Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Characteristics
2.2. DNA Extraction
2.3. Detection of HPV by Polymerase Chain Reaction (PCR) Amplification
2.4. Real-time PCR Amplification of HPV Type-Specific DNA
2.5. RNA Extraction
2.6. Real-Time PCR for MIF mRNA Quantification
2.7. Immunohistochemistry of p16
2.8. Immunohistochemistry for MIF Protein Detection
2.9. Computer-Assisted Morphometry
2.10. LC MS/MS Analysis and Data Processing
2.11. Cell Culture
2.12. MIF Inhibitor
2.13. Determination of MIF Concentrations
2.14. IC50 Determination for the MIF Inhibitor by a Crystal Violet Stain Assay
2.15. Western Blotting for the Evaluation of HIF-1α and MIF Expression
2.16. Animal Study
2.17. Cell Sample Preparation for NMR Analysis
2.18. H-NMR Sample Preparation and Spectroscopy
2.19. Statistical Analyses
3. Results
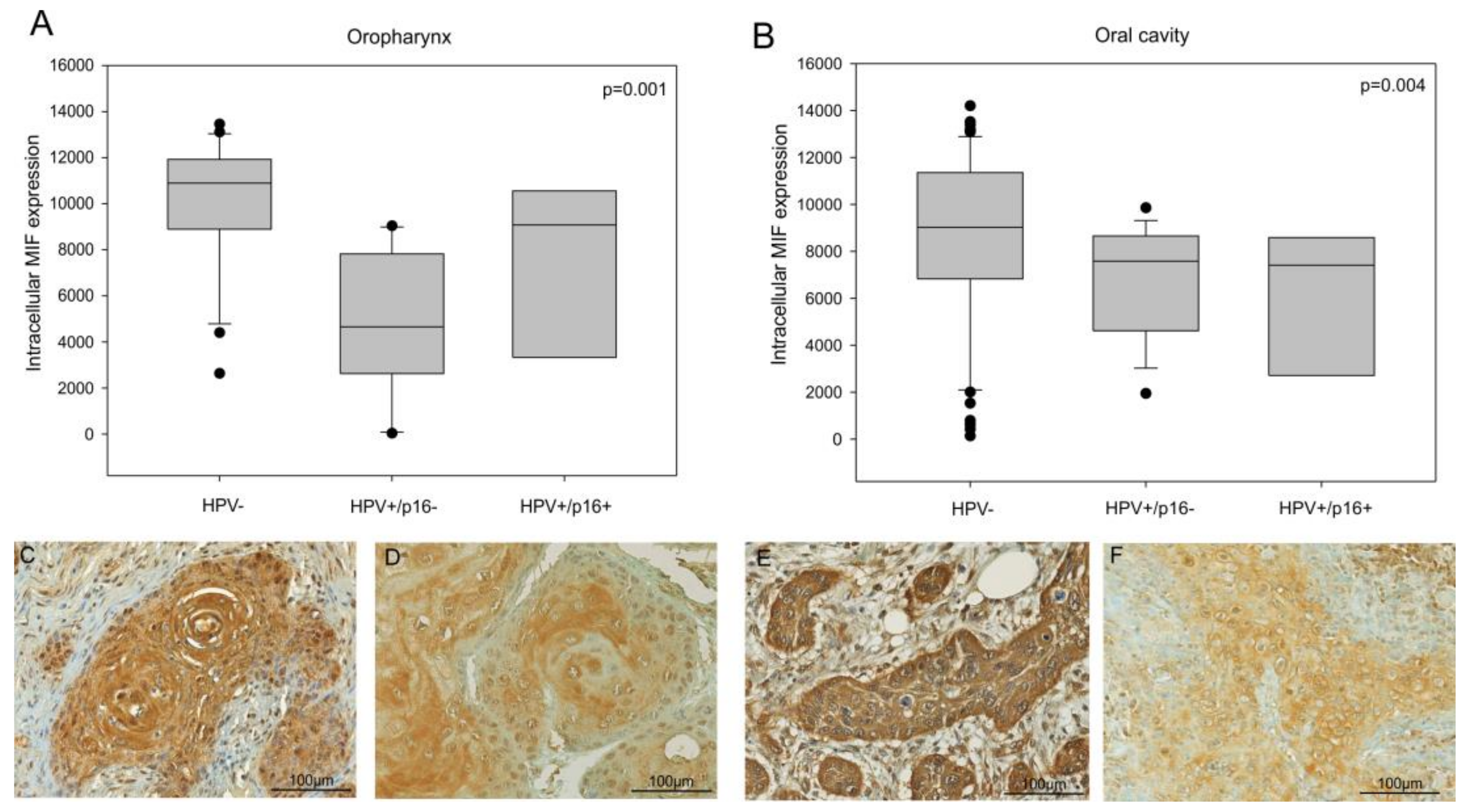
3.1. Tissue MIF Expression is Decreased in the Oral Cavity and in Oropharyngeal Carcinomas Infected with HPV
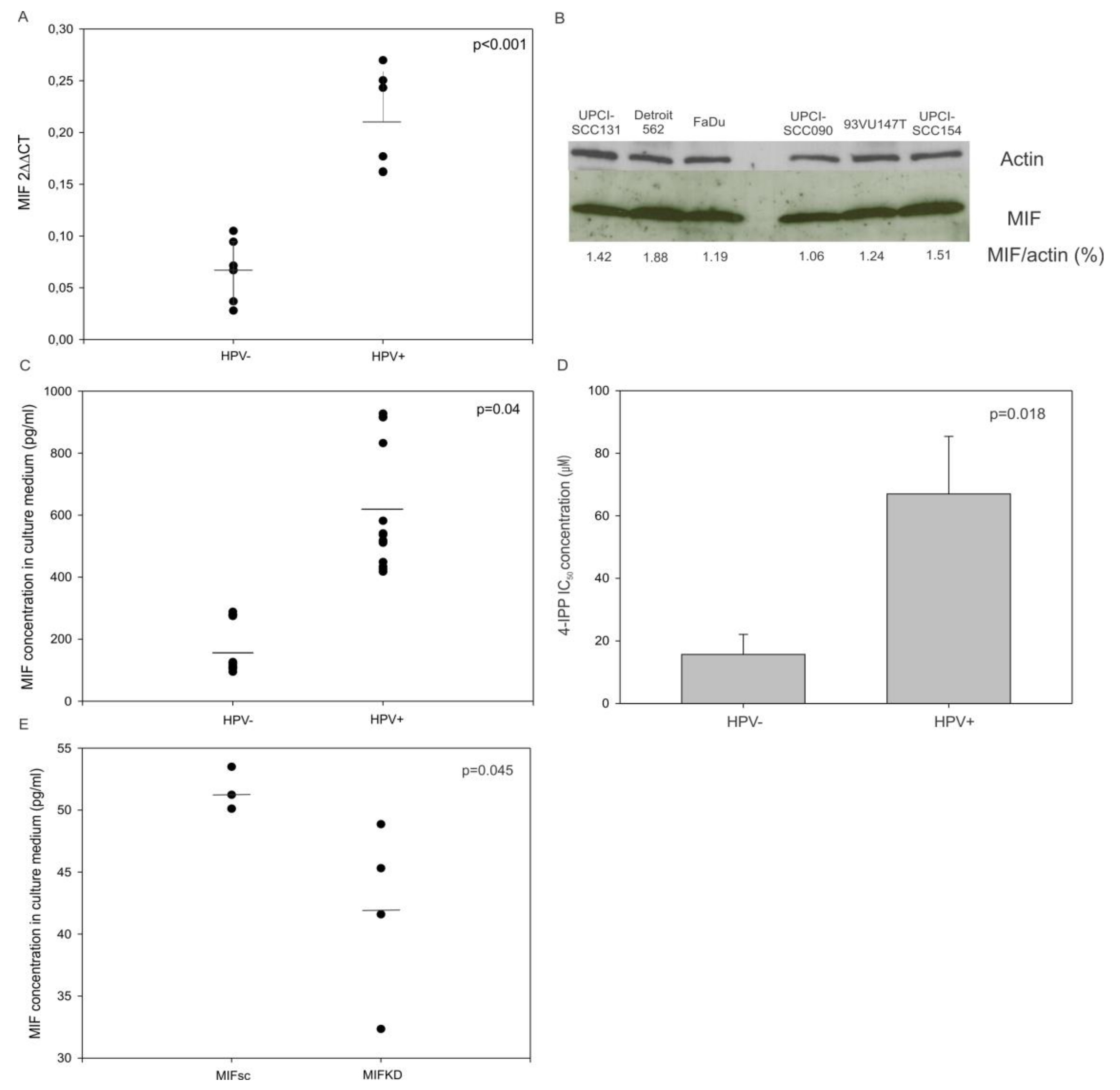
3.2. MIF mRNA Synthesis and Protein Secretion is Increased in HPV-Infected Human Head and Neck Cancer Cell Lines
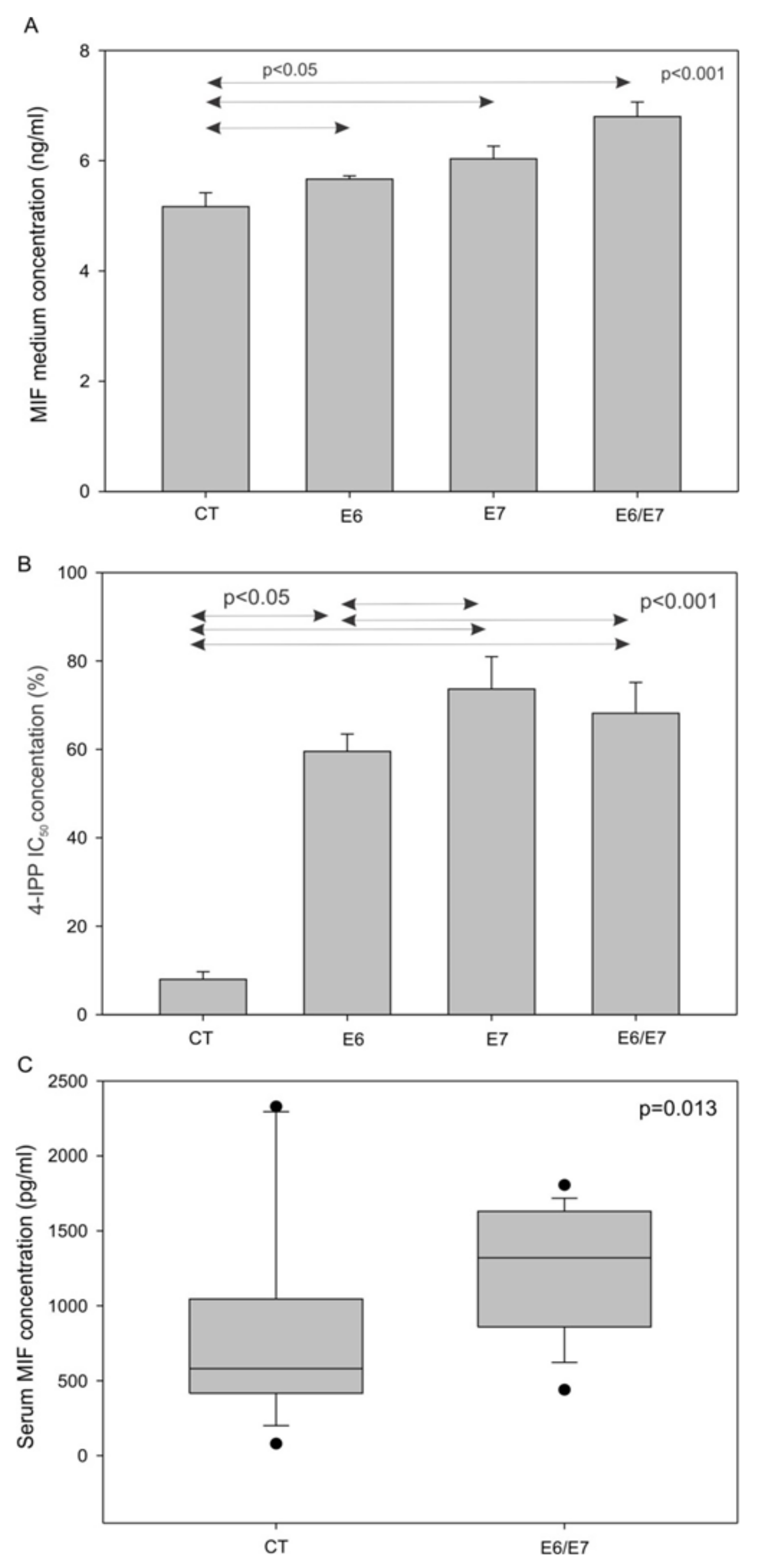
3.3. MIF Secretion is Increased in Murine Head and Neck Cancer Cell Lines Transfected with E6/E7 Oncoproteins In Vitro and In Vivo
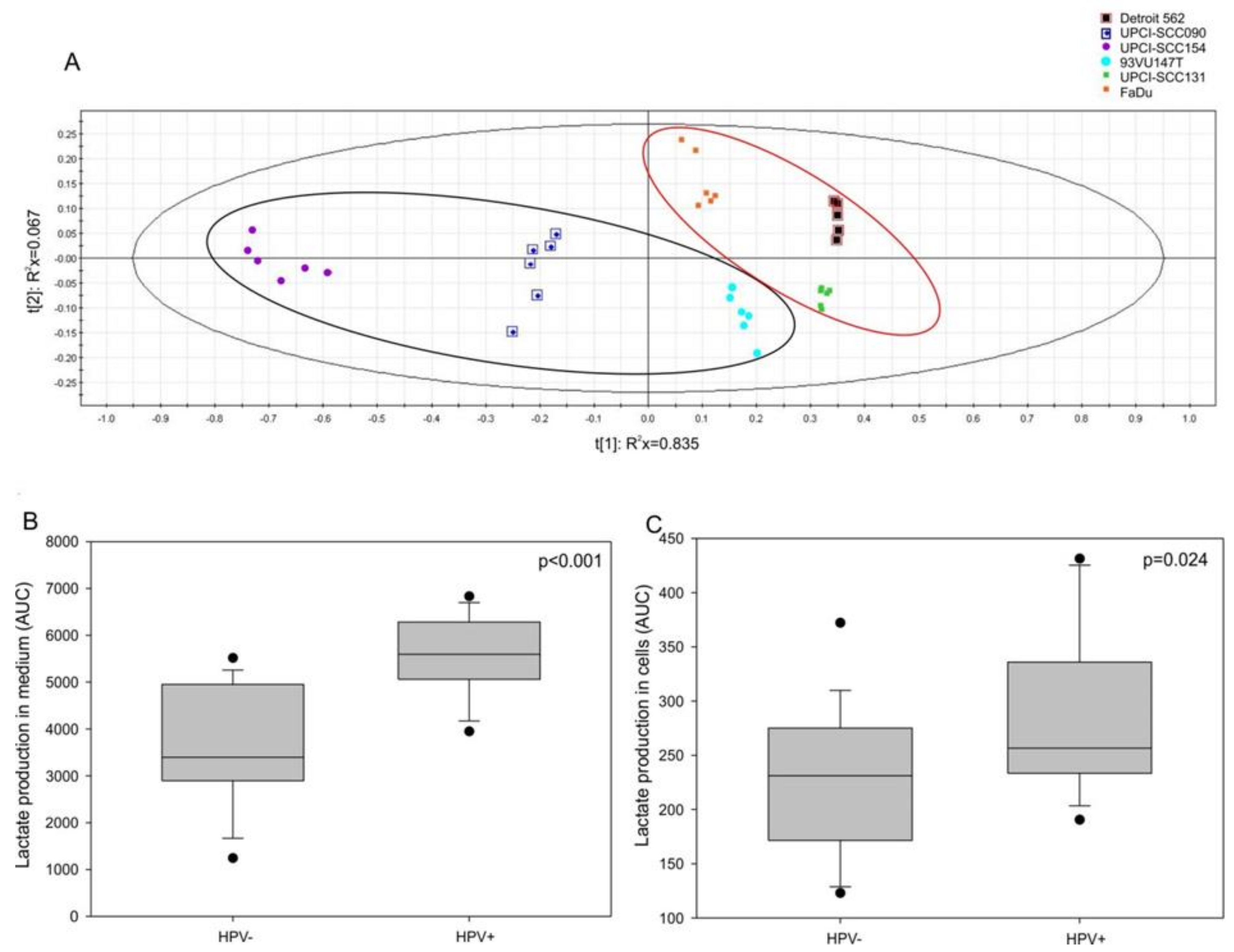
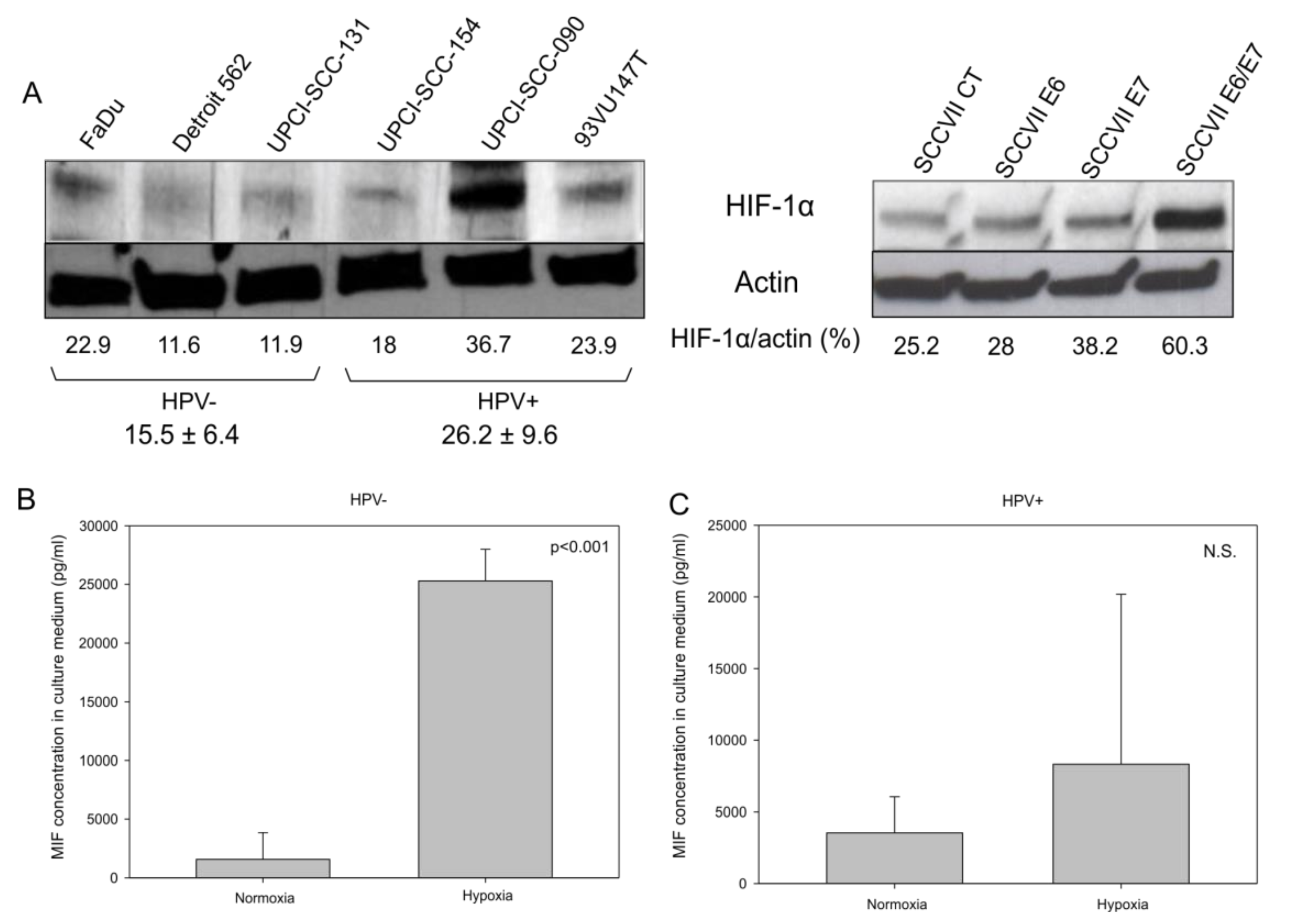
3.4. Lactate Production and HIF-1α Expression are Increased in HPV-Positive Cell Lines
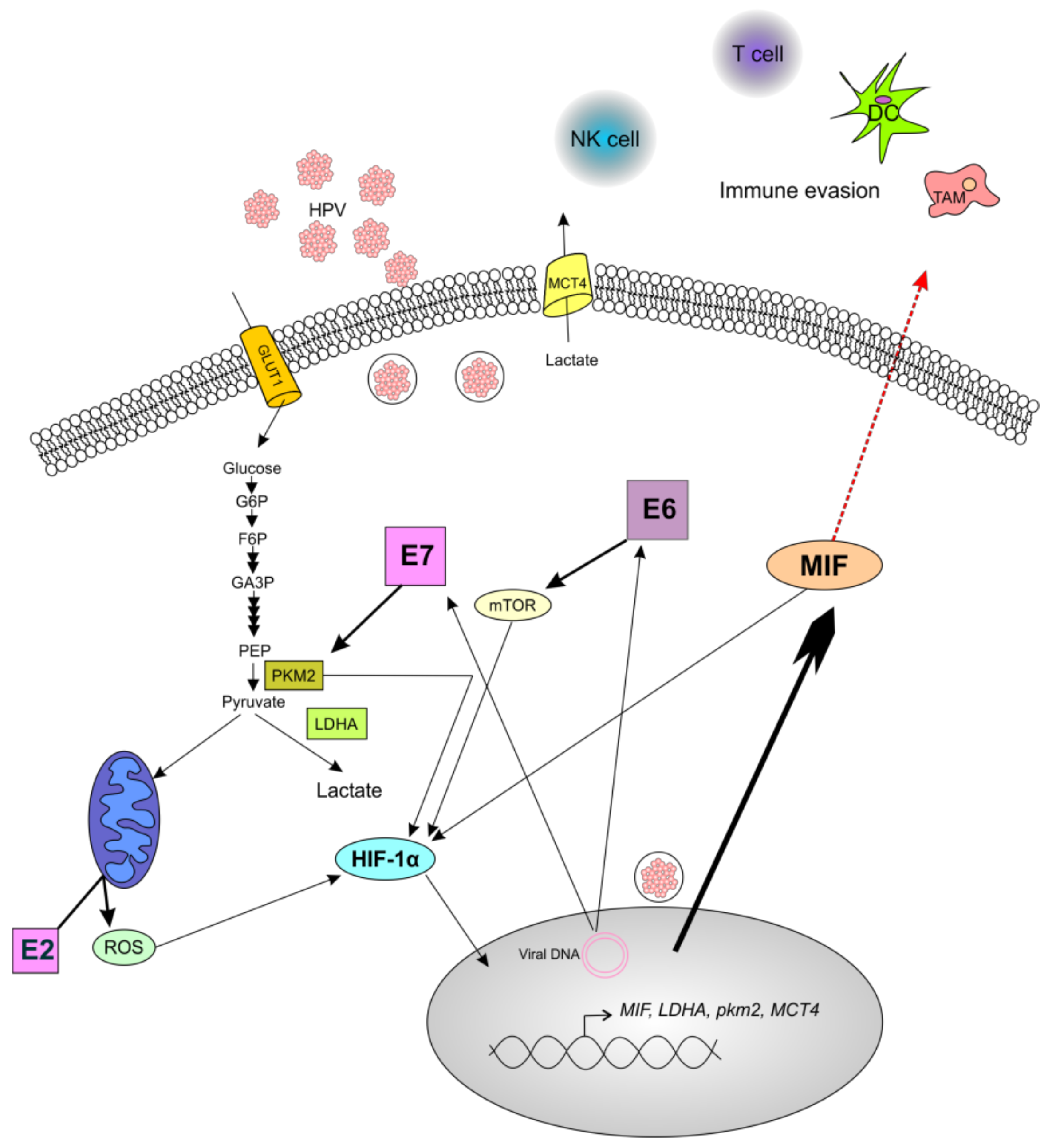
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments:
Conflicts of Interest
References
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2017, 3, 524–548. [Google Scholar]
- Elrefaey, S.; Massaro, M.A.; Chiocca, S.; Chiesa, F.; Ansarin, M. HPV in oropharyngeal cancer: The basics to know in clinical practice. Acta Otorhinolaryngol. Ital. 2014, 34, 299–309. [Google Scholar] [PubMed]
- Saraiya, M.; Unger, E.R.; Thompson, T.D.; Lynch, C.F.; Hernandez, B.Y.; Lyu, C.W.; Steinau, M.; Watson, M.; Wilkinson, E.J.; Hopenhayn, C.; et al. US assessment of HPV types in cancers: Implications for current and 9-valent HPV vaccines. J. Natl. Cancer Inst. 2015, 107, djv086. [Google Scholar] [CrossRef]
- Chera, B.S.; Kumar, S.; Shen, C.; Amdur, R.J.; Dagan, R.; Weiss, J.; Grilley-Olson, J.; Zanation, A.; Hackman, T.; Blumberg, J.; et al. Plasma Circulating Tumor HPV DNA for the Surveillance of Cancer Recurrence in HPV-associated Oropharyngeal Cancer. Int. J. Radiat. Oncol. 2018, 102, 1605–1606. [Google Scholar] [CrossRef]
- Gupta, G.P.; Kumar, S.; Marron, D.; Amdur, R.J.; Hayes, D.N.; Weiss, J.; Grilley-Olson, J.; Zanation, A.; Hackman, T.; Zevallos, J.P.; et al. Circulating Tumor HPV16 DNA as a Biomarker of Tumor Genomics and Disease Control in HPV-associated Oropharyngeal Squamous Cell Carcinoma. Int. J. Radiat. Oncol. 2018, 100, 1310–1311. [Google Scholar] [CrossRef]
- Anantharaman, D.; Abedi-Ardekani, B.; Beachler, D.C.; Gheit, T.; Olshan, A.F.; Wisniewski, K.; Wunsch-Filho, V.; Toporcov, T.N.; Tajara, E.H.; Levi, J.E.; et al. Geographic heterogeneity in the prevalence of human papillomavirus in head and neck cancer. Int. J. Cancer 2017, 140, 1968–1975. [Google Scholar] [CrossRef] [PubMed]
- Näsman, A.; Attner, P.; Hammarstedt, L.; Du, J.; Eriksson, M.; Giraud, G.; Ahrlund-Richter, S.; Marklund, L.; Romanitan, M.; Lindquist, D.; et al. Incidence of human papillomavirus (HPV) positive tonsillar carcinoma in Stockholm, Sweden: An epidemic of viral-induced carcinoma? Int. J. Cancer 2009, 125, 362–366. [Google Scholar] [CrossRef] [Green Version]
- Vlantis, A.C. Human Papilloma Virus and Oropharyngeal Carcinoma-Lessons from History. Chin. J. Dent. Res. Off. J. Sci. Sect. Chin. Stomatol. Assoc. CSA 2016, 19, 9–16. [Google Scholar]
- Descamps, G.; Karaca, Y.; Lechien, J.R.; Kindt, N.; Decaestecker, C.; Remmelink, M.; Larsimont, D.; Andry, G.; Hassid, S.; Rodriguez, A.; et al. Classical risk factors, but not HPV status, predict survival after chemoradiotherapy in advanced head and neck cancer patients. J. Cancer Res. Clin. Oncol. 2016, 142, 2185–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filleul, O.; Preillon, J.; Crompot, E.; Lechien, J.; Saussez, S. Incidence of head and neck cancers in Belgium: Comparison with world wide and French data. Bull. Cancer (Paris) 2011, 98, 1173–1183. [Google Scholar]
- Bloom, B.R.; Bennett, B. Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science 1966, 153, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Kindt, N.; Journe, F.; Laurent, G.; Saussez, S. Involvement of macrophage migration inhibitory factor in cancer and novel therapeutic targets. Oncol. Lett. 2016, 12, 2247–2253. [Google Scholar] [CrossRef] [Green Version]
- Kindt, N.; Preillon, J.; Kaltner, H.; Gabius, H.-J.; Chevalier, D.; Rodriguez, A.; Johnson, B.D.; Megalizzi, V.; Decaestecker, C.; Laurent, G.; et al. Macrophage migration inhibitory factor in head and neck squamous cell carcinoma: Clinical and experimental studies. J. Cancer Res. Clin. Oncol. 2013, 139, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Duray, A.; Descamps, G.; Decaestecker, C.; Sirtaine, N.; Gilles, A.; Khalifé, M.; Chantrain, G.; Depuydt, C.E.; Delvenne, P.; Saussez, S. Human papillomavirus predicts the outcome following concomitant chemoradiotherapy in patients with head and neck squamous cell carcinomas. Oncol. Rep. 2013, 30, 371–376. [Google Scholar] [CrossRef] [PubMed]
- PrimerBank. Available online: https://pga.mgh.harvard.edu/primerbank/ (accessed on 4 October 2018).
- Sawicka, M.; Pawlikowski, J.; Wilson, S.; Ferdinando, D.; Wu, H.; Adams, P.D.; Gunn, D.A.; Parish, W. The specificity and patterns of staining in human cells and tissues of p16INK4a antibodies demonstrate variant antigen binding. PLoS ONE 2013, 8, e53313. [Google Scholar] [CrossRef] [PubMed]
- Cludts, S.; Decaestecker, C.; Johnson, B.; Lechien, J.; Leroy, X.; Kindt, N.; Kaltner, H.; André, S.; Gabius, H.-J.; Saussez, S. Increased expression of macrophage migration inhibitory factor during progression to hypopharyngeal squamous cell carcinoma. Anticancer Res. 2010, 30, 3313–3319. [Google Scholar] [PubMed]
- Kindt, N.; Descamps, G.; Seminerio, I.; Bellier, J.; Lechien, J.R.; Mat, Q.; Pottier, C.; Delvenne, P.; Journé, F.; Saussez, S. High stromal Foxp3-positive T cell number combined to tumor stage improved prognosis in head and neck squamous cell carcinoma. Oral Oncol. 2017, 67, 183–191. [Google Scholar] [CrossRef]
- Zhou, Q.; Yan, X.; Gershan, J.; Orentas, R.J.; Johnson, B.D. Expression of macrophage migration inhibitory factor by neuroblastoma leads to the inhibition of antitumor T cell reactivity in vivo. J. Immunol. 2008, 181, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Hou, W.-J.; Zhao, Y.-J.; Liu, S.-L.; Qiu, X.-S.; Wang, E.-H.; Wu, G.-P. Overexpression of HPV16 E6/E7 mediated HIF-1α upregulation of GLUT1 expression in lung cancer cells. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 4655–4663. [Google Scholar] [CrossRef] [PubMed]
- Coppock, J.D.; Lee, J.H. mTOR, metabolism, and the immune response in HPV-positive head and neck squamous cell cancer. World J. Otorhinolaryngol. Head Neck Surg. 2016, 2, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Kindt, N.; Lechien, J.; Decaestecker, C.; Rodriguez, A.; Chantrain, G.; Remmelink, M.; Laurent, G.; Gabius, H.-J.; Saussez, S. Expression of macrophage migration-inhibitory factor is correlated with progression in oral cavity carcinomas. Anticancer Res. 2012, 32, 4499–4505. [Google Scholar] [PubMed]
- Howard, J.D.; Chung, C.H. Biology of Human Papillomavirus–Related Oropharyngeal Cancer. Semin. Radiat. Oncol. 2012, 22, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zhang, Q.; Nishitani, J.; Brown, J.; Shi, S.; Le, A.D. Overexpression of human papillomavirus type 16 oncoproteins enhances hypoxia-inducible factor 1 alpha protein accumulation and vascular endothelial growth factor expression in human cervical carcinoma cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 2568–2576. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Meng, X.; Ma, J.; Zheng, Y.; Wang, Q.; Wang, Y.; Shang, H. Human papillomavirus 16 E6 contributes HIF-1α induced Warburg effect by attenuating the VHL-HIF-1α interaction. Int. J. Mol. Sci. 2014, 15, 7974–7986. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Ojo, D.; Yan, J.; Tang, D. PKM2 contributes to cancer metabolism. Cancer Lett. 2015, 356, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Tan, C.L.; Gunaratne, J.; Quek, L.S.; Nei, W.; Thierry, F.; Bellanger, S. Localization of HPV-18 E2 at mitochondrial membranes induces ROS release and modulates host cell metabolism. PLoS ONE 2013, 8, e75625. [Google Scholar] [CrossRef]
- Pinheiro, C.; Garcia, E.A.; Morais-Santos, F.; Scapulatempo-Neto, C.; Mafra, A.; Steenbergen, R.D.M.; Boccardo, E.; Villa, L.L.; Baltazar, F.; Longatto-Filho, A. Lactate transporters and vascular factors in HPV-induced squamous cell carcinoma of the uterine cervix. BMC Cancer 2014, 14, 751. [Google Scholar] [CrossRef]
- Gupta, Y.; Pasupuleti, V.; Du, W.; Welford, S.M. Macrophage Migration Inhibitory Factor Secretion Is Induced by Ionizing Radiation and Oxidative Stress in Cancer Cells. PLoS ONE 2016, 11, e0146482. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Luo, F.; Yang, L.; Wu, W.; Liu, X. Hypoxia stimulates the expression of macrophage migration inhibitory factor in human vascular smooth muscle cells via HIF-1alpha dependent pathway. BMC Cell Biol. 2010, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.; Oda, T.; Nishi, K.; Takabuchi, S.; Wakamatsu, T.; Tanaka, T.; Adachi, T.; Fukuda, K.; Semenza, G.L.; Hirota, K. Macrophage migration inhibitory factor activates hypoxia-inducible factor in a p53-dependent manner. PLoS ONE 2008, 3, e2215. [Google Scholar] [CrossRef]
- No, Y.R.; Lee, S.-J.; Kumar, A.; Yun, C.C. HIF1α-Induced by Lysophosphatidic Acid Is Stabilized via Interaction with MIF and CSN5. PLoS ONE 2015, 10, e0137513. [Google Scholar] [CrossRef] [PubMed]
- Lechner, A.; Schlößer, H.; Rothschild, S.I.; Thelen, M.; Reuter, S.; Zentis, P.; Shimabukuro-Vornhagen, A.; Theurich, S.; Wennhold, K.; Garcia-Marquez, M.; et al. Characterization of tumor-associated T-lymphocyte subsets and immune checkpoint molecules in head and neck squamous cell carcinoma. Oncotarget 2017, 8, 44418–44433. [Google Scholar] [CrossRef] [PubMed]
- Kindt, N.; Descamps, G.; Seminerio, I.; Bellier, J.; Lechien, J.R.; Pottier, C.; Larsimont, D.; Journé, F.; Delvenne, P.; Saussez, S. Langerhans cell number is a strong and independent prognostic factor for head and neck squamous cell carcinomas. Oral Oncol. 2016, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.; Bellile, E.; Thomas, D.; McHugh, J.; Rozek, L.; Virani, S.; Peterson, L.; Carey, T.E.; Walline, H.; Moyer, J.; et al. Tumor infiltrating lymphocytes and survival in patients with head and neck squamous cell carcinoma. Head Neck 2016, 38, 1074–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Karp, J.E.; Emadi, A. Elevated lactate dehydrogenase (LDH) can be a marker of immune suppression in cancer: Interplay between hematologic and solid neoplastic clones and their microenvironments. Cancer Biomark. Sect. Dis. Markers 2017, 19, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.; Smith, J.; Rocher-Ros, V.; Nadkarni, S.; Montero-Melendez, T.; D’Acquisto, F.; Bland, E.J.; Bombardieri, M.; Pitzalis, C.; Perretti, M.; et al. Lactate Regulates Metabolic and Pro-inflammatory Circuits in Control of T Cell Migration and Effector Functions. PLoS Biol. 2015, 13, e1002202. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.A.; Gholaman, H.; Trellakis, S.; Bruderek, K.; Dominas, N.; Gu, X.; Bankfalvi, A.; Whiteside, T.L.; Lang, S.; Brandau, S. Tumor-derived macrophage migration inhibitory factor modulates the biology of head and neck cancer cells via neutrophil activation. Int. J. Cancer 2011, 129, 859–869. [Google Scholar] [CrossRef] [Green Version]
- Krockenberger, M.; Dombrowski, Y.; Weidler, C.; Ossadnik, M.; Hönig, A.; Häusler, S.; Voigt, H.; Becker, J.C.; Leng, L.; Steinle, A.; et al. Macrophage migration inhibitory factor contributes to the immune escape of ovarian cancer by down-regulating NKG2D. J. Immunol. 2008, 180, 7338–7348. [Google Scholar] [CrossRef]
- Ullah, M.S.; Davies, A.J.; Halestrap, A.P. The plasma membrane lactate transporter MCT4, but not MCT1, is up-regulated by hypoxia through a HIF-1alpha-dependent mechanism. J. Biol. Chem. 2006, 281, 9030–9037. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Cases | |
|---|---|
| Age (years) | |
| Median | 57 |
| Range | 44–90 |
| Gender | |
| Male | 31 |
| Female | 8 |
| Localization | |
| Tonsils | 13 |
| Soft palate | 11 |
| Base of tongue | 6 |
| Posterior wall | 3 |
| Unknown | 6 |
| Histological grade | |
| Well differentiated | 14 |
| Moderately differentiated | 12 |
| Poorly differentiated | 6 |
| Unknown | 7 |
| TNM stage | |
| T1-2 | 19 |
| T3-4 | 10 |
| Unknown | 10 |
| Risk factors | |
| Alcohol | |
| Non-drinker | 3 |
| Drinker | 24 |
| Unknown | 12 |
| Tobacco | |
| Non-smoker | 2 |
| Smoker | 25 |
| Unknown | 12 |
| HPV status | |
| HPV−ve | 21 |
| HPV+ve/p16+ | 3 |
| HPV+ve/p16− | 10 |
| HPV+ve/p16 unknown | 5 |
| Recurrence | |
| Yes | 15 |
| None | 15 |
| Unknown | 9 |
| Clinical follow-up (months) | |
| Median | 33 |
| Range | 1–121 |
| Number of Cases | |
|---|---|
| Age (years) | |
| Median | 58 |
| Range | 23–87 |
| Gender | |
| Male | 97 |
| Female | 20 |
| Localization | |
| Tongue | 27 |
| Floor of the mouth | 35 |
| Gingivae | 12 |
| Buccal mucosa | 7 |
| Retromolar trigone | 3 |
| Lips | 2 |
| Hard palate | 1 |
| Unknown | 30 |
| Histological grade | |
| Well differentiated | 44 |
| Moderately differentiated | 53 |
| Poorly differentiated | 19 |
| Unknown | 1 |
| TNM stage | |
| T1-2 | 79 |
| T3-4 | 31 |
| Unknown | 7 |
| Risk factors | |
| Alcohol | |
| Non-drinker | 47 |
| Drinker | 49 |
| Unknown | 21 |
| Tobacco | |
| Non-smoker | 40 |
| Smoker | 57 |
| Unknown | 20 |
| HPV status | |
| HPV−ve | 65 |
| HPV+ve/p16+ | 6 |
| HPV+ve/p16- | 42 |
| HPV+ve/p16 unknown | 4 |
| Recurrence | |
| Yes | 44 |
| None | 59 |
| Unknown | 14 |
| Clinical follow-up (months) | |
| Median | 32 |
| Range | 1–188 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kindt, N.; Descamps, G.; Lechien, J.R.; Remmelink, M.; Colet, J.-M.; Wattiez, R.; Berchem, G.; Journe, F.; Saussez, S. Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma. J. Clin. Med. 2019, 8, 75. https://doi.org/10.3390/jcm8010075
Kindt N, Descamps G, Lechien JR, Remmelink M, Colet J-M, Wattiez R, Berchem G, Journe F, Saussez S. Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma. Journal of Clinical Medicine. 2019; 8(1):75. https://doi.org/10.3390/jcm8010075
Chicago/Turabian StyleKindt, Nadège, Géraldine Descamps, Jérôme R. Lechien, Myriam Remmelink, Jean-Marie Colet, Ruddy Wattiez, Guy Berchem, Fabrice Journe, and Sven Saussez. 2019. "Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma" Journal of Clinical Medicine 8, no. 1: 75. https://doi.org/10.3390/jcm8010075
APA StyleKindt, N., Descamps, G., Lechien, J. R., Remmelink, M., Colet, J.-M., Wattiez, R., Berchem, G., Journe, F., & Saussez, S. (2019). Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma. Journal of Clinical Medicine, 8(1), 75. https://doi.org/10.3390/jcm8010075