The Beneficial Effect of Acute Exercise on Motor Memory Consolidation is Modulated by Dopaminergic Gene Profile

 ,
,  , and
, and
Abstract
:1. Introduction
Dopamine Transmission Influences Memory Consolidation
2. Study Design and Data Analysis
3. Results and Discussions
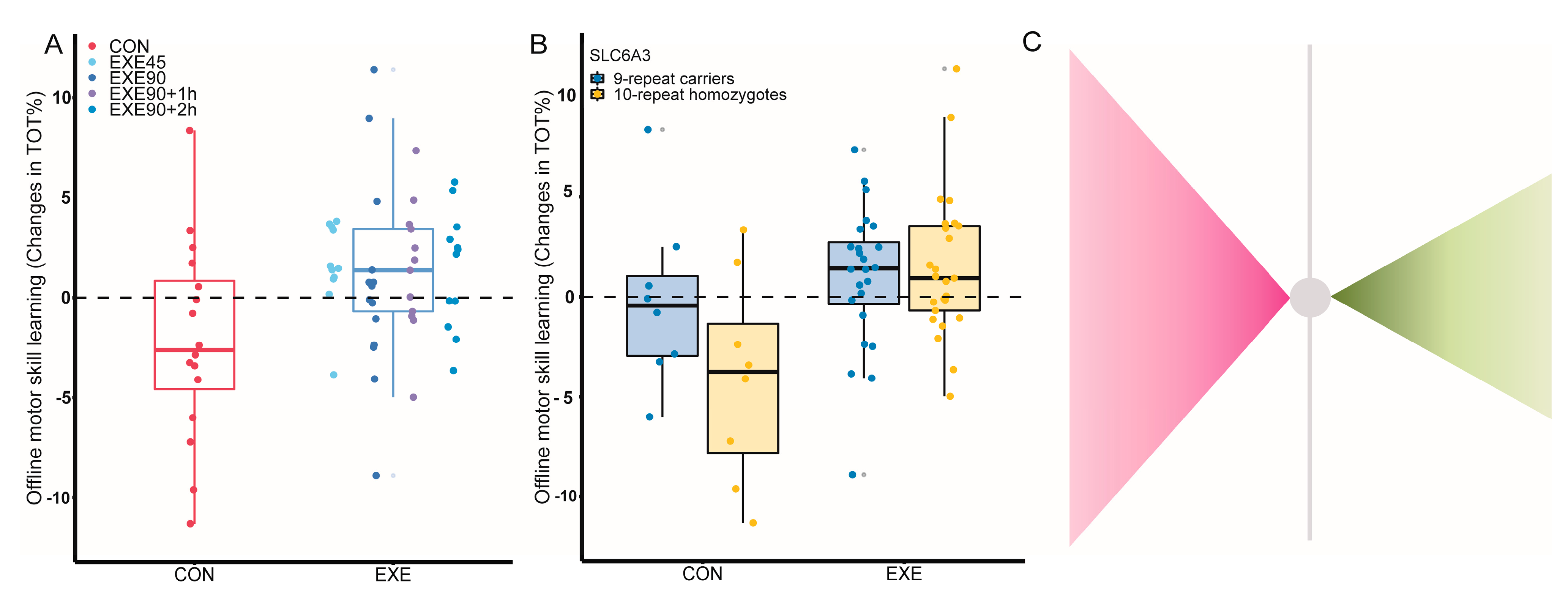
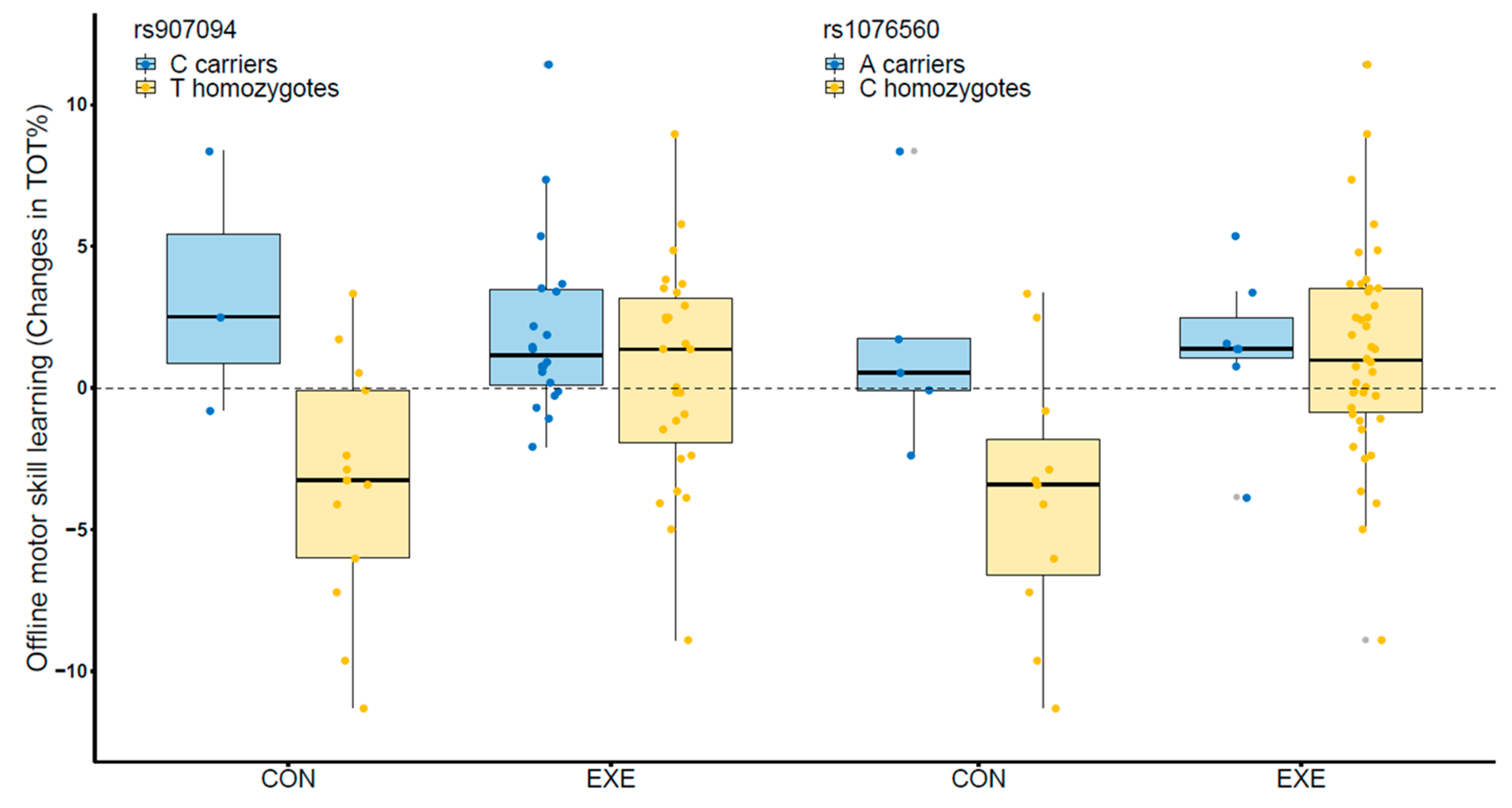
3.1. Variations in DRD2, PPP1R1B, and SLC6A3 Influence the Effects of Exercise
3.2. Exercise Benefits Individuals with Allele Combinations Associated with Lower Consolidation
3.3. Val66Met Polymorphism Did Not Influence Consolidation or Interact with Exercise
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Participants

{kind=link}
{kind=link}
{kind=link}
| Group | Control | Exercise | |||
|---|---|---|---|---|---|
| EXE90 | EXE45 | EXE90 + 1 h | EXE90 + 2 h | ||
| Number of participants | 16 | 14 | 11 | 12 | 12 |
| Age (years) | 24.4 ± 2.8 | 25.4 ± 2.7 | 23.6 ± 2.4 | 24.1 ± 2.3 | 23.6 ± 2.0 |
| Weight (kg) | 78.33 ± 8.3 | 78.9 ± 11.1 | 80.6 ± 6.7 | 80.4 ± 6.7 | 78.8 ± 13.1 |
| Height (cm) | 185 ± 7.0 | 182 ± 5 | 187 ± 7 | 184 ± 8 | 182 ± 7 |
| BMI (kg/m2) | 22.9 ± 2.0 | 23.8 ± 2.5 | 23.2 ± 1.3 | 23.8 ± 1.9 | 23.6 ± 2.8 |
| VO2peak (mL O2· kg−1·min−1) | 51.99 ± 4.0 | 49.5 ± 7.4 | 49.6 ± 3.9 | 49.0 ± 5.6 | 50.4 ± 6.9 |
| Baseline motor performance (VAT) | 44.6 ± 11.6 | 43.8 ± 12.4 | 51.3 ± 9.6 | 52.5 ± 8.8 | 50.9 ± 7.5 |
| Post-acquisition motor performance (VAT) | 68.1 ± 6.3 | 68.6 ± 5.2 | 71.9 ± 4.3 | 72.8 ± 4.8 | 71.9 ± 4.8 |
Appendix A.2. Study Design
Appendix A.3. Graded Maximal Exercise Test
Appendix A.4. Visuomotor Accuracy Tracking Task (VAT)
Appendix A.5. Exercise Protocol
Appendix A.6. DNA Purification and Preparation
Appendix A.7. Genotype Data Analysis
| Gene ID | SNP ID | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|---|
| COMT | rs4680 | ACGTTGGATGACCATCGAGATCAACCCCG | ACGTTGGATGTTTTCCAGGTCTGACAACGG |
| DRD2 | rs6277 | ACGTTGGATGCATTCTTCTCTGGTTTGGCG | ACGTTGGATGACCAGCTGACTCTCCCCGA |
| DRD3 | rs6280 | ACGTTGGATGCATAGTAGGCATGTGGGCG | ACGTTGGATGCTCTGGGCTATGGCATCTCT |
| DRD2 | rs1076560 | ACGTTGGATGTAAAGCCGGACAAGTTCCCA | ACGTTGGATGTGTGGTGTTTGCAGGAGTCT |
| DRD1 | rs686 | ACGTTGGATGAGAGTCTCACCGTACCTTAG | ACGTTGGATGCCTGAACTCGCAGATGAATC |
| BDNF | rs6265 | ACGTTGGATGTTGTTTTCTTCATTGGGCCG | ACGTTGGATGGCTTGACATCATTGGCTGAC |
| ANKK1 | rs1800497 | ACGTTGGATGTCAAGGGCAACACAGCCATC | ACGTTGGATGGACATGATGCCCTGCTTTCG |
| PPP1R1B | rs907094 | ACGTTGGATGTGAAGGTCATCAGGCAGTCT | ACGTTGGATGGGACGTCCTCGTATACTCAA |
Appendix A.8. Statistical Analysis
| SNP ID | rs1076560 | rs1800497 | rs907094 | rs28363170 | ||||
|---|---|---|---|---|---|---|---|---|
| Allelic combination | C/C | C/A + A/A | C/C | C/T + T/T | T/T | C/T + CC | 10/10 | 10/9 + 9/9 |
| CON | 11 | 5 | 10 | 4 | 13 | 3 | 8 | 8 |
| EX90 | 13 | 1 | 13 | 1 | 5 | 8 | 7 | 6 |
| EX45 | 7 | 4 | 7 | 4 | 7 | 3 | 5 | 5 |
| EX90 + 1 h | 11 | 1 | 9 | 2 | 7 | 5 | 7 | 7 |
| EX90 + 2 h | 11 | 1 | 10 | 2 | 8 | 4 | 5 | 7 |
| Not Identified | 0 | N/A | 3 | N/A | 2 | N/A | 0 | N/A |

References
- McGaugh, J.L. Memory—A Century of Consolidation. Science 2000, 287, 248–251. [Google Scholar] [CrossRef]
- Hötting, K.; Schickert, N.; Kaiser, J.; Röder, B.; Schmidt-Kassow, M. The effects of acute physical exercise on memory, peripheral BDNF, and cortisol in young adults. Neural Plast. 2016, 2016, 6860573. [Google Scholar] [CrossRef] [PubMed]
- Delancey, D.; Frith, E.; Sng, E.; Loprinzi, P.D. Randomized Controlled Trial Examining the Long-Term Memory Effects of Acute Exercise During the Memory Consolidation Stage of Memory Formation. J. Cogn. Enhanc. 2018. [Google Scholar] [CrossRef]
- Perini, R.; Bortoletto, M.; Capogrosso, M.; Fertonani, A.; Miniussi, C. Acute effects of aerobic exercise promote learning. Sci. Rep. 2016, 6, 25440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambourne, K.; Tomporowski, P. The effect of exercise-induced arousal on cognitive task performance: A meta-regression analysis. Brain Res. 2010, 1341, 12–24. [Google Scholar] [CrossRef]
- Frank, M.J.; Doll, B.B.; Oas-Terpstra, J.; Moreno, F. Prefrontal and striatal dopaminergic genes predict individual differences in exploration and exploitation. Nat. Neurosci. 2009, 12, 1062. [Google Scholar] [CrossRef]
- Seidler, R.; Carson, R. Sensorimotor learning: Neurocognitive mechanisms and individual differences. J. Neuroeng. Rehabil. 2017, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; Campbell, K.L.; Ross, C.J.; Boyd, L.A. Promoting neuroplasticity for motor rehabilitation after stroke: Considering the effects of aerobic exercise and genetic variation on brain-derived neurotrophic factor. Phys. Ther. 2013, 93, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-K.; Labban, J.; Gapin, J.; Etnier, J.L. The effects of acute exercise on cognitive performance: A meta-analysis. Brain Res. 2012, 1453, 87–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loprinzi, P.D. Intensity-specific effects of acute exercise on human memory function: Considerations for the timing of exercise and the type of memory. Health Promot. Perspect. 2018, 8, 255–262. [Google Scholar] [CrossRef]
- Ferrer-Uris, B.; Busquets, A.; Lopez-Alonso, V.; Fernandez-del-Olmo, M.; Angulo-Barroso, R. Enhancing consolidation of a rotational visuomotor adaptation task through acute exercise. PLoS ONE 2017, 12, e0175296. [Google Scholar] [CrossRef] [PubMed]
- Tomporowski, P.D.; Pendleton, D.M. Effects of the timing of acute exercise and movement complexity on young adults’ psychomotor learning. J. Sport Exerc. Psychol. 2018, 40, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Uris, B.; Busquets, A.; Angulo-Barroso, R. Adaptation and Retention of a Perceptual-Motor Task in Children: Effects of a Single Bout of Intense Endurance Exercise. J. Sport Exerc. Psychol. 2018, 40, 1–9. [Google Scholar] [CrossRef]
- Thomas, R.; Beck, M.M.; Lind, R.R.; Korsgaard Johnsen, L.; Geertsen, S.S.; Christiansen, L.; Ritz, C.; Roig, M.; Lundbye-Jensen, J. Acute exercise and motor memory consolidation: The role of exercise timing. Neural Plast. 2016, 2016, 6205452. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Johnsen, L.K.; Geertsen, S.S.; Christiansen, L.; Ritz, C.; Roig, M.; Lundbye-Jensen, J. Acute exercise and motor memory consolidation: The role of exercise intensity. PLoS ONE 2016, 11, e0159589. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Flindtgaard, M.; Skriver, K.; Geertsen, S.S.; Christiansen, L.; Korsgaard Johnsen, L.; Busk, D.V.P.; Bojsen-Møller, E.; Madsen, M.; Ritz, C. Acute exercise and motor memory consolidation: Does exercise type play a role? Scand. J. Med. Sci. Sports 2017, 27, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Roig, M.; Skriver, K.; Lundbye-Jensen, J.; Kiens, B.; Nielsen, J.B. A single bout of exercise improves motor memory. PLoS ONE 2012, 7, e44594. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.S.; Chen, J.; Riechman, S.; Roig, M.; Wright, D.L. The protective effects of acute cardiovascular exercise on the interference of procedural memory. Psychol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ostadan, F.; Centeno, C.; Daloze, J.-F.; Frenn, M.; Lundbye-Jensen, J.; Roig, M. Changes in corticospinal excitability during consolidation predict acute exercise-induced off-line gains in procedural memory. Neurobiol. Learn. Mem. 2016, 136, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Dal Maso, F.; Desormeau, B.; Boudrias, M.-H.; Roig, M. Acute cardiovascular exercise promotes functional changes in cortico-motor networks during the early stages of motor memory consolidation. NeuroImage 2018, 174, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Skriver, K.; Roig, M.; Lundbye-Jensen, J.; Pingel, J.; Helge, J.W.; Kiens, B.; Nielsen, J.B. Acute exercise improves motor memory: Exploring potential biomarkers. Neurobiol. Learn. Mem. 2014, 116, 46–58. [Google Scholar] [CrossRef]
- Segal, S.K.; Cotman, C.W.; Cahill, L.F. Exercise-induced noradrenergic activation enhances memory consolidation in both normal aging and patients with amnestic mild cognitive impairment. J. Alzheimer’s Dis. 2012, 32, 1011–1018. [Google Scholar] [CrossRef]
- Eddy, M.C.; Stansfield, K.J.; Green, J.T. Voluntary exercise improves performance of a discrimination task through effects on the striatal dopamine system. Learn. Mem. 2014, 21, 334–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyes, M.P.; Garnett, E.S.; Coates, G. Nigrostriatal dopaminergic activity is increased during exhaustive exercise stress in rats. Life Sci. 1988, 42, 1537–1542. [Google Scholar] [CrossRef]
- Koch, G.; Johansson, U.; Arvidsson, E. Radioenzymatic determination of epinephrine, norepinephrine and dopamine in 0.1 mL plasma samples. Plasma catecholamine response to submaximal and near maximal exercise. Clin. Chem. Lab. Med. 1980, 18, 367–372. [Google Scholar] [CrossRef]
- Van Loon, G.R.; Schwartz, L.; Sole, M.J. Plasma dopamine responses to standing and exercise in man. Life Sci. 1979, 24, 2273–2277. [Google Scholar] [CrossRef]
- Winter, B.; Breitenstein, C.; Mooren, F.C.; Voelker, K.; Fobker, M.; Lechtermann, A.; Krueger, K.; Fromme, A.; Korsukewitz, C.; Floel, A.; et al. High impact running improves learning. Neurobiol. Learn. Mem. 2007, 87, 597–609. [Google Scholar] [CrossRef]
- Meeusen, R.; Smolders, I.; Sarre, S.; De Meirleir, K.; Keizer, H.; Serneels, M.; Ebinger, G.; Michotte, Y. Endurance training effects on neurotransmitter release in rat striatum: An in vivo microdialysis study. Acta Physiol. Scand. 1997, 159, 335–341. [Google Scholar] [CrossRef]
- Hattori, S.; Naoi, M.; Nishino, H. Striatal dopamine turnover during treadmill running in the rat: Relation to the speed of running. Brain Res. Bull. 1994, 35, 41–49. [Google Scholar] [CrossRef]
- Cestari, V.; Castellano, C.; Cabib, S.; Puglisi-Allegra, S. Strain-dependent effects of post-training dopamine receptor agonists and antagonists on memory storage in mice. Behav. Neural Biol. 1992, 58, 58–63. [Google Scholar] [CrossRef]
- Pearson-Fuhrhop, K.M.; Minton, B.; Acevedo, D.; Shahbaba, B.; Cramer, S.C. Genetic variation in the human brain dopamine system influences motor learning and its modulation by L-Dopa. PLoS ONE 2013, 8, e61197. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; McEwen, L.M.; MacIsaac, J.L.; Snow, N.J.; Campbell, K.L.; Kobor, M.S.; Ross, C.J.; Boyd, L.A. Exploring genetic influences underlying acute aerobic exercise effects on motor learning. Sci. Rep. 2017, 7, 12123. [Google Scholar] [CrossRef] [Green Version]
- White, N.M.; Packard, M.G.; Seamans, J. Memory enhancement by post-training peripheral administration of low doses of dopamine agonists: Possible autoreceptor effect. Behav. Neural Biol. 1993, 59, 230–241. [Google Scholar] [CrossRef]
- Packard, M.G.; White, N.M. Memory facilitation produced by dopamine agonists: Role of receptor subtype and mnemonic requirements. Pharmacol. Biochem. Behav. 1989, 33, 511–518. [Google Scholar] [CrossRef]
- Gozzani, J.L.; Izquierdo, I. Possible peripheral adrenergic and central dopaminergic influences in memory consolidation. Psychopharmacology 1976, 49, 109–111. [Google Scholar]
- Williams, C.L.; Packard, M.G.; McGaugh, J.L. Amphetamine facilitation of win-shift radial-arm maze retention: The involvement of peripheral adrenergic and central dopaminergic systems. Psychobiology 1994, 22, 141–148. [Google Scholar]
- Levesque, M.; Bedard, M.A.; Courtemanche, R.; Tremblay, P.L.; Scherzer, P.; Blanchet, P.J. Raclopride-induced motor consolidation impairment in primates: Role of the dopamine type-2 receptor in movement chunking into integrated sequences. Exp. Brain Res. 2007, 182, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.J.; Fossella, J.A. Neurogenetics and Pharmacology of Learning, Motivation, and Cognition. Neuropsychopharmacology 2010, 36, 133. [Google Scholar] [CrossRef]
- Schultz, W. Dopamine reward prediction-error signalling: A two-component response. Nat. Rev. Neurosci. 2016, 17, 183. [Google Scholar] [CrossRef]
- Abe, M.; Schambra, H.; Wassermann, E.M.; Luckenbaugh, D.; Schweighofer, N.; Cohen, L.G. Reward improves long-term retention of a motor memory through induction of offline memory gains. Curr. Biol. 2011, 21, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Galea, J.M.; Mallia, E.; Rothwell, J.; Diedrichsen, J. The dissociable effects of punishment and reward on motor learning. Nat. Neurosci. 2015, 18, 597. [Google Scholar] [CrossRef] [PubMed]
- Hosp, J.; Molina-Luna, K.; Hertler, B.; Atiemo, C.O.; Luft, A. Dopaminergic modulation of motor maps in rat motor cortex: An in vivo study. Neuroscience 2009, 159, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Hosp, J.A.; Pekanovic, A.; Rioult-Pedotti, M.S.; Luft, A.R. Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning. J. Neurosci. 2011, 31, 2481–2487. [Google Scholar] [CrossRef] [PubMed]
- Molina-Luna, K.; Pekanovic, A.; Röhrich, S.; Hertler, B.; Schubring-Giese, M.; Rioult-Pedotti, M.-S.; Luft, A.R. Dopamine in motor cortex is necessary for skill learning and synaptic plasticity. PLoS ONE 2009, 4, e7082. [Google Scholar] [CrossRef]
- Sheynikhovich, D.; Otani, S.; Arleo, A. Dopaminergic Control of Long-Term Depression/Long-Term Potentiation Threshold in Prefrontal Cortex. J. Neurosci. 2013, 33, 13914–13926. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Smith, A.; McGinty, J. A single injection of amphetamine or methamphetamine induces dynamic alterations in c-fos, zif/268 and preprodynorphin messenger RNA expression in rat forebrain. Neuroscience 1995, 68, 83–95. [Google Scholar] [CrossRef]
- Mao, L.; Wang, J.Q. Group I metabotropic glutamate receptor-mediated calcium signalling and immediate early gene expression in cultured rat striatal neurons. Eur. J. Neurosci. 2003, 17, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; McGinty, J. Scopolamine augments c-fos and zif/268 messenger RNA expression induced by the full D1 dopamine receptor agonist SKF-82958 in the intact rat striatum. Neuroscience 1996, 72, 601–616. [Google Scholar] [CrossRef]
- Fleischmann, A.; Hvalby, O.; Jensen, V.; Strekalova, T.; Zacher, C.; Layer, L.E.; Kvello, A.; Reschke, M.; Spanagel, R.; Sprengel, R. Impaired long-term memory and NR2A-type NMDA receptor-dependent synaptic plasticity in mice lacking c-Fos in the CNS. J. Neurosci. 2003, 23, 9116–9122. [Google Scholar] [CrossRef]
- Fazio, L.; Blasi, G.; Taurisano, P.; Papazacharias, A.; Romano, R.; Gelao, B.; Ursini, G.; Quarto, T.; Lo Bianco, L.; Di Giorgio, A.; et al. D2 receptor genotype and striatal dopamine signaling predict motor cortical activity and behavior in humans. NeuroImage 2011, 54, 2915–2921. [Google Scholar] [CrossRef]
- Jocham, G.; Klein, T.A.; Neumann, J.; von Cramon, D.Y.; Reuter, M.; Ullsperger, M. Dopamine DRD2 polymorphism alters reversal learning and associated neural activity. J. Neurosci. 2009, 29, 3695–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolino, A.; Taurisano, P.; Pisciotta, N.M.; Blasi, G.; Fazio, L.; Romano, R.; Gelao, B.; Bianco, L.L.; Lozupone, M.; Di Giorgio, A.; et al. Genetically Determined Measures of Striatal D2 Signaling Predict Prefrontal Activity during Working Memory Performance. PLoS ONE 2010, 5, e9348. [Google Scholar] [CrossRef]
- Zhang, Y.; Bertolino, A.; Fazio, L.; Blasi, G.; Rampino, A.; Romano, R.; Lee, M.-L.T.; Xiao, T.; Papp, A.; Wang, D. Polymorphisms in human dopamine D2 receptor gene affect gene expression, splicing, and neuronal activity during working memory. Proc. Natl. Acad. Sci. USA 2007, 104, 20552–20557. [Google Scholar] [CrossRef] [Green Version]
- Fuke, S.; Suo, S.; Takahashi, N.; Koike, H.; Sasagawa, N.; Ishiura, S. The VNTR polymorphism of the human dopamine transporter (DAT1) gene affects gene expression. Pharm. J. 2001, 1, 152. [Google Scholar] [CrossRef]
- Ouimet, C.; Miller, P.; Hemmings, H.; Walaas, S.I.; Greengard, P. DARPP-32, a dopamine-and adenosine 3′:5′-monophosphate-regulated phosphoprotein enriched in dopamine-innervated brain regions. III. Immunocytochemical localization. J. Neurosci. 1984, 4, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Walaas, S.I.; Greengard, P. DARPP-32, a dopamine-and adenosine 3′:5′-monophosphate-regulated phosphoprotein enriched in dopamine-innervated brain regions. I. Regional and cellular distribution in the rat brain. J. Neurosci. 1984, 4, 84–98. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Straub, R.E.; Lipska, B.K.; Verchinski, B.A.; Goldberg, T.; Callicott, J.H.; Egan, M.F.; Huffaker, S.S.; Mattay, V.S.; Kolachana, B.; et al. Genetic evidence implicating DARPP-32 in human frontostriatal structure, function, and cognition. J. Clin. Investig. 2007, 117, 672–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabresi, P.; Gubellini, P.; Centonze, D.; Picconi, B.; Bernardi, G.; Chergui, K.; Svenningsson, P.; Fienberg, A.A.; Greengard, P. Dopamine and cAMP-Regulated Phosphoprotein 32 kDa Controls Both Striatal Long-Term Depression and Long-Term Potentiation, Opposing Forms of Synaptic Plasticity. J. Neurosci. 2000, 20, 8443–8451. [Google Scholar] [CrossRef] [PubMed]
- Snyder, G.L.; Fienberg, A.A.; Huganir, R.L.; Greengard, P. A dopamine/D1 receptor/protein kinase A/dopamine-and cAMP-regulated phosphoprotein (M r 32 kDa)/protein phosphatase-1 pathway regulates dephosphorylation of the NMDA receptor. J. Neurosci. 1998, 18, 10297–10303. [Google Scholar] [CrossRef] [PubMed]
- Jay, T.M. Dopamine: A potential substrate for synaptic plasticity and memory mechanisms. Prog. Neurobiol. 2003, 69, 375–390. [Google Scholar] [CrossRef]
- Svenningsson, P.; Nishi, A.; Fisone, G.; Girault, J.-A.; Nairn, A.C.; Greengard, P. DARPP-32: An integrator of neurotransmission. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 269–296. [Google Scholar] [CrossRef]
- Kleim, J.A.; Chan, S.; Pringle, E.; Schallert, K.; Procaccio, V.; Jimenez, R.; Cramer, S.C. BDNF val66met polymorphism is associated with modified experience-dependent plasticity in human motor cortex. Nat. Neurosci. 2006, 9, 735. [Google Scholar] [CrossRef]
- Cirillo, J.; Hughes, J.; Ridding, M.; Thomas, P.Q.; Semmler, J.G. Differential modulation of motor cortex excitability in BDNF Met allele carriers following experimentally induced and use-dependent plasticity. Eur. J. Neurosci. 2012, 36, 2640–2649. [Google Scholar] [CrossRef]
- Canivet, A.; Albinet, C.T.; André, N.; Pylouster, J.; Rodríguez-Ballesteros, M.; Kitzis, A.; Audiffren, M. Effects of BDNF polymorphism and physical activity on episodic memory in the elderly: A cross sectional study. Eur. Rev. Aging Phys. Act. 2015, 12, 15. [Google Scholar] [CrossRef]
- Petzinger, G.M.; Fisher, B.E.; McEwen, S.; Beeler, J.A.; Walsh, J.P.; Jakowec, M.W. Exercise-enhanced neuroplasticity targeting motor and cognitive circuitry in Parkinson’s disease. Lancet Neurol. 2013, 12, 716–726. [Google Scholar] [CrossRef]
- Goodwin, V.A.; Richards, S.H.; Taylor, R.S.; Taylor, A.H.; Campbell, J.L. The effectiveness of exercise interventions for people with Parkinson’s disease: A systematic review and meta-analysis. Mov. Disord. 2008, 23, 631–640. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Salmoni, A.W.; Schmidt, R.A.; Walter, C.B. Knowledge of results and motor learning: A review and critical reappraisal. Psychol. Bull. 1984, 95, 355. [Google Scholar] [CrossRef]
- Grego, F.; Vallier, J.; Collardeau, M.; Rousseu, C.; Cremieux, J.; Brisswalter, J. Influence of exercise duration and hydration status on cognitive function during prolonged cycling exercise. Int. J. Sports Med. 2005, 26, 27–33. [Google Scholar] [CrossRef]
- Cian, C.; Koulmann, N.; Barraud, P.; Raphel, C.; Jimenez, C.; Melin, B. Influences of variations in body hydration on cognitive function: Effect of hyperhydration, heat stress, and exercise-induced dehydration. J. Psychophysiol. 2000, 14, 29. [Google Scholar] [CrossRef]
- Piil, J.F.; Lundbye-Jensen, J.; Christiansen, L.; Ioannou, L.; Tsoutsoubi, L.; Dallas, C.N.; Mantzios, K.; Flouris, A.D.; Nybo, L. High prevalence of hypohydration in occupations with heat stress—Perspectives for performance in combined cognitive and motor tasks. PLoS ONE 2018, 13, e0205321. [Google Scholar] [CrossRef]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Huertas, E.; Bühler, K.-M.; Echeverry-Alzate, V.; Giménez, T.; López-Moreno, J.A. C957T polymorphism of the dopamine D2 receptor gene is associated with motor learning and heart rate. Genesbrain Behav. 2012, 11, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Baetu, I.; Burns, N.R.; Urry, K.; Barbante, G.G.; Pitcher, J.B. Commonly-occurring polymorphisms in the COMT, DRD1 and DRD2 genes influence different aspects of motor sequence learning in humans. Neurobiol. Learn. Mem. 2015, 125, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Noohi, F.; Boyden, N.B.; Kwak, Y.; Humfleet, J.; Burke, D.T.; Müller, M.L.T.M.; Bohnen, N.I.; Seidler, R.D. Association of COMT val158met and DRD2 G>T genetic polymorphisms with individual differences in motor learning and performance in female young adults. J. Neurophysiol. 2014, 111, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.J.; Moustafa, A.A.; Haughey, H.M.; Curran, T.; Hutchison, K.E. Genetic triple dissociation reveals multiple roles for dopamine in reinforcement learning. Proc. Natl. Acad. Sci. USA 2007, 104, 16311–16316. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. arXiv preprint 2014, arXiv:1406.5823. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. Multi-Model Inference. Package ‘MuMIn’. Version 1.43.6. 2019-04-08. Available online: https://r-forge.r-project.org/R/?group_id=346 (accessed on 26 April 2019).


| Gene | Locus | ~ SNP | ~ Time × SNP | ~ Exercise × Time × SNP |
|---|---|---|---|---|
| DRD2 | rs1076560 | 0.16 | 0.03 | 0.04 |
| DRD2 | rs6277 | 0.18 | 0.36 | 0.77 |
| ANKK1 | rs1800497 | 0.26 | 0.02 | 0.09 |
| DRD1 | rs686 | 0.09 | 0.15 | 0.63 |
| DRD3 | rs6280 | 0.12 | 0.48 | 0.74 |
| COMT | rs4680 | 0.71 | 0.44 | 0.42 |
| PPP1R1B | rs907094 | 0.56 | 0.003 | 0.05 |
| SLC6A3 / DAT1 | rs28363170 | 0.78 | 0.13 | 0.05 |
| BDNF | rs6265 | 0.52 | 0.33 | 0.78 |
| Fitted Models | AIC | LMM R2(m) | LMM R2(c) |
|---|---|---|---|
| ~ Time × SNP1 + Time × SNP2 + … + … | 728 | 0.04 | 0.80 |
| ~ Exercise × Time | 717 | 0.10 | 0.78 |
| ~ Exercise × Time × SNP1 + Exercise × Time × SNP2 + … + … | 720 | 0.19 | 0.84 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christiansen, L.; Thomas, R.; Beck, M.M.; Pingel, J.; Andersen, J.D.; Mang, C.S.; Madsen, M.A.J.; Roig, M.; Lundbye-Jensen, J. The Beneficial Effect of Acute Exercise on Motor Memory Consolidation is Modulated by Dopaminergic Gene Profile. J. Clin. Med. 2019, 8, 578. https://doi.org/10.3390/jcm8050578
Christiansen L, Thomas R, Beck MM, Pingel J, Andersen JD, Mang CS, Madsen MAJ, Roig M, Lundbye-Jensen J. The Beneficial Effect of Acute Exercise on Motor Memory Consolidation is Modulated by Dopaminergic Gene Profile. Journal of Clinical Medicine. 2019; 8(5):578. https://doi.org/10.3390/jcm8050578
Chicago/Turabian StyleChristiansen, Lasse, Richard Thomas, Mikkel M. Beck, Jessica Pingel, Jeppe D. Andersen, Cameron S. Mang, Mads A. J. Madsen, Marc Roig, and Jesper Lundbye-Jensen. 2019. "The Beneficial Effect of Acute Exercise on Motor Memory Consolidation is Modulated by Dopaminergic Gene Profile" Journal of Clinical Medicine 8, no. 5: 578. https://doi.org/10.3390/jcm8050578
APA StyleChristiansen, L., Thomas, R., Beck, M. M., Pingel, J., Andersen, J. D., Mang, C. S., Madsen, M. A. J., Roig, M., & Lundbye-Jensen, J. (2019). The Beneficial Effect of Acute Exercise on Motor Memory Consolidation is Modulated by Dopaminergic Gene Profile. Journal of Clinical Medicine, 8(5), 578. https://doi.org/10.3390/jcm8050578