb, S.; Humphreys, G.W. Separating Neural Correlates of Allocentric and Egocentric Neglect: Distinct Cortical Sites and Common White Matter Disconnections. Cogn. Neuropsychol. 2010, 27, 277–303. [Google Scholar] [CrossRef]
- The MathWorks Inc. MATLAB, Version: 7.14.0 (R2012a); The MathWorks Inc.: Natick, MA, USA, 2012. Available online: https://www.mathworks.com(accessed on 28 May 2024).
- Ashburner, J.; Friston, K.J. Voxel-Based Morphometry—The Methods. Neuroimage 2000, 11, 805–821. [Google Scholar] [CrossRef]
- Kiebel, S.; Holmes, A. The General Linear Model; Elsevier: Amsterdam, The Netherlands, 2007; Chapter 8. [Google Scholar]
- Duvernoy, H.M. The Human Brain: Surface, Three-Dimensional Sectional Anatomy with MRI, and Blood Supply; Springer Science & Business Media: Vienna, Austria, 1999. [Google Scholar]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Woolsey, T.A.; Hanaway, J.; Gado, M.H. The Brain Atlas: A Visual Guide to the Human Central Nervous System; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Thiebaut de Schotten, M.; Ffytche, D.H.; Bizzi, A.; Dell’Acqua, F.; Allin, M.; Walshe, M.; Murray, R.; Williams, S.C.; Murphy, D.G.; Catani, M. Atlasing Location, Asymmetry and Inter-Subject Variability of White Matter Tracts in the Human Brain with MR Diffusion Tractography. Neuroimage 2010, 54, 49–59. [Google Scholar] [CrossRef] [PubMed]
- de Schotten, M.T.; Dell’Acqua, F.; Forkel, S.J.; Simmons, A.; Vergani, F.; Murphy, D.G.M.; Catani, M. A Lateralized Brain Network for Visuospatial Attention. Nat. Neurosci. 2011, 14, 1245–1246. [Google Scholar] [CrossRef] [PubMed]
- Rojkova, K.; Volle, E.; Urbanski, M.; Humbert, F.; Dell’Acqua, F.; de Schotten, M.T. Atlasing the frontal lobe connections and their variability due to age and education: A spherical deconvolution tractography study. Brain Struct. Funct. 2016, 221, 1751–1766. [Google Scholar] [CrossRef]
- Sui, J. Self-Reference Acts as a Golden Thread in Binding. Trends Cogn. Sci. 2016, 20, 482–483. [Google Scholar] [CrossRef]
- Sui, J.; Humphreys, G.W. The Integrative Self: How Self-Reference Integrates Perception and Memory. Trends Cogn. Sci. 2015, 19, 719–728. [Google Scholar] [CrossRef]
- Stendardi, D.; Biscotto, F.; Bertossi, E.; Ciaramelli, E. Present and Future Self in Memory: The Role of vmPFC in the Self-Reference Effect. Soc. Cogn. Affect. Neurosci. 2021, 16, 1205–1213. [Google Scholar] [CrossRef]
- Philippi, C.L.; Duff, M.C.; Denburg, N.L.; Tranel, D.; Rudrauf, D. Medial PFC Damage Abolishes the Self-Reference Effect. J. Cogn. Neurosci. 2012, 24, 475–481. [Google Scholar] [CrossRef]
- Wagner, D.D.; Haxby, J.V.; Heatherton, T.F. The Representation of Self and Person Knowledge in the Medial Prefrontal Cortex. WIREs Cogn. Sci. 2012, 3, 451–470. [Google Scholar] [CrossRef]
- Pizzagalli, D.A.; Roberts, A.C. Prefrontal Cortex and Depression. Neuropsychopharmacology 2022, 47, 225–246. [Google Scholar] [CrossRef]
- George, M.S.; Ketter, T.A.; Post, R.M. Prefrontal Cortex Dysfunction in Clinical Depression. Depression 1994, 2, 59–72. [Google Scholar] [CrossRef]
- Murray, E.A.; Wise, S.P.; Drevets, W.C. Localization of Dysfunction in Major Depressive Disorder: Prefrontal Cortex and Amygdala. Biol. Psychiatry 2011, 69, e43–e54. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, Y.; Li, M.; Qin, W.; Xie, Y.; Zhao, G.; Wang, Y.; Yang, C.; Zhang, B.; Jing, Y. Childhood Sexual Abuse Related to Brain Activity Abnormalities in Right Inferior Temporal Gyrus among Major Depressive Disorder. Neurosci. Lett. 2023, 806, 137196. [Google Scholar] [CrossRef]
- Liu, Y.S.; Song, Y.; Lee, N.A.; Bennett, D.M.; Button, K.S.; Greenshaw, A.; Cao, B.; Sui, J. Depression Screening Using a Non-Verbal Self-Association Task: A Machine-Learning Based Pilot Study. J. Affect. Disord. 2022, 310, 87–95. [Google Scholar] [CrossRef]
- Sui, J.; Humphreys, G.W. The Self Survives Extinction: Self-Association Biases Attention in Patients with Visual Extinction. Cortex 2017, 95, 248–256. [Google Scholar] [CrossRef]
- Molnar-Szakacs, I.; Uddin, L.Q. Self-Processing and the Default Mode Network: Interactions with the Mirror Neuron System. Front. Hum. Neurosci. 2013, 7, 571. [Google Scholar] [CrossRef]
- Knyazev, G. EEG Correlates of Self-Referential Processing. Front. Hum. Neurosci. 2013, 7, 264. [Google Scholar] [CrossRef]
- Tian, Y.; Margulies, D.S.; Breakspear, M.; Zalesky, A. Topographic Organization of the Human Subcortex Unveiled with Functional Connectivity Gradients. Nat. Neurosci. 2020, 23, 1421–1432. [Google Scholar] [CrossRef]
- Caverzasi, E.; Papinutto, N.; Amirbekian, B.; Berger, M.S.; Henry, R.G. Q-Ball of Inferior Fronto-Occipital Fasciculus and Beyond. PLoS ONE 2014, 9, e100274. [Google Scholar] [CrossRef]
- Martino, J.; Brogna, C.; Robles, S.G.; Vergani, F.; Duffau, H. Anatomic Dissection of the Inferior Fronto-Occipital Fasciculus Revisited in the Lights of Brain Stimulation Data. Cortex 2010, 46, 691–699. [Google Scholar] [CrossRef]
- Zhang, F.; Daducci, A.; He, Y.; Schiavi, S.; Seguin, C.; Smith, R.E.; Yeh, C.-H.; Zhao, T.; O’Donnell, L.J. Quantitative Mapping of the Brain’s Structural Connectivity Using Diffusion MRI Tractography: A Review. Neuroimage 2022, 249, 118870. [Google Scholar] [CrossRef]
- Sui, J.; Humphreys, G.W. Self-Referential Processing Is Distinct from Semantic Elaboration: Evidence from Long-Term Memory Effects in a Patient with Amnesia and Semantic Impairments. Neuropsychologia 2013, 51, 2663–2673. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Sui, J. Decoding Individual Differences in Self-Prioritization from the Resting-State Functional Connectome. NeuroImage 2023, 276, 120205. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Size (Voxels) | Z-Score | Coordinates (X, Y, Z) | Brain Structure |
|---|---|---|---|---|
| Automatic self-reference | ||||
| 1384 | 3.44 | 34 26 −42 | Right ITG 1 | |
| 50 | 3.41 | −10 50 4 | Left vmPFC 2 | |
| Control processing | ||||
| 405 | 3.93 | 44 −46 62 | Right IPL 3 | |
| 275 | 3.74 | −28 −66 52 | Left IPL 3 | |
| 343 | 3.39 | −46 −10 4 | Left AIC 4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, J.; Rotshtein, P.; Lu, Z.; Chechlacz, M. Causal Roles of Ventral and Dorsal Neural Systems for Automatic and Control Self-Reference Processing: A Function Lesion Mapping Study. J. Clin. Med. 2024, 13, 4170. https://doi.org/10.3390/jcm13144170
Sui J, Rotshtein P, Lu Z, Chechlacz M. Causal Roles of Ventral and Dorsal Neural Systems for Automatic and Control Self-Reference Processing: A Function Lesion Mapping Study. Journal of Clinical Medicine. 2024; 13(14):4170. https://doi.org/10.3390/jcm13144170
Chicago/Turabian StyleSui, Jie, Pia Rotshtein, Zhuoen Lu, and Magdalena Chechlacz. 2024. "Causal Roles of Ventral and Dorsal Neural Systems for Automatic and Control Self-Reference Processing: A Function Lesion Mapping Study" Journal of Clinical Medicine 13, no. 14: 4170. https://doi.org/10.3390/jcm13144170