Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Evaluation of Patients
2.2. Selection of Patients for the Analysis of Cytokine and Chemokine Levels in Serum and CSF
2.3. Cytokine and Chemokine Determinations
2.4. Statistical Analyses
3. Results
3.1. Comparison of Cytokine and Chemokine Levels in Serum and CSF
3.2. Cytokine and Chemokine Levels According to White Cell Counts in Blood or CSF
3.3. Comparison of Cytokine and Chemokine Levels in CSF and Serum According to Duration of Illness
3.4. Association of Inflammatory Mediators with Severity of Acute Illness
3.5. Association of Inflammatory Mediators with Levels of IgG Antibodies against TBEV in CSF and Serum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Taba, P.; Schmutzhard, E.; Forsberg, P.; Lutsar, I.; Ljøstad, U.; Mygland, Å.; Levchenko, I.; Strle, F.; Steiner, I. EAN consensus review on prevention, diagnosis and management of tick-borne encephalitis. Eur. J. Neurol. 2017, 24, 1214–1261. [Google Scholar] [CrossRef]
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Bogovič, P.; Stupica, D.; Rojko, T.; Lotrič-Furlan, S.; Avšič-Županc, T.; Kastrin, A.; Lusa, L.; Strle, F. The long-term outcome of tick-borne encephalitis in Central Europe. Ticks Tick-Borne Dis. 2018, 9, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus—A review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Bogovic, P.; Logar, M.; Avsic-Zupanc, T.; Strle, F.; Lotric-Furlan, S. Quantitative evaluation of the severity of acute illness in adult patients with tick-borne encephalitis. Biomed. Res. Int. 2014, 2014, 841027. [Google Scholar] [CrossRef]
- Westfall, P.H.; Young, S.S. Resampling-Based Multiple Testing; Wiley: New York, NY, USA, 1993; ISBN 978-0-471-55761-6. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 24 October 2018).
- Kröger, A.; Överby, A.K. Pathogenesis of TBE with a focus on molecular mechanisms. In Tick-Borne Encephalitis, 1st ed.; Dobler, G., Erber, W., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2018; pp. 58–69. ISBN 978-981-11-1903-3. [Google Scholar]
- Blom, K.; Cuapio, A.; Sandberg, J.T.; Varnaite, R.; Michaëlsson, J.; Björkström, N.K.; Sandberg, J.K.; Klingström, J.; Lindquist, L.; Gredmark Russ, S.; et al. Cell-Mediated Immune Responses and Immunopathogenesis of Human Tick-Borne Encephalitis Virus-Infection. Front. Immunol. 2018, 9, 2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, F.X.; Berger, R.; Tuma, W.; Kunz, C. Location of immunodominant antigenic determinants on fragments of the tick-borne encephalitis virus glycoprotein: Evidence for two different mechanisms by which antibodies mediate neutralization and hemagglutination inhibition. Virology 1983, 130, 485–501. [Google Scholar] [CrossRef]
- Kreil, T.R.; Eibl, M.M. Pre- and postexposure protection by passive immunoglobulin but no enhancement of infection with a flavivirus in a mouse model. J. Virol. 1997, 71, 2921–2927. [Google Scholar]
- Niedrig, M.; Klockmann, U.; Lang, W.; Roeder, J.; Burk, S.; Modrow, S.; Pauli, G. Monoclonal antibodies directed against tick-borne encephalitis virus with neutralizing activity in vivo. Acta Virol. 1994, 38, 141–149. [Google Scholar] [PubMed]
- Phillpotts, R.J.; Stephenson, J.R.; Porterfield, J.S. Passive immunization of mice with monoclonal antibodies raised against tick-borne encephalitis virus. Brief report. Arch. Virol. 1987, 3, 295–301. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K. The molecular and antigenic structure of TBEV. In Tick-Borne Encephalitis, 1st ed.; Dobler, G., Erber, W., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2018; pp. 27–40. ISBN 978-981-11-1903-3. [Google Scholar]
- Hofmann, H.; Frisch-Niggemeyer, W.; Kunz, C. Protection of mice against tick-borne encephalitis by different classes of immunoglobulins. Infection 1978, 6, 154–157. [Google Scholar] [CrossRef]
- Palus, M.; Vojtíšková, J.; Salát, J.; Kopecký, J.; Grubhoffer, L.; Lipoldová, M.; Demant, P.; Růžek, D. Mice with different susceptibility to tick-borne encephalitis virus infection show selective neutralizing antibody response and inflammatory reaction in the central nervous system. J. Neuroinflammation 2013, 10, 77. [Google Scholar] [CrossRef]
- Dörrbecker, B.; Dobler, G.; Spiegel, M.; Hufert, F.T. Tick-borne encephalitis virus and the immune response of the mammalian host. Travel Med. Infect. Dis. 2010, 8, 213–222. [Google Scholar] [CrossRef]
- Kaiser, R.; Holzmann, H. Laboratory findings in tick-borne encephalitis—Correlation with clinical outcome. Infection 2000, 28, 78–84. [Google Scholar] [CrossRef]
- Bogovič, P.; Lotrič-Furlan, S.; Avšič-Županc, T.; Lusa, L.; Strle, F. Factors associated with severity of tick-borne encephalitis: A prospective observational study. Travel Med. Infect. Dis. 2018, 26, 25–31. [Google Scholar] [CrossRef]
- Selinger, M.; Wilkie, G.S.; Tong, L.; Gu, Q.; Schnettler, E.; Grubhoffer, L.; Kohl, A. Analysis of tick-borne encephalitis virus-induced host responses in human cells of neuronal origin and interferon-mediated protection. J. Gen. Virol. 2017, 98, 2043–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palus, M.; Bílý, T.; Elsterová, J.; Langhansová, H.; Salát, J.; Vancová, M.; Růžek, D. Infection and injury of human astrocytes by tick-borne encephalitis virus. J. Gen. Virol. 2014, 95, 411–2426. [Google Scholar] [CrossRef]
- Gelpi, E.; Preusser, M.; Laggner, U.; Garzuly, F.; Holzmann, H.; Heinz, F.X.; Budka, H. Inflammatory response in human tick-borne encephalitis: Analysis of postmortem brain tissue. J. Neurovirol. 2006, 12, 322–327. [Google Scholar] [CrossRef]
- Růžek, D.; Salát, J.; Singh, S.K.; Kopecký, J. Breakdown of the blood-brain barrier during tick-borne encephalitis in mice is not dependent on CD8+ T-cells. PLoS ONE 2011, 6, e20472. [Google Scholar] [CrossRef]
- Palus, M.; Formanová, P.; Salát, J.; Žampachová, E.; Elsterová, J.; Růžek, D. Analysis of serum levels of cytokines, chemokines, growth factors, and monoamine neurotransmitters in patients with tick-borne encephalitis: Identification of novel inflammatory markers with implications for pathogenesis. J. Med. Virol. 2015, 87, 885–892. [Google Scholar] [CrossRef]
- Gelpi, E.; Preusser, M.; Garzuly, F.; Holzmann, H.; Heinz, F.X.; Budka, H. Visualization of Central European tick-borne encephalitis infection in fatal human cases. J. Neuropathol. Exp. Neurol. 2005, 64, 506–512. [Google Scholar] [CrossRef]
- Kondrusik, M.; Pancewicz, S.; Zajkowska, J.; Hermanowska-Szpakowicz, T. Tumor necrosis factor alpha and interleukin 1-beta in serum of patients with tick-borne encephalitis. Pol. Merkur. Lekarski. 2001, 11, 26–28. (In Polish) [Google Scholar]
- Atrasheuskaya, A.V.; Fredeking, T.M.; Ignatyev, G.M. Changes in immune parameters and their correction in human cases of tick-borne encephalitis. Clin. Exp. Immunol. 2003, 131, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Lepej, S.Z.; Misić-Majerus, L.; Jeren, T.; Rode, O.D.; Remenar, A.; Sporec, V.; Vince, A. Chemokines CXCL10 and CXCL11 in the cerebrospinal fluid of patients with tick-borne encephalitis. Acta Neurol. Scand. 2007, 115, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Zajkowska, J.; Moniuszko-Malinowska, A.; Pancewicz, S.A.; Muszyńska-Mazur, A.; Kondrusik, M.; Grygorczuk, S.; Swierzbińska-Pijanowska, R.; Dunaj, J.; Czupryna, P. Evaluation of CXCL10, CXCL11, CXCL12 and CXCL13 chemokines in serum and cerebrospinal fluid in patients with tick borne encephalitis (TBE). Adv. Med. Sci. 2011, 56, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Cerar, T.; Ogrinc, K.; Lotric-Furlan, S.; Kobal, J.; Levicnik-Stezinar, S.; Strle, F.; Ruzić-Sabljic, E. Diagnostic value of cytokines and chemokines in Lyme neuroborreliosis. Clin. Vaccine Immunol. 2013, 20, 1578–1584. [Google Scholar] [CrossRef]
- Pietikäinen, A.; Maksimow, M.; Kauko, T.; Hurme, S.; Salmi, M.; Hytönen, J. Cerebrospinal fluid cytokines in Lyme neuroborreliosis. J. Neuroinflammation 2016, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Markowicz, M.; Schötta, A.M.; Kundi, M.; Bogovič, P.; Ogrinc, K.; Strle, F.; Stanek, G. CXCL13 concentrations in cerebrospinal fluid of patients with Lyme neuroborreliosis and other neurological disorders determined by Luminex and ELISA. Ticks Tick-Borne Dis. 2018, 9, 1137–1142. [Google Scholar] [CrossRef]
- Grygorczuk, S.; Parczewski, M.; Świerzbińska, R.; Czupryna, P.; Moniuszko, A.; Dunaj, J.; Kondrusik, M.; Pancewicz, S. The increased concentration of macrophage migration inhibitory factor in serum and cerebrospinal fluid of patients with tick-borne encephalitis. J. Neuroinflammation 2017, 14, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygorczuk, S.; Świerzbińska, R.; Kondrusik, M.; Dunaj, J.; Czupryna, P.; Moniuszko, A.; Siemieniako, A.; Pancewicz, S. The intrathecal expression and pathogenetic role of Th17 cytokines and CXCR2-binding chemokines in tick-borne encephalitis. J. Neuroinflammation 2018, 15, 115. [Google Scholar] [CrossRef]
- Günther, G.; Haglund, M.; Lindquist, L.; Forsgren, M.; Andersson, J.; Andersson, B.; Sköldenberg, B. Tick-borne encephalitis is associated with low levels of interleukin-10 in cerebrospinal fluid. Infect. Ecol. Epidemiol. 2011, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettelli, E.; Korn, T.; Oukka, M.; Kuchroo, V.K. Induction and effector functions of T(H)17 cells. Nature 2008, 453, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Strle, K.; Sulka, K.B.; Pianta, A.; Crowley, J.T.; Arvikar, S.L.; Anselmo, A.; Sadreyev, R.; Steere, A.C. T-Helper 17 Cell Cytokine Responses in Lyme Disease Correlate With Borrelia burgdorferi Antibodies During Early Infection and With Autoantibodies Late in the Illness in Patients With Antibiotic-Refractory Lyme Arthritis. Clin. Infect. Dis. 2017, 64, 930–938. [Google Scholar] [CrossRef] [PubMed]
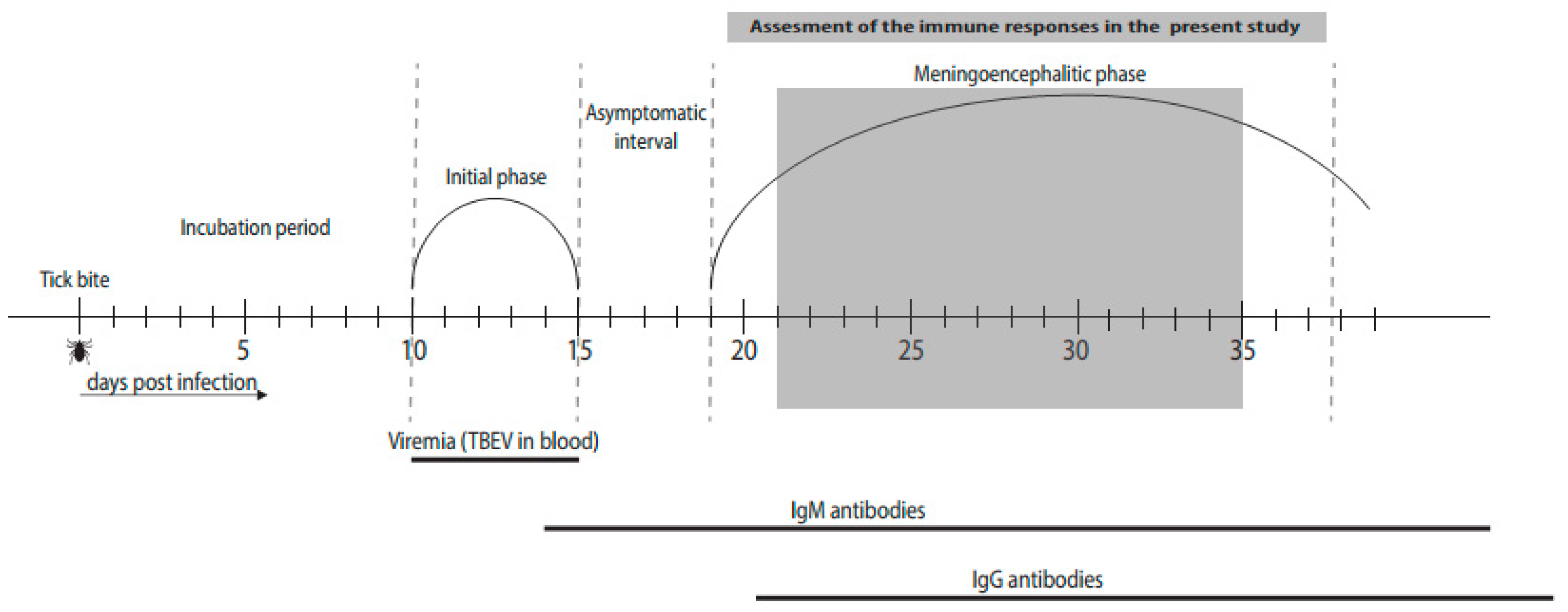



{kind=link}
{kind=link}
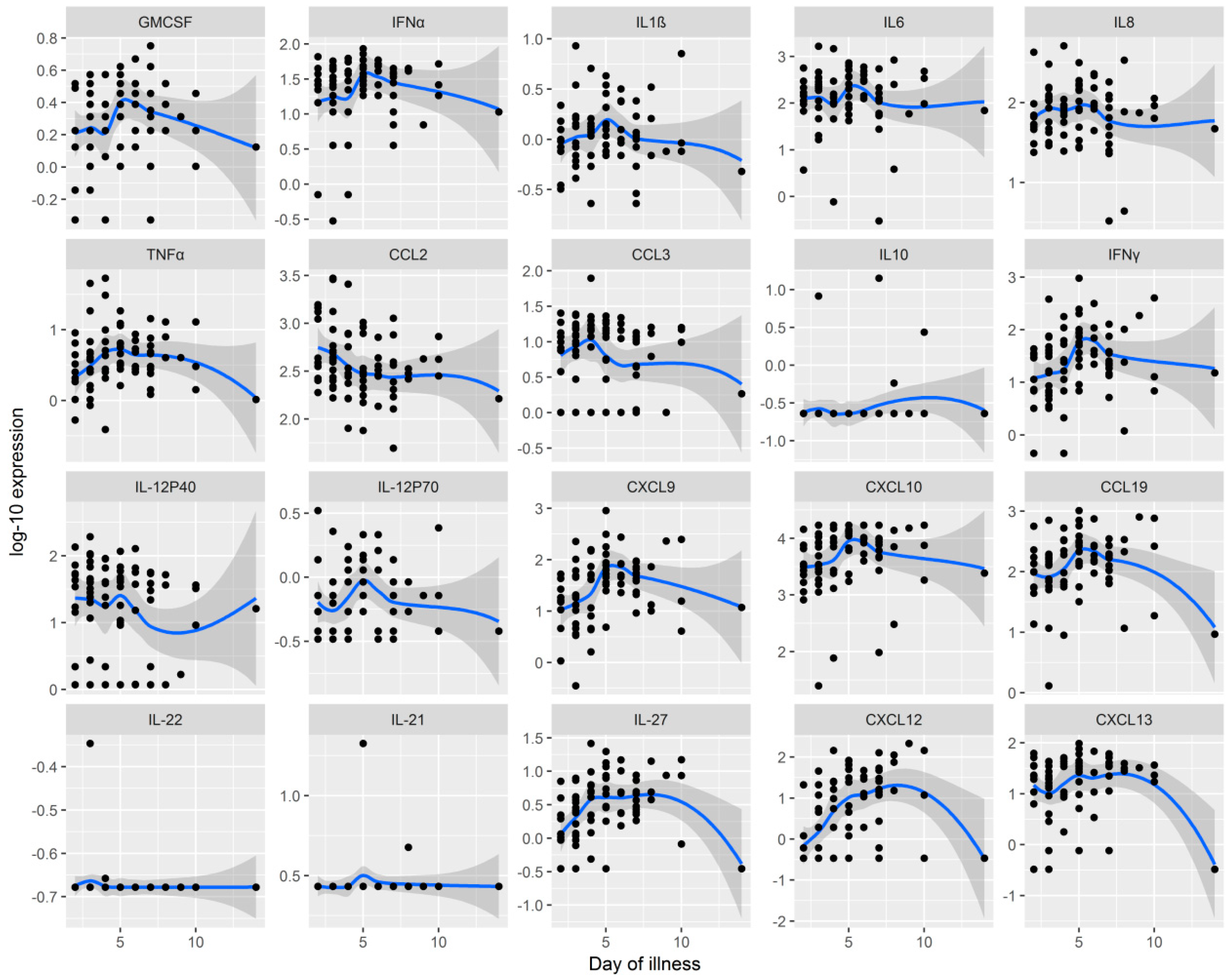{kind=link}
| Characteristic | Number (%, 95% CI) or Median (IQR) |
|---|---|
| Male sex | 40 (49.4; 38.1–60.7) |
| Age (years) | 56 (43–63) |
| Males | 58 (43–62.5) |
| Females | 55 (45–63) |
| Underlying illnesses | 39 (48.1; 36.9–59.5) |
| Monophasic course of illness | 35 (43.2; 32.2–54.7) |
| Clinical presentation | |
| Meningitis | 33 (40.7; 30.0–52.2) |
| Meningoencephalitis | 40 (49.4; 38.1–60.7) |
| Meningoencephalomyelitis | 8 (9.9; 4.4–18.5) |
| Severity of illness | |
| According to clinical assessment | |
| -Mild (meningitis) | 33 (40.7; 30.0–52.2) |
| -Severe (meningoencephalitis or meningoencephalomyelitis) | 48 (59.3; 47.8–70.1) |
| According to severity score | 12 (4–23) |
| Treatment in intensive care unit | 7 (8.6; 3.6–17.0) |
| Duration (days) | 5 (3.5–9) |
| Artificial ventilation: number; duration (days) | 2 (28.6; 3.7–71.0); 6 (5–7) |
| Duration of illness before CSF and blood sample obtained (days) a | 5 (3–6) b |
| Blood leukocyte count (× 109 cells/L) | 9.9 (8.2–12.4) c |
| Neutrophils (× 109 cells/L) | 7.7 (6.0–9.8) |
| Lymphocytes (× 109 cells/L) | 1.4 (1.1–1.8) |
| Monocytes (× 109 cells/L) | 0.7 (0.5–0.9) |
| CSF leukocyte count (× 106 cells/L) | 76 (37–134) |
| Neutrophils (× 106 cells/L) | 23 (11–48) |
| Lymphocytes (× 106 cells/L) | 43 (20–76) |
| Monocytes (× 106 cells/L) | 2 (0–5) |
| CSF protein concentration (g/L) | 0.70 (0.53–0.91) |
| Elevated (> 0.45 g/L) | 67 (82.7; 72.7–90.2) |
| CSF glucose concentration (mmol/L) | 2.9 (2.6–3.3) c |
| CSFglu/Sglu < 0.33 | 0/79 (0; 0–4.6) |
| Albumin quotient (× 10−3) d | 10.56 (7.98–12.92) b |
| IgG quotient (× 10−3) d | 5.18 (3.98–6.62) b |
| Concomitant Lyme neuroborreliosis e | 3/78 (3.8; 0.8–10.8) |
| Positive B. burgdorferi sensu lato IgG antibodies f | 7/78 (9.0; 3.7–17.6) |
| Cytokine/Chemokine | Concentrations (pg/mL) a Median (IQR) | pb Value | pc Value Adjusted | |
|---|---|---|---|---|
| CSF | Serum | |||
| Innate | ||||
| GMCSF | 2 (1.3–2.9) | 0.87 (0.31–1.68) | 1.8 × 10−9 | 0.0001 |
| IFNα | 310 (18–45) | 0.71 (0.30–6.2) | 1.3 × 10−11 | 0.0001 |
| IL-1β | 1 (0.8–1.6) | 0.32 (0.23–0.69) | 4.2 × 10−5 | 0.0003 |
| IL-6 | 154 (95–319) | 0.30 (0.30–0.76) | 9.0 × 10−15 | 0.0001 |
| IL-8 | 85 (49–117) | 1.9 (0.62–15.1) | 5.3 × 10−11 | 0.0001 |
| TNFα | 4 (2.1–6.4) | 6.4 (4.4–16.4) | 3.0 × 10−10 | 0.0001 |
| CCL2 | 325 (214–585) | 205 (158–259) | 6.1 × 10−13 | 0.0001 |
| CCL3 | 12 (3.4–15.4) | 1.01 (1.0–8.8) | 0.21 | 0.95 |
| IL-10 | 0.2 (0.2–0.2) | 0.2 (0.2 –0.2) | 1 | 1 |
| Th1 | ||||
| IFNγ | 28 (11–62) | 1.1 (0.45–2.42) | 1.7 × 10−14 | 0.0001 |
| IL-12P40 | 37 (9–54) | 1.2 (1.2–1.2) | 1.1 × 10−12 | 0.0001 |
| IL-12P70 | 0.7 (0.4–0.9) | 0.38 (0.33–0.92) | 0.0956 | 0.72 |
| CXCL9 | 39 (12–72) | 22 (12–41) | 0.0002 | 0.0013 |
| CXCL10 | 7082 (2730–10451) | 91 (60–147) | 7.1 × 10−15 | 0.0001 |
| CCL19 | 145 (67–259) | 24 (11–31) | 2.5 × 10−13 | 0.0001 |
| Th17 | ||||
| IL-17F d | 320 (320–320) | 320 (320–320) | 0.0059 | 0.0711 |
| IL-17A d | 0.31 (0.31–0.31) | 0.31 (0.31–0.31) | 0.0011 | 0.0115 |
| IL-22 | 0.21 (0.21–0.21) | 0.22 (0.21–0.45) | 3.8 × 10−7 | 0.0001 |
| IL-21 | 2.7 (2.7–2.7) | 5.71 (2.7–14.11) | 4.7 × 10−11 | 0.0001 |
| IL-23 d | 250 (250–250) | 250 (250–250) | 0.0017 | 0.0210 |
| IL-25 d | 320 (320–320) | 320 (320–320) | 0.0591 | 0.53 |
| IL-27 | 3000 (1500–6100) | 1500 (900–2100) | 0.10 | 0.75 |
| B cell | ||||
| CXCL12 | 5.5 (0.34–26) | 31 (19–53) | 1.3 × 10−9 | 0.0001 |
| CXCL13 | 26 (11–39) | 26 (19–36) | 0.59 | 1 |
| Immune Response Cytokine/Chemokine | Correlation with Leukocyte Count | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CSF Leucocytes | Serum Leucocytes | |||||||||||||||||
| Neutrophils | Lymphocytes | Monocytes | Neutrophils | Lymphocytes | Monocytes | |||||||||||||
| Rho a | p Value b | p Value c | Rho a | p Value b | p Value c | Rho a | p Value b | p Value c | Rho a | p Value b | p Value c | Rho a | p Value b | p Value c | Rho a | p Value b | p Value c | |
| Innate | ||||||||||||||||||
| GMCSF | 0.2511 | 0.02 | 0.88 | 0.2059 | 0.0652 | 0.99 | 0.1764 | 0.12 | 1 | −0.1838 | 0.10 | 1 | −0.0896 | 0.43 | 1 | −0.1911 | 0.0896 | 0.99 |
| IFNα | 0.2003 | 0.0730 | 0.99 | 0.3575 | 0.0010 | 0.11 | 0.2576 | 0.0202 | 0.84 | 0.0040 | 0.97 | 1 | −0.0060 | 0.96 | 1 | 0.0682 | 0.55 | 1 |
| IL-1β | 0.4768 | <0.0001 | 0.0008 | 0.3007 | 0.0064 | 0.45 | 0.1801 | 0.11 | 1 | 0.0313 | 0.78 | 1 | −0.0386 | 0.73 | 1 | −0.0645 | 0.57 | 1 |
| IL-6 | 0.5435 | <0.0001 | 0.0001 | 0.1948 | 0.0814 | 0.99 | 0.1496 | 0.18 | 1 | 0.1231 | 0.28 | 1 | −0.1520 | 0.18 | 1 | −0.0391 | 0.73 | 1 |
| IL-8 | 0.5858 | <0.0001 | 0.0001 | 0.1346 | 0.23 | 1 | 0.0279 | 0.80 | 1 | 0.0481 | 0.67 | 1 | 0.0675 | 0.55 | 1 | 0.0218 | 0.85 | 1 |
| TNFα | 0.2669 | 0.0160 | 0.76 | 0.5344 | <0.0001 | 0.0001 | 0.2009 | 0.0721 | 0.99 | −0.0903 | 0.43 | 1 | −0.0338 | 0.77 | 1 | −0.0232 | 0.84 | 1 |
| CCL2 | 0.3096 | 0.0049 | 0.37 | −0.1752 | 0.12 | 1 | 0.0016 | 0.99 | 1 | −0.2827 | 0.0110 | 0.63 | −0.1539 | 0.17 | 1 | −0.3189 | 0.0039 | 0.32 |
| CCL3 | 0.1486 | 0.19 | 1 | 0.1244 | 0.27 | 1 | 0.2016 | 0.0711 | 0.99 | −0.0532 | 0.64 | 1 | −0.0740 | 0.51 | 1 | 0.0333 | 0.77 | 1 |
| IL-10 | 0.1308 | 0.24 | 1 | 0.1025 | 0.36 | 1 | −0.1588 | 0.16 | 1 | −0.1134 | 0.32 | 1 | −0.0579 | 0.61 | 1 | −0.0989 | 0.38 | 1 |
| Th1 | ||||||||||||||||||
| IFNγ | 0.2344 | 0.0352 | 0.96 | 0.5011 | <0.0001 | 0.0002 | 0.2369 | 0.0333 | 0.95 | −0.0671 | 0.55 | 1 | −0.0518 | 0.65 | 1 | −0.1119 | 0.32 | 1 |
| IL-12P40 | 0.2011 | 0.0719 | 0.99 | 0.0345 | 0.76 | 1 | 0.2047 | 0.0668 | 0.99 | −0.1502 | 0.18 | 1 | −0.2486 | 0.0262 | 0.91 | −0.1120 | 0.32 | 1 |
| IL-12P70 | 0.2195 | 0.0490 | 0.99 | 0.3109 | 0.0047 | 0.36 | 0.2907 | 0.0085 | 0.54 | 0.0095 | 0.93 | 1 | −0.1261 | 0.26 | 1 | −0.0548 | 0.63 | 1 |
| CXCL9 | 0.0573 | 0.61 | 1 | 0.4607 | <0.0001 | 0.0018 | 0.2011 | 0.0719 | 0.99 | −0.2458 | 0.0280 | 0.92 | −0.2009 | 0.0739 | 0.99 | 0.0720 | 0.53 | 1 |
| CXCL10 | 0.1951 | 0.0809 | 0.99 | 0.5968 | <0.0001 | 0.0001 | 0.385 | 0.0004 | 0.0422 | −0.2878 | 0.0096 | 0.59 | −0.1808 | 0.11 | 1 | −0.0943 | 0.41 | 1 |
| CCL19 | 0.1533 | 0.17 | 1 | 0.4759 | <0.0001 | 0.0008 | 0.3084 | 0.0051 | 0.38 | −0.0325 | 0.77 | 1 | 0.0993 | 0.38 | 1 | 0.0163 | 0.89 | 1 |
| Th17 | ||||||||||||||||||
| IL-17F | NA | NA | NA | NA | NA | NA | NA | NA | NA | −0.0994 | 0.38 | 1 | −0.0194 | 0.86 | 1 | −0.1770 | 0.12 | 1 |
| IL-17A | NA | NA | NA | NA | NA | NA | NA | NA | NA | 0.0080 | 0.94 | 1 | 0.0802 | 0.48 | 1 | −0.0079 | 0.94 | 1 |
| IL-22 | 0.2278 | 0.0408 | 0.98 | 0.0228 | 0.84 | 1 | −0.1850 | 0.0982 | 1 | −0.0333 | 0.77 | 1 | 0.0484 | 0.67 | 1 | 0.0969 | 0.39 | 1 |
| IL-21 | −0.0937 | 0.41 | 1 | 0.13 | 0.25 | 1 | −0.1850 | 0.0982 | 1 | 0.0154 | 0.89 | 1 | −0.1430 | 0.21 | 1 | 0.0438 | 0.70 | 1 |
| IL-23 | NA | NA | NA | NA | NA | NA | NA | NA | NA | −0.0372 | 0.74 | 1 | 0.1385 | 0.22 | 1 | −0.0618 | 0.59 | 1 |
| IL-25 | NA | NA | NA | NA | NA | NA | NA | NA | NA | −0.0365 | 0.75 | 1 | −0.0726 | 0.52 | 1 | -0.2147 | 0.0558 | 0.99 |
| IL-27 | 0.2642 | 0.0172 | 0.79 | 0.4776 | <0.0001 | 0.0008 | 0.1193 | 0.29 | 1 | −0.0429 | 0.71 | 1 | −0.2505 | 0.0250 | 0.90 | −0.0811 | 0.47 | 1 |
| B cell | ||||||||||||||||||
| CXCL12 | −0.0106 | 0.93 | 1 | 0.5593 | <0.0001 | 0.0001 | 0.1604 | 0.15 | 1 | 0.0936 | 0.41 | 1 | 0.1521 | 0.18 | 1 | −0.1018 | 0.37 | 1 |
| CXCL13 | −0.1671 | 0.14 | 1 | 0.1607 | 0.15 | 1 | 0.1365 | 0.22 | 1 | −0.1137 | 0.32 | 1 | 0.0316 | 0.78 | 1 | 0.0103 | 0.93 | 1 |
| Immune Response Cytokine/Chemokine | Correlation with Severity * of Tick-Borne Encephalitis | |||||
|---|---|---|---|---|---|---|
| CSF | Serum | |||||
| Rho a | Unadjusted p Value b | Adjusted p Value c | Rho a | Unadjusted p Value b | Adjusted p Value c | |
| Innate | ||||||
| GMCSF | 0.3402 | 0.0019 | 0.0570 | 0.3212 | 0.0035 | 0.0989 |
| IFNα | 0.2376 | 0.0327 | 0.62 | 0.2122 | 0.0572 | 0.82 |
| IL-1β | 0.3703 | 0.0007 | 0.0207 | 0.1461 | 0.19 | 0.99 |
| IL-6 | 0.1368 | 0.22 | 0.99 | 0.1431 | 0.20 | 0.99 |
| IL-8 | 0.1026 | 0.36 | 1 | 0.1159 | 0.30 | 1 |
| TNFα | 0.2507 | 0.0240 | 0.50 | 0.0518 | 0.65 | 1 |
| CCL2 | 0.0049 | 0.97 | 1 | 0.215 | 0.0539 | 0.80 |
| CCL3 | 0.363 | 0.0009 | 0.0273 | 0.0846 | 0.45 | 1 |
| IL-10 | 0.082 | 0.47 | 1 | −0.0834 | 0.46 | 1 |
| Th1 | ||||||
| IFNγ | 0.0572 | 0.61 | 1 | 0.1581 | 0.16 | 0.99 |
| IL-12P40 | 0.3871 | 0.0004 | 0.0128 | −0.009 | 0.94 | 1 |
| IL-12P70 | 0.3591 | 0.0010 | 0.0319 | 0.2118 | 0.0577 | 0.83 |
| CXCL9 | 0.1887 | 0.0916 | 0.94 | 0.4345 | 0.0001 | 0.0022 |
| CXCL10 | 0.04 | 0.72 | 1 | 0.3516 | 0.0013 | 0.0404 |
| CCL19 | 0.0316 | 0.78 | 1 | 0.0518 | 0.65 | 1 |
| Th17 | ||||||
| IL-17F | NA | NA | NA | −0.0218 | 0.85 | 1 |
| IL-17A | NA | NA | NA | −0.0112 | 0.92 | 1 |
| IL-22 | 0.0664 | 0.56 | 1 | −0.1627 | 0.15 | 0.99 |
| IL-21 | 0.0087 | 0.94 | 1 | −0.1155 | 0.30 | 1 |
| IL-23 | NA | NA | NA | 0.0217 | 0.85 | 1 |
| IL-25 | NA | NA | NA | 0.0177 | 0.88 | 1 |
| IL-27 | −0.0481 | 0.67 | 1 | −0.0087 | 0.94 | 1 |
| B cell | ||||||
| CXCL12 | −0.1087 | 0.33 | 1 | −0.1067 | 0.34 | 1 |
| CXCL13 | −0.0879 | 0.44 | 1 | −0.132 | 0.24 | 0.99 |
| Immune Response Cytokine/Chemokine | Correlation with the Levels of Igg Antibodies against Tick-Borne Encephalitis Virus | |||||
|---|---|---|---|---|---|---|
| CSF | Serum | |||||
| Rho a | Unadjusted p Value b | Adjusted p Value c | Rho a | Unadjusted p Value b | Adjusted p Value c | |
| Innate | ||||||
| GMCSF | −0.13 | 0.25 | 1 | −0.1914 | 0.0869 | 0.94 |
| IFNα | 0.0072 | 0.95 | 1 | −0.1998 | 0.0736 | 0.91 |
| IL-1β | −0.1316 | 0.24 | 1 | −0.2310 | 0.0380 | 0.70 |
| IL-6 | −0.0627 | 0.58 | 1 | −0.2476 | 0.0258 | 0.54 |
| IL-8 | −0.0253 | 0.82 | 1 | −0.2409 | 0.0302 | 0.62 |
| TNFα | 0.1695 | 0.13 | 0.99 | −0.2561 | 0.0210 | 0.47 |
| CCL2 | −0.2938 | 0.0078 | 0.22 | −0.1659 | 0.14 | 0.99 |
| CCL3 | −0.3074 | 0.0052 | 0.15 | −0.3493 | 0.0014 | 0.0447 |
| IL-10 | 0.1548 | 0.17 | 0.99 | 0.0238 | 0.83 | 1 |
| Th1 | ||||||
| IFNγ | 0.2535 | 0.0224 | 0.49 | 0.0240 | 0.83 | 1 |
| IL-12P40 | −0.339 | 0.0020 | 0.0606 | −0.0362 | 0.75 | 1 |
| IL-12P70 | −0.0756 | 0.50 | 1 | −0.1534 | 0.17 | 0.9974 |
| CXCL9 | 0.2650 | 0.0168 | 0.40 | 0.0093 | 0.93 | 1 |
| CXCL10 | 0.2698 | 0.0149 | 0.36 | −0.2431 | 0.0288 | 0.59 |
| CCL19 | 0.2342 | 0.0353 | 0.68 | 0.2432 | 0.0287 | 0.59 |
| Th17 | ||||||
| IL-17F | NA | NA | NA | −0.1104 | 0.33 | 1 |
| IL-17A | NA | NA | NA | −0.1576 | 0.16 | 0.9957 |
| IL-22 | −0.1712 | 0.13 | 0.98 | −0.0893 | 0.43 | 1 |
| IL-21 | 0.1292 | 0.25 | 0.99 | −0.1909 | 0.0878 | 0.94 |
| IL-23 | NA | NA | NA | −0.1310 | 0.24 | 1 |
| IL-25 | NA | NA | NA | −0.0364 | 0.75 | 1 |
| IL-27 | 0.3563 | 0.0011 | 0.0369 | −0.0941 | 0.40 | 1 |
| B cell | ||||||
| CXCL12 | 0.4462 | 0.0000 | 0.0013 | −0.0805 | 0.47 | 1 |
| CXCL13 | 0.1867 | 0.0952 | 0.95 | −0.0528 | 0.64 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogovič, P.; Lusa, L.; Korva, M.; Pavletič, M.; Resman Rus, K.; Lotrič-Furlan, S.; Avšič-Županc, T.; Strle, K.; Strle, F. Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis. J. Clin. Med. 2019, 8, 731. https://doi.org/10.3390/jcm8050731
Bogovič P, Lusa L, Korva M, Pavletič M, Resman Rus K, Lotrič-Furlan S, Avšič-Županc T, Strle K, Strle F. Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis. Journal of Clinical Medicine. 2019; 8(5):731. https://doi.org/10.3390/jcm8050731
Chicago/Turabian StyleBogovič, Petra, Lara Lusa, Miša Korva, Miša Pavletič, Katarina Resman Rus, Stanka Lotrič-Furlan, Tatjana Avšič-Županc, Klemen Strle, and Franc Strle. 2019. "Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis" Journal of Clinical Medicine 8, no. 5: 731. https://doi.org/10.3390/jcm8050731
APA StyleBogovič, P., Lusa, L., Korva, M., Pavletič, M., Resman Rus, K., Lotrič-Furlan, S., Avšič-Županc, T., Strle, K., & Strle, F. (2019). Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis. Journal of Clinical Medicine, 8(5), 731. https://doi.org/10.3390/jcm8050731