Intact Stimulus–Response Conflict Processing in ADHD—Multilevel Evidence and Theoretical Implications



Abstract
1. Introduction
2. Experimental Section
2.1. Samples and Power Considerations
2.2. Task
2.3. EEG Recording and Analysis
2.4. Residue Iteration Decomposition (RIDE)
2.5. Statistics
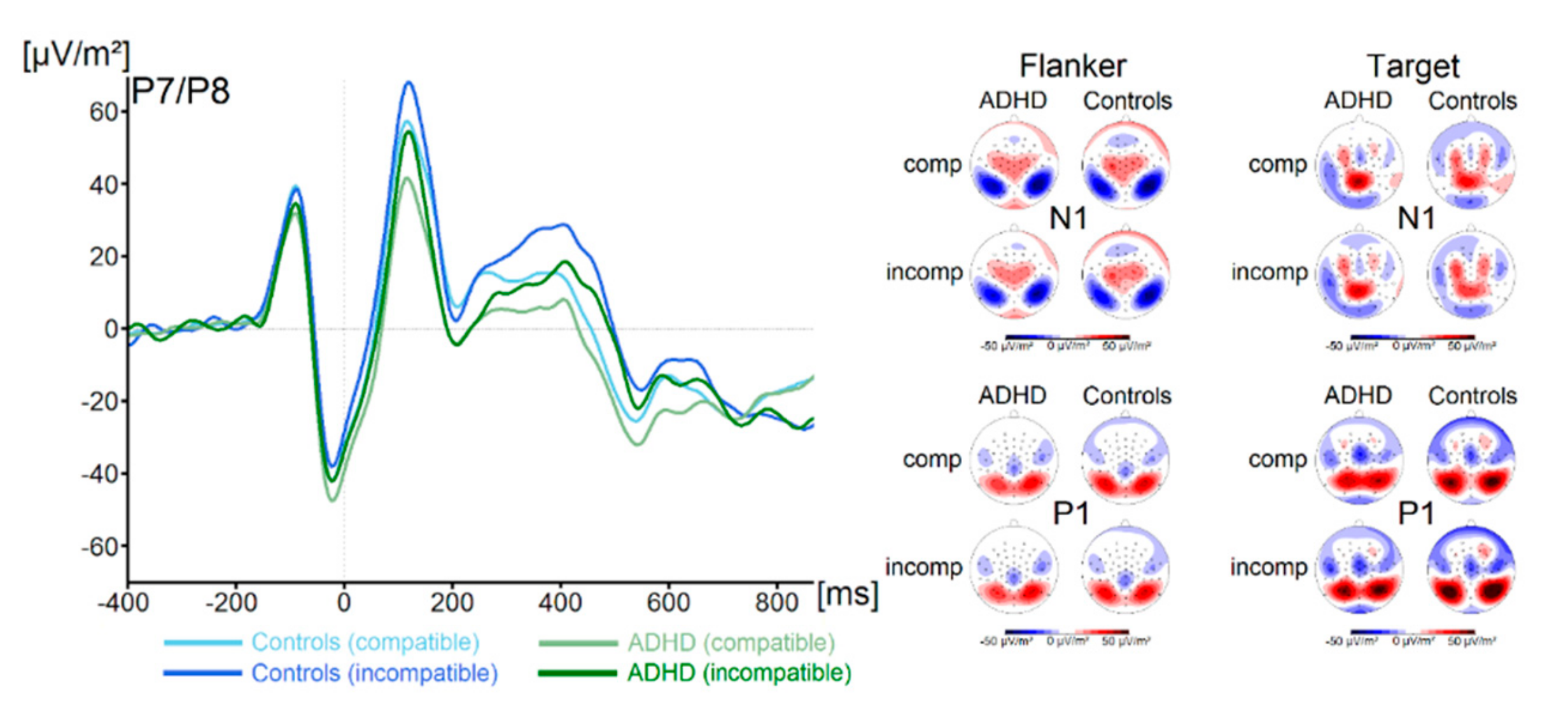
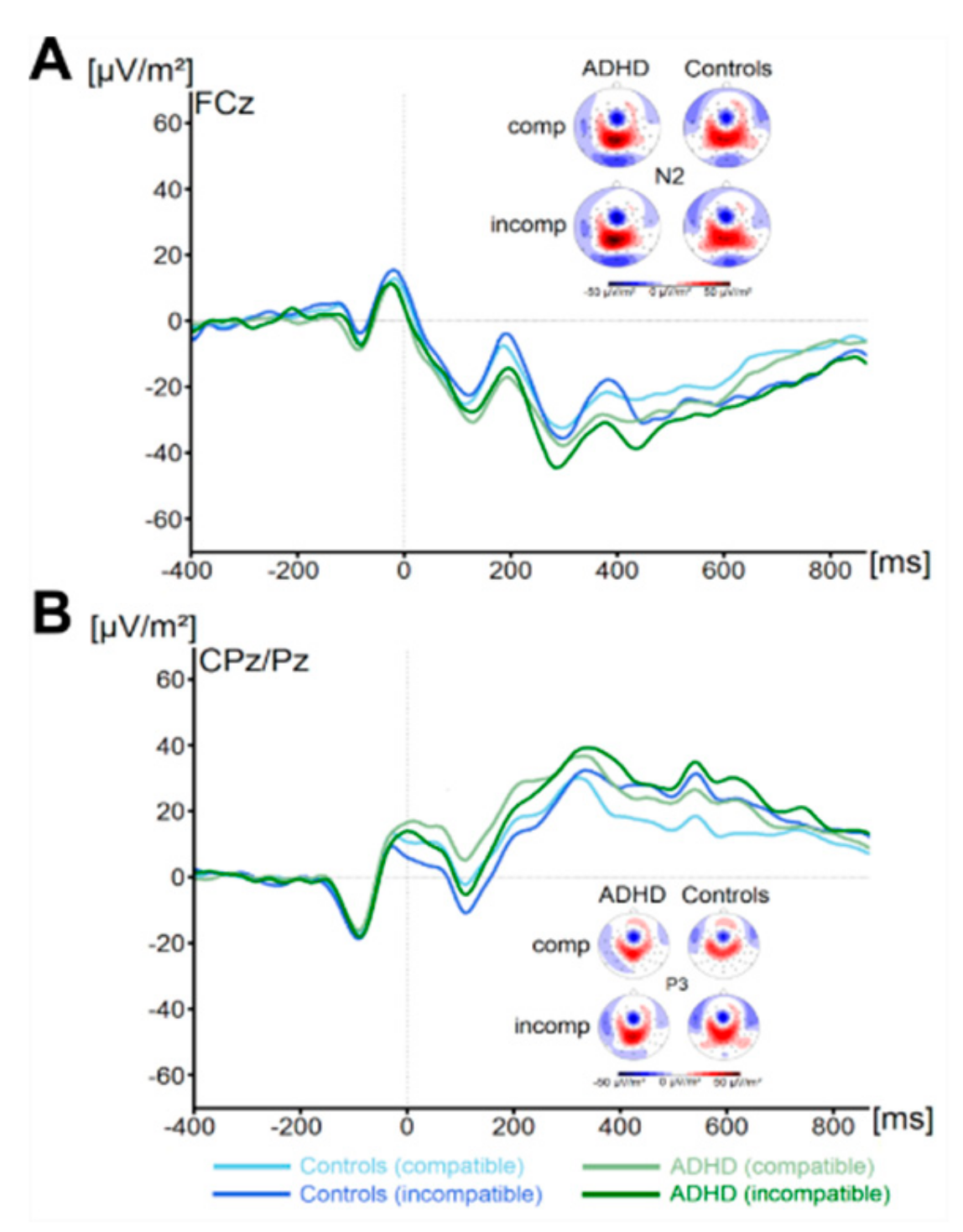
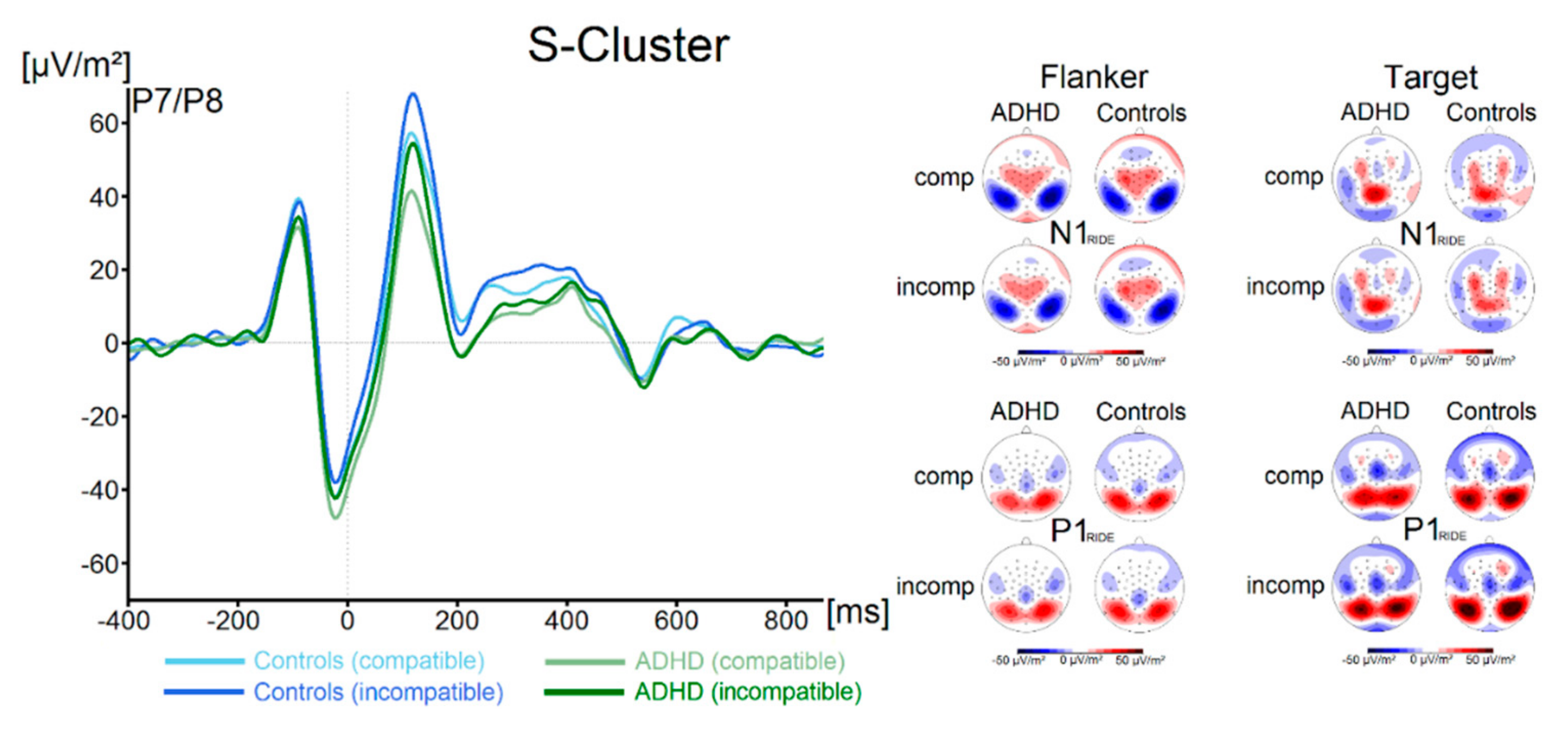
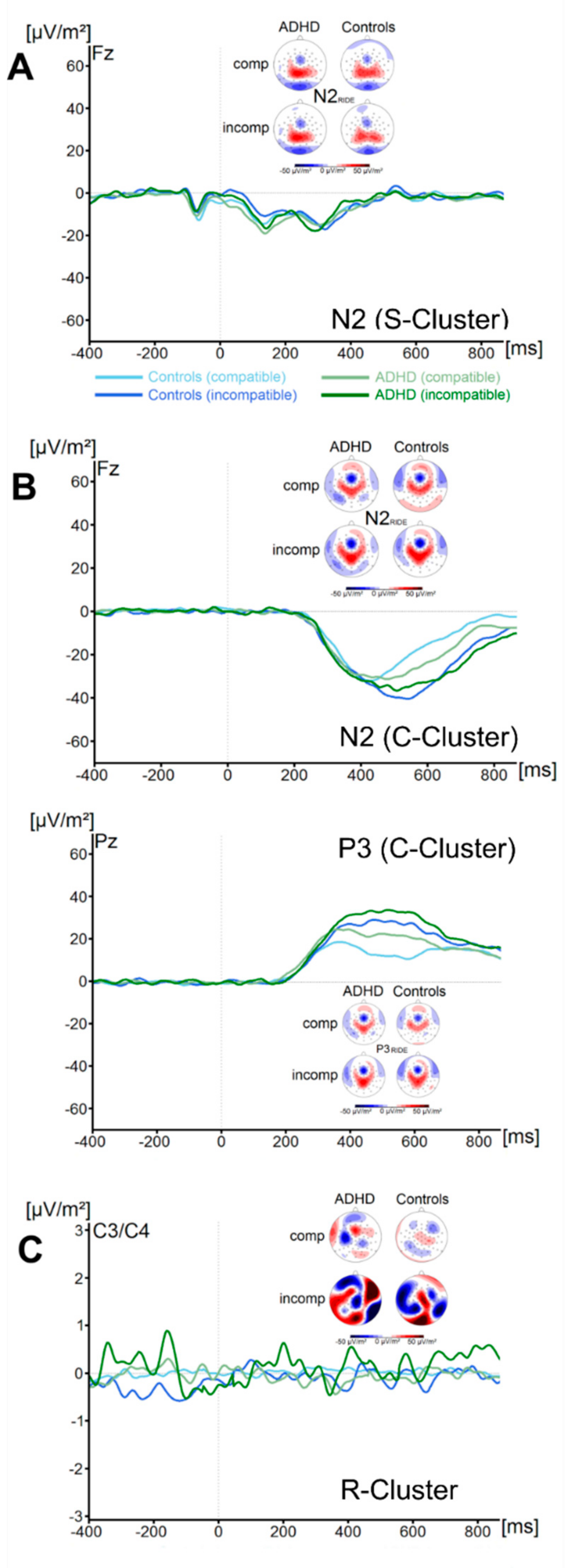
3. Results
3.1. Behavioural Data
3.2. Neurophysiological Data
3.2.1. Standard ERP Analysis
3.2.2. RIDE Analysis
S-Cluster
C-Cluster
R-Cluster
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ahmadi, N.; Mohammadi, M.R.; Araghi, S.M.; Zarafshan, H. Neurocognitive profile of children with attention deficit hyperactivity disorders (ADHD): A comparison between subtypes. Iran. J. Psychiatry 2014, 9, 197–202. [Google Scholar] [PubMed]
- Arnsten, A.F.T.; Rubia, K. Neurobiological circuits regulating attention, cognitive control, motivation, and emotion: Disruptions in neurodevelopmental psychiatric disorders. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Oosterlaan, J.; Sergeant, J.A. Response inhibition and response re-engagement in attention-deficit/hyperactivity disorder, disruptive, anxious and normal children. Behav. Brain Res. 1998, 94, 33–43. [Google Scholar] [CrossRef]
- Randall, K.D.; Brocki, K.C.; Kerns, K.A. Cognitive control in children with ADHD-C: How efficient are they? Child Neuropsychol. J. Norm. Abnorm. Dev. Child. Adolesc. 2009, 15, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Stroux, D.; Shushakova, A.; Geburek-Höfer, A.J.; Ohrmann, P.; Rist, F.; Pedersen, A. Deficient interference control during working memory updating in adults with ADHD: An event-related potential study. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2016, 127, 452–463. [Google Scholar] [CrossRef]
- Urcelay, G.P.; Dalley, J.W. Linking ADHD, impulsivity, and drug abuse: A neuropsychological perspective. Curr. Top. Behav. Neurosci. 2012, 9, 173–197. [Google Scholar]
- Van Rooij, D.; Hartman, C.A.; Mennes, M.; Oosterlaan, J.; Franke, B.; Rommelse, N.; Heslenfeld, D.; Faraone, S.V.; Buitelaar, J.K.; Hoekstra, P.J. Altered neural connectivity during response inhibition in adolescents with attention-deficit/hyperactivity disorder and their unaffected siblings. NeuroImage Clin. 2015, 7, 325–335. [Google Scholar] [CrossRef]
- Diamond, A. Executive functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2008, 45, 152–170. [Google Scholar] [CrossRef]
- Shenhav, A.; Botvinick, M.M.; Cohen, J.D. The expected value of control: An integrative theory of anterior cingulate cortex function. Neuron 2013, 79, 217–240. [Google Scholar] [CrossRef]
- Carter, C.S.; van Veen, V. Anterior cingulate cortex and conflict detection: An update of theory and data. Cogn. Affect. Behav. Neurosci. 2007, 7, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Larson, M.J.; Clayson, P.E.; Clawson, A. Making sense of all the conflict: A theoretical review and critique of conflict-related ERPs. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2014, 93, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, W.X.; Mückschel, M.; Beste, C. Response selection codes in neurophysiological data predict conjoint effects of controlled and automatic processes during response inhibition. Hum. Brain Mapp. 2018, 39, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Mückschel, M.; Chmielewski, W.; Ziemssen, T.; Beste, C. The norepinephrine system shows information-content specific properties during cognitive control—Evidence from EEG and pupillary responses. NeuroImage 2017, 149, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Mückschel, M.; Dippel, G.; Beste, C. Distinguishing stimulus and response codes in theta oscillations in prefrontal areas during inhibitory control of automated responses. Hum. Brain Mapp. 2017, 38, 5681–5690. [Google Scholar] [CrossRef] [PubMed]
- Faraone, S.V. The pharmacology of amphetamine and methylphenidate: Relevance to the neurobiology of attention-deficit/hyperactivity disorder and other psychiatric comorbidities. Neurosci. Biobehav. Rev. 2018, 87, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Friedrich, J.; Schreiter, M.L.; Roessner, V.; Beste, C. A comparative study on the neurophysiological mechanisms underlying effects of methylphenidate and neurofeedback on inhibitory control in attention deficit hyperactivity disorder. NeuroImage Clin. 2018, 20, 1191–1203. [Google Scholar] [CrossRef]
- Bluschke, A.; von der Hagen, M.; Papenhagen, K.; Roessner, V.; Beste, C. Response inhibition in attention deficit disorder and neurofibromatosis type 1-clinically similar, neurophysiologically different. Sci. Rep. 2017, 7, 43929. [Google Scholar] [CrossRef]
- Chmielewski, W.; Bluschke, A.; Bodmer, B.; Wolff, N.; Roessner, V.; Beste, C. Evidence for an altered architecture and a hierarchical modulation of inhibitory control processes in ADHD. Dev. Cogn. Neurosci. 2019, 36, 100623. [Google Scholar] [CrossRef]
- Albrecht, B.; Brandeis, D.; Uebel, H.; Heinrich, H.; Mueller, U.C.; Hasselhorn, M.; Steinhausen, H.-C.; Rothenberger, A.; Banaschewski, T. Action monitoring in boys with attention-deficit/hyperactivity disorder, their nonaffected siblings, and normal control subjects: Evidence for an endophenotype. Biol. Psychiatry 2008, 64, 615–625. [Google Scholar] [CrossRef][Green Version]
- Dockstader, C.; Gaetz, W.; Cheyne, D.; Tannock, R. Abnormal neural reactivity to unpredictable sensory events in attention-deficit/hyperactivity disorder. Biol. Psychiatry 2009, 66, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Karatekin, C.; Bingham, C.; White, T. Regulation of cognitive resources during an n-back task in youth-onset psychosis and attention-deficit/hyperactivity disorder (ADHD). Int. J. Psychophysiol. 2009, 73, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, J.A.; van der Meere, J. What happens after a hyperactive child commits an error? Psychiatry Res. 1988, 24, 157–164. [Google Scholar] [CrossRef]
- Spinelli, S.; Joel, S.; Nelson, T.E.; Vasa, R.A.; Pekar, J.J.; Mostofsky, S.H. Different neural patterns are associated with trials preceding inhibitory errors in children with and without attention-deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry 2011, 50, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Van Meel, C.S.; Heslenfeld, D.J.; Oosterlaan, J.; Sergeant, J.A. Adaptive control deficits in attention-deficit/hyperactivity disorder (ADHD): The role of error processing. Psychiatry Res. 2007, 151, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Chmielewski, W.X.; Roessner, V.; Beste, C. Intact context-dependent modulation of conflict monitoring in childhood ADHD. J. Atten. Disord. 2016. [CrossRef]
- Gmehlin, D.; Fuermaier, A.B.M.; Walther, S.; Debelak, R.; Rentrop, M.; Westermann, C.; Sharma, A.; Tucha, L.; Koerts, J.; Tucha, O.; et al. Intraindividual variability in inhibitory function in adults with ADHD—An ex-Gaussian approach. PLoS ONE 2014, 9, e112298. [Google Scholar] [CrossRef]
- Henríquez-Henríquez, M.P.; Billeke, P.; Henríquez, H.; Zamorano, F.J.; Rothhammer, F.; Aboitiz, F. Intra-individual response variability assessed by ex-gaussian analysis may be a new endophenotype for attention-deficit/hyperactivity disorder. Front. Psychiatry 2014, 5, 197. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Hwang-Gu, S.-L.; Gau, S.S.-F. Intra-individual reaction time variability based on ex-Gaussian distribution as a potential endophenotype for attention-deficit/hyperactivity disorder. Acta Psychiatr. Scand. 2015, 132, 39–50. [Google Scholar] [CrossRef]
- Plessen, K.J.; Allen, E.A.; Eichele, H.; van Wageningen, H.; Høvik, M.F.; Sørensen, L.; Worren, M.K.; Hugdahl, K.; Eichele, T. Reduced error signalling in medication-naive children with ADHD: Associations with behavioural variability and post-error adaptations. J. Psychiatry Neurosci. 2016, 41, 77–87. [Google Scholar] [CrossRef]
- Saville, C.W.N.; Feige, B.; Kluckert, C.; Bender, S.; Biscaldi, M.; Berger, A.; Fleischhaker, C.; Henighausen, K.; Klein, C. Increased reaction time variability in attention-deficit hyperactivity disorder as a response-related phenomenon: Evidence from single-trial event-related potentials. J. Child Psychol. Psychiatry 2015, 56, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Alba, G.; Pereda, E.; Mañas, S.; Méndez, L.D.; Duque, M.R.; González, A.; González, J.J. The variability of EEG functional connectivity of young ADHD subjects in different resting states. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2016, 127, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Gohil, K.; Petzold, M.; Roessner, V.; Beste, C. Neural mechanisms underlying successful and deficient multi-component behavior in early adolescent ADHD. NeuroImage Clin. 2018, 18, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Chmielewski, W.X.; Mückschel, M.; Roessner, V.; Beste, C. Neuronal intra-individual variability masks response selection differences between ADHD subtypes—A need to change perspectives. Front. Hum. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Gonen-Yaacovi, G.; Arazi, A.; Shahar, N.; Karmon, A.; Haar, S.; Meiran, N.; Dinstein, I. Increased ongoing neural variability in ADHD. Cortex J. Devoted Study Nerv. Syst. Behav. 2016, 81, 50–63. [Google Scholar] [CrossRef]
- Lazzaro, I.; Anderson, J.; Gordon, E.; Clarke, S.; Leong, J.; Meares, R. Single trial variability within the P300 (250–500 ms) processing window in adolescents with attention deficit hyperactivity disorder. Psychiatry Res. 1997, 73, 91–101. [Google Scholar] [CrossRef]
- Ouyang, G.; Sommer, W.; Zhou, C. A toolbox for residue iteration decomposition (RIDE)—A method for the decomposition, reconstruction, and single trial analysis of event related potentials. J. Neurosci. Methods 2015, 250, 7–21. [Google Scholar] [CrossRef]
- Ouyang, G.; Herzmann, G.; Zhou, C.; Sommer, W. Residue iteration decomposition (RIDE): A new method to separate ERP components on the basis of latency variability in single trials. Psychophysiology 2011, 48, 1631–1647. [Google Scholar] [CrossRef]
- Hasshim, N.; Parris, B.A. Assessing stimulus–stimulus (semantic) conflict in the Stroop task using saccadic two-to-one color response mapping and preresponse pupillary measures. Atten. Percept. Psychophys. 2015, 77, 2601–2610. [Google Scholar] [CrossRef]
- Hommel, B. The Simon effect as tool and heuristic. Acta Psychol. (Amst) 2011, 136, 189–202. [Google Scholar] [CrossRef]
- Kornblum, S.; Hasbroucq, T.; Osman, A. Dimensional overlap: Cognitive basis for stimulus-response compatibility—A model and taxonomy. Psychol. Rev. 1990, 97, 253–270. [Google Scholar] [CrossRef] [PubMed]
- De Houwer, J. On the role of stimulus-response and stimulus-stimulus compatibility in the Stroop effect. Mem. Cognit. 2003, 31, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Lansbergen, M.M.; Kenemans, J.L.; van Engeland, H. Stroop interference and attention-deficit/hyperactivity disorder: A review and meta-analysis. Neuropsychology 2007, 21, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Mullane, J.C.; Corkum, P.V.; Klein, R.M.; McLaughlin, E. Interference control in children with and without ADHD: A systematic review of Flanker and Simon task performance. Child Neuropsychol. J. Norm. Abnorm. Dev. Child. Adolesc. 2009, 15, 321–342. [Google Scholar] [CrossRef]
- Coderre, E.L.; van Heuven, W.J.B. Modulations of the executive control network by stimulus onset asynchrony in a Stroop task. BMC Neurosci. 2013, 14, 79. [Google Scholar] [CrossRef]
- Lei, H.; Yi, J.; Wang, H.; Zhang, X.; Dong, J.; Zhou, C.; Fan, J.; Zhong, M.; Zhu, X. Inhibitory deficit in semantic conflict in obsessive-compulsive disorder: An event-related potential study. Neurosci. Lett. 2013, 552, 162–167. [Google Scholar] [CrossRef]
- Li, Q.; Yang, G.; Li, Z.; Qi, Y.; Cole, M.W.; Liu, X. Conflict detection and resolution rely on a combination of common and distinct cognitive control networks. Neurosci. Biobehav. Rev. 2017, 83, 123–131. [Google Scholar] [CrossRef]
- Hart, H.; Radua, J.; Nakao, T.; Mataix-Cols, D.; Rubia, K. Meta-analysis of functional magnetic resonance imaging studies of inhibition and attention in attention-deficit/hyperactivity disorder: Exploring task-specific, stimulant medication, and age effects. JAMA Psychiatry 2013, 70, 185–198. [Google Scholar] [CrossRef]
- Rubia, K.; Alegria, A.A.; Cubillo, A.I.; Smith, A.B.; Brammer, M.J.; Radua, J. Effects of stimulants on brain function in attention-deficit/hyperactivity disorder: A systematic review and meta-analysis. Biol. Psychiatry 2014, 76, 616–628. [Google Scholar] [CrossRef]
- Chmielewski, W.X.; Beste, C. Stimulus feature conflicts enhance motor inhibitory control processes in the lateral prefrontal Cortex. J. Cogn. Neurosci. 2019, 31, 1430–1442. [Google Scholar] [CrossRef]
- Chmielewski, W.X.; Beste, C. Neurophysiological mechanisms underlying the modulation of cognitive control by simultaneous conflicts. Cortex J. Devoted Study Nerv. Syst. Behav. 2019, 115, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A.G. Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Döpfner, M.; Görtz-Dorten, A.; Lehmkuhl, G. Diagnostik-System für Psychische Störungen im Kindes- und Jugendalter nach ICD-10 und DSM-IV, DISYPS-II; Huber: Bern, Switzerland, 2008. [Google Scholar]
- Nunez, P.L.; Pilgreen, K.L. The spline-Laplacian in clinical neurophysiology: A method to improve EEG spatial resolution. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 1991, 8, 397–413. [Google Scholar] [CrossRef]
- Mückschel, M.; Stock, A.-K.; Beste, C. Psychophysiological mechanisms of interindividual differences in goal activation modes during action cascading. Cereb. Cortex 2014, 24, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.; Sommer, W.; Zhou, C. Updating and validating a new framework for restoring and analyzing latency-variable ERP components from single trials with residue iteration decomposition (RIDE). Psychophysiology 2015, 52, 839–856. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.; Schacht, A.; Zhou, C.; Sommer, W. Overcoming limitations of the ERP method with Residue Iteration Decomposition (RIDE): A demonstration in go/no-go experiments. Psychophysiology 2013, 50, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Masson, M.E.J. A tutorial on a practical Bayesian alternative to null-hypothesis significance testing. Behav. Res. Methods 2011, 43, 679–690. [Google Scholar] [CrossRef]
- Raftery, A.E. Bayesian model selection in social research. Sociol. Methodol. 1995, 25, 111–163. [Google Scholar] [CrossRef]
- Sebastian, A.; Jung, P.; Krause-Utz, A.; Lieb, K.; Schmahl, C.; Tüscher, O. Frontal dysfunctions of impulse control—A systematic review in borderline personality disorder and attention-deficit/hyperactivity disorder. Front. Hum. Neurosci. 2014, 8, 698. [Google Scholar] [CrossRef]
- Zhang, H.; Kornblum, S. The effects of stimulus–response mapping and irrelevant stimulus–response and stimulus–stimulus overlap in four-choice Stroop tasks with single-carrier stimuli. J. Exp. Psychol. Hum. Percept. Perform. 1998, 24, 3–19. [Google Scholar] [CrossRef]
- Schmidt, J.R.; Cheesman, J. Dissociating stimulus-stimulus and response-response effects in the stroop task. Can. J. Exp. Psychol. Can. Psychol. Expérimentale 2005, 59, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Bari, A.; Robbins, T.W. Inhibition and impulsivity: Behavioral and neural basis of response control. Prog. Neurobiol. 2013, 108, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.R.; Cai, W.; Badre, D.; Robbins, T.W. Evidence supports specific braking function for inferior PFC. Trends Cogn. Sci. 2015, 19, 711–712. [Google Scholar] [CrossRef] [PubMed]
- West, R. Neural correlates of cognitive control and conflict detection in the Stroop and digit-location tasks. Neuropsychologia 2003, 41, 1122–1135. [Google Scholar] [CrossRef]
- West, R.; Jakubek, K.; Wymbs, N.; Perry, M.; Moore, K. Neural correlates of conflict processing. Exp. Brain Res. 2005, 167, 38–48. [Google Scholar] [CrossRef]
- Chmielewski, W.X.; Tiedt, A.; Bluschke, A.; Dippel, G.; Roessner, V.; Beste, C. Effects of multisensory stimuli on inhibitory control in adolescent ADHD: It is the content of information that matters. NeuroImage Clin. 2018, 19, 527–537. [Google Scholar] [CrossRef]
- Fallgatter, A.J.; Ehlis, A.-C.; Rösler, M.; Strik, W.K.; Blocher, D.; Herrmann, M.J. Diminished prefrontal brain function in adults with psychopathology in childhood related to attention deficit hyperactivity disorder. Psychiatry Res. 2005, 138, 157–169. [Google Scholar] [CrossRef]
- Fallgatter, A.J.; Ehlis, A.-C.; Seifert, J.; Strik, W.K.; Scheuerpflug, P.; Zillessen, K.E.; Herrmann, M.J.; Warnke, A. Altered response control and anterior cingulate function in attention-deficit/hyperactivity disorder boys. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2004, 115, 973–981. [Google Scholar] [CrossRef]
- Paul-Jordanov, I.; Bechtold, M.; Gawrilow, C. Methylphenidate and if-then plans are comparable in modulating the P300 and increasing response inhibition in children with ADHD. Atten. Deficit Hyperact. Disord. 2010, 2, 115–126. [Google Scholar] [CrossRef]
- Pliszka, S.R.; Liotti, M.; Bailey, B.Y.; Perez, R., III; Glahn, D.; Semrud-Clikeman, M. Electrophysiological effects of stimulant treatment on inhibitory control in children with attention-deficit/hyperactivity disorder. J. Child Adolesc. Psychopharmacol. 2007, 17, 356–366. [Google Scholar] [CrossRef]
- Seifert, J.; Scheuerpflug, P.; Zillessen, K.-E.; Fallgatter, A.; Warnke, A. Electrophysiological investigation of the effectiveness of methylphenidate in children with and without ADHD. J. Neural Transm. 2003, 110, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Van der Oord, S.; Geurts, H.M.; Prins, P.J.M.; Emmelkamp, P.M.G.; Oosterlaan, J. Prepotent response inhibition predicts treatment outcome in attention deficit/hyperactivity disorder. Child Neuropsychol. J. Norm. Abnorm. Dev. Child. Adolesc. 2012, 18, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Broschwitz, F.; Kohl, S.; Roessner, V.; Beste, C. The neuronal mechanisms underlying improvement of impulsivity in ADHD by theta/beta neurofeedback. Sci. Rep. 2016, 6, 31178. [Google Scholar] [CrossRef] [PubMed]
- Bluschke, A.; Roessner, V.; Beste, C. Specific cognitive-neurophysiological processes predict impulsivity in the childhood attention-deficit/hyperactivity disorder combined subtype. Psychol. Med. 2016, 46, 1277–1287. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Navarra, R.; Graf, R.; Huang, Y.; Logue, S.; Comery, T.; Hughes, Z.; Day, M. Effects of atomoxetine and methylphenidate on attention and impulsivity in the 5-choice serial reaction time test. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Bedard, A.-C.V.; Stein, M.A.; Halperin, J.M.; Krone, B.; Rajwan, E.; Newcorn, J.H. Diffenrtial impact of methylphenidate and atomoxetine on sustained attention in youth with attention-deficit/hyperactivity disorder. J. Child. Psychol. Psychiatry 2015, 56, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, Y.; Ota, T.; Iida, J.; Yamamuro, K.; Kishimotor, N.; Okzaki, K.; Kishimoto, T. Differential therapeutic effects of atomoxetine and methylphenidate in childhood attention deficit/hyperactivity disorder as measured by near-infrared spectroscopy. Child. Adolesc. Psychiatry Ment. Health 2017, 11, 26. [Google Scholar] [CrossRef]
- Schulz, K.P.; Fan, J.; Bedard, A.C.V.; Clerkin, S.M.; Ivanov, I.; Tang, C.; Halperin, J.; Newcorn, J.H. Common and unique therapeutic mechanisms of stimulant and nonstimulant treatments for attention-deficit/hyperactivity disorder. Arch. Gen. Psychiatry 2012, 69, 952–961. [Google Scholar] [CrossRef]
- Hermens, D.F.; Rowe, D.L.; Gordon, E.; Williams, L.M. Integrative neuroscience approach to predict ADHD stimulant response. Expert Rev. Neurother. 2006, 6, 753–763. [Google Scholar] [CrossRef]
- Stein, M.A.; MCGough, J.J. The pharmacogenomic era: Promise for personalizing ADHD therapy. Child. Adolesc. Psychiatr. Clin. N. Am. 2008, 17, 475–490. [Google Scholar] [CrossRef][Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Controls (n = 33) | Patients with ADHD (n = 36) | Group Comparison | |
|---|---|---|---|
| n male | 21 | 24 | Χ2(1) = 0.07, p = 0.79 |
| mean age ± SE | 11.7 ± 0.4 years | 11.7 ± 0.3 years | t(68) = 0.11; p = 0.92 |
| mean IQ ± SE | 106 ± 1.3 | 102 ± 1.7 | t(68) = −1.8; p = 0.08 |
| ADHD symptom checklist inattention (mean ± SE) | 0.5 ± 0.1 | 2.0 ± 0.1 | t(62) = 11.2; p ≤ 0.001 |
| ADHD symptom checklist hyperactivity (mean ± SE) | 0.09 ± 0.05 | 1.1 ± 0.1 | t(62) = 7.9; p ≤ 0.001 |
| ADHD symptom checklist impulsivity (mean ± SE) | 0.3 ± 0.09 | 1.5 ± 0.2 | t(62) = 6.5; p ≤ 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bluschke, A.; Mückschel, M.; Roessner, V.; Beste, C. Intact Stimulus–Response Conflict Processing in ADHD—Multilevel Evidence and Theoretical Implications. J. Clin. Med. 2020, 9, 234. https://doi.org/10.3390/jcm9010234
Bluschke A, Mückschel M, Roessner V, Beste C. Intact Stimulus–Response Conflict Processing in ADHD—Multilevel Evidence and Theoretical Implications. Journal of Clinical Medicine. 2020; 9(1):234. https://doi.org/10.3390/jcm9010234
Chicago/Turabian StyleBluschke, Annet, Moritz Mückschel, Veit Roessner, and Christian Beste. 2020. "Intact Stimulus–Response Conflict Processing in ADHD—Multilevel Evidence and Theoretical Implications" Journal of Clinical Medicine 9, no. 1: 234. https://doi.org/10.3390/jcm9010234
APA StyleBluschke, A., Mückschel, M., Roessner, V., & Beste, C. (2020). Intact Stimulus–Response Conflict Processing in ADHD—Multilevel Evidence and Theoretical Implications. Journal of Clinical Medicine, 9(1), 234. https://doi.org/10.3390/jcm9010234