Effects of Oral Anticoagulant Therapy on Gene Expression in Crosstalk between Osteogenic Progenitor Cells and Endothelial Cells

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Experimental Section
2.1. Subjects
2.2. Biochemical Parameters
2.3. Isolation of Circulating Progenitor Cells
2.4. RNA Extraction and Reverse Transcription
2.5. Real Time PCR (RT-PCR)
2.6. Cell Cultures and Co-Cultures Preparation
2.7. Immunofluorescence
2.8. Alizarin Staining
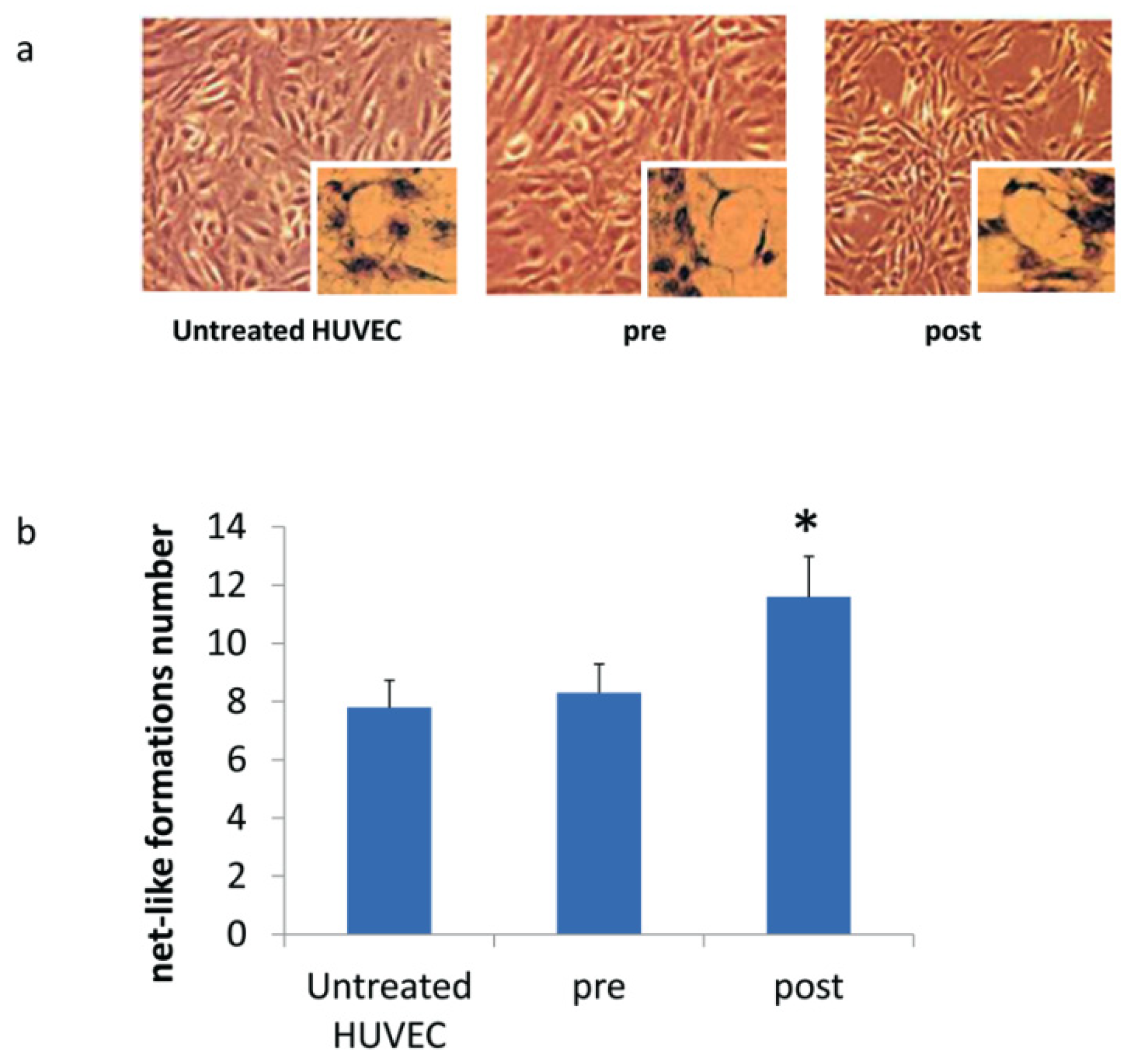
2.9. Net-Like Formation Assay
2.10. Statistical Analysis
3. Results
3.1. Clinical and Biochemical Data
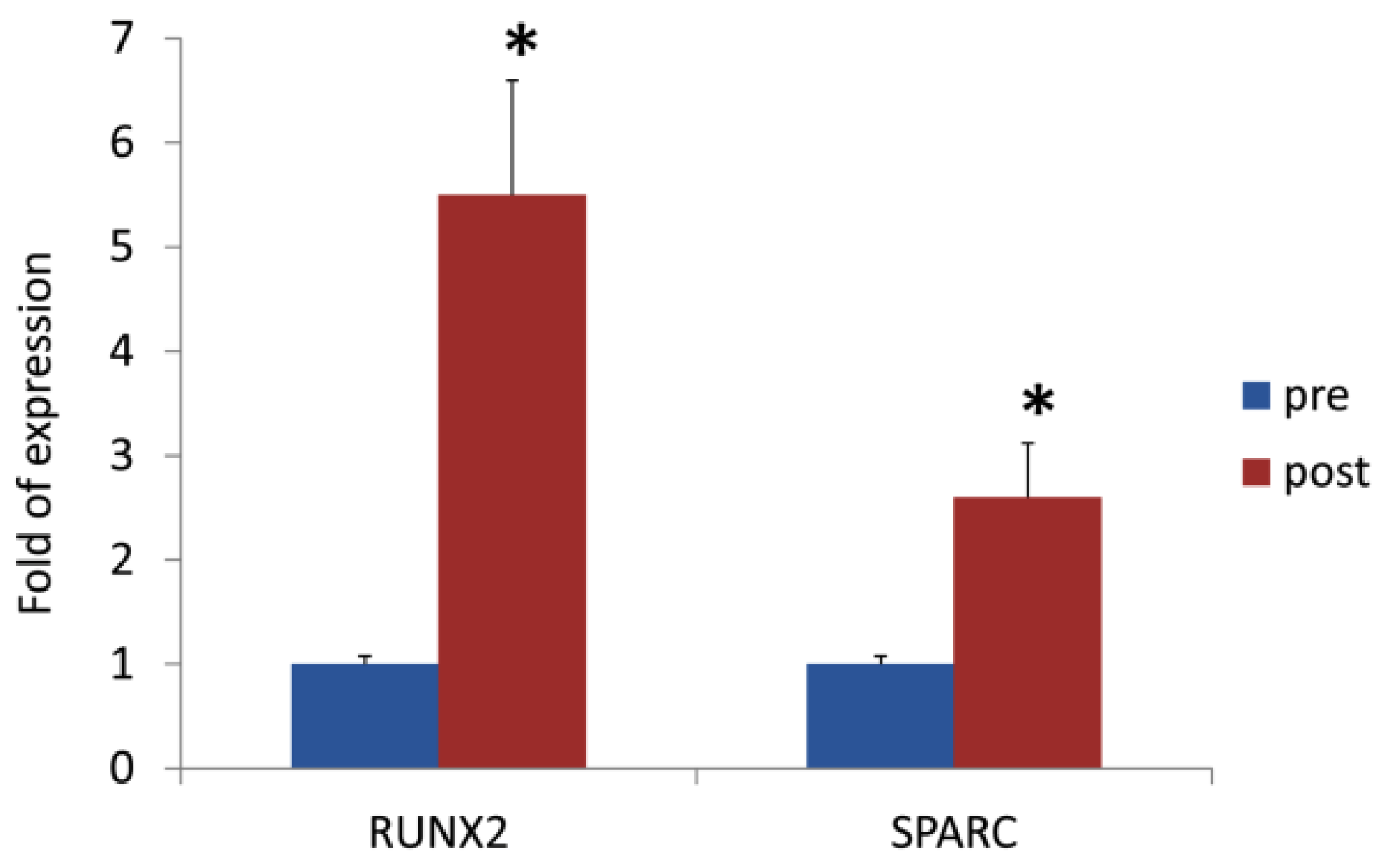
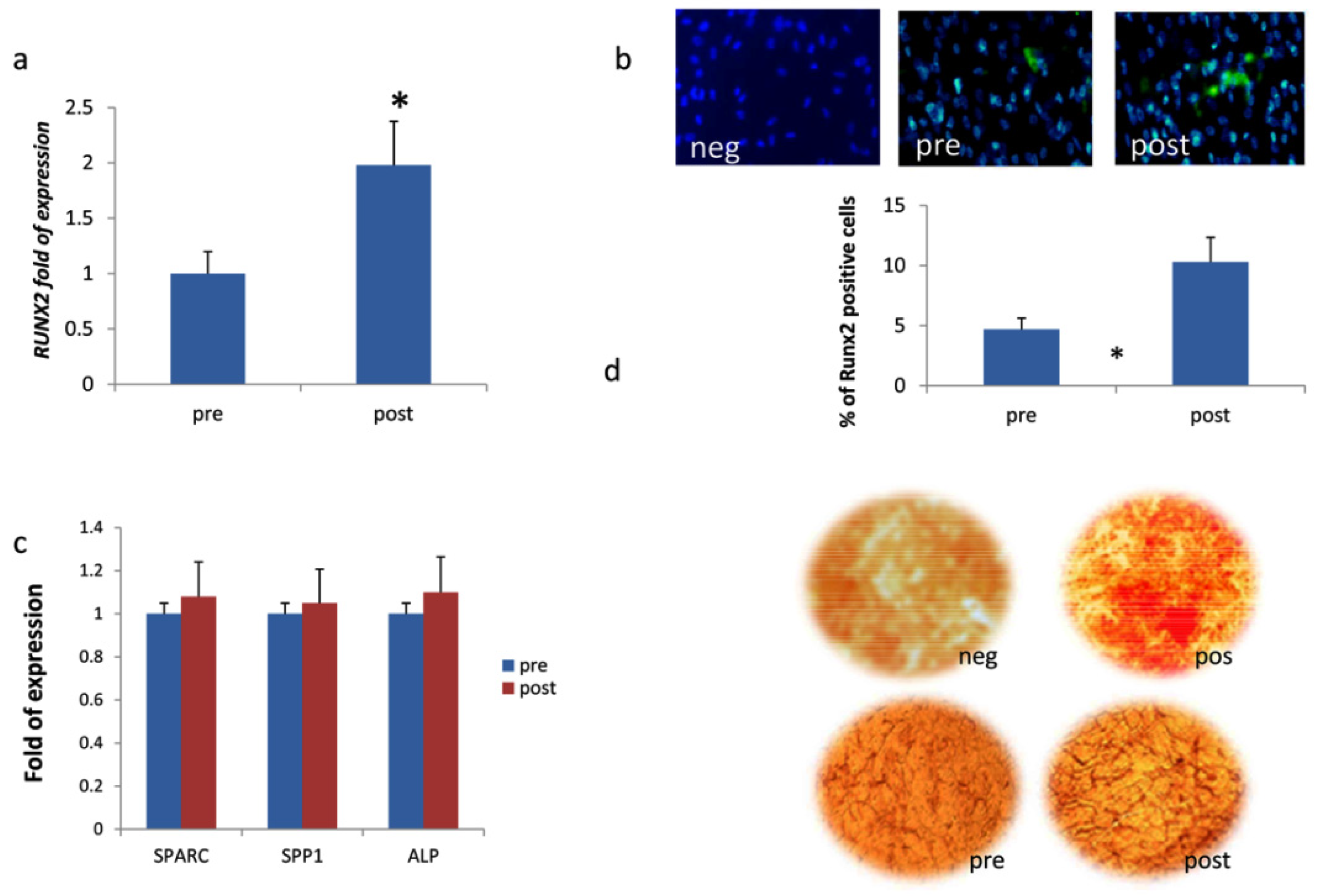
3.2. DOACs Enhance Osteogenic Commitment
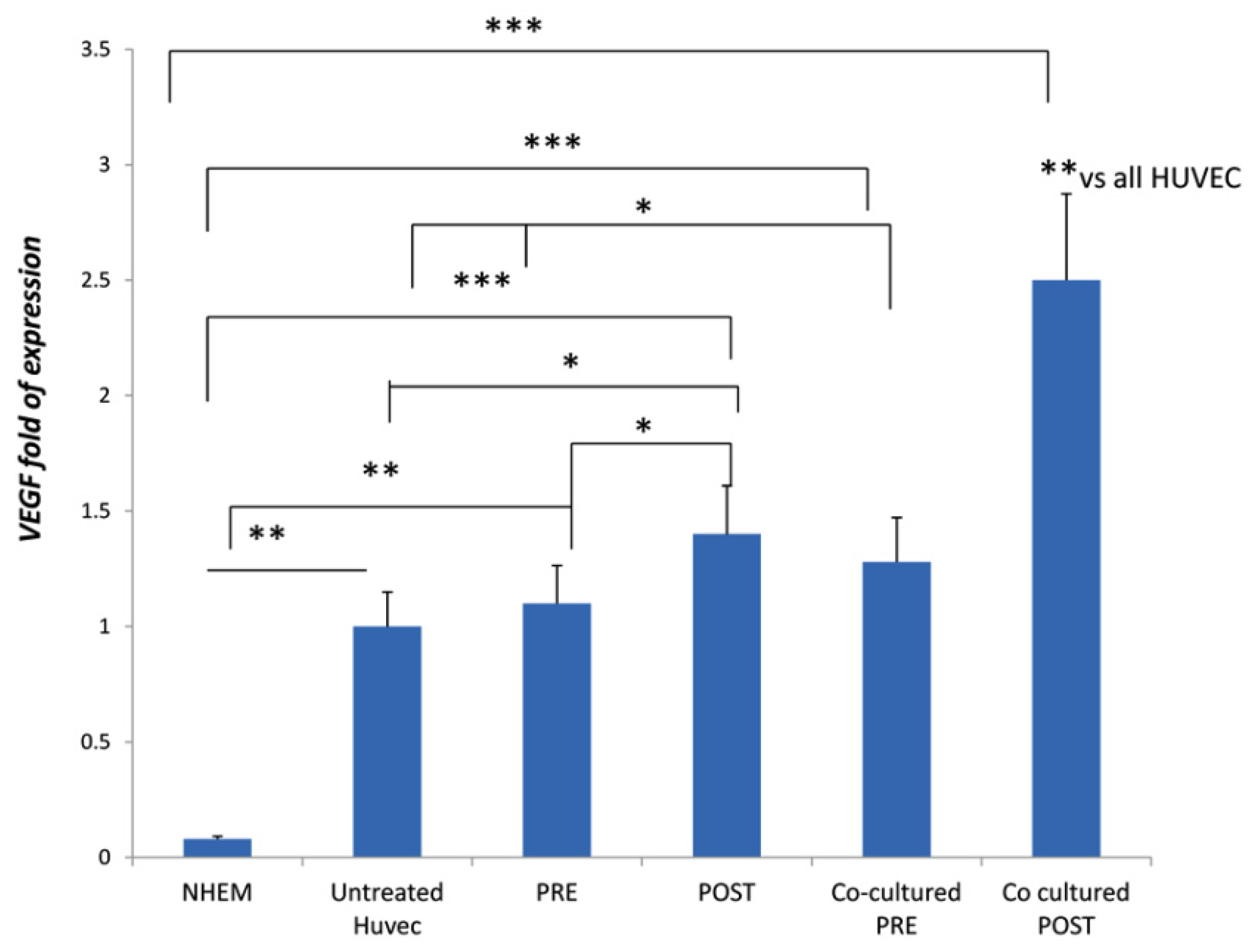
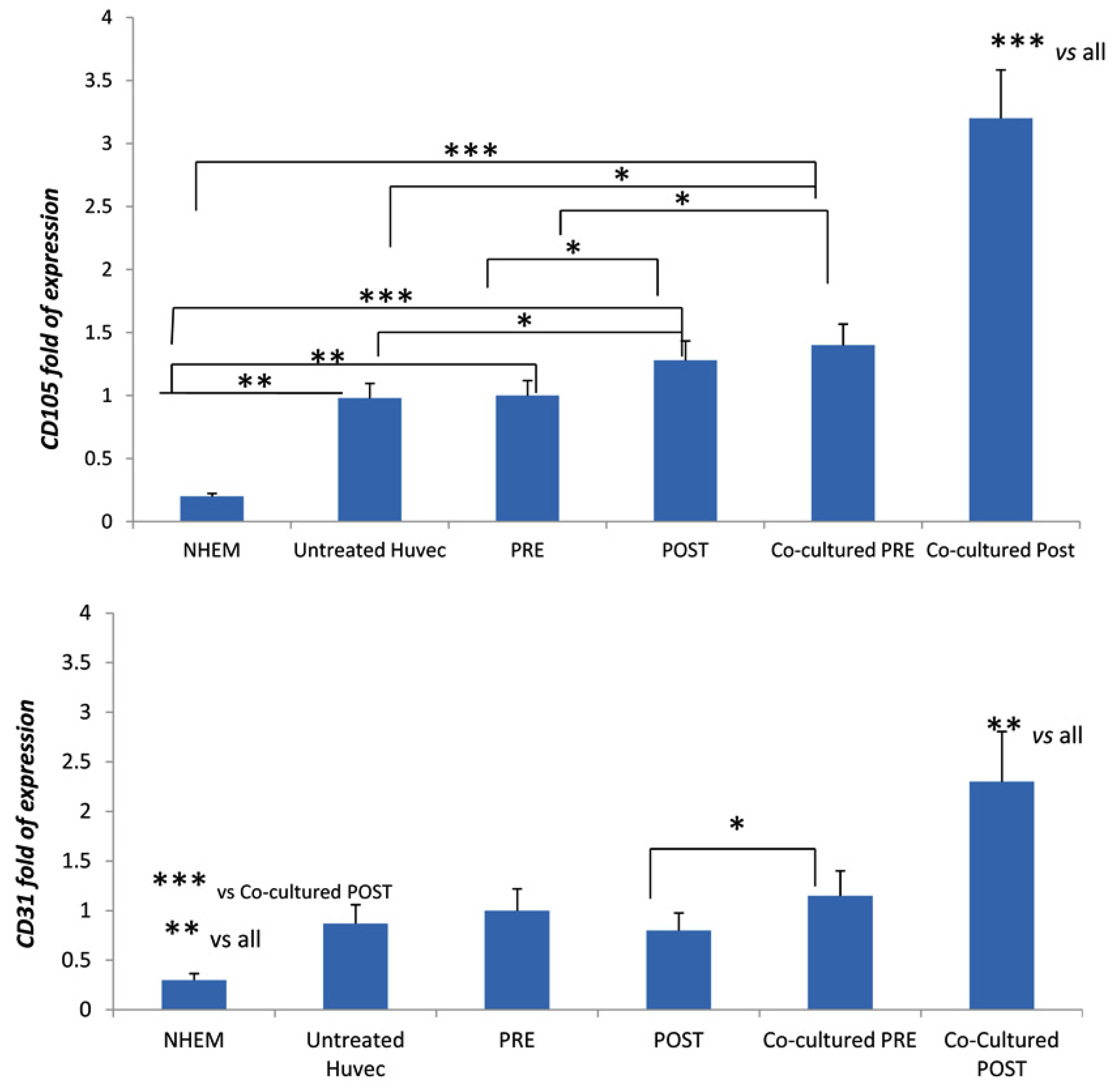
3.3. DOACs Increase Expression of Key Angiogenic Markers in Presence of MSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tummala, R.; Kavtaradze, A.; Gupta, A.; Ghosh, R.K. Specific antidotes against direct oral anticoagulants: A comprehensive review of clinical trials data. Int. J. Cardiol. 2016, 214, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.T.; Lip, G.Y.; De Caterina, R.; Heidbuchel, H.; Zamorano, J.L.; Agnelli, G.; Verheugt, F.; Camm, A.J. State of play and future direction with NOACs: An expert consensus. Vascul. Pharmacol. 2018, 106, 9–21. [Google Scholar] [CrossRef]
- Becattini, C.; Vedovati, M.C.; Agnelli, G. Old and new oral anticoagulants for venous thromboembolism and atrial fibrillation: A review of the literature. Thromb. Res. 2012, 129, 392–400. [Google Scholar] [CrossRef]
- Spronk, H.M.; de Jong, A.M.; Crijns, H.J.; Schotten, U.; Van Gelder, I.C.; Ten Cate, H. Pleiotropic effects of factor Xa and thrombin: What to expect from novel anticoagulants. Cardiovasc. Res. 2014, 101, 344–351. [Google Scholar] [CrossRef]
- Choi, H.J.; Kim, N.E.; Kim, J.; An, S.; Yang, S.H.; Ha, J.; Cho, S.; Kwon, I.; Kim, Y.D.; Nam, H.S.; et al. Dabigatran reduces endothelial permeability through inhibition of thrombin-induced cytoskeleton reorganization. Thromb. Res. 2018, 167, 165–171. [Google Scholar] [CrossRef]
- Lopez-Farre, A.J.; Rodriguez-Sierra, P.; Modrego, J.; Segura, A.; Martin-Palacios, N.; Saiz, A.M.; Zamorano-Leon, J.J.; Duarte, J.; Serrano, J.; Monux, G. Effects of factor Xa on the expression of proteins in femoral arteries from type 2 diabetic patients. Br. J. Clin. Pharmacol. 2014, 78, 1366–1377. [Google Scholar] [CrossRef] [Green Version]
- Sanada, F.; Muratsu, J.; Otsu, R.; Shimizu, H.; Koibuchi, N.; Uchida, K.; Taniyama, Y.; Yoshimura, S.; Rakugi, H.; Morishita, R. Local Production of Activated Factor X in Atherosclerotic Plaque Induced Vascular Smooth Muscle Cell Senescence. Sci. Rep. 2017, 7, 17172. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.C.; Chan, J.S.; Lee, C.Y.; Leu, H.B.; Huang, P.H.; Chen, J.S.; Lin, S.J.; Chen, J.W. Rivaroxaban, a factor Xa inhibitor, improves neovascularization in the ischemic hindlimb of streptozotocin-induced diabetic mice. Cardiovasc. Diabetol. 2015, 14, 81. [Google Scholar] [CrossRef] [Green Version]
- Villari, A.; Giurdanella, G.; Bucolo, C.; Drago, F.; Salomone, S. Apixaban Enhances Vasodilatation Mediated by Protease-Activated Receptor 2 in Isolated Rat Arteries. Front. Pharmacol. 2017, 8, 480. [Google Scholar] [CrossRef]
- Iba, T.; Aihara, K.; Yamada, A.; Nagayama, M.; Tabe, Y.; Ohsaka, A. Rivaroxaban attenuates leukocyte adhesion in the microvasculature and thrombus formation in an experimental mouse model of type 2 diabetes mellitus. Thromb. Res. 2014, 133, 276–280. [Google Scholar] [CrossRef]
- Bidarra, S.J.; Barrias, C.C.; Barbosa, M.A.; Soares, R.; Amedee, J.; Granja, P.L. Phenotypic and proliferative modulation of human mesenchymal stem cells via crosstalk with endothelial cells. Stem Cell Res. 2011, 7, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.G.; Zhao, X.; Yang, Q.; Li, Y.; Ge, C.; Zhao, G.; Franceschi, R.T. Physical and functional interactions between Runx2 and HIF-1alpha induce vascular endothelial growth factor gene expression. J. Cell. Biochem. 2011, 112, 3582–3593. [Google Scholar] [CrossRef]
- Kim, I.S.; Otto, F.; Zabel, B.; Mundlos, S. Regulation of chondrocyte differentiation by Cbfa1. Mech. Dev. 1999, 80, 159–170. [Google Scholar] [CrossRef]
- Bronckers, A.L.; Sasaguri, K.; Cavender, A.C.; D’Souza, R.N.; Engelse, M.A. Expression of Runx2/Cbfa1/Pebp2alphaA during angiogenesis in postnatal rodent and fetal human orofacial tissues. J. Bone Miner. Res. 2005, 20, 428–437. [Google Scholar] [CrossRef]
- Namba, K.; Abe, M.; Saito, S.; Satake, M.; Ohmoto, T.; Watanabe, T.; Sato, Y. Indispensable role of the transcription factor PEBP2/CBF in angiogenic activity of a murine endothelial cell MSS31. Oncogene 2000, 19, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Carbonare, L.D.; Mottes, M.; Malerba, G.; Mori, A.; Zaninotto, M.; Plebani, M.; Dellantonio, A.; Valenti, M.T. Enhanced Osteogenic Differentiation in Zoledronate-Treated Osteoporotic Patients. Int. J. Mol. Sci. 2017, 18, 1261. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Verstegen, M.M.; Engela, A.U.; Korevaar, S.S.; Roemeling-van Rhijn, M.; Merino, A.; Franquesa, M.; de Jonge, J.; Ijzermans, J.N.; Weimar, W.; et al. No evidence for circulating mesenchymal stem cells in patients with organ injury. Stem Cells Dev. 2014, 23, 2328–2335. [Google Scholar] [CrossRef]
- Xu, L.; Li, G. Circulating mesenchymal stem cells and their clinical implications. J. Orthop. Transl. 2014, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Valenti, M.T.; Mottes, M.; Cheri, S.; Deiana, M.; Micheletti, V.; Cosaro, E.; Davi, M.V.; Francia, G.; Carbonare, L.D. Runx2 overexpression compromises bone quality in acromegalic patients. Endocr.-Relat. Cancer 2018, 25, 269–277. [Google Scholar] [CrossRef]
- Thoma, S.J.; Lamping, C.P.; Ziegler, B.L. Phenotype analysis of hematopoietic CD34+ cell populations derived from human umbilical cord blood using flow cytometry and cDNA-polymerase chain reaction. Blood 1994, 83, 2103–2114. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Khodarev, N.N.; Yu, J.; Labay, E.; Darga, T.; Brown, C.K.; Mauceri, H.J.; Yassari, R.; Gupta, N.; Weichselbaum, R.R. Tumour-endothelium interactions in co-culture: Coordinated changes of gene expression profiles and phenotypic properties of endothelial cells. J. Cell Sci. 2003, 116, 1013–1022. [Google Scholar] [CrossRef]
- Liu, Z.; Lebrin, F.; Maring, J.A.; van den Driesche, S.; van der Brink, S.; van Dinther, M.; Thorikay, M.; Martin, S.; Kobayashi, K.; Hawinkels, L.J.; et al. ENDOGLIN is dispensable for vasculogenesis, but required for vascular endothelial growth factor-induced angiogenesis. PLoS ONE 2014, 9, e86273. [Google Scholar] [CrossRef]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial functions of platelet/endothelial cell adhesion molecule-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef]
- Gong, M.; Yu, B.; Wang, J.; Wang, Y.; Liu, M.; Paul, C.; Millard, R.W.; Xiao, D.S.; Ashraf, M.; Xu, M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget 2017, 8, 45200–45212. [Google Scholar] [CrossRef] [Green Version]
- Morishima, Y.; Kamisato, C.; Honda, Y.; Furugohri, T.; Shibano, T. The effects of warfarin and edoxaban, an oral direct factor Xa inhibitor, on gammacarboxylated (Gla-osteocalcin) and undercarboxylated osteocalcin (uc-osteocalcin) in rats. Thromb. Res. 2013, 131, 59–63. [Google Scholar] [CrossRef]
- Jeong, H.M.; Cho, D.H.; Jin, Y.H.; Chung, J.O.; Chung, M.Y.; Chung, D.J.; Lee, K.Y. Inhibition of osteoblastic differentiation by warfarin and 18-alpha-glycyrrhetinic acid. Arch. Pharm. Res. 2011, 34, 1381–1387. [Google Scholar] [CrossRef]
- Namba, S.; Yamaoka-Tojo, M.; Kakizaki, R.; Nemoto, T.; Fujiyoshi, K.; Hashikata, T.; Kitasato, L.; Hashimoto, T.; Kameda, R.; Meguro, K.; et al. Effects on bone metabolism markers and arterial stiffness by switching to rivaroxaban from warfarin in patients with atrial fibrillation. Heart Vessel. 2017, 32, 977–982. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Previous Therapy | No. of Patients |
|---|---|---|
| NVAF | No treatment | 0 |
| Warfarin | 12 | |
| Enoxaparin | 2 | |
| ASA | 1 | |
| VTE | No treatment | 3 |
| Warfarin | 2 | |
| enoxaparin/fondaparinux | 3 | |
| ASA | 0 | |
| Ischemic stroke | No treatment | 3 |
| Warfarin | 2 | |
| Enoxaparin | 0 | |
| ASA | 6 |
| Parameter | Baseline | 3 Months | p-Value |
|---|---|---|---|
| Hemoglobin (g/dL) | 13.4 ± 1.7 | 13.1 ± 2.0 | n.s. |
| Hematocrit, (%) | 41 ± 5 | 40 ± 6 | n.s. |
| Platelets, (109/L) | 209 ± 71 | 217 ± 65 | n.s. |
| PT-INR | 1.71 ± 0.65 | 1.27 ± 0.23 | n.s. |
| aPTT, ratio | 1.08 ± 0.20 | 1.18 ± 0.44 | n.s. |
| Cholesterol, (mg/dL) | 160 ± 30 | 162 ± 38 | n.s. |
| HDL, (mg/dL) | 52 ± 14 | 53 ± 14 | n.s. |
| Triglycerides, (mg/dL) | 111 ± 42 | 105 ± 30 | n.s. |
| LDL, (mg/dL) | 79 ± 24 | 83 ± 33 | n.s. |
| Creatinine, (mg/dL) | 1.09 ± 0.44 | 1.16 ± 0.45 | n.s. |
| eGFR, (mL/min) | 61 ± 22 | 57 ± 21 | n.s. |
| AST, (U/L) | 24 ± 14 | 19 ± 7 | n.s. |
| ALT, (U/L) | 23 ± 16 | 18 ± 8 | n.s. |
| Calcium, (mg/dL) | 8.7 ± 0.3 | 9.4 ± 0.4 | n.s. |
| Vitamin D (25-OH), (ng/mL) | 17 ± 7 | 25 ± 14 | n.s. |
| CTX, (ng/mL) | 0.31 ± 0.09 | 0.36 ± 0.12 | <0.05 |
| Parameter | NVAF | VTE | Ischemic Stroke | p Value |
|---|---|---|---|---|
| PT-INR (baseline) | 2.2 ± 0.5 | 1.5 ± 0.5 | 1.3 ± 0.3 | <0.05 |
| Cluster Differentiation Transcript | Pre DOACS | Post DOACS | p-Value |
|---|---|---|---|
| CD105 | 65 ± 0.6% | 64% (±0.5) | n.s |
| CD 73 | 73% (±0.3) | 72 (±0.2) | n.s |
| CD3 | 0% | 0% | n.s |
| CD14 | 0.3% (±0.04) | 0.4% (±0.06) | n.s |
| CD19 | 0% | 0% | n.s |
| CD45 | 1.6% (±0.3) | 1.8% (±0.4) | n.s |
| CD34 | low level | low level | n.s |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalle Carbonare, L.; Mottes, M.; Brunelli, A.; Deiana, M.; Cheri, S.; Suardi, S.; Valenti, M.T. Effects of Oral Anticoagulant Therapy on Gene Expression in Crosstalk between Osteogenic Progenitor Cells and Endothelial Cells. J. Clin. Med. 2019, 8, 329. https://doi.org/10.3390/jcm8030329
Dalle Carbonare L, Mottes M, Brunelli A, Deiana M, Cheri S, Suardi S, Valenti MT. Effects of Oral Anticoagulant Therapy on Gene Expression in Crosstalk between Osteogenic Progenitor Cells and Endothelial Cells. Journal of Clinical Medicine. 2019; 8(3):329. https://doi.org/10.3390/jcm8030329
Chicago/Turabian StyleDalle Carbonare, Luca, Monica Mottes, Anna Brunelli, Michela Deiana, Samuele Cheri, Silvia Suardi, and Maria Teresa Valenti. 2019. "Effects of Oral Anticoagulant Therapy on Gene Expression in Crosstalk between Osteogenic Progenitor Cells and Endothelial Cells" Journal of Clinical Medicine 8, no. 3: 329. https://doi.org/10.3390/jcm8030329
APA StyleDalle Carbonare, L., Mottes, M., Brunelli, A., Deiana, M., Cheri, S., Suardi, S., & Valenti, M. T. (2019). Effects of Oral Anticoagulant Therapy on Gene Expression in Crosstalk between Osteogenic Progenitor Cells and Endothelial Cells. Journal of Clinical Medicine, 8(3), 329. https://doi.org/10.3390/jcm8030329