Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. ICA Animal Model and hMSCs Transplantation
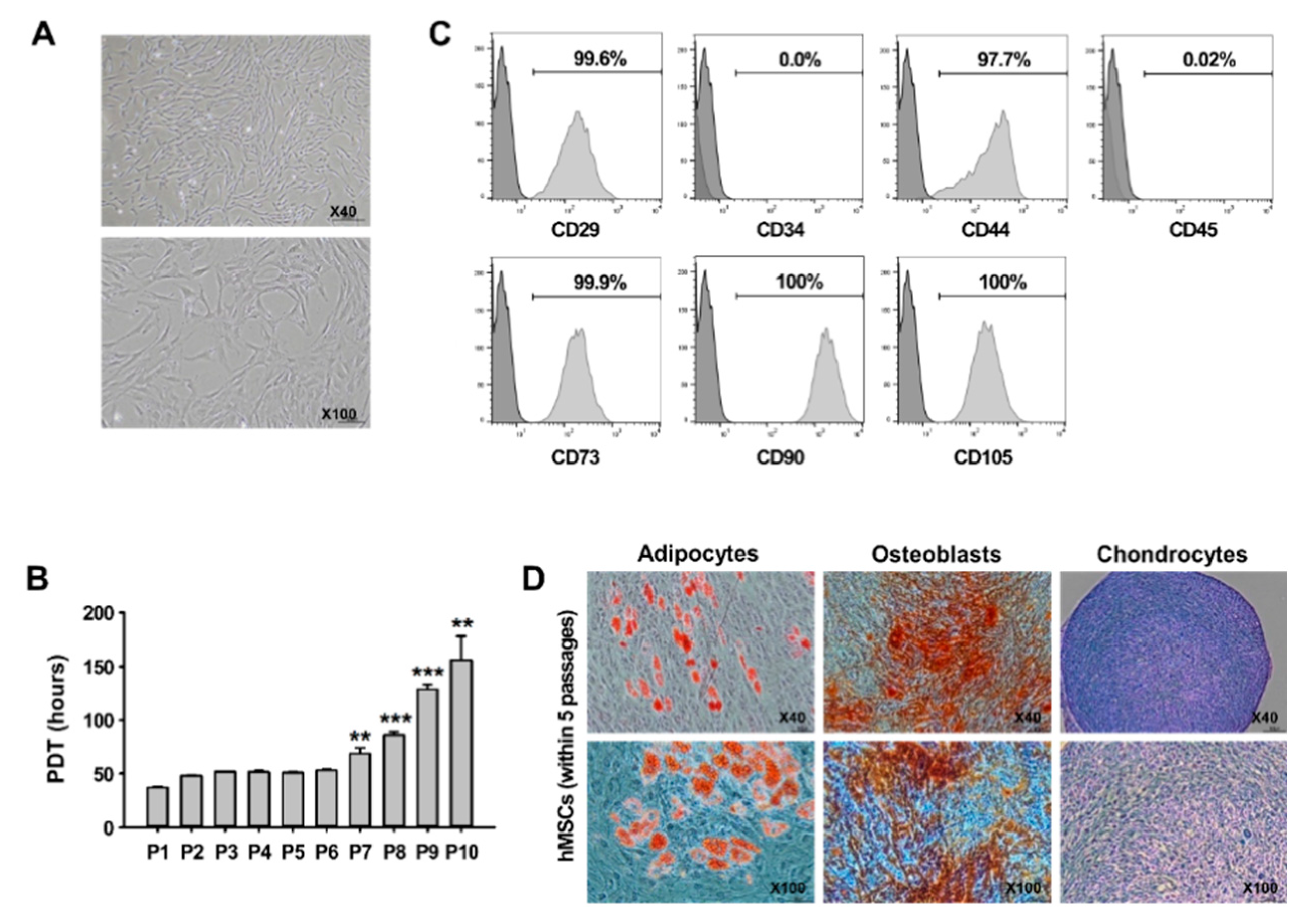
2.3. hMSC Isolation and Characterization
2.4. Behavioral Tests
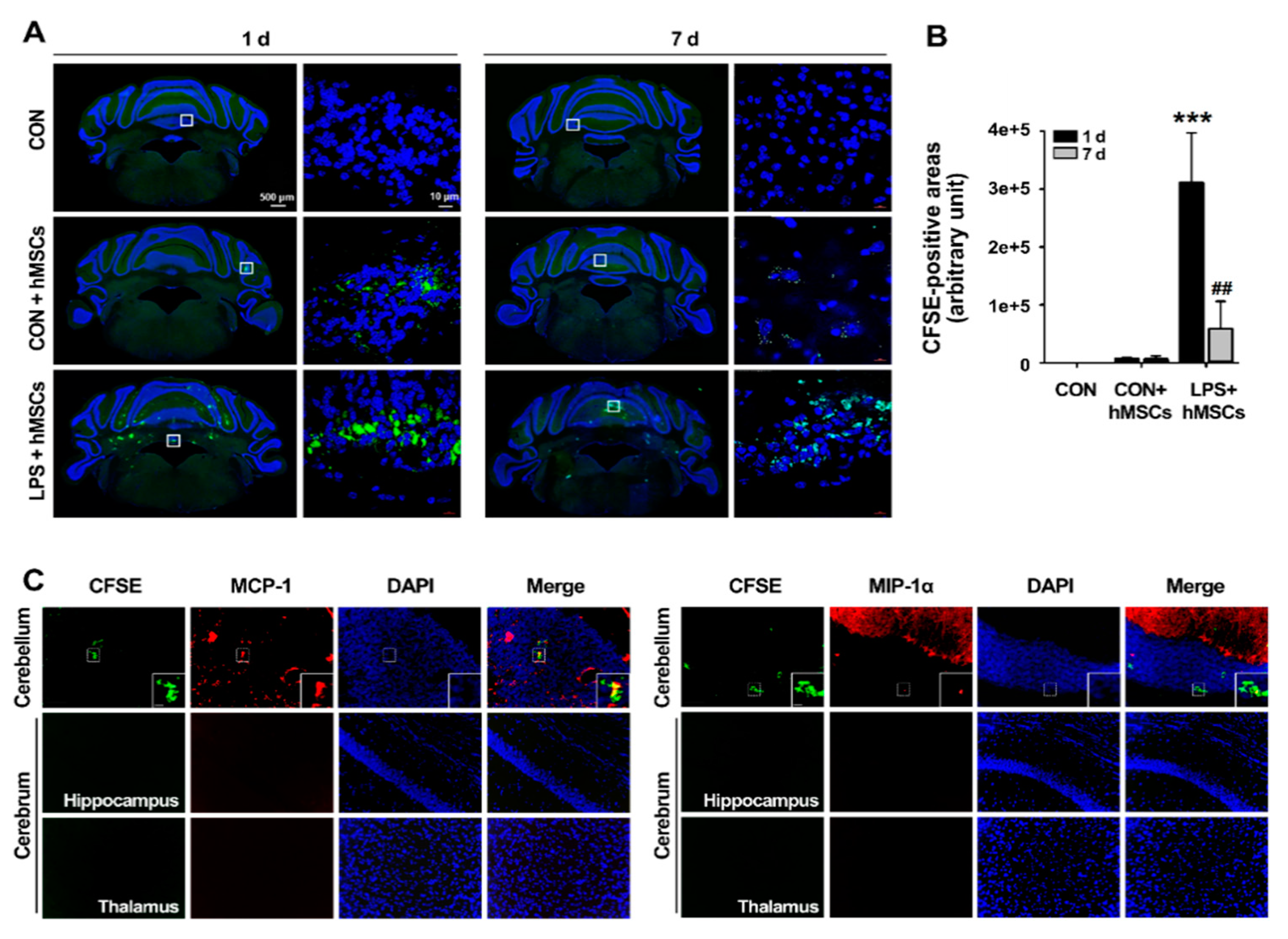
2.5. Carboxyfluorescein Succinimidyl Ester (CFSE) Labeling of hMSCs for Transplantation and Biodistribution of hMSCs in the Inflamed Cerebellum
2.6. Immunofluorescence Staining
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Characterization of hMSCs In Vitro
3.2. Migration of hMSCs into the LPS-Exposed Cerebellum after Intrathecal Implantation via the Cisterna Magna
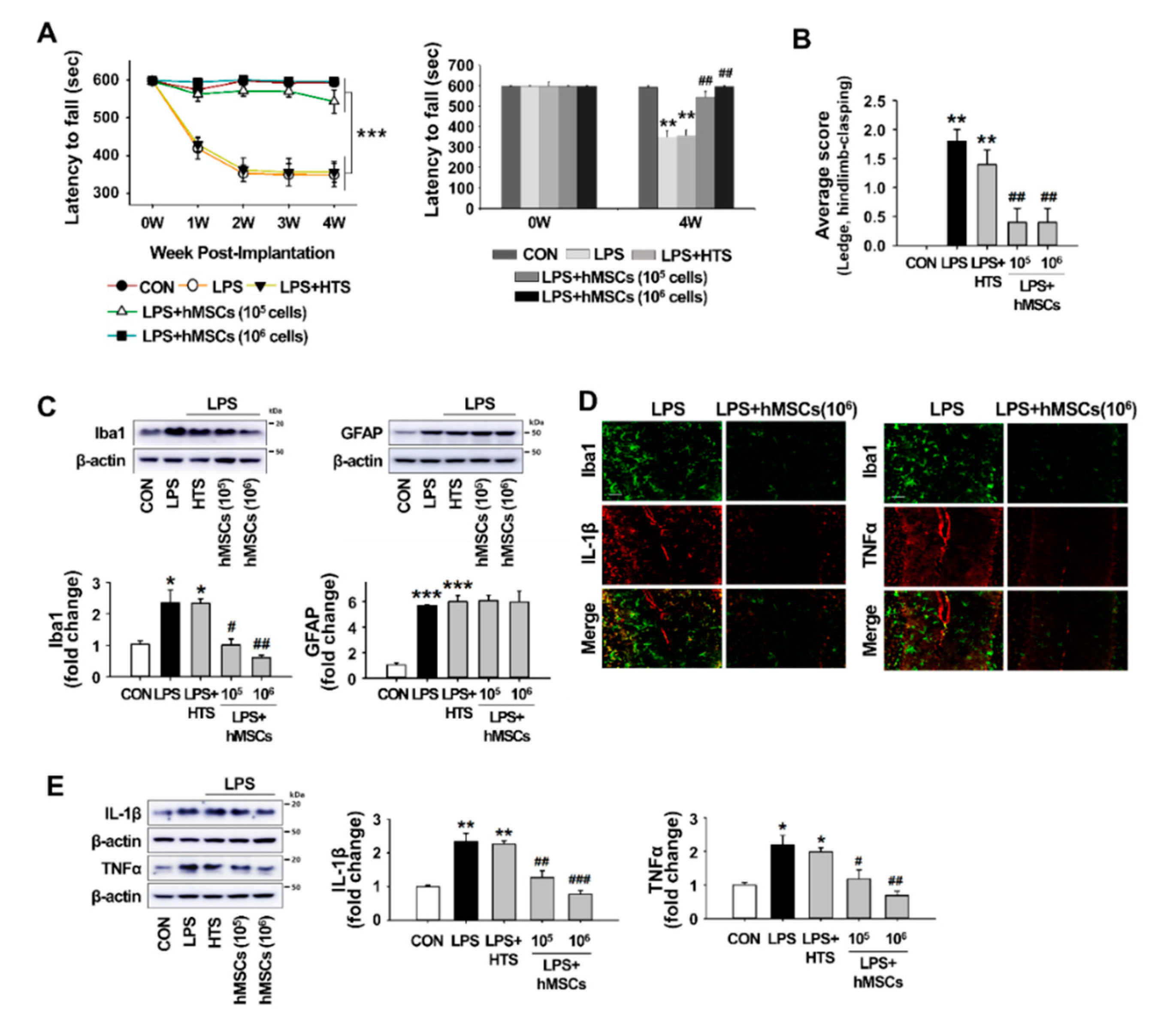
3.3. Effects of hMSCs Treatment on the Impaired Behaviors of the LPS-Induced ICA Mice
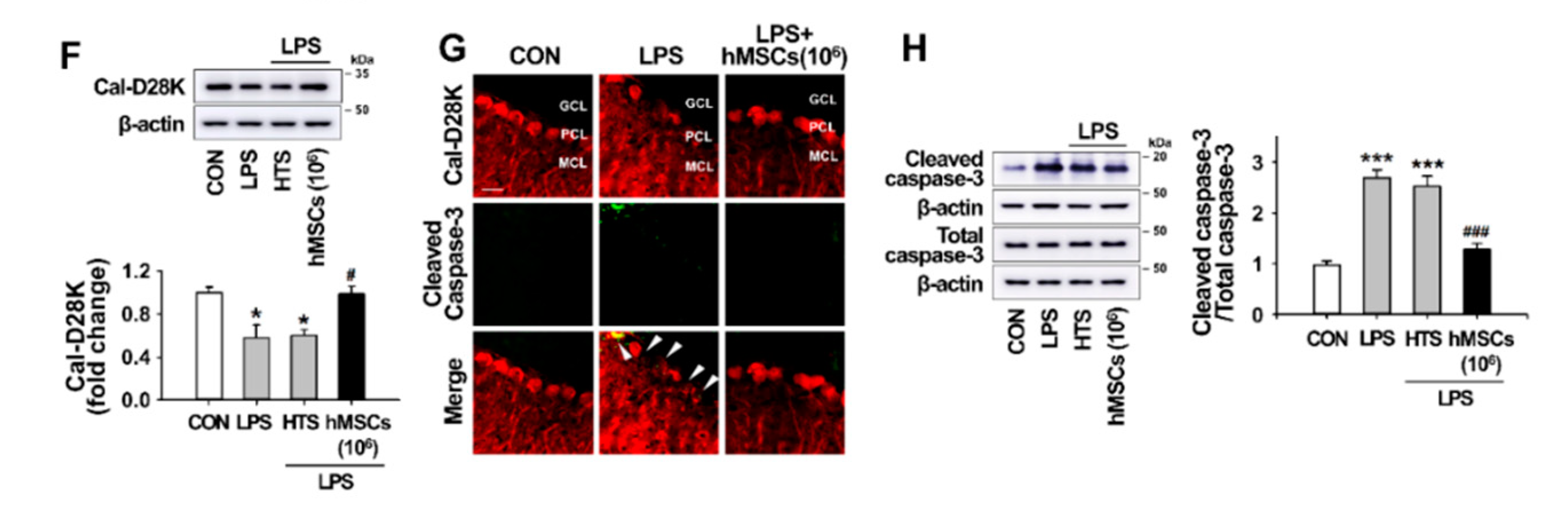
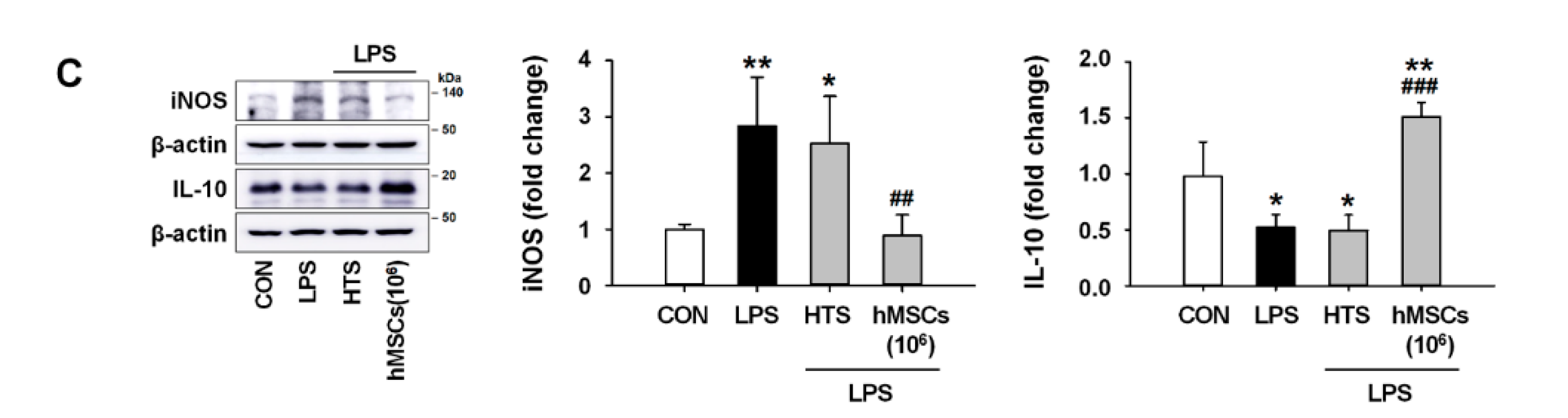
3.4. Effects of hMSC Transplantation on Neuroinflammation and Purkinje Cell Damage in the Cerebellum of ICA Mice
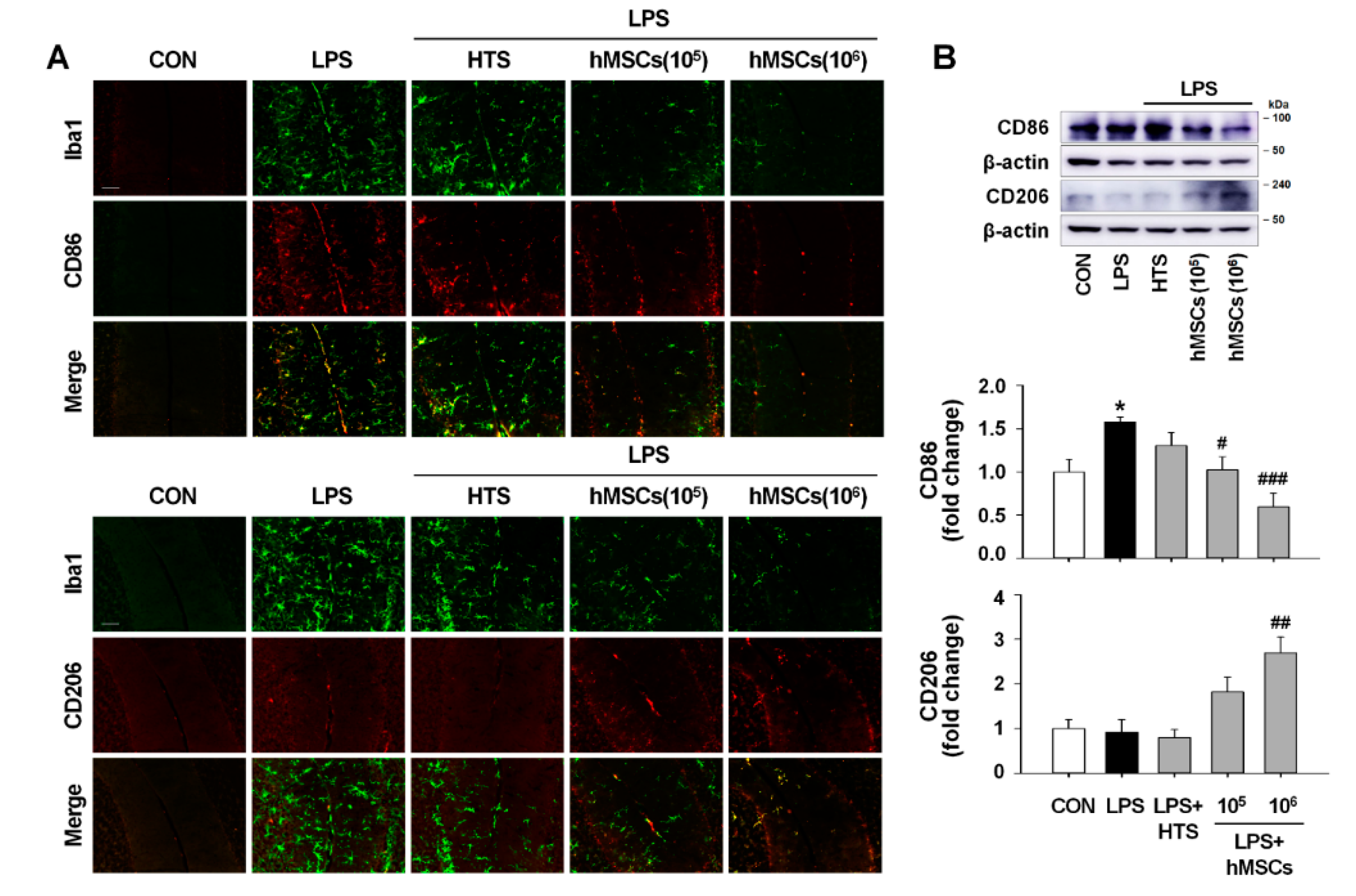
3.5. Effects of hMSCs on Microglial Polarization in the Cerebellum of LPS-Induced ICA Mouse
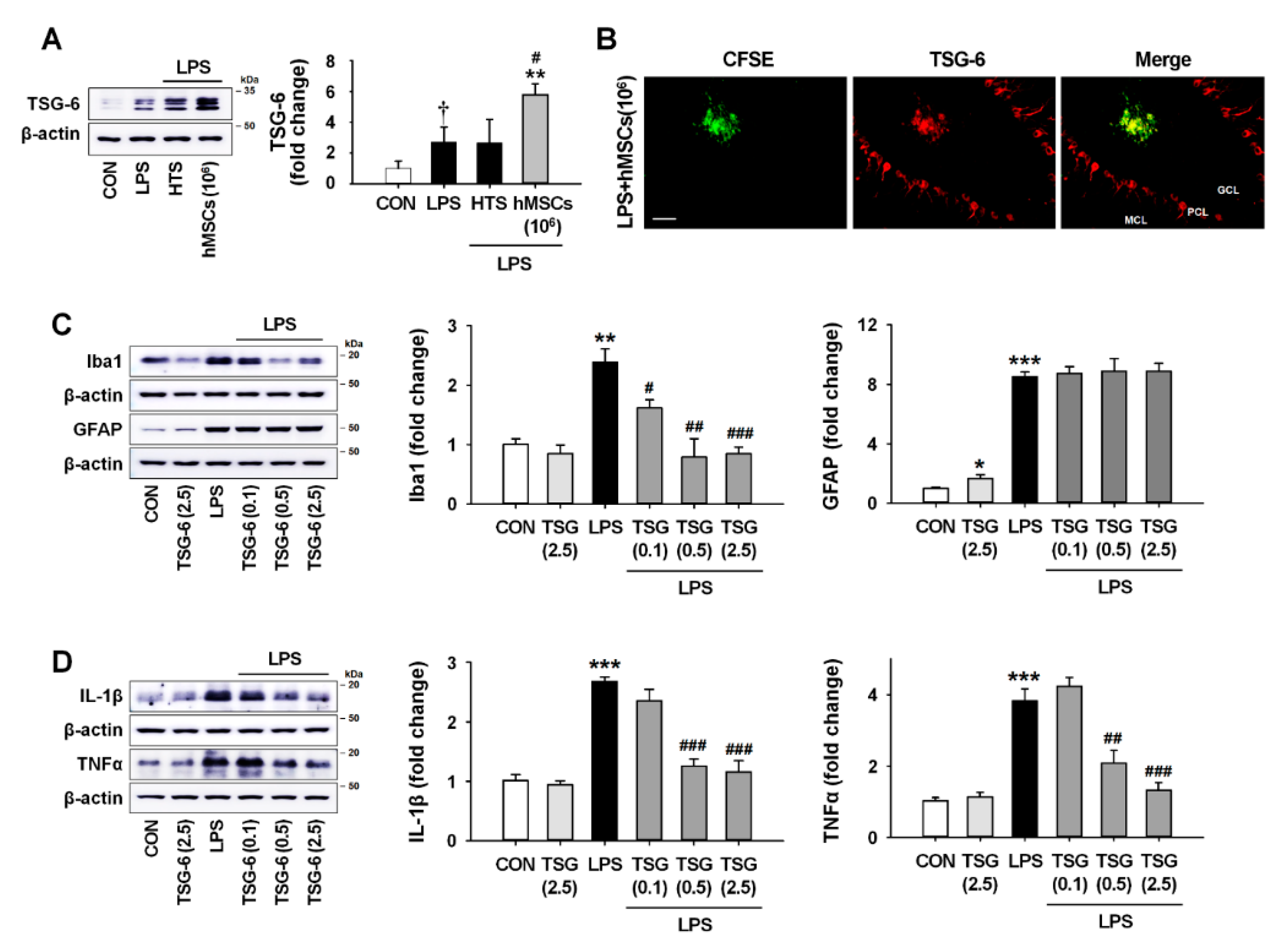
3.6. TSG-6 Upregulation by hMSC Treatment and its Anti-Inflammatory Effects in the Mouse Cerebellum
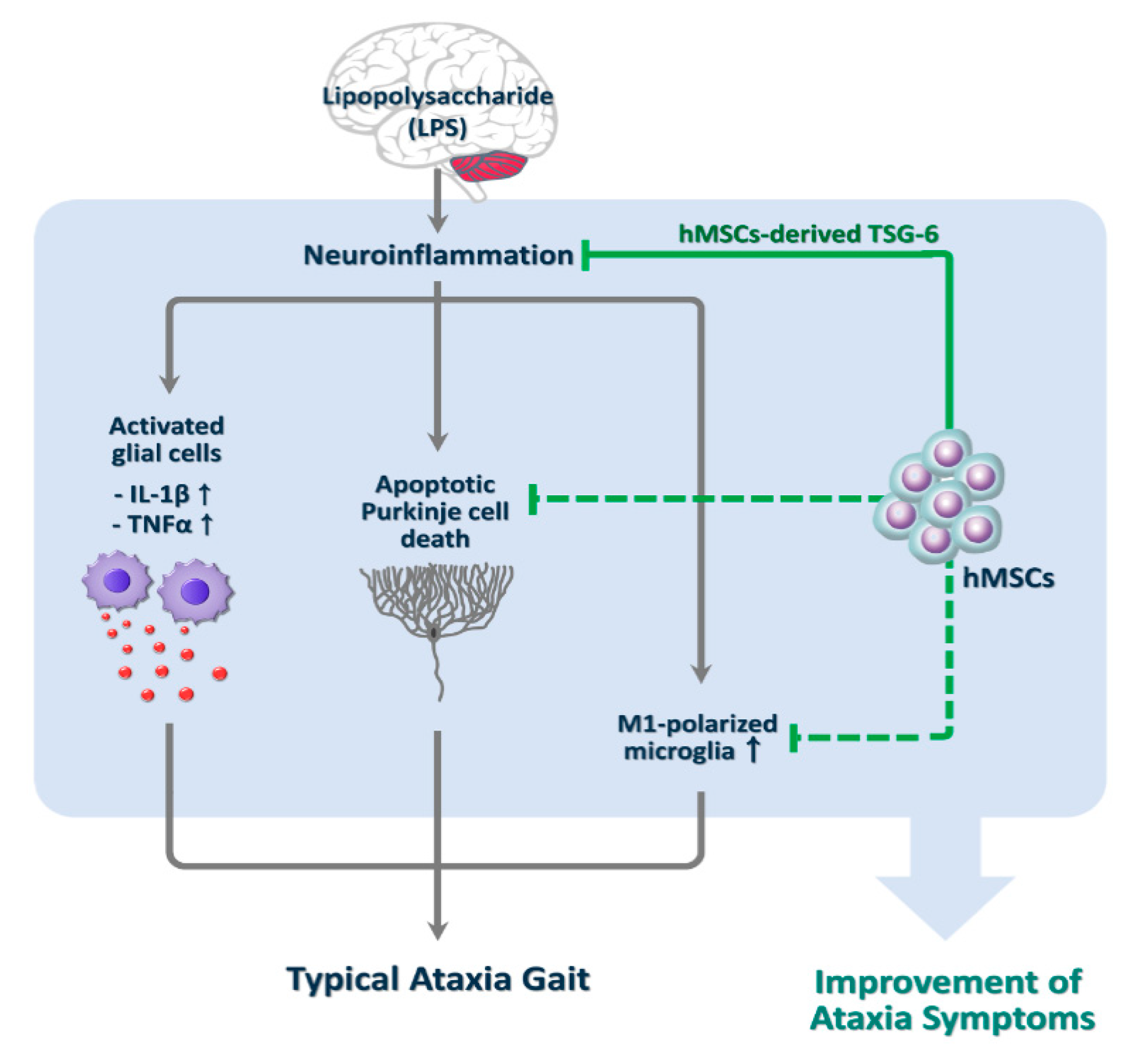
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klockgether, T. Sporadic ataxia with adult onset: Classification and diagnostic criteria. Lancet Neurol. 2010, 9, 94–104. [Google Scholar] [CrossRef]
- Teive, H.A. Non-progressive cerebellar ataxia with previous acute cerebellar injury of undetermined origin: A puzzling neurological disorder. Arq. Neuropsiquiatr. 2015, 73, 819–820. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Zamora, A.; Zeigler, W.; Desai, N.; Biller, J. Treatable causes of cerebellar ataxia. Mov. Disord. 2015, 30, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Gabsi, S.; Gouider-Khouja, N.; Belal, S.; Fki, M.; Kefi, M.; Turki, I.; Ben Hamida, M.; Kayden, H.; Mebazaa, R.; Hentati, F. Effect of vitamin E supplementation in patients with ataxia with vitamin E deficiency. Eur. J. Neurol. 2001, 8, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Hadjivassiliou, M.; Davies-Jones, G.A.; Sanders, D.S.; Grunewald, R.A. Dietary treatment of gluten ataxia. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- Fogel, B.L.; Perlman, S. An approach to the patient with late-onset cerebellar ataxia. Nat. Clin. Pract. Neurol. 2006, 2, 629–635. [Google Scholar] [CrossRef]
- Rossi, A.; Martinetti, C.; Morana, G.; Severino, M.; Tortora, D. Neuroimaging of Infectious and Inflammatory Diseases of the Pediatric Cerebellum and Brainstem. Neuroimaging Clin. N. Am. 2016, 26, 471–487. [Google Scholar] [CrossRef]
- Cvetanovic, M.; Ingram, M.; Orr, H.; Opal, P. Early activation of microglia and astrocytes in mouse models of spinocerebellar ataxia type 1. Neuroscience 2015, 289, 289–299. [Google Scholar] [CrossRef]
- Aikawa, T.; Mogushi, K.; Iijima-Tsutsui, K.; Ishikawa, K.; Sakurai, M.; Tanaka, H.; Mizusawa, H.; Watase, K. Loss of MyD88 alters neuroinflammatory response and attenuates early Purkinje cell loss in a spinocerebellar ataxia type 6 mouse model. Hum. Mol. Genet. 2015, 24, 4780–4791. [Google Scholar] [CrossRef]
- Evert, B.O.; Vogt, I.R.; Kindermann, C.; Ozimek, L.; de Vos, R.A.; Brunt, E.R.; Schmitt, I.; Klockgether, T.; Wullner, U. Inflammatory genes are upregulated in expanded ataxin-3-expressing cell lines and spinocerebellar ataxia type 3 brains. J. Neurosci. 2001, 21, 5389–5396. [Google Scholar] [CrossRef]
- Cutando, L.; Busquets-Garcia, A.; Puighermanal, E.; Gomis-Gonzalez, M.; Delgado-Garcia, J.M.; Gruart, A.; Maldonado, R.; Ozaita, A. Microglial activation underlies cerebellar deficits produced by repeated cannabis exposure. J. Clin. Investig. 2013, 123, 2816–2831. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Agarwalla, S.K.; Nanda, A.K. Case reports on atypical presentation of Plasmodium falciparum malaria in pediatric patients. J. Nat. Sci. Biol. Med. 2015, 6, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Marmolino, D.; Manto, M. Past, present and future therapeutics for cerebellar ataxias. Curr. Neuropharmacol. 2010, 8, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Yoon, D.; Nam, Y.; Seo, D.; Kim, J.H.; Kim, M.S.; Lee, T.Y.; Kim, K.S.; Ko, P.W.; Lee, H.W.; et al. Lipopolysaccharide administration for a mouse model of cerebellar ataxia with neuroinflammation. Sci. Rep. 2020, 10, 13337. [Google Scholar] [CrossRef]
- Sarugaser, R.; Hanoun, L.; Keating, A.; Stanford, W.L.; Davies, J.E. Human mesenchymal stem cells self-renew and differentiate according to a deterministic hierarchy. PLoS ONE 2009, 4, e6498. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, Y.; Bi, Y.; Liu, J.; Tan, B.; Gong, M.; Li, T.; Chen, J. Mesenchymal stem cells suppress neuronal apoptosis and decrease IL-10 release via the TLR2/NFkappaB pathway in rats with hypoxic-ischemic brain damage. Mol. Brain 2015, 8, 65. [Google Scholar] [CrossRef]
- Li, L.; Chopp, M.; Ding, G.L.; Qu, C.S.; Li, Q.J.; Lu, M.; Wang, S.; Nejad-Davarani, S.P.; Mahmood, A.; Jiang, Q. MRI measurement of angiogenesis and the therapeutic effect of acute marrow stromal cell administration on traumatic brain injury. J. Cereb. Blood Flow Metab. 2012, 32, 2023–2032. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Y.; Yan, K.; Chen, L.; Chen, X.R.; Li, P.; Chen, F.F.; Jiang, X.D. Anti-inflammatory and immunomodulatory mechanisms of mesenchymal stem cell transplantation in experimental traumatic brain injury. J. Neuroinflamm. 2013, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Joyce, N.; Annett, G.; Wirthlin, L.; Olson, S.; Bauer, G.; Nolta, J.A. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen. Med. 2010, 5, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Turgeman, G. The therapeutic potential of mesenchymal stem cells in Alzheimer’s disease: Converging mechanisms. Neural Regener. Res. 2015, 10, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Mesenchymal stem cell therapy in Parkinson’s disease animal models. Curr. Res. Transl. Med. 2017, 65, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Hsuan, Y.C.; Lin, C.H.; Chang, C.P.; Lin, M.T. Mesenchymal stem cell-based treatments for stroke, neural trauma, and heat stroke. Brain Behav. 2016, 6, e00526. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, S.; Shuvaev, A.N.; Iizuka, A.; Nakamura, K.; Hirai, H. Mesenchymal stem cells ameliorate cerebellar pathology in a mouse model of spinocerebellar ataxia type 1. Cerebellum 2014, 13, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Chen, M.H.; Chiang, Y.H.; Chen, Y.F.; Ma, W.H.; Tseng, C.Y.; Soong, B.W.; Ho, J.H.; Lee, O.K. Mesenchymal stem cell transplantation ameliorates motor function deterioration of spinocerebellar ataxia by rescuing cerebellar Purkinje cells. J. Biomed. Sci. 2011, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Sun, J.J.; Qian, L.; Liu, Z.; Zhang, Z.; Cao, W.; Li, W.; Xu, Y. Human umbilical mesenchymal stem cells enhance the expression of neurotrophic factors and protect ataxic mice. Brain Res. 2011, 1402, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.J.; Yeh, C.C.; Huang, W.J.; Min, M.Y.; Huang, T.H.; Ko, T.L.; Huang, P.Y.; Chen, T.H.; Hsu, S.P.C.; Soong, B.W.; et al. Xenografting of human umbilical mesenchymal stem cells from Wharton’s jelly ameliorates mouse spinocerebellar ataxia type 1. Transl. Neurodegener. 2019, 8, 29. [Google Scholar] [CrossRef]
- Sawada, Y.; Konno, A.; Nagaoka, J.; Hirai, H. Inflammation-induced reversible switch of the neuron-specific enolase promoter from Purkinje neurons to Bergmann glia. Sci. Rep. 2016, 6, 27758. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, H.; Oh, K.W.; Oh, S.I.; Koh, S.H.; Baik, W.; Noh, M.Y.; Kim, K.S.; Kim, S.H. Biological markers of mesenchymal stromal cells as predictors of response to autologous stem cell transplantation in patients with amyotrophic lateral sclerosis: An investigator-initiated trial and in vivo study. Stem Cells 2014, 32, 2724–2731. [Google Scholar] [CrossRef]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow, adipose tissue, and umbilical cord blood as sources of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef]
- Payne, N.L.; Dantanarayana, A.; Sun, G.; Moussa, L.; Caine, S.; McDonald, C.; Herszfeld, D.; Bernard, C.C.; Siatskas, C. Early intervention with gene-modified mesenchymal stem cells overexpressing interleukin-4 enhances anti-inflammatory responses and functional recovery in experimental autoimmune demyelination. Cell Adhes. Migr. 2012, 6, 179–189. [Google Scholar] [CrossRef]
- Wagner, W.; Wein, F.; Seckinger, A.; Frankhauser, M.; Wirkner, U.; Krause, U.; Blake, J.; Schwager, C.; Eckstein, V.; Ansorge, W.; et al. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp. Hematol. 2005, 33, 1402–1416. [Google Scholar] [CrossRef] [PubMed]
- Calio, M.L.; Marinho, D.S.; Ko, G.M.; Ribeiro, R.R.; Carbonel, A.F.; Oyama, L.M.; Ormanji, M.; Guirao, T.P.; Calio, P.L.; Reis, L.A.; et al. Transplantation of bone marrow mesenchymal stem cells decreases oxidative stress, apoptosis, and hippocampal damage in brain of a spontaneous stroke model. Free Radic. Biol. Med. 2014, 70, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Hong, J.; Moon, G.J.; Jung, U.J.; Won, S.Y.; Kim, S.R. Morin Prevents Granule Cell Dispersion and Neurotoxicity via Suppression of mTORC1 in a Kainic Acid-induced Seizure Model. Exp. Neurobiol. 2018, 27, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jung, U.J.; Oh, Y.S.; Jeon, M.T.; Kim, H.J.; Shin, W.H.; Hong, J.; Kim, S.R. Beneficial Effects of Silibinin Against Kainic Acid-induced Neurotoxicity in the Hippocampus in vivo. Exp. Neurobiol. 2017, 26, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-kappaB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef]
- Lee, R.H.; Yu, J.M.; Foskett, A.M.; Peltier, G.; Reneau, J.C.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. TSG-6 as a biomarker to predict efficacy of human mesenchymal stem/progenitor cells (hMSCs) in modulating sterile inflammation in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 16766–16771. [Google Scholar] [CrossRef]
- Day, A.J.; Milner, C.M. TSG-6: A multifunctional protein with anti-inflammatory and tissue-protective properties. Matrix Biol. 2019, 78–79, 60–83. [Google Scholar] [CrossRef]
- Li, R.; Liu, W.; Yin, J.; Chen, Y.; Guo, S.; Fan, H.; Li, X.; Zhang, X.; He, X.; Duan, C. TSG-6 attenuates inflammation-induced brain injury via modulation of microglial polarization in SAH rats through the SOCS3/STAT3 pathway. J. Neuroinflamm. 2018, 15, 231. [Google Scholar] [CrossRef]
- Fogel, B.L. Childhood cerebellar ataxia. J. Child Neurol. 2012, 27, 1138–1145. [Google Scholar] [CrossRef]
- Manto, M. Toxic agents causing cerebellar ataxias. Handb. Clin. Neurol. 2012, 103, 201–213. [Google Scholar] [PubMed]
- Shanmugarajah, P.D.; Hoggard, N.; Currie, S.; Aeschlimann, D.P.; Aeschlimann, P.C.; Gleeson, D.C.; Karajeh, M.; Woodroofe, N.; Grunewald, R.A.; Hadjivassiliou, M. Alcohol-related cerebellar degeneration: Not all down to toxicity? Cerebellum Ataxias 2016, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Binan, L.; Ajji, A.; De Crescenzo, G.; Jolicoeur, M. Approaches for neural tissue regeneration. Stem Cell Rev. Rep. 2014, 10, 44–59. [Google Scholar] [CrossRef]
- Sakthiswary, R.; Raymond, A.A. Stem cell therapy in neurodegenerative diseases: From principles to practice. Neural Regener. Res. 2012, 7, 1822–1831. [Google Scholar]
- Sarukhan, A.; Zanotti, L.; Viola, A. Mesenchymal stem cells: Myths and reality. Swiss Med. Wkly. 2015, 145, w14229. [Google Scholar] [CrossRef]
- Belema-Bedada, F.; Uchida, S.; Martire, A.; Kostin, S.; Braun, T. Efficient homing of multipotent adult mesenchymal stem cells depends on FROUNT-mediated clustering of CCR2. Cell Stem Cell 2008, 2, 566–575. [Google Scholar] [CrossRef]
- Song, C.H.; Honmou, O.; Furuoka, H.; Horiuchi, M. Identification of chemoattractive factors involved in the migration of bone marrow-derived mesenchymal stem cells to brain lesions caused by prions. J. Virol. 2011, 85, 11069–11078. [Google Scholar] [CrossRef]
- Jiang, L.; Newman, M.; Saporta, S.; Chen, N.; Sanberg, C.; Sanberg, P.R.; Willing, A.E. MIP-1alpha and MCP-1 Induce Migration of Human Umbilical Cord Blood Cells in Models of Stroke. Curr. Neurovasc. Res. 2008, 5, 118–124. [Google Scholar] [CrossRef][Green Version]
- Yun, H.M.; Kim, H.S.; Park, K.R.; Shin, J.M.; Kang, A.R.; Il Lee, K.; Song, S.; Kim, Y.B.; Han, S.B.; Chung, H.M.; et al. Placenta-derived mesenchymal stem cells improve memory dysfunction in an Abeta1-42-infused mouse model of Alzheimer’s disease. Cell Death Dis. 2013, 4, e958. [Google Scholar] [CrossRef]
- Rossignol, J.; Fink, K.; Davis, K.; Clerc, S.; Crane, A.; Matchynski, J.; Lowrance, S.; Bombard, M.; Dekorver, N.; Lescaudron, L.; et al. Transplants of adult mesenchymal and neural stem cells provide neuroprotection and behavioral sparing in a transgenic rat model of Huntington’s disease. Stem Cells 2014, 32, 500–509. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, H.J.; Lee, G.; Bang, O.Y.; Ahn, Y.H.; Joe, E.; Kim, H.O.; Lee, P.H. Neuroprotective effects of human mesenchymal stem cells on dopaminergic neurons through anti-inflammatory action. Glia 2009, 57, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, M.Y.; Zhang, D.F.; Zhong, X.; Du, K.; Qian, P.; Gao, H.; Wei, M.J. Natural products as a potential modulator of microglial polarization in neurodegenerative diseases. Pharmacol. Res. 2019, 145, 104253. [Google Scholar] [CrossRef]
- Kudlik, G.; Hegyi, B.; Czibula, A.; Monostori, E.; Buday, L.; Uher, F. Mesenchymal stem cells promote macrophage polarization toward M2b-like cells. Exp. Cell Res. 2016, 348, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, R.; Yan, K.; Chen, F.; Huang, W.; Lv, B.; Sun, C.; Xu, L.; Li, F.; Jiang, X. Mesenchymal stem cells inhibit lipopolysaccharide-induced inflammatory responses of BV2 microglial cells through TSG-6. J. Neuroinflamm. 2014, 11, 135. [Google Scholar] [CrossRef]
- Ichiseki, T.; Shimazaki, M.; Ueda, Y.; Ueda, S.; Tsuchiya, M.; Souma, D.; Kaneuji, A.; Kawahara, N. Intraarticularly-Injected Mesenchymal Stem Cells Stimulate Anti-Inflammatory Molecules and Inhibit Pain Related Protein and Chondrolytic Enzymes in a Monoiodoacetate-Induced Rat Arthritis Model. Int. J. Mol. Sci. 2018, 19, 203. [Google Scholar] [CrossRef]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef]
- Qi, Y.; Jiang, D.; Sindrilaru, A.; Stegemann, A.; Schatz, S.; Treiber, N.; Rojewski, M.; Schrezenmeier, H.; Vander Beken, S.; Wlaschek, M.; et al. TSG-6 released from intradermally injected mesenchymal stem cells accelerates wound healing and reduces tissue fibrosis in murine full-thickness skin wounds. J. Investig. Dermatol. 2014, 134, 526–537. [Google Scholar] [CrossRef]
- Kong, D.; Zhu, J.; Liu, Q.; Jiang, Y.; Xu, L.; Luo, N.; Zhao, Z.; Zhai, Q.; Zhang, H.; Zhu, M.; et al. Mesenchymal stem cells protect neurons against hypoxic-ischemic injury via inhibiting parthanatos, necroptosis, and apoptosis, but not autophagy. Cell. Mol. Neurobiol. 2017, 37, 303–313. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Name | Source | Identifier | Dilution |
|---|---|---|---|
| Rabbit anti-ionized calcium-binding adapter molecule 1 | Wako Pure Chemical Industries, Osaka, Japan | 019-19741 | IF 1:2000 |
| WB 1:1500 | |||
| Rabbit anti-glial fibrillary acidic protein | Millipore, Billerica, MA, USA | AB5804 | IF 1:2000 |
| WB 1:2000 | |||
| Rabbit anti-interleukin 1 beta | Abcam, Cambridge, UK | Ab9722 | IF 1:500 |
| WB 1:1000 | |||
| Mouse anti-tumor necrosis factor alpha | Abcam, Cambridge, UK | Ab6671 | IF 1:500 |
| WB 1:1000 | |||
| Rat anti-monocyte chemoattractant protein 1 | Abcam, Cambridge, UK | Ab8101 | IF 1:100 |
| WB 1:500 | |||
| Rat anti-macrophage inflammatory protein 1 alpha | R&D Systems, Minneapolis, MN, USA | MAB4501 | IF 1:100, |
| WB 1:1000 | |||
| Mouse anti-calbindin-D-28K | Sigma, St. Louis, MO, USA | C9848 | IF 1:500 |
| WB 1:2000 | |||
| Rabbit anti-cleaved caspase-3 | Cell Signaling, Beverly, MA, USA | CST 9664 | IF 1:400 |
| WB 1:1000 | |||
| Rabbit anti-caspase-3 | Cell Signaling, Beverly, MA, USA | CST 9662 | WB 1:1000 |
| Rabbit anti-cluster of differentiation 86 | Invitrogen, Carlsbad, CA, USA | 14-0863-81 | IF 1:200, |
| WB 1:1000 | |||
| Goat anti-cluster of differentiation 206 | R&D Systems, Minneapolis, MN, USA | AF2535 | IF 1:200 |
| WB 1:1000 | |||
| Rabbit anti-inducible nitric oxide synthase | Abcam, Cambridge, UK | Ab3523 | WB 1:1000 |
| Rabbit anti-interleukin 10 | Abcam, Cambridge, MA, USA | Ab9969 | WB 1:1000 |
| Rabbit anti-TNFα stimulated gene-6 | GeneTex, Irvine, CA, USA | GTX55175 | WB 1:1000 |
| Mouse anti-β-actin | Santa cruz biotechnology, Santa Cruz, CA, USA | sc-47778 | WB 1:2000 |
| Texas Red-conjugated anti-mouse IgG antibody | Vector Laboratories, Burlingame, CA, USA | TI-2000 | IF 1:400 |
| Texas Red-conjugated anti-Goat IgG antibody | Vector Laboratories, Burlingame, CA, USA | TI-5000 | IF 1:400 |
| FITC-labeled IgG antibody | Jackson ImmunoResearch, West Grove, PA, USA | FI-2000 | IF 1:400 |
| horseradish peroxidase (HRP)-conjugated antibody | Thermo Fisher Scientific, Piscataway, NJ, USA | 61-6520 | WB 1:4000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, Y.; Yoon, D.; Hong, J.; Kim, M.S.; Lee, T.Y.; Kim, K.S.; Lee, H.-W.; Suk, K.; Kim, S.R. Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation. J. Clin. Med. 2020, 9, 3654. https://doi.org/10.3390/jcm9113654
Nam Y, Yoon D, Hong J, Kim MS, Lee TY, Kim KS, Lee H-W, Suk K, Kim SR. Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation. Journal of Clinical Medicine. 2020; 9(11):3654. https://doi.org/10.3390/jcm9113654
Chicago/Turabian StyleNam, Youngpyo, Dongyeong Yoon, Jungwan Hong, Min Sung Kim, Tae Yong Lee, Kyung Suk Kim, Ho-Won Lee, Kyoungho Suk, and Sang Ryong Kim. 2020. "Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation" Journal of Clinical Medicine 9, no. 11: 3654. https://doi.org/10.3390/jcm9113654
APA StyleNam, Y., Yoon, D., Hong, J., Kim, M. S., Lee, T. Y., Kim, K. S., Lee, H.-W., Suk, K., & Kim, S. R. (2020). Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation. Journal of Clinical Medicine, 9(11), 3654. https://doi.org/10.3390/jcm9113654