1. Introduction
Obstructive sleep apnea (OSA) is one of the most common forms of sleep-disordered breathing, which is characterized by repetitive narrowing and opening of the upper airway during sleep, resulting in a cyclical breathing pattern accompanied by hypoxia and reoxygenation (intermittent hypoxia), and brief awakenings from sleep (sleep fragmentation) [
1]. Intermittent hypoxia and sleep fragmentation can induce chemical and structural neuronal injury, subsequently leading to prefrontal cortex dysfunction [
2,
3,
4]. As such, OSA has been reported to affect multiple domains of frontal cognitive function, especially attention, vigilance, and executive function [
5].
Action-monitoring function, an important domain of frontal cognitive function, plays a major role in adaptive behavior for early response-conflict monitoring, error detection, and then late behavioral adjustments. Therefore, action-monitoring function is important for humans to recognize errors, adjust behaviors, and optimize subsequent performances [
6,
7]. Action-monitoring behaviors are mediated by the anterior cingulate cortex (ACC) in the medial frontal lobe [
8]. Using the modified Flanker task, action-monitoring dysfunction has been reported in patients with OSA [
9]. Despite our previous reports suggesting behavioral action-monitoring dysfunction in patients with OSA, relatively less is known about the associated cerebral substrate underlying this dysfunction in people with different severities of OSA. An additional investigation using both neuroelectrophysiological response and functional connectivity is necessary on order to classify the pathological progression of action-monitoring dysfunction in patients with OSA.
Electrophysiological study is an informative and dynamic method for evaluating the neural activities underlying behaviors. Error-related negativity (ERN) and error positivity (Pe), two components of event-related potential (ERP) proposed by Falkenstein et al., are believed to reflect action-monitoring function and error processing [
10]. ERN, a negative electroencephalogram (EEG) deflection that is the largest over the frontal and central midline electrodes and generally peaks at 100 milliseconds (ms) after errors, is believed to be generated in the caudal ACC [
11]. Functionally, ERN is reportedly an early process of response-conflict monitoring [
12,
13,
14]. Following ERN, a positive deflection, Pe, generated from the rostral ACC and the parietal cortex, is observed with a centroparietal maximum at approximately 300–350 ms after errors [
11]. Pe is believed to reflect an independent process distinct from ERN, and it is associated with conscious error processing and late remedial action [
15,
16,
17]. Experimentally extended wakefulness and sleep deprivation can result in decreased amplitudes of ERN and Pe, indicating impaired action-monitoring function [
18,
19,
20,
21].
Although electrophysiological study offers excellent temporal resolution, the spatial resolution achieved using the technique is not satisfactory. Functional magnetic resonance imaging (fMRI) is another practical tool with excellent spatial resolution that can be used to evaluate changes in the cerebral response and the functional connectivity underlying behaviors. Studies using fMRI and cognitive tasks pertaining to attention, mismatch, and response inhibition have shown that patients with OSA exhibited altered cerebral activation in the frontal region and the ACC [
22,
23,
24]. Among fMRI techniques, resting-state fMRI provides a convenient and fundamental assessment tool to measure brain activity [
25].
However, to the best of our knowledge, no study has yet investigated the differences in neural activities and functional connectivity underlying action-monitoring dysfunction in patients with different severities of OSA using electrophysiology (i.e., ERN and Pe) and neuroimaging (i.e., fMRI). Considering that patients with OSA exhibit behavioral action-monitoring dysfunction, we hypothesized that patients with different severities of OSA would exhibit different patterns of neural activities and functional connectivity underlying action-monitoring dysfunction. We tested our hypothesis using the modified Flanker task to evaluate the action-monitoring function and to obtain electrophysiological measurements, namely, ERN and Pe, and fMRI, for examining neural activities and the functional connectivity in cerebral regions related to action-monitoring in patients with OSA of different severities.
2. Materials and Methods
2.1. Participants
A cross-sectional case–control design was adopted in this study. A total of 27 patients with moderate-to-severe OSA (apnea–hypopnea index (AHI) ≥ 15 counts/h) were recruited from Kaohsiung Medical University Hospital in Southern Taiwan, and 14 healthy controls were recruited from the general community between 2012 and 2015. All study participants were categorized into the following three groups based on their AHI values: Control (
Cont) participants with AHI < 5 counts/h, participants with moderate OSA (
mOSA) with 15 counts/h ≤ AHI ≤ 30 counts/h, and participants with severe OSA (
sOSA) with AHI > 30 counts/h. The AHI ranges of the three groups were defined according to the American Academy of Sleep Medicine [
26].
After including the participants in the study, each participant received a comprehensive medical evaluation, including cognitive assessment conducted using the Chinese version of the Cognitive Abilities Screening Instrument (CASI) [
27], and a complete overnight polysomnography (PSG) examination. All participants were qualified with CASI cognitive evaluation for normality with a total score of at least above 85. To avoid inclusion of participants with potential OSA in the
Cont group, all
Cont individuals with AHI ≥ 5 counts/h were excluded. When participants had completed the overnight PSG examination, they underwent the modified Flanker task to evaluate their action-monitoring function, and meanwhile, ERPs were acquired using EEG. Then, a resting-state fMRI was performed for each participant with instruction to lie in the MRI scanner without any specific thoughts and to be awake.
We excluded participants with a history, or clinical evidence, of neurological or psychiatric disease, those with major organ failure, those receiving medical therapy that might affect their cognitive function (e.g., a prescription of benzodiazepines, anticholinergics, antipsychotics, or other potential sedatives), and those with CASI scores lower than the cutoff value (adjusted for age and level of education) [
28]. In addition, all participants were right-handed users and claimed that they have not had any medical experience due to of sleep-disordered breathing. None of them consumed alcohol, caffeine, nicotine, or any cognition-enhancing medications on the day of testing. This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Kaohsiung Medical University Hospital (KMUHIRB-2012-03-09(II)). Participation was voluntary, and all participants provided written informed consent.
2.2. Modified Flanker Task and Assessments of Error-Related ERPs
The modified Flanker task was performed to elicit ERPs in this study. Briefly, five visual stimuli were presented in the middle of a computer monitor, and participants were instructed to indicate the direction of a central target stimulus. In congruent trials, the nontarget stimuli corresponded to the same direction as the target (<<<<< or >>>>>), whereas in incongruent trials, the nontarget stimuli corresponded to the direction opposite to that of the target (<<><< or >><>>). Finally, visual feedback was provided to all participants based on the accuracy of their responses. The formal task involved five sessions, each comprising 100 trials. The order of stimuli randomly differed in each session.
During the task, the participants responded to congruent and incongruent trials, which were presented randomly in a trial ratio of 1:4. Meanwhile, electrophysiological signals were recorded by continuous EEG, which was performed using Ag/AgCl electrodes placed on six locations of the central scalp (Fz, FCz, Cz, CPz, Pz, and Oz) based on the international 10–20 system montage; all electrodes were referenced to linked earlobes. Electrode impedance was maintained at lower than 5 kΩ. The EEG signals were filtered with a 0.01–40 Hz bandpass filter using a SynAmps amplifier (NeuroScan Inc., Charlotte, NC, USA). Continuous EEG recordings acquired at a sampling rate of 256 Hz were stored for further offline analysis.
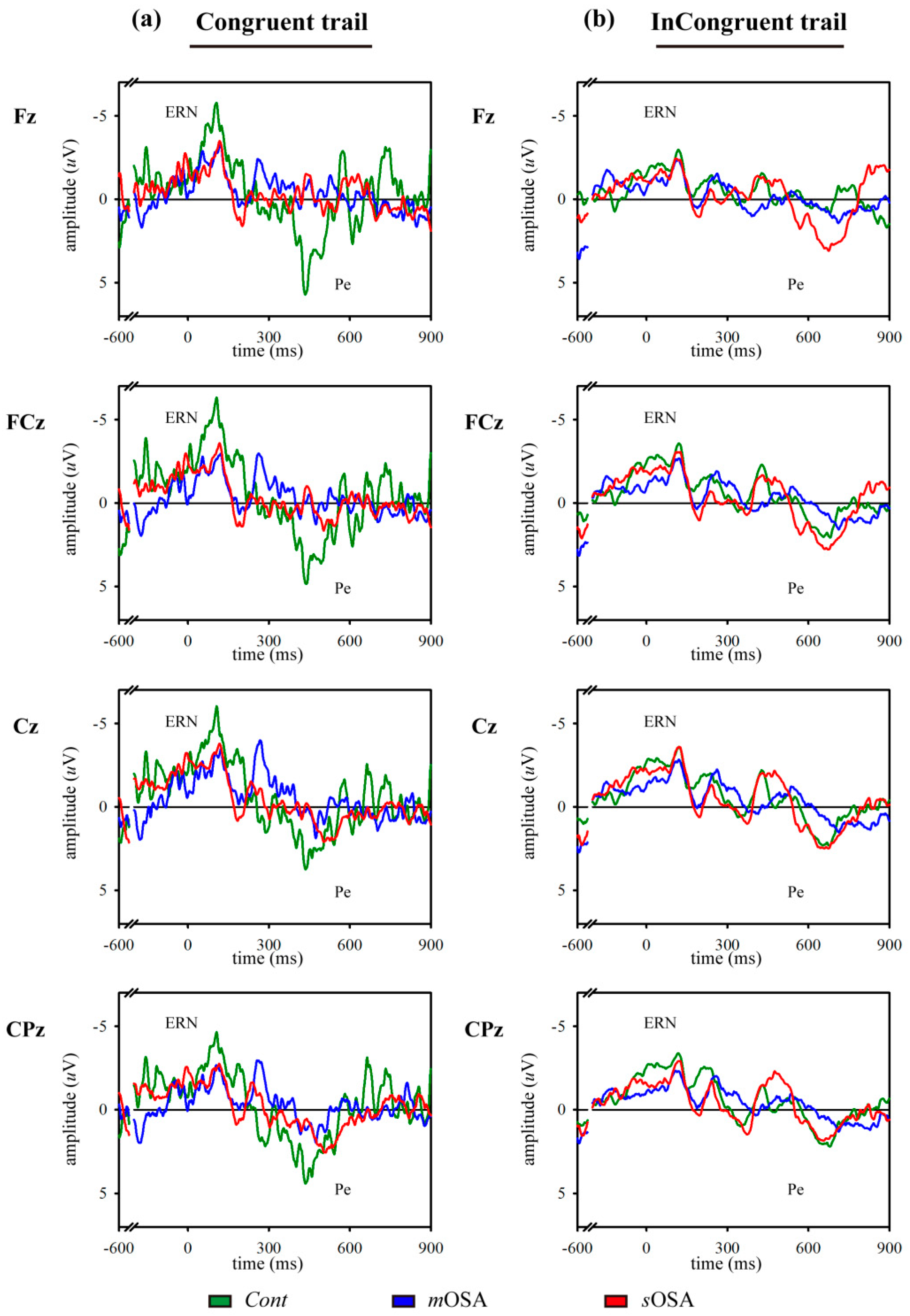
ERPs obtained under the congruent and incongruent conditions were extracted from the continuous EEG signals and averaged separately. Response-locked ERPs, namely, ERN and Pe, were detected when participants responded incorrectly to the trials corresponding to the congruent and incongruent conditions. When a participant responded incorrectly, the ERN response was detected as the first negative peak, following errors in the time window of 50–200 ms, and the Pe response was observed as the positive component following ERN, with a latency of 200–700 ms after the error event.
2.3. Polysomnography
A standard overnight full-channel PSG using two validated machines (Nicolet Ultrasom, Madison, WI, USA and Respironics Alice 5, Philips, Eindhoven, Netherlands) was applied to every participant. Physiological parameters, including pulse oximetry, an EEG, an electrooculogram, an electromyogram, an electrocardiogram, nasal and oral air flow measurements, and thoracic and abdominal respiratory inductive plethysmography, were recorded. Sleep stages and sleep-related respiratory events were scored based on the criteria of the American Academy of Sleep Medicine [
26]. Respiratory parameters, including the AHI, oxygen desaturation index, arousal index, percentage of time with blood oxygen saturation (SpO
2) < 90% (SpO
2 value < 90%), and the lowest SpO
2, were recorded. Hypopnea was defined as a drop in peak signal excursions by ≥30% of the pre-event baseline when nasal pressure is applied for ≥10 seconds with ≥4% oxygen desaturation or a drop in peak signal excursions by ≥50% for ≥10 seconds with ≥3% oxygen desaturation or associated arousal.
2.4. fMRI Scan Protocol
A 6 min fMRI scan was performed to acquire the functional response of the brain in the resting state. Functional imaging covering the entire brain was acquired using a 3-Tesla GE scanner (Signa HDx, GE, Milwaukee, WI, United States) with an echo planar imaging (EPI)-based blood oxygen level-dependent sequence in the axial plane with the following parameters: Repetition time (2000 ms), echo time (35 ms), flip angle (80°), field of view (220 × 220 mm2), acquisition matrix (64 × 64), slice thickness (3.4 mm), slice number (28 slices/volume), and volume number (180 time points). As neuron firing is induced by endogenous or exogenous stimuli, the intensity of fMRI signals corresponds to the induced magnetic variation due to the flow of deoxyhemoglobin in blood. Furthermore, high-resolution T1-weighted images were collected along the anterior commissure–posterior commissure line using a three-dimensional spoiled gradient recalled sequence (3D-SPGR) with the following parameters: Repetition time (2.5 ms), echo time (4.38 ms), flip angle (8°), field of view (240 × 240 mm2), acquisition matrix (256 × 256), slice thickness (1 mm), and slice number (124 slices/volume). This T1 imaging was used as the anatomical template underlaid by functional imaging.
All participants were instructed to lie still with eyes closed, think nonspecific thoughts, and not fall asleep in the scanner. Earplugs were used to isolate the participants from the operating noise of the fMRI scanner. After the scan, the participants reported that they had not fallen asleep during scanning.
2.5. fMRI Data Analysis
All functional data were preprocessed using the Data Processing and Analysis of Brain Imaging (DPABI,
http://rfmri.org/DPABI) suite in the MATLAB environment [
29]. The first 10 volumes of imaging data were discarded for maintaining the stability of the signals. The preprocessing steps included slice timing adjustment to the middle slice between the slices of a volume, registration for motion correction between the volumes of a scanning session to the first volume, and coregistration and normalization to a standardized Montreal Neurological Institute (MNI) template averaged by 512 individuals, followed by registration to the participant’s T1-weighted anatomical image using exponentiated Lie algebra. Thereafter, the normalized data were spatially smoothed using a 4 mm isotropic Gaussian kernel, temporally filtered with a high-pass filter of 128 s, and resampled at a resolution of 3 × 3 × 3 mm
3. After preprocessing, a linear detrending was applied to remove systematic drifting artifact before a bandpass filter was used to reserve imaging data in the spontaneous resting frequency (0.01–0.1 Hz).
Afterward, feature extraction based on the calculation of the connection intensity of the ACC functional network was performed on the preprocessed imaging data. Predefined ACC seeds were coordinated at (±7, 19, 30) in the MNI domain, according to an event-related fMRI study that conducted a modified Flanker task [
30]. The reference time course of the seed was defined as the averaged time course of a spherical 8 mm region around the seed. Thereafter, the functional connectivity over the brain was calculated between the reference time course and the remaining voxels in the brain, and the connectivity was presented as the correlation intensity based on the values of the Pearson correlation coefficients. After the calculation of the correlation, an overall functional map of connection intensity was generated after thresholding the correlation intensity whose value was above statistical significance (
p < 0.05) on an averaged map of two ACC-seeded networks.
In addition to presenting individual stationary functional network, we analyzed the interregional connectivity between regions of interest, which were extracted from the examination of the functional variation over groups. Here, we selected three action-monitoring-related regions [
31,
32,
33], namely, the ACC, the supplementary motor area (SMA), and the precuneus, for effective connectivity analysis. Interregional interaction of the regions of interest, that is, determination of effective connectivity considering the interactive influence bidirectionally, was performed between the target regions [
34]. This method proposed by the Granger causal model considers that the relationship of responsive time course between two regions (or two voxels) is bidirectionally influenced, that is,
x →
y and
y →
x. For example, the current signal (at time point
t) at region
x is composed of self-contribution and the contributions of the neighboring region, such as region
y. The self-influence originates from the temporal dependence of the last signal of region
x at time point (
t − ∆
t). The influence of the neighboring region originates from the functional dependence between two regions with high function correlation; that is, these regions might proceed with the same activity in response to a specific external stimulus at time point
t. Finally, the ratio of the contributions of the neighboring region to that of self-contribution was used to represent the directional influence from region
y to region
x (
y →
x). The directional influence from region
x to region
y (
x →
y) was calculated using the same scheme used for processing thoughts.
2.6. Statistical Analysis
Statistical analysis was performed using SPSS (IBM SPSS 19.0, Armonk, NY, USA). Analysis of variance (ANOVA), followed by homogeneity of variance between groups (using Levene’s test), was used for continuous variables, and the chi-squared test was used to analyze categorical variables to assess the differences among the three groups in terms of demographic data, PSG parameters, behavioral performance, and error-related ERP parameters. Normality of each sample data were also examined by the Kolmogorov–Smirnov test. These checks of data quality within and between data according to items ensured the statistical validity in subsequent statistical examinations. All statistical tests were two-tailed, and an alpha value of 0.05 indicated statistical significance. In all ANOVA comparisons, we additionally considered the clinical impact with effect size (ES) given as partial eta square (
ηp2), which was defined as the sum of squares effect (
SSbetween) over the sum of squares effect (
SSbetween) plus the sum of squares error (
SSerror), that is,
[
35]. ES is less limited by the sample size rather than by the
p value significance tests, and it is appropriate to explain the magnitude of the relationship in the population. The value corresponded to small, medium, and large ESs, respectively, as follows: 0.01 ≤
ηp2 < 0.06, 0.06 ≤
ηp2 < 0.14, and
ηp2 ≥ 0.14 [
35,
36].
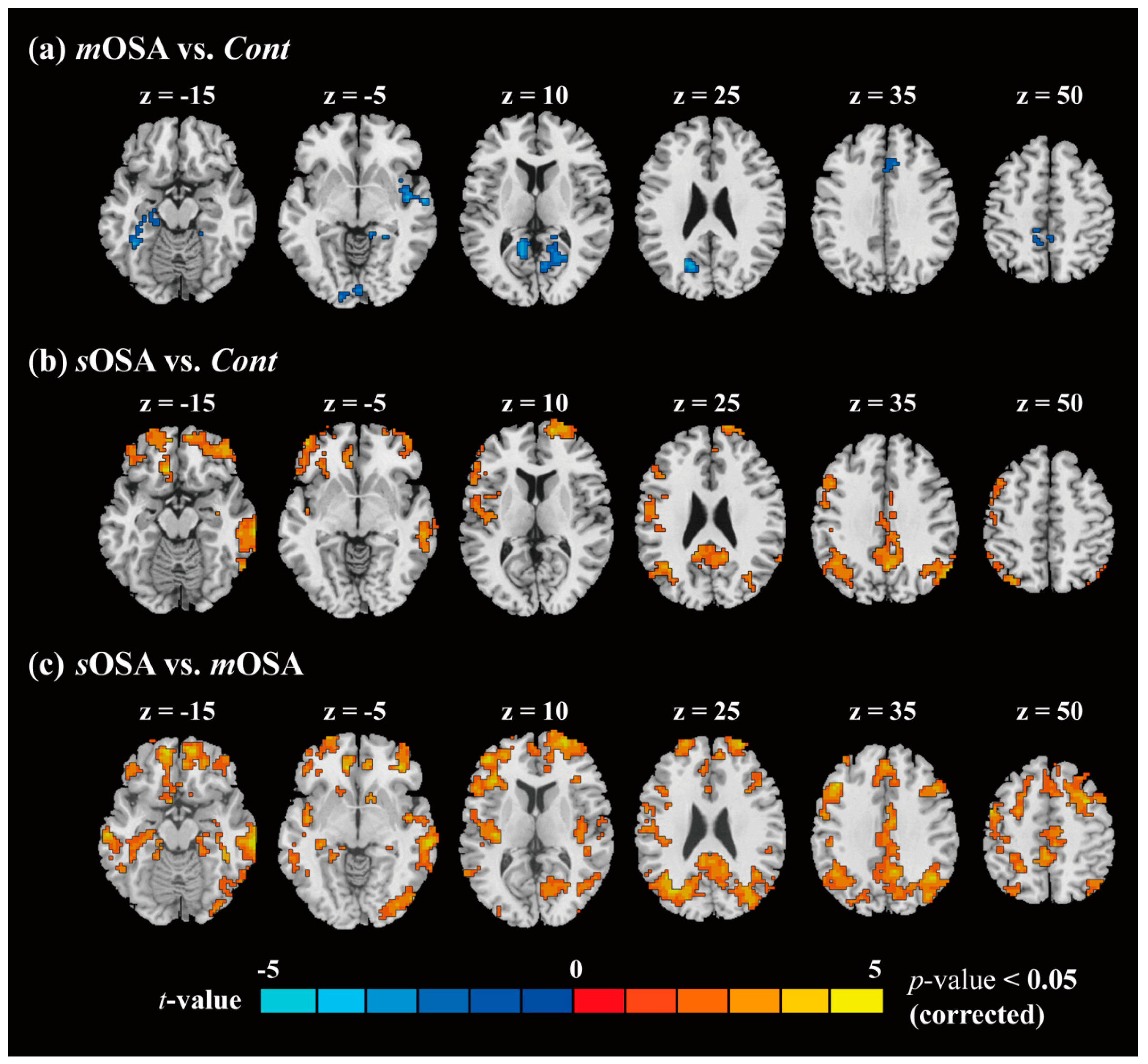
In the comparison of functional connectivity among the groups, we applied two-sample t-tests following the extent spatial constraint for activation cluster (with alpha simulation) to examine the difference between each two groups, for example, mOSA vs. Cont, sOSA vs. Cont, and sOSA vs. mOSA, followed by assessment of normality using the Kolmogorov–Smirnov test. The resulting statistical parametric maps (t value) helped us observe the functional contrast between two different levels of OSA severity. Later, by conjoining these contrast maps, we were able to perform further regional analysis across the three groups. Three regions related to action-monitoring function were selected, including the ACC, the SMA, and the precuneus. The functional difference across the three groups was analyzed by one-factor ANOVA, and a p value of 0.05 was considered for statistical significance.
4. Discussion
To the best of our knowledge, this case–control study is the first investigation to examine the action-monitoring function in patients with OSA using comprehensive behavioral, neurophysiological, and functional image evaluation. The primary findings are consistent with our hypothesis that patients with varying severities of OSA exhibit varying alterations in behavioral and functional performance underlying the action-monitoring function. In our study, patients with
mOSA exhibited reduced amplitudes of both ERN and Pe accompanied by impaired performance in the modified Flanker task, which indicated action-monitoring dysfunction in experimental sleep research [
19,
21,
37]. Consistently, the resting-state fMRI data also provided evidence of action-monitoring dysfunction in patients with
mOSA on the ACC functional network in our study. Using fMRI, we demonstrated that
mOSA is associated with decreased cerebral activation in multiple ACC functional-connected areas, compared with that in the
Cont group, suggesting that a compromised cerebral function in response to cognitive challenges underlies the action-monitoring dysfunction in patients with
mOSA.
Among factors responsible for action-monitoring dysfunction, sleep fragmentation and intermittent hypoxia may contribute to the dysfunction observed in patients with OSA. Sleep fragmentation disrupts sleep architecture and reduces the efficacy of restorative processes in sleep [
2]. Intermittent hypoxia increases oxidative stress and results in neuroinflammation in the frontal lobe [
38]. It has been reported that the combined drawbacks of sleep fragmentation and intermittent hypoxia result in decreased regional cerebral blood flow in the bilateral cingulate gyrus [
39]. Furthermore, the duration in hypoxia and the frequency of sleep fragmentation could contribute to decreased ACC activations in patients with OSA [
22,
23]. Based on the abovementioned concept, cerebral gray matter deficits, impaired white matter integrity, and functional disconnection in the ACC were apparently detected in patients with OSA [
40,
41,
42]. Overall, the pathophysiological impact of intermittent hypoxia and sleep fragmentation induces ACC damage, thereby contributing to the changes in ERN and Pe, altered ACC network, and finally leading directly to action-monitoring dysfunction in the moderate stage of OSA.
Another innovative finding observed in this study is that patients with
sOSA exhibited increased amplitudes of ERN and Pe and increased cerebral activation in multiple ACC functional-connected areas in the resting-state fMRI compared to those in patients with
mOSA. Increased neural activities and functional connectivity in the ACC in patients with
sOSA can be interpreted either as compensatory neural recruitment to maintain action-monitoring performance or as nonselective recruitment interfering with action-monitoring performance [
43,
44]. To address this issue, the action-monitoring performances of patients with
mOSA and those with
sOSA were examined and compared. A trend of deterioration in performances was observed in the modified Flanker task between patients with
mOSA and those with
sOSA, as shown in
Table 1. However, the post hoc analysis demonstrated no significant difference in the error response rate and the posterror correction rate in both the congruent and incongruent modified Flanker tasks between these two groups, which supports the finding that increased activation and functional connectivity in the ACC in patients with
sOSA may represent effective recruitment of neurons beyond those used by patients with
mOSA, to compensate the trend of deterioration in the action-monitoring function. Therefore, we suggest that increased activation and functional connectivity in the ACC-associated cerebral regions to maintain action-monitoring performance in patients with
sOSA represents a compensatory response.
The functional recruitment in patients with moderately severe OSA is supported by other cognitive evidence such as default mode networking, verbal learning skill, and working memory capability [
45,
46,
47]. Patients with OSA (mean AHI = 54.7) have been shown to demonstrate increased functional connectivity of the default mode in the posterior cingulate cortex, which might reflect neural compensation for cognitive damage [
45]. Furthermore, patients with OSA (mean AHI = 35.1) exhibited intact performance in verbal learning, accompanied by increased cerebral activations in the associated cerebral regions compared with those in healthy subjects [
46]. In an evaluation of working memory in another study, the results showed that with an increase in severity (mean AHI = 40.1), there was an increase in functional activations in the right parietal lobe, which may potentially reflect compensatory neural responses in patients with moderately severe OSA [
47]. To summarize, the findings in the literature together with our findings lead us to interpret that these increased cerebral activations in functional connectivity serve to compensate for natural neuronal activity to maintain the action-monitoring function in patients with moderately severe OSA.
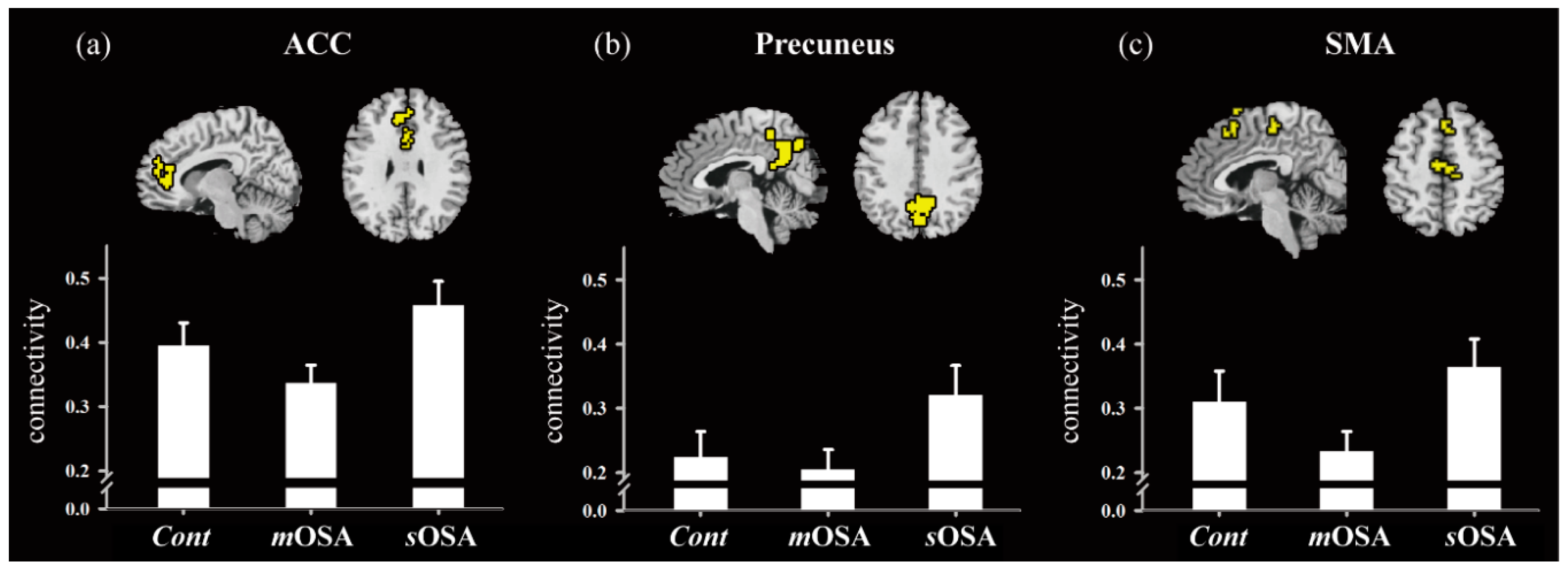
In addition to the changes in functional performance in specific regions, this study demonstrated the relationships among action-monitoring-related regions in the functional network. It has been reported that patients with OSA suffer from impaired network organization, resulting in dysfunctions of autonomic, affective, executive, sensorimotor, and cognitive functions [
48]. However, the functional network underlying the action-monitoring function is not well known. We selected the ACC, the precuneus, and the SMA for analyzing the functional connectivity because their functional roles have been specified in the essence of the action-monitoring function. SMA plays the leading role in early response-conflict monitoring by censoring the responses to correct or erroneous behavioral actions [
49]. Driven along the path of signal migration from the SMA, the ACC is closely involved in error processing [
31]. Furthermore, it has been reported that the activation response of the precuneus is associated with the adaptation or adjustment of successive actions in error processing, and this is required to shift attention to prevent successive error responses [
32,
33,
50]. In summary, a highly functional link exists between the SMA, the ACC, and the precuneus in the processing of action-monitoring.
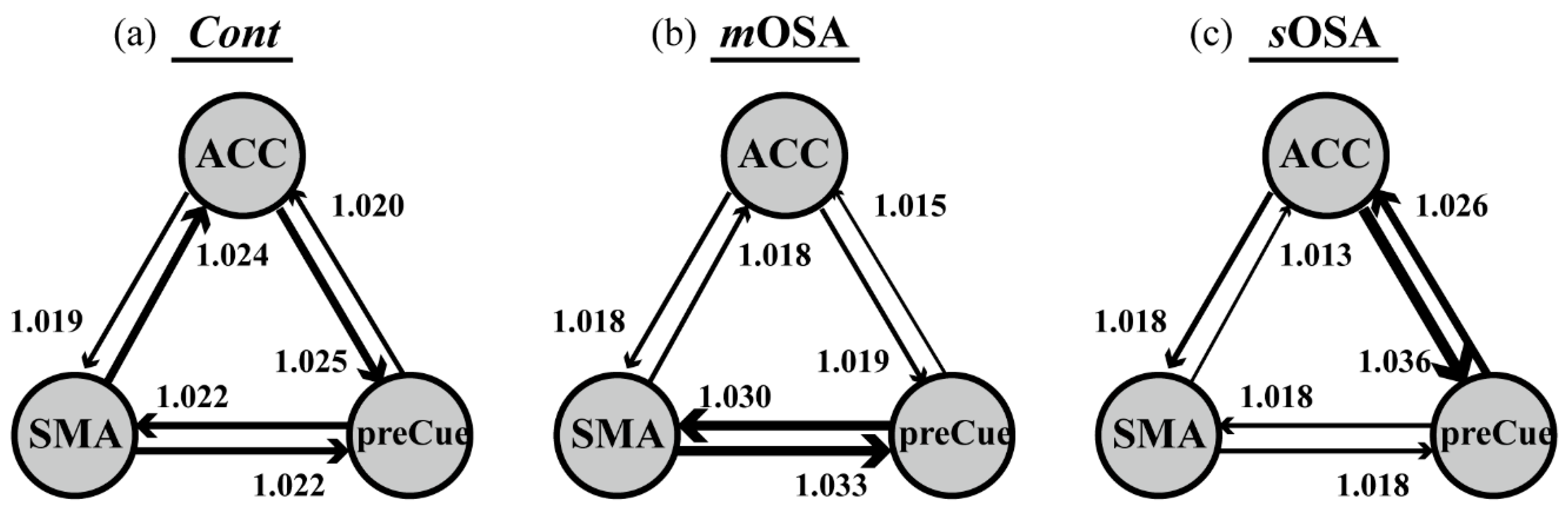
Our findings emphasized the role of the ACC in the action-monitoring function in the progression of OSA pathology; that is, the connection of the ACC to the SMA and the precuneus declines in the stage of moderate OSA, but it reflects reinforced connection to the precuneus in the stage of severe OSA (shown in
Figure 4). Our study also demonstrated altered activation in functional connectivity among the SMA, the ACC, and the precuneus based on the severity of OSA. In patients with
mOSA, the ACC had the most significantly reduced functional connections with the other two regions. In contrast, in patients with
sOSA, the connections between the ACC and the precuneus were re-enhanced most significantly. Based on the finding of deactivation and disrupted connections of the ACC in patients with
mOSA and the regained connections of the ACC in patients with
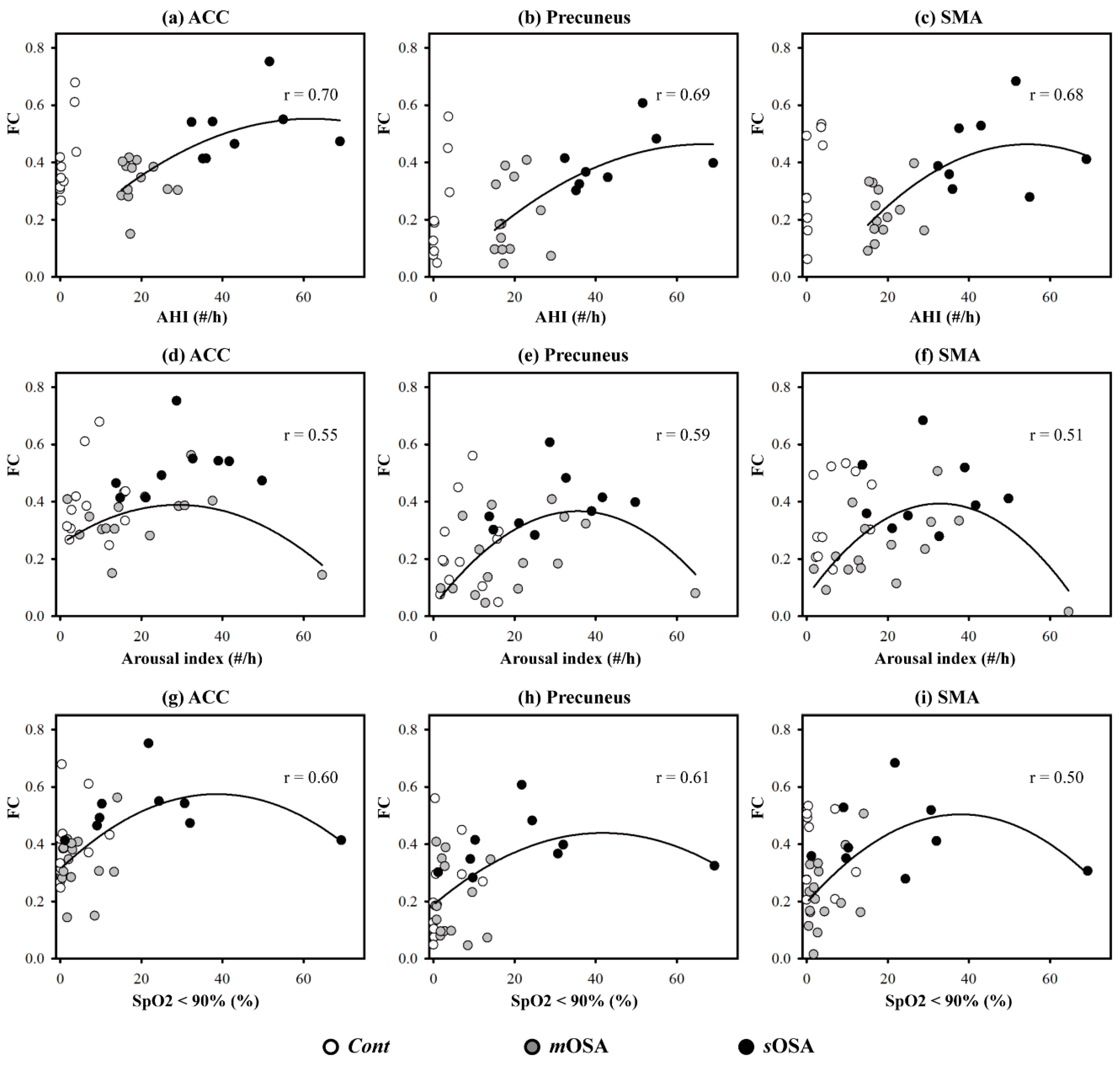
sOSA, it could be inferred that the ACC plays a vital role in terms of action-monitoring dysfunction, and that it is adaptable in the pathological progression of OSA. Furthermore, we have shown that the phenomenon of functional compensation might drop out as the AHI value exceeds 60 based on our regression model of function connectivity along with AHI values (shown in
Figure 5).
To the best of our knowledge, our study might be the first to examine the error-related ERPs in patients with OSA. Moreover, ACC networking associated with the action-monitoring function has never been reported in patients with moderate to moderately severe OSA. Our study results presented the possible occurrence of compensatory neural recruitment to compensate for the action-monitoring dysfunction in patients with moderately severe OSA based on comprehensive behavioral, electrophysiological, and functional image evaluation and supported an additional insight into neuronal plasticity for patients with OSA.
However, we should acknowledge the limitations of this study. First, although our data have passed the homogeneity of variance test on baseline information among the study groups using Levene’s test, it is also important to consider the issue of performance validity due to the small sample size in future work. Furthermore, there was a substantial female predominance in the healthy control group compared with the OSA groups. The impact of sex on cognitive impairment in OSA has not been determined before. Due to the small sample size, it was not possible to perform subgroup analysis stratified by sex in this study. Second, our study was aimed at investigating the action-monitoring function. To avoid the possible interference of global cognitive function in the action-monitoring function, we excluded subjects with CASI score below the cutoff value. This inclusion criterion limited our participants to moderately severe OSA, because very severe OSA could contribute to obvious multiple cognitive impairment. Third, we did not include patients with mild OSA in this study. Previous studies have reported that patients with mild OSA might not show appreciable deficits in overall cognition and neuropsychological tests [
51,
52]. Moreover, due to the hospital-based study design, it was difficult to enroll adequate number of patients with mild OSA. The symptoms of mild OSA might be too subtle to make these patients seek medical advice. Therefore, we were not able to determine a dose-dependent response for action-monitoring dysfunction over the complete range of severity of OSA. Further longitudinal studies addressing these issues could be considered to identify and confirm the changes in cerebral responses for the action-monitoring function in patients over the complete range of severity of OSA.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}