
MicroRNA-224 Induces G1/S Checkpoint Release in Liver Cancer

Abstract
:1. Introduction
2. Experimental Section
2.1. Human Tissues
2.2. Quantitative Real-Time RT-PCR (qRT-PCR)
2.3. Cell Lines and Cell Culture
2.4. Transfection with miR-224 Mimic or Non-Specific Mimic
2.5. Retroviral Vectors, Viral Supernatant Production, and Viral Transduction
2.6. Cell Growth Assay
2.7. cDNA Microarrays and Filtering Genes
2.8. Cell Cycle Analysis by Flow Cytometry
2.9. Luciferase Reporter Assay
2.10. Western Blotting
2.11. Statistical Analysis
3. Results and Discussion
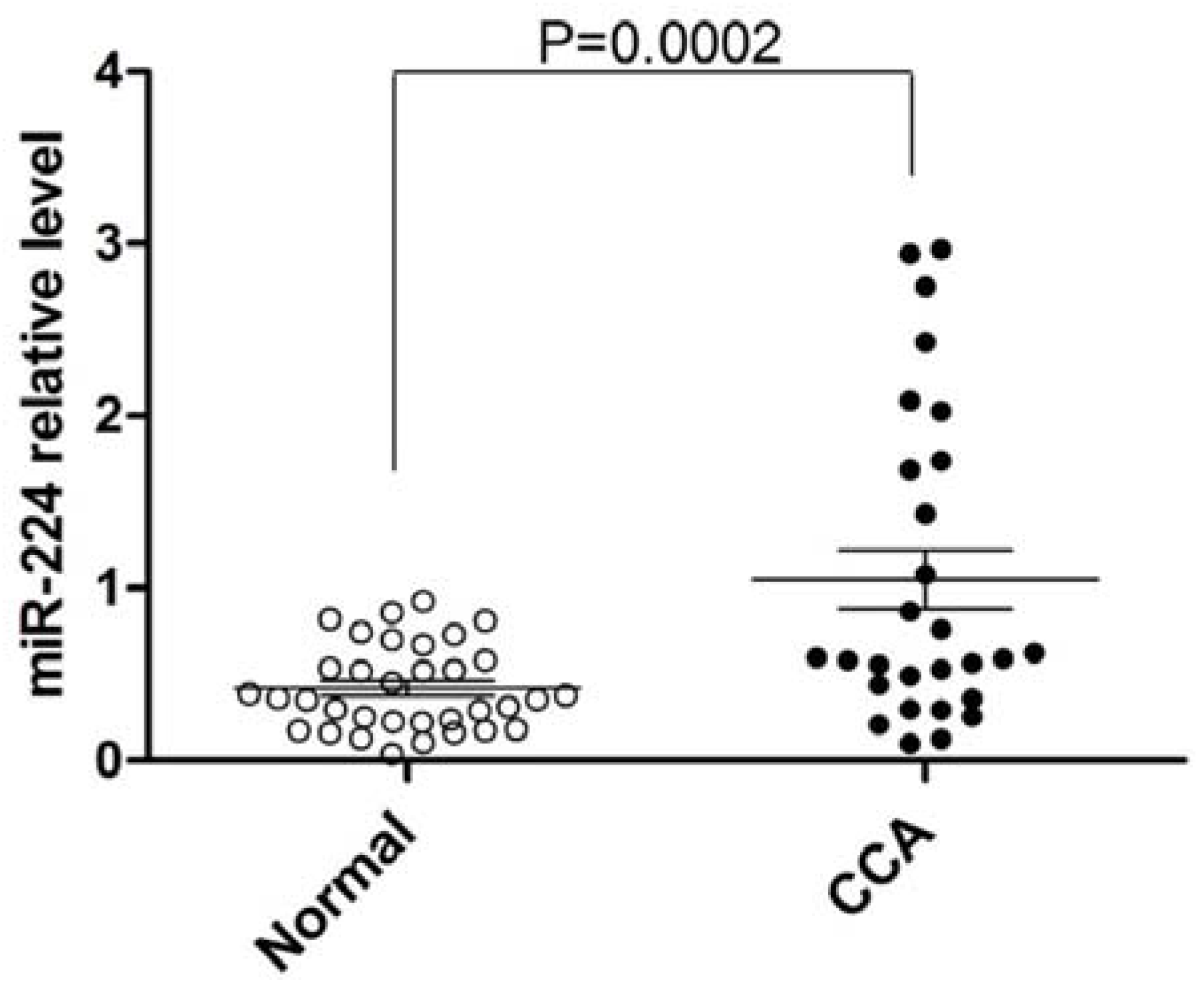
3.1. miR-224 is Upregulated in Human CCA vs. Normal Tissues

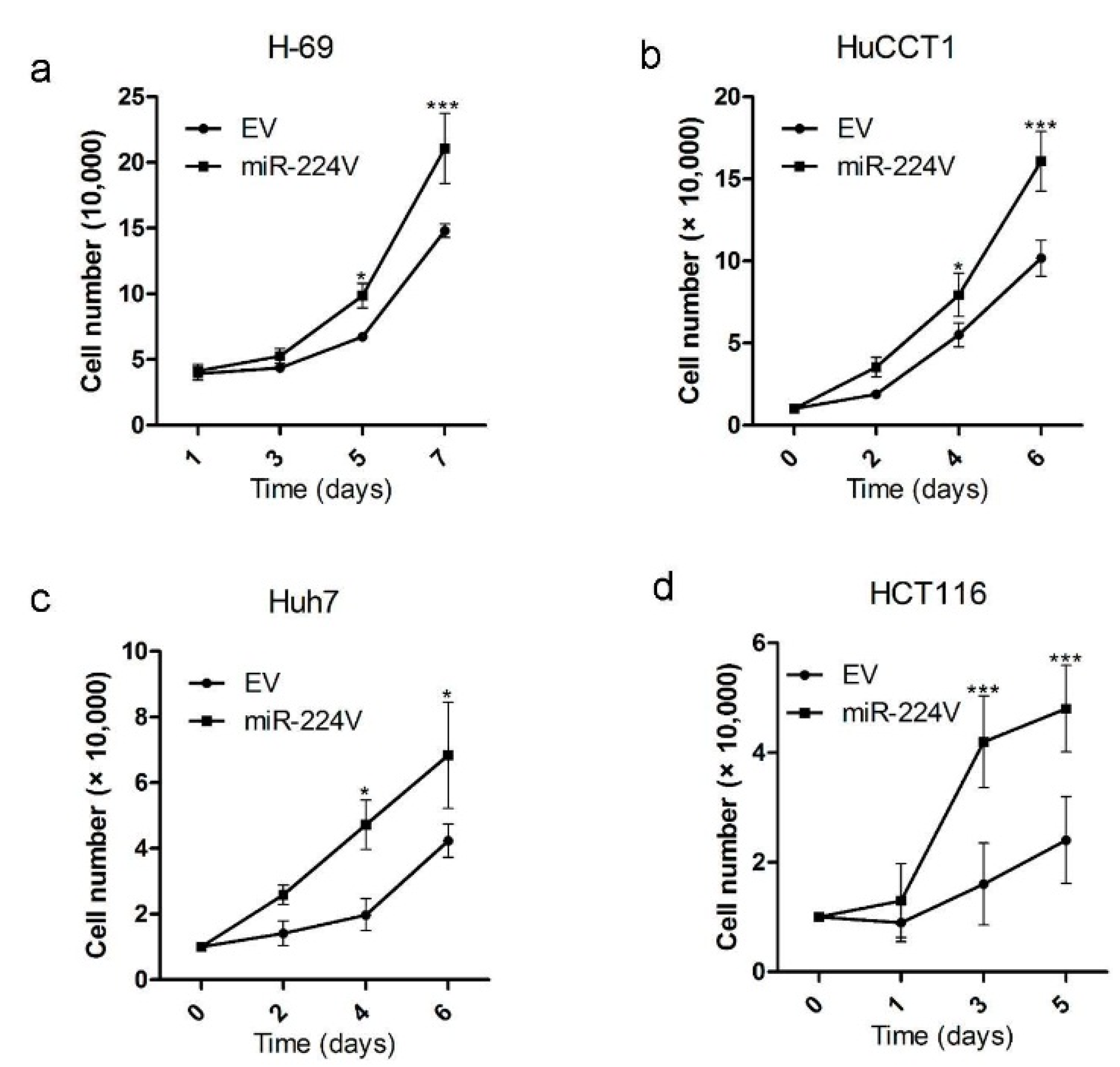
3.2. miR-224 Induces Cell Growth

3.3. miR-224 Functions and Signaling Pathways Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diseases and Disorders | ||
| Name | p-Value | Molecules Number |
| Cancer | 1.24E-05–1.35E--02 | 188 |
| Immunological Disease | 2.15E-05–1.01E--02 | 34 |
| Neurological Disease | 1.05E-04–1.33E--02 | 179 |
| Dermatological Diseases and Conditions | 1.11E-04–1.27E--02 | 76 |
| Genetic Disorder | 1.39E-45–1.27E--02 | 113 |
| Molecular and Cellular Functions | ||
| Name | p-Value | Molecules Number |
| Gene Expression | 5.20E-07–1.15E--02 | 137 |
| Cell Cycle | 8.78E-07–1.36E--02 | 90 |
| Cell Death | 2.71E-06–1.12E--02 | 129 |
| Cellular Growth and Proliferation | 9.88E-06–1.40E--02 | 162 |
| Small Molecule Biochemistry | 5.00E-05–1.18E--02 | 61 |
| Physiological System Development and Function | ||
| Name | p-Value | Molecules Number |
| Connective Tissue Development and Function | 1.28E-04–1.26E--02 | 58 |
| Embryonic Development | 2.22E-04–9.57E--03 | 33 |
| Hematological System Development and Function | 2.32E-04–1.38E--03 | 72 |
| Hematopoiesis | 2.32E-04–1.38E--03 | 48 |
| Top Canonical Pathways | ||
| Name | p-Value | Ratio |
| Regulation of Actin-based Motility by Rho | 7.22E-05 | 1/19(0.121) |
| Cell Cycle Regulation by BTG Family Proteins | 1.01E-04 | 7/36(0.197) |
| Pancreatic Adenocarcinoma Signaling | 5.76E-04 | 11/119(0.092) |
| Crosstalk between Dendritic Cells and Natural Killer Cells | 5.91E-04 | 10/96(0.104) |
| Type 1 Diabetes Mellitus Signaling | 7.29E-04 | 11/121(0.091) |
| Top Tox Lists | ||
| Name | p-Value | Ratio |
| Renal Necrosis/Cell Death | 1.04E-06 | 28/314(0.089) |
| Oxidative Stress | 2.17E-03 | 7/57(0.123) |
| Cell Cycle:G1/S Checkpoint Regulation | 9.47E-03 | 6/57(0.105) |
| Aryl Hydrocarbon Receptor Signaling | 1.22E-02 | 11/157(0.07) |
| VDR/RXR Activation | 1.23E-02 | 7/78(0.09) |
| Clinical Chemistry and Hematology | ||
| Name | p-Value | Molecules Number |
| Increased Levels of AST | 1.40E-02–1.40E--02 | 2 |
| Increased Levels of Albumin | 3.20E-02–3.20E--02 | 1 |
| Increased Levels of Alkaline Phosphatase | 6.29E-02–3.79E--02 | 3 |
| Increased Levels of Potassium | 6.29E-02–6.29E--02 | 1 |
| Decreased Levels of Albumin | 1.77E-01–1.77E--01 | 1 |
| Hepatotoxicity | ||
| Name | p-Value | Molecules Number |
| Liver Damage | 5.86E-03–1.41E--01 | 5 |
| Liver Necrosis/Cell Death | 1.15E-02–3.45E--01 | 12 |
| Liver Cholestasis | 2.51E-02–1.19E--01 | 7 |
| Biliary Hyperplasia | 3.20E-02–3.20E--01 | 1 |
| Liver Degradation | 3.20E-02–3.20E--02 | 1 |
3.4. miR-224 Induces G1/S Checkpoint Release in Normal and Cancer Cells

3.5. miR-224 Regulates Multiple Proteins Involved in G1/S Checkpoint


3.6. miR-224 Binds to p21 mRNA 3′UTR

4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Braconi, C.; Henry, J.C.; Kogure, T.; Schmittgen, T.; Patel, T. The role of microRNAs in human liver cancers. Semin. Oncol. 2011, 38, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Pineau, P.; Volinia, S.; McJunkin, K.; Marchio, A.; Battiston, C.; Terris, B.; Mazzaferro, V.; Lowe, S.W.; Croce, C.M.; Dejean, A. miR-221 overexpression contributes to liver tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Mott, J.L.; Kobayashi, S.; Bronk, S.F.; Gores, G.J. mir-29 regulates Mcl-1 protein expression and apoptosis. Oncogene 2007, 26, 6133–6140. [Google Scholar] [CrossRef] [PubMed]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.W.; Chang, T.C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R.; et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Razumilava, N.; Bronk, S.F.; Smoot, R.L.; Fingas, C.D.; Werneburg, N.W.; Roberts, L.R.; Mott, J.L. miR-25 targets TNF-related apoptosis inducing ligand (TRAIL) death receptor-4 and promotes apoptosis resistance in cholangiocarcinoma. Hepatology 2012, 55, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Braconi, C.; Huang, N.; Patel, T. MicroRNA-dependent regulation of DNA methyltransferase-1 and tumor suppressor gene expression by interleukin-6 in human malignant cholangiocytes. Hepatology 2010, 51, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Henson, R.; Lang, M.; Wehbe, H.; Maheshwari, S.; Mendell, J.T.; Jiang, J.; Schmittgen, T.D.; Patel, T. Involvement of human micro-RNA in growth and response to chemotherapy in human cholangiocarcinoma cell lines. Gastroenterology 2006, 130, 2113–2129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lee, A.T.; Ma, J.Z.; Wang, J.; Ren, J.; Yang, Y.; Tantoso, E.; Li, K.B.; Ooi, L.L.; Tan, P.; Lee, C.G. Profiling microRNA expression in hepatocellular carcinoma reveals microRNA-224 up-regulation and apoptosis inhibitor-5 as a microRNA-224-specific target. J. Biol. Chem. 2008, 283, 13205–13215. [Google Scholar] [CrossRef] [PubMed]
- Ladeiro, Y.; Couchy, G.; Balabaud, C.; Bioulac-Sage, P.; Pelletier, L.; Rebouissou, S.; Zucman-Rossi, J. MicroRNA profiling in hepatocellular tumors is associated with clinical features and oncogene/tumor suppressor gene mutations. Hepatology 2008, 47, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Wong, C.C.; Tung, E.K.; Lee, J.M.; Wong, C.M.; Ng, I.O. Deregulation of microRNA expression occurs early and accumulates in early stages of HBV-associated multistep hepatocarcinogenesis. J. Hepatol. 2011, 54, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Grubman, S.A.; Perrone, R.D.; Lee, D.W.; Murray, S.L.; Rogers, L.C.; Wolkoff, L.I.; Mulberg, A.E.; Cherington, V.; Jefferson, D.M. Regulation of intracellular pH by immortalized human intrahepatic biliary epithelial cell lines. Am. J. Physiol. 1994, 266, G1060–G1070. [Google Scholar] [PubMed]
- Cummins, J.M.; He, Y.; Leary, R.J.; Pagliarini, R.; Diaz, L.A., Jr.; Sjoblom, T.; Barad, O.; Bentwich, Z.; Szafranska, A.E.; Labourier, E.; et al. The colorectal microRNAome. Proc. Natl. Acad. Sci. USA 2006, 103, 3687–3692. [Google Scholar] [CrossRef] [PubMed]
- Ghiaur, G.; Lee, A.; Bailey, J.; Cancelas, J.A.; Zheng, Y.; Williams, D.A. Inhibition of RhoA GTPase activity enhances hematopoietic stem and progenitor cell proliferation and engraftment. Blood 2006, 108, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Wahlers, A.; Schwieger, M.; Li, Z.; Meier-Tackmann, D.; Lindemann, C.; Eckert, H.G.; von Laer, D.; Baum, C. Influence of multiplicity of infection and protein stability on retroviral vector-mediated gene expression in hematopoietic cells. Gene Ther. 2001, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linsley, P.S.; Schelter, J.; Burchard, J.; Kibukawa, M.; Martin, M.M.; Bartz, S.R.; Johnson, J.M.; Cummins, J.M.; Raymond, C.K.; Dai, H.; et al. Transcripts targeted by the microRNA-16 family cooperatively regulate cell cycle progression. Mol. Cell. Biol. 2007, 27, 2240–2252. [Google Scholar] [CrossRef] [PubMed]
- Patel, T. Cholangiocarcinoma—Controversies and challenges. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, G.; Shan, J.L.; Yang, Z.X.; Wang, H.Z.; Feng, J.; Zhen, J.J.; Chen, C.; Zhang, Z.M.; Xu, W.; et al. MicroRNA-224 is upregulated in HepG2 cells and involved in cellular migration and invasion. J. Gastroenterol. Hepatol. 2010, 25, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lee, C.G. Role of miR-224 in hepatocellular carcinoma: A tool for possible therapeutic intervention? Epigenomics 2011, 3, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ding, C.; Chen, C.; Zhang, Z.; Xiao, H.; Xie, F.; Lei, L.; Chen, Y.; Mao, B.; Jiang, M.; et al. miR-224 promotion of cell migration and invasion by targeting Homeobox D 10 gene in human hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2014, 29, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Maudsley, S.; Chadwick, W.; Wang, L.; Zhou, Y.; Martin, B.; Park, S.S. Bioinformatic approaches to metabolic pathways analysis. Methods Mol. Biol. 2011, 756, 99–130. [Google Scholar] [PubMed]
- Gusev, Y. Computational methods for analysis of cellular functions and pathways collectively targeted by differentially expressed microRNA. Methods 2008, 44, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhu, L.J.; Yang, Y.C.; Wang, Z.X.; Wang, R. MiR-224 promotes the chemoresistance of human lung adenocarcinoma cells to cisplatin via regulating G(1)/S transition and apoptosis by targeting p21(WAF1/CIP1). Br. J. Cancer 2014, 111, 339–354. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, F.; Olaru, A.V.; Mezey, E.; Xie, Q.; Li, L.; Piontek, K.B.; Selaru, F.M. MicroRNA-224 Induces G1/S Checkpoint Release in Liver Cancer. J. Clin. Med. 2015, 4, 1713-1728. https://doi.org/10.3390/jcm4091713
An F, Olaru AV, Mezey E, Xie Q, Li L, Piontek KB, Selaru FM. MicroRNA-224 Induces G1/S Checkpoint Release in Liver Cancer. Journal of Clinical Medicine. 2015; 4(9):1713-1728. https://doi.org/10.3390/jcm4091713
Chicago/Turabian StyleAn, Fangmei, Alexandru V. Olaru, Esteban Mezey, Qing Xie, Ling Li, Klaus B. Piontek, and Florin M. Selaru. 2015. "MicroRNA-224 Induces G1/S Checkpoint Release in Liver Cancer" Journal of Clinical Medicine 4, no. 9: 1713-1728. https://doi.org/10.3390/jcm4091713
APA StyleAn, F., Olaru, A. V., Mezey, E., Xie, Q., Li, L., Piontek, K. B., & Selaru, F. M. (2015). MicroRNA-224 Induces G1/S Checkpoint Release in Liver Cancer. Journal of Clinical Medicine, 4(9), 1713-1728. https://doi.org/10.3390/jcm4091713