
Dietary and Nutritional Strategies to Prevent Uremic Toxin Formation and Slow the Progression of Diabetic Kidney Disease

 ,
,  , ,
, , 
Abstract
1. Introduction
- Renin–Angiotensin–Aldosterone System (RAAS) Inhibition: ACE inhibitors or ARBs are recommended in patients with albuminuria to reduce proteinuria and control blood pressure. A RAAS blockade remains foundational in slowing DKD progression.
- Sodium–glucose co-transporter 2 inhibitors are recommended for all patients with proteinuric CKD, irrespective of their diabetes status, due to their proven nephroprotective and cardioprotective effects.
- Non-steroidal Mineralocorticoid Receptor Antagonist (n-MRA): finerenone, as the only approved agent in this class for DKD, is recommended for patients with T2DM and DKD. It provides an additional renal and cardiovascular benefit beyond RAAS inhibition.
- Glucagon-like peptide-1 receptor agonists are advised in patients with type 2 diabetes and DKD, particularly when glycemic control, weight reduction, or further cardiovascular protection is indicated.
1.1. Uremic Toxins—The Role of the Gut–Kidney Axis
1.2. Intestinal Dysbiosis
1.3. Targeted Supplementation
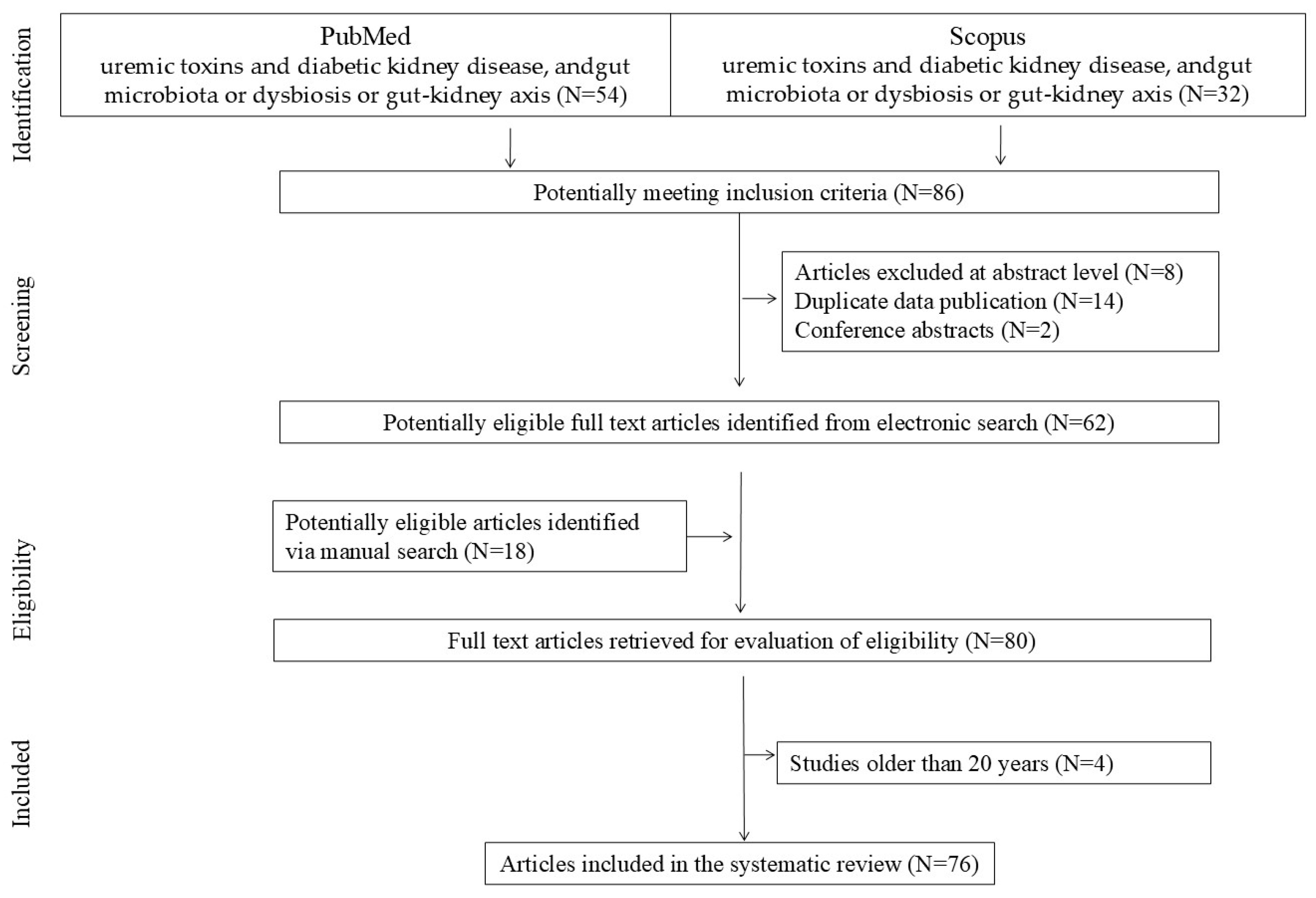
2. Materials and Methods
3. Therapeutic Dietary Interventions for Microbiota in DKD
3.1. Prebiotics
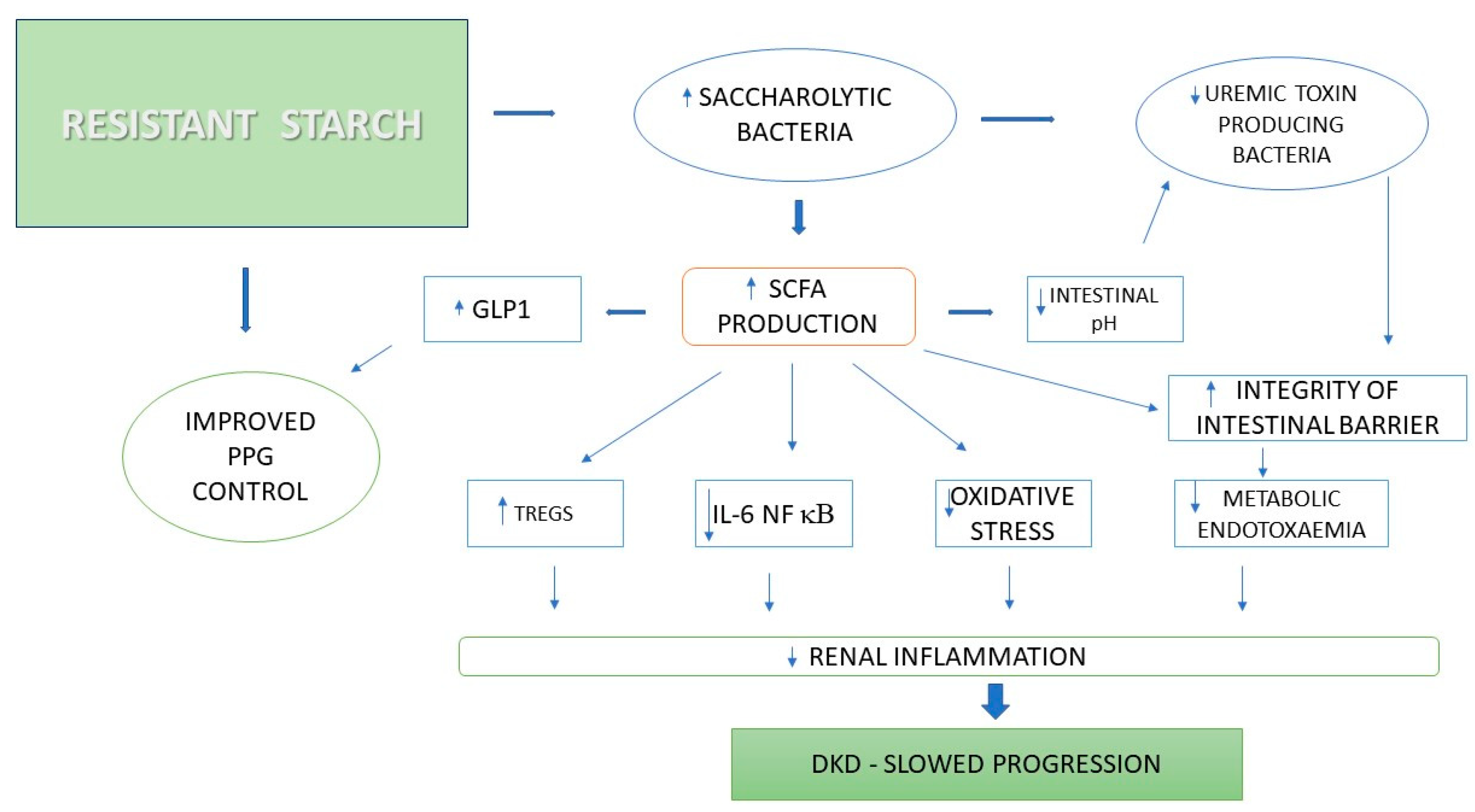
3.1.1. Resistant Starch
3.1.2. Fructooligosaccharides
3.1.3. Arabic Gum, Polydextrose, and Glucomannan
3.2. Research Directions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jager, K.J.; Kovesdy, C.; Langham, R.; Rosenberg, M.; Jha, V.; Zoccali, C. A single number for advocacy and communication-worldwide more than 850 million individuals have kidney diseases. Kidney Int. 2019, 96, 1048–1050. [Google Scholar] [CrossRef] [PubMed]
- Hoogeveen, E.K. The epidemiology of diabetic kidney disease. Kidney Dial. 2022, 2, 433–442. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C.; et al. IDF diabetes atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Koye, D.N.; Shaw, J.E.; Reid, C.M.; Atkins, R.C.; Reutens, A.T.; Magliano, D.J. Incidence of chronic kidney disease among people with diabetes: A systematic review of observational studies. Diabet. Med. 2017, 34, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.T.; Xu, X.; Lim, P.S.; Hung, K.Y. Worldwide epidemiology of diabetes-related end-stage renal disease, 2000–2015. Diabetes Care 2021, 44, 89–97. [Google Scholar] [CrossRef]
- Levin, A.; Ahmed, S.B.; Carrero, J.J.; Foster, B.; Francis, A.; Hall, R.K.; Herrington, W.G.; Hill, G.; Inker, L.A.; Kazancıoğlu, R.; et al. KDIGO 2024 clinical practice guideline for the management of diabetes in chronic kidney disease. Kidney Int. 2024, 105 (Suppl. 1), S1–S127. [Google Scholar] [CrossRef]
- Lewis, E.J.; Hunsicker, L.G.; Clarke, W.R.; Berl, T.; Pohl, M.A.; Lewis, J.B.; Ritz, E.; Atkins, R.C.; Rohde, R.; Raz, I.; et al. Renoprotective effect of the angiotensin-receptor antagonist irbesartan in patients with nephropathy due to type 2 diabetes. N. Engl. J. Med. 2001, 345, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.M.; Cooper, M.E.; De Zeeuw, D.; Keane, W.F.; Mitch, W.E.; Parving, H.-H.; Remuzzi, G.; Snapinn, S.M.; Zhang, Z.; Shahinfar, S. Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathy. N. Engl. J. Med. 2001, 345, 861–869. [Google Scholar] [CrossRef]
- Patel, A. ADVANCE collaborative group. Effects of a fixed combination of perindopril and indapamide on macrovascular and microvascular outcomes in patients with type 2 diabetes mellitus (the ADVANCE trial): A randomised controlled trial. Lancet 2007, 370, 829–840. [Google Scholar] [CrossRef]
- Neal, B.; Perkovic, V.; Mahaffey, K.W.; de Zeeuw, D.; Fulcher, G.; Erondu, N.; Shaw, W.; Law, G.; Desai, M.; Matthews, D.R.; et al. Canagliflozin and cardiovascular and renal events in type 2 diabetes. N. Engl. J. Med. 2017, 377, 644–657. [Google Scholar] [CrossRef]
- Heerspink, H.J.L.; Stefánsson, B.V.; Correa-Rotter, R.; Chertow, G.M.; Greene, T.; Hou, F.F.; Mann, J.F.E.; Mcmurray, J.J.V.; Lindberg, M.; Rossing, P.; et al. DAPA-CKD trial committees and investigators. Dapagliflozin in patients with chronic kidney disease. N. Engl. J. Med. 2020, 383, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- The EMPA-KIDNEY Collaborative Group; Herrington, W.G.; Staplin, N.; Wanner, C.; Green, J.B.; Hauske, S.J.; Emberson, J.R.; Preiss, D.; Judge, P.; Mayne, K.J.; et al. Empagliflozin in patients with chronic kidney disease. N. Engl. J. Med. 2023, 388, 117–127. [Google Scholar]
- Bakris, G.L.; Agarwal, R.; Anker, S.D.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Nowack, C.; Schloemer, P.; Joseph, A.; et al. Effect of finerenone on chronic kidney disease outcomes in type 2 diabetes. N. Engl. J. Med. 2020, 383, 2219–2229. [Google Scholar] [CrossRef]
- Perkovic, V.; Tuttle, K.R.; Rossing, P.; Mahaffey, K.W.; Mann, J.F.E.; Bakris, G.; Baeres, F.M.M.; Idorn, T.; Bosch-Traberg, H.; Lausvig, N.L.; et al. Effects of semaglutide on chronic kidney disease in patients with type 2 diabetes. N. Engl. J. Med. 2024, 391, 109–121. [Google Scholar] [CrossRef]
- Bosch, C.C.S.; Soler, M.J.; Ortiz, A.; Fernandez-Fernandez, B. Tirzepatide and prevention of chronic kidney disease. Clin. Kidney J. 2022, 16, sfac274. [Google Scholar] [CrossRef]
- Naaman, S.C.; Bakris, G.L. Diabetic nephropathy: Update on pillars of therapy slowing progression. Diabetes Care 2023, 46, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, K. Kidney disease: Improving global outcomes diabetes working group. KDIGO 2022 clinical practice guideline for diabetes management in chronic kidney disease. Kidney Int. 2022, 102, S1–S127. [Google Scholar]
- Gao, Q.; Tan, N.C.; Lee, M.L.; Hsu, W.; Choo, J. Comparative effectiveness of first-line antihypertensive drug classes on the maintenance of estimated glomerular filtration rate (egfr) in real world primary care. Sci. Rep. 2023, 13, 21223. [Google Scholar] [CrossRef]
- Watanabe, K.; Sato, E.; Mishima, E.; Miyazaki, M.; Tanaka, T. What’s new in the molecular mechanisms of diabetic kidney disease: Recent advances. Int. J. Mol. Sci. 2023, 24, 570. [Google Scholar] [CrossRef]
- Paulmann, N.; Grohmann, M.; Voigt, J.-P.; Bert, B.; Vowinckel, J.; Bader, M.; Skelin, M.; Jevšek, M.; Fink, H.; Rupnik, M.; et al. Intracellular serotonin modulates insulin secretion from pancreatic beta-cells by protein serotonylation. PLoS Biol. 2009, 7, e1000229. [Google Scholar] [CrossRef]
- Allalou, A.; Nalla, A.; Prentice, K.J.; Liu, Y.; Zhang, M.; Dai, F.F.; Ning, X.; Osborne, L.R.; Cox, B.J.; Gunderson, E.P.; et al. A predictive metabolic signature for the transition from gestational diabetes mellitus to type 2 diabetes. Diabetes 2016, 65, 2529–2539. [Google Scholar] [CrossRef]
- López-Tenorio, I.I.; Aguilar-Villegas, Ó.R.; Espinoza-Palacios, Y.; Segura-Real, L.; Peña-Aparicio, B.; Amedei, A.; Aguirre-García, M.M. Primary prevention strategy for non-communicable diseases (ncds) and their risk factors: The role of intestinal microbiota. Biomedicines 2024, 12, 2529. [Google Scholar] [CrossRef] [PubMed]
- Mudimela, S.; Vishwanath, N.K.; Pillai, A.; Morales, R.; Marrelli, S.P.; Barichello, T.; Giridharan, V.V. Clinical significance and potential role of trimethylamine n-oxide in neurological and neuropsychiatric disorders. Drug Discov. Today 2022, 27, 103334. [Google Scholar] [CrossRef] [PubMed]
- Cheng, E.; Hung, S.C.; Lin, T.Y. Association of trimethylamine n-oxide and metabolites with kidney function decline in patients with chronic kidney disease. Clin. Nutr. 2025, 44, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Linnane, A.W.; Gobe, G.C. From the gastrointestinal tract (GIT) to the kidneys: Live bacterial cultures (probiotics) mediating reductions of uremic toxin levels via free radical signaling. Toxins 2013, 5, 2042–2057. [Google Scholar] [CrossRef]
- Graboski, A.L.; Redinbo, M.R. Gut-derived protein-bound uremic toxins. Toxins 2020, 12, 590. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; Desantis, T.Z.; Ni, Z.; Nguyen, T.-H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef]
- Ramezani, A.; Raj, D.S. The gut microbiome, kidney disease, and targeted interventions. J. Am. Soc. Nephrol. 2013, 25, 657–670. [Google Scholar] [CrossRef]
- Vaziri, N.D. CKD impairs barrier function and alters microbial flora of the intestine: A major link to inflammation and uremic toxicity. Curr. Opin. Nephrol. Hypertens. 2012, 21, 587–592. [Google Scholar] [CrossRef]
- Barrios, C.; Beaumont, M.; Pallister, T.; Villar, J.; Goodrich, J.K.; Clark, A.; Pascual, J.; Ley, R.E.; Spector, T.D.; Bell, J.T.; et al. Gut-microbiota-metabolite axis in early renal function decline. PLoS ONE 2015, 10, e0134311. [Google Scholar] [CrossRef]
- Fukuuchi, F.; Hida, M.; Aiba, Y.; Koga, Y.; Endoh, M.; Kurokawa, K.; Sakai, H. Intestinal bacteria-derived putrefactants in chronic renal failure. Clin. Exp. Nephrol. 2002, 6, 99–104. [Google Scholar] [CrossRef]
- Strid, H.; Simrén, M.; Stotzer, P.O.; Ringström, G.; Abrahamsson, H.; Björnsson, E.S. Patients with chronic renal failure have abnormal small intestinal motility and a high prevalence of small intestinal bacterial overgrowth. Digestion 2003, 67, 129–137. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, P.; Jiang, H.; Cheng, S. Gut bacterial translocation contributes to microinflammation in experimental uremia. Dig. Dis. Sci. 2012, 57, 2856–2862. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Piceno, Y.M.; Desantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2015, 39, 230–237. [Google Scholar] [CrossRef]
- Peters, B.A.; Qi, Q.; Usyk, M.; Daviglus, M.L.; Cai, J.; Franceschini, N.; Lash, J.P.; Gellman, M.D.; Yu, B.; Boerwinkle, E.; et al. Association of the gut microbiome with kidney function and damage in the hispanic community health study/study of latinos (HCHS/SOL). Gut Microbes. 2023, 15, 2186685. [Google Scholar] [CrossRef]
- Gargiulo, S.; Barone, V.; Bonente, D.; Tamborrino, T.; Inzalaco, G.; Gherardini, L.; Bertelli, E.; Chiariello, M. Integrated ultrasound characterization of the diet-induced obesity (DIO) model in young adult c57bl/6j mice: Assessment of cardiovascular, renal and hepatic changes. J. Imaging. 2024, 10, 217. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Wang, Y.; Xu, L.-X.; Yang, J.; Zhao, Y.; Qiao, J.; Li, N.; Li, Y.; Lv, D.-Q.; Sun, W.-Y. Relationship between dietary patterns and diabetic microvascular complications in patients with type 2 diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 8780–8794. [Google Scholar] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The international scientific association of probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Cornall, L.M.; Mathai, M.L.; Hryciw, D.H.; Mcainch, A.J. The therapeutic potential of GPR43: A novel role in modulating metabolic health. Cell Mol. Life Sci. 2013, 70, 4759–4770. [Google Scholar] [CrossRef]
- Jiao, A.; Zhao, Y.; Chu, L.; Yang, Y.; Jin, Z. A review on animal and plant proteins in regulating diabetic kidney disease: Mechanism of action and future perspectives. J. Funct. Foods 2024, 119, 106353. [Google Scholar] [CrossRef]
- Guida, B.; Germanò, R.; Trio, R.; Russo, D.; Memoli, B.; Grumetto, L.; Barbato, F.; Cataldi, M. Effect of short-term synbiotic treatment on plasma p-cresol levels in patients with chronic renal failure: A randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Poesen, R.; Evenepoel, P.; de Loor, H.; Delcour, J.A.; Courtin, C.M.; Kuypers, D.; Augustijns, P.; Verbeke, K.; Meijers, B. The influence of prebiotic arabinoxylan oligosaccharides on microbiota derived uremic retention solutes in patients with chronic kidney disease: A randomized controlled trial. PLoS ONE 2016, 11, e0153893. [Google Scholar] [CrossRef]
- Meijers, B.K.; de Preter, V.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. P-cresyl sulfate serum concentrations in haemodialysis patients are reduced by the prebiotic oligofructose-enriched inulin. Nephrol. Dial. Transplant. 2010, 25, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Esgalhado, M.; Kemp, J.A.; Azevedo, R.; Paiva, B.R.; Stockler-Pinto, M.B.; Dolenga, C.J.; Borges, N.A.; Nakao, L.S.; Mafra, D. Could resistant starch supplementation improve inflammatory and oxidative stress biomarkers and uremic toxins levels in hemodialysis patients? A pilot randomized controlled trial. Food Funct. 2018, 9, 6508–6516. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; Plummer, N.S.; Gardner, C.D.; Hostetter, T.H.; Meyer, T.W. Effect of increasing dietary fiber on plasma levels of colon-derived solutes in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2014, 9, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Ren. Physiol. 2016, 310, 857–871. [Google Scholar] [CrossRef]
- Salarolli, R.T.; Alvarenga, L.; Cardozo, L.F.M.F.; Teixeira, K.T.R.; Moreira, L.d.S.G.; Lima, J.D.; Rodrigues, S.D.; Nakao, L.S.; Fouque, D.; Mafra, D. Can curcumin supplementation reduce plasma levels of gut-derived uremic toxins in hemodialysis patients? A pilot randomized, double-blind, controlled study. Int. Urol. Nephrol. 2021, 53, 1231–1238. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Tenore, G.C.; Novellino, E. Effects of grape pomace polyphenolic extract (taurisolo®) in reducing TMAO serum levels in humans: Preliminary results from a randomized, placebo-controlled, cross-over study. Nutrients 2019, 11, 139. [Google Scholar] [CrossRef]
- Tenore, G.C.; Caruso, D.; Buonomo, G.; D’avino, M.; Ciampaglia, R.; Maisto, M.; Schisano, C.; Bocchino, B.; Novellino, E. Lactofermented annurca apple puree as a functional food indicated for the control of plasma lipid and oxidative amine levels: Results from a randomised clinical trial. Nutrients 2019, 11, 122. [Google Scholar] [CrossRef]
- Wang, Z.; Peters, B.A.; Yu, B.; Grove, M.L.; Wang, T.; Xue, X.; Thyagarajan, B.; Daviglus, M.L.; Boerwinkle, E.; Hu, G.; et al. Gut microbiota and blood metabolites related to fiber intake and type 2 diabetes. Circ. Res. 2024, 134, 842–854. [Google Scholar] [CrossRef]
- Conlon, M.A.; Kerr, C.A.; Mcsweeney, C.S.; Dunne, R.A.; Shaw, J.M.; Kang, S.; Bird, A.R.; Morell, M.K.; Lockett, T.J.; Molloy, P.L.; et al. Odporne skrobie chronią przed uszkodzeniem kolonicznego DNA i zmieniają mikroflorę i ekspresję genów u szczurów karmionych zachodnią dietą. J. Nutr. 2012, 142, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Kellow, N.J.; Coughlan, M.T. Modulation of the gut microbiota by resistant starch as a treatment of chronic kidney diseases: Evidence of efficacy and mechanistic insights. Adv. Nutr. 2019, 10, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Szczuko, M.; Kikut, J.; Maciejewska, D.; Kulpa, D.; Celewicz, Z.; Ziętek, M. The associations of SCFA with anthropometric parameters and carbohydrate metabolism in pregnant women. Int. J. Mol. Sci. 2020, 21, 9212. [Google Scholar] [CrossRef]
- Shamloo, M.; Mollard, R.; Wang, H.; Kingra, K.; Tangri, N.; Mackay, D. A randomized double-blind cross-over trial to study the effects of resistant starch prebiotic in chronic kidney disease (respeckd). Trials 2022, 23, 72. [Google Scholar] [CrossRef] [PubMed]
- Metzler-zebeli, B.U.; Canibe, N.; Montagne, L.; Freire, J.; Bosi, P.; Prates, J.A.M.; Tanghe, S.; Trevisi, P. Resistant starch reduces large intestinal ph and promotes fecal lactobacilli and bifidobacteria in pigs. Animal 2019, 13, 64–73. [Google Scholar] [CrossRef]
- Sybille, T.; June, Z.; Michael, K.; Roy, M.; Maria, L.M. The intestinal microbiota in aged mice is modulated by dietary resistant starch and correlated with improvements in host responses. FEMS Microbiol. Ecol. 2013, 83, 299–309. [Google Scholar]
- Tian, E.; Wang, F.; Zhao, L.; Sun, Y.; Yang, J. The pathogenic role of intestinal flora metabolites in diabetic nephropathy. Front Physiol. 2023, 14, 1231621. [Google Scholar] [CrossRef]
- Tao, S.; Li, L.; Li, L.; Liu, Y.; Ren, Q.; Shi, M.; Liu, J.; Jiang, J.; Ma, H.; Huang, Z.; et al. Understanding the gut–kidney axis among biopsy-proven diabetic nephropathy, type 2 diabetes mellitus and healthy controls: An analysis of the gut microbiota composition. Acta Diabetol. 2019, 56, 581–592. [Google Scholar] [CrossRef]
- Martínez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant starches types 2 and 4 have differential effects on the composition of the fecal microbiota in human subjects. PLoS ONE 2010, 5, e15046. [Google Scholar] [CrossRef]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M.; Blaser, M.J.; Britton, R.; Walter, J. Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. Mbio 2019, 10, 10–128. [Google Scholar] [CrossRef]
- Venkataraman, A.; Sieber, J.R.; Schmidt, A.W.; Waldron, C.; Theis, K.R.; Schmidt, T.M. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome 2016, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Alfa, M.J.; Strang, D.; Tappia, P.S.; Graham, M.; van Domselaar, G.; Forbes, J.D.; Laminman, V.; Olson, N.; Degagne, P.; Bray, D.; et al. A randomized trial to determine the impact of a digestion resistant starch composition on the gut microbiome in older and mid-age adults. Clin. Nutr. 2018, 37, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Drake, A.M.; Coughlan, M.T.; Christophersen, C.T.; Snelson, M. Resistant starch as a dietary intervention to limit the progression of diabetic kidney disease. Nutrients 2022, 14, 4547. [Google Scholar] [CrossRef]
- Jia, X.H.; Wang, S.Y.; Sun, A.Q. Dietary fiber intake and its association with diabetic kidney disease in american adults with diabetes: A cross-sectional study. World J. Diabetes 2024, 15, 475–487. [Google Scholar] [CrossRef]
- Pengrattanachot, N.; Thongnak, L.; Lungkaphin, A. The impact of prebiotic fructooligosaccharides on gut dysbiosis and inflammation in obesity and diabetes related kidney disease. Food Funct. 2022, 13, 5925–5945. [Google Scholar] [CrossRef]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bauerlein, M.; Frohburg, C.; Landschutze, V.; Gibson, G.R. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long chain inulin extracted from globe artichoke (cynara scolymus) in healthy subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef]
- Ramnani, P.; Gaudier, E.; Bingham, M.; van Bruggen, P.; Tuohy, K.M.; Gibson, G.R. Prebiotic effect of fruit and vegetable shots containing jerusalem artichoke inulin: A human intervention study. Br. J. Nutr. 2010, 104, 233–240. [Google Scholar] [CrossRef]
- Calame, W.; Weseler, A.R.; Viebke, C.; Flynn, C.; Siemensma, A.D. Gum arabic establishes prebiotic functionality in healthy human volunteers in a dose-dependent manner. Br. J. Nutr. 2008, 100, 1269–1275. [Google Scholar] [CrossRef]
- Mirmiran, P.; Houshialsadat, Z.; Gaeini, Z.; Bahadoran, Z.; Azizi, F. Functional properties of beetroot (beta vulgaris) in management of cardio-metabolic diseases. Nutr. Metab. 2020, 17, 3. [Google Scholar] [CrossRef]
- Jie, Z.; Bang-yao, J.; Ming-jie, X.; Hai-wei, L.; Zu-kang, Z.; Ting-song, W.; Craig, S.A.S. Studies on the effects of polydextrose intake on physiologic functions in chinese people. Am. J. Clin. Nutr. 2000, 72, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Hengst, C.; Ptok, S.; Roessler, A.; Fechner, A.; Jahreis, G. Effects of polydextrose supplementation on different faecal parameters in healthy volunteers. Int. J. Food Sci. Nutr. 2009, 60, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lefranc-millot, C.; Gruerin-deremaux, L.; Wils, D.; Neut, C.; Miller, L.E.; Saniez-degrave, M.H. Impact of a resistant dextrin on intestinal ecology: How altering the digestive ecosystem with NUTRIOSE, a soluble fiber with prebiotic properties, may be beneficial for health. J. Int. Med. Res. 2012, 40, 211–224. [Google Scholar] [CrossRef]
- Chen, H.; Nie, Q.; Hu, J.; Huang, X.; Zhang, K.; Nie, S. Glucomannans alleviated the progression of diabetic kidney disease by improving kidney metabolic disturbance. Mol. Nutr. Food Res. 2019, 63, e1801008. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhou, T.; He, Y.; Xie, Y.; Xu, Y.; Huang, W. The role and mechanism of butyrate in the prevention and treatment of diabetic kidney disease. Front. Microbiol. 2022, 13, 961536. [Google Scholar] [CrossRef]
- Mafra, D.; Kemp, J.A.; Borges, N.A.; Wong, M.; Stenvinkel, P. Gut microbiota interventions to retain residual kidney function. Toxins 2023, 15, 499. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| RS 1 | Physically inaccessible or undigestible resistant starch |
| Food sources: Legumes Seeds Wholegrains | |
| RS 2 | Resistant starch is inaccessible to enzymes due to starch conformation |
| Food sources: Raw potato Unripe bananas High-Amylose Maize Starch (HAMS) | |
| RS 3 | Resistant starch is created through the process of retrogradation when starch becomes less soluble after being heated and dissolved in water and then cooled |
| Food sources: Heated and then cooled rice, pasta, potatoes, bread | |
| RS 4 | Resistant starch is created by the chemical modification of starch molecules and resists digestion |
| Food sources: Food in which the modified starches have been used (breads, cakes) | |
| RS 5 | Starches that are complexed with lipids |
| Food sources: Food with high amylose content (e.g., peas, maize, potatoes, wheat) |
| Study | Substance | Subjects and Duration of Supplementation | Duration of Study Results |
|---|---|---|---|
| Martinez et al., 2010 [59] | RS 2 or RS 4 about 30 g RS/day | 13 healthy adults 17 weeks Analysis of fecal microbiota by PCR-DGGE | RS2 and RS4 show functional differences in the effect on human microbiota RS2—Ruminococcus bromii and Eubacterium rectale RS4—Bifidobacterium adolescentis and Parabacteroides distasonis |
| Baxter et al., 2019 [60] | RPS about 31 g/day (70%RS2) RMS about 22 g/day (50%RS2) Inulin 20 g/day Starch 40 g/day | 2 weeks 172 healthy adults N = 39 N = 43 N = 41 N = 49 | RPS resulted in the greatest increase in total SCFA, including butyrate RMS and inulin induced distinct changes in fecal communities but did not generate significant increases in fecal butyrate levels |
| Venkataraman et al., 2016 [61] | RS (50%RS2) | 10 days 20 healthy young adults | increased abundance of the butyrogenic microorganism Eubacterium rectale |
| Alfa et al., 2018 [62] | RS2 | 12 weeks 42 elderly patient | increased numbers of Bifidobacterium |
| Walker et al., 2011 [63] | RS | 10 weeks 12 overweight men | increased in the number of Eubacterium rectale |
| Xin-Hua Jia et al., 2024 [65] | Dietary fiber intake | 2316 patients with DKD | increased dietary fiber intake is associated with a reduced incidence of DKD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kędzierska-Kapuza, K.; Grudniewska, A.; Durma, A.; Małecki, R.; Franek, E.; Szczuko, M. Dietary and Nutritional Strategies to Prevent Uremic Toxin Formation and Slow the Progression of Diabetic Kidney Disease. J. Clin. Med. 2025, 14, 4701. https://doi.org/10.3390/jcm14134701
Kędzierska-Kapuza K, Grudniewska A, Durma A, Małecki R, Franek E, Szczuko M. Dietary and Nutritional Strategies to Prevent Uremic Toxin Formation and Slow the Progression of Diabetic Kidney Disease. Journal of Clinical Medicine. 2025; 14(13):4701. https://doi.org/10.3390/jcm14134701
Chicago/Turabian StyleKędzierska-Kapuza, Karolina, Anna Grudniewska, Anna Durma, Robert Małecki, Edward Franek, and Małgorzata Szczuko. 2025. "Dietary and Nutritional Strategies to Prevent Uremic Toxin Formation and Slow the Progression of Diabetic Kidney Disease" Journal of Clinical Medicine 14, no. 13: 4701. https://doi.org/10.3390/jcm14134701
APA StyleKędzierska-Kapuza, K., Grudniewska, A., Durma, A., Małecki, R., Franek, E., & Szczuko, M. (2025). Dietary and Nutritional Strategies to Prevent Uremic Toxin Formation and Slow the Progression of Diabetic Kidney Disease. Journal of Clinical Medicine, 14(13), 4701. https://doi.org/10.3390/jcm14134701