Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Experimental Section
2.1. Patients
Ethics Consideration
2.2. Samples
2.3. Bioinformatics Analysis
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
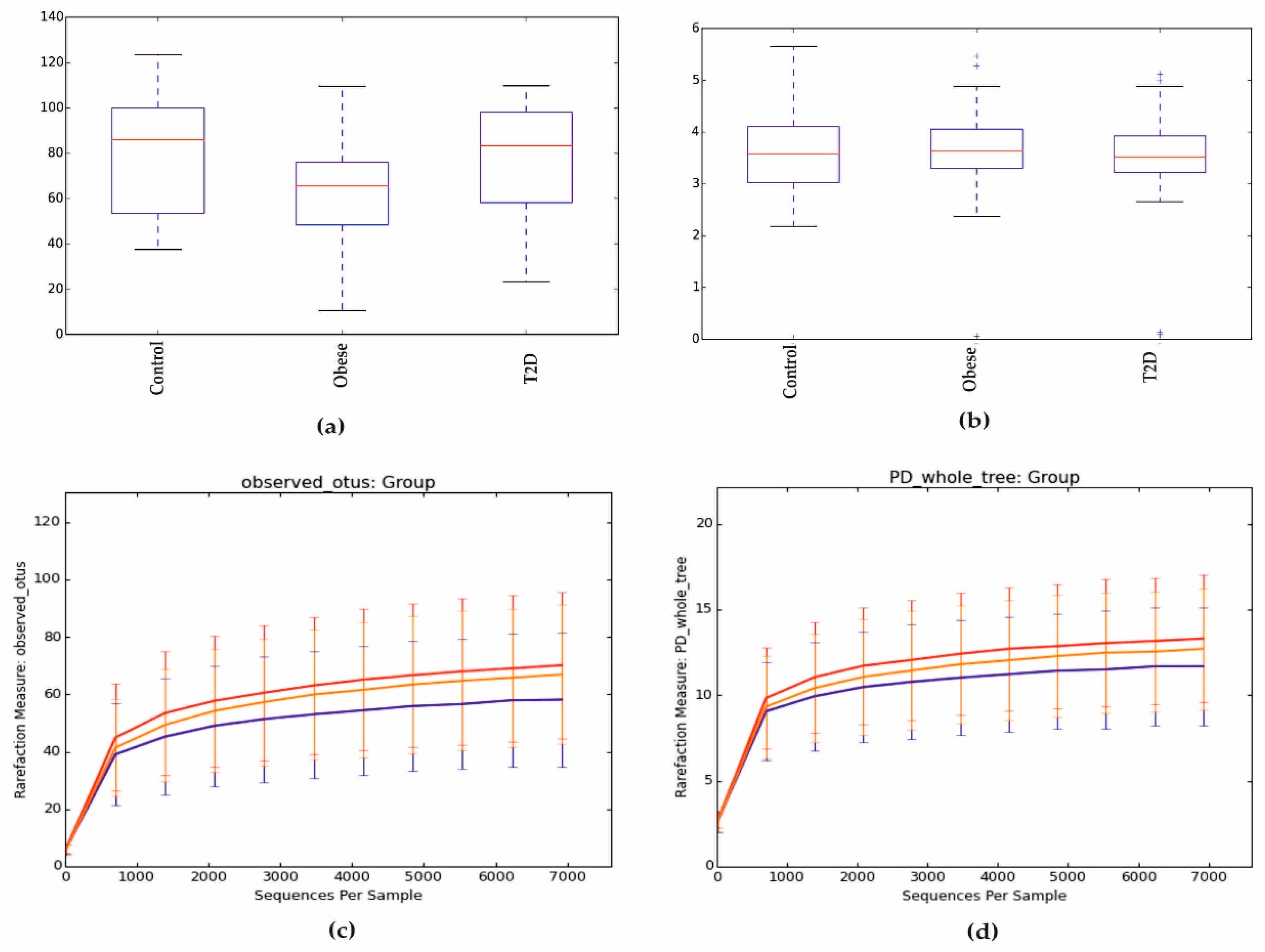
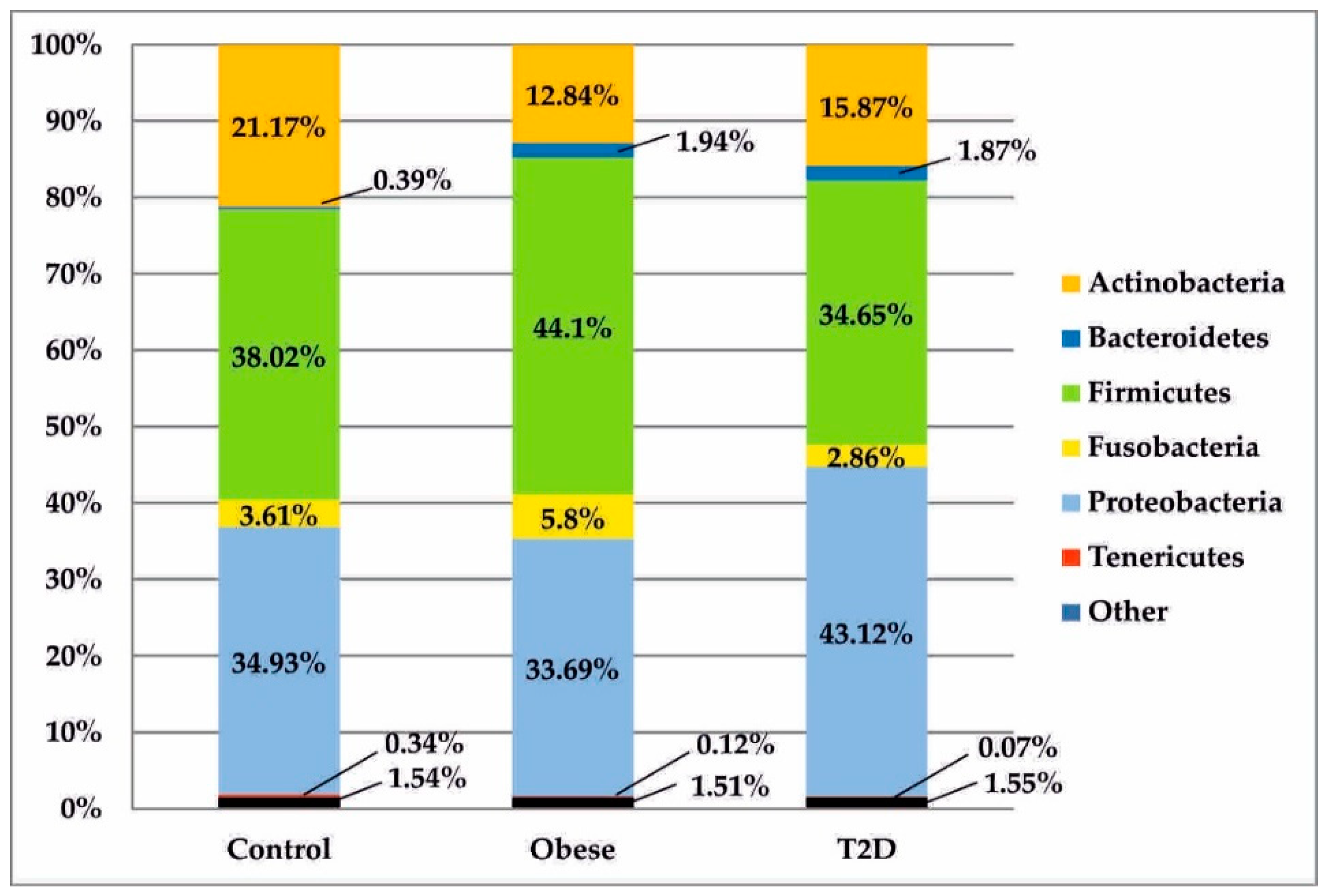
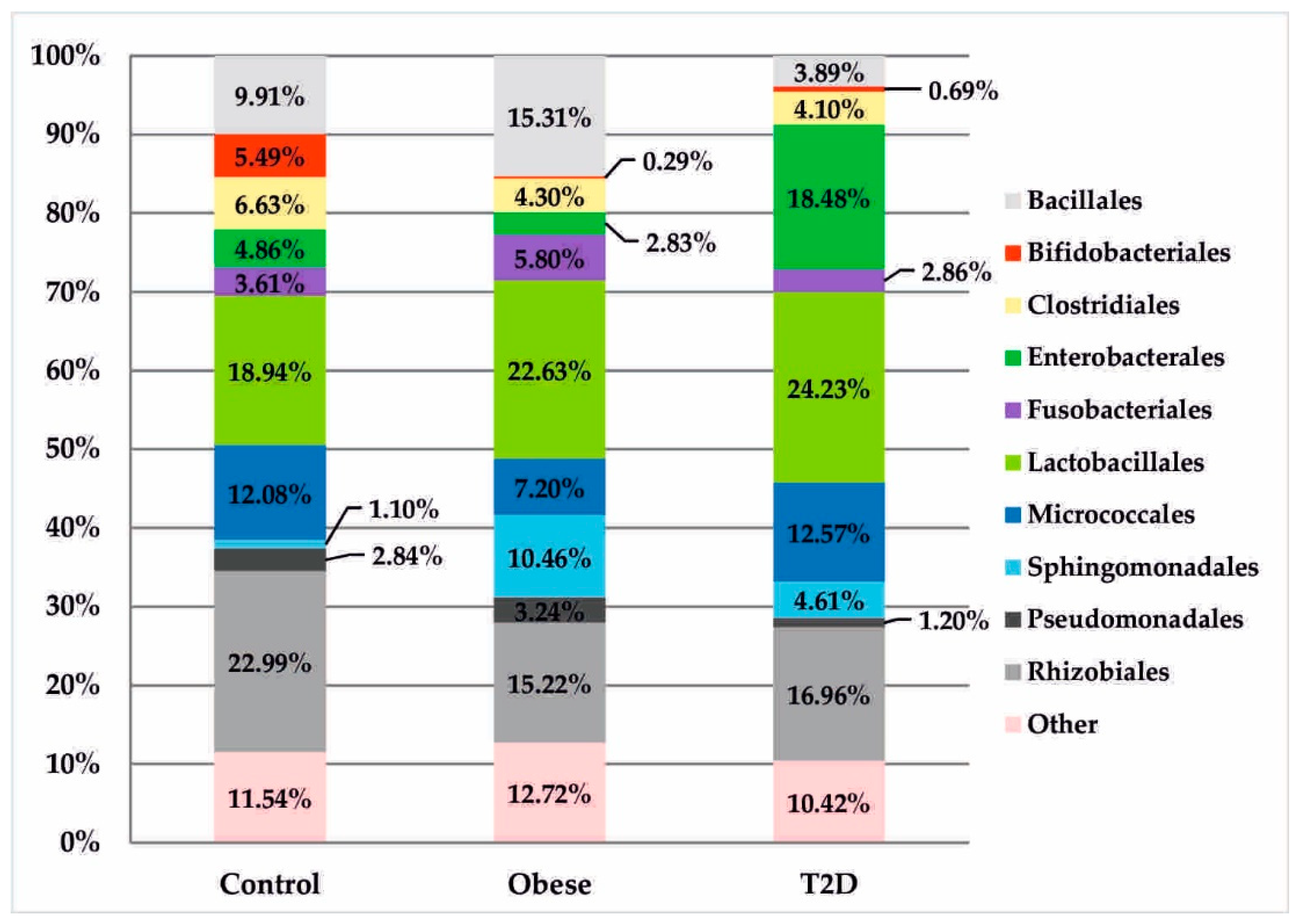
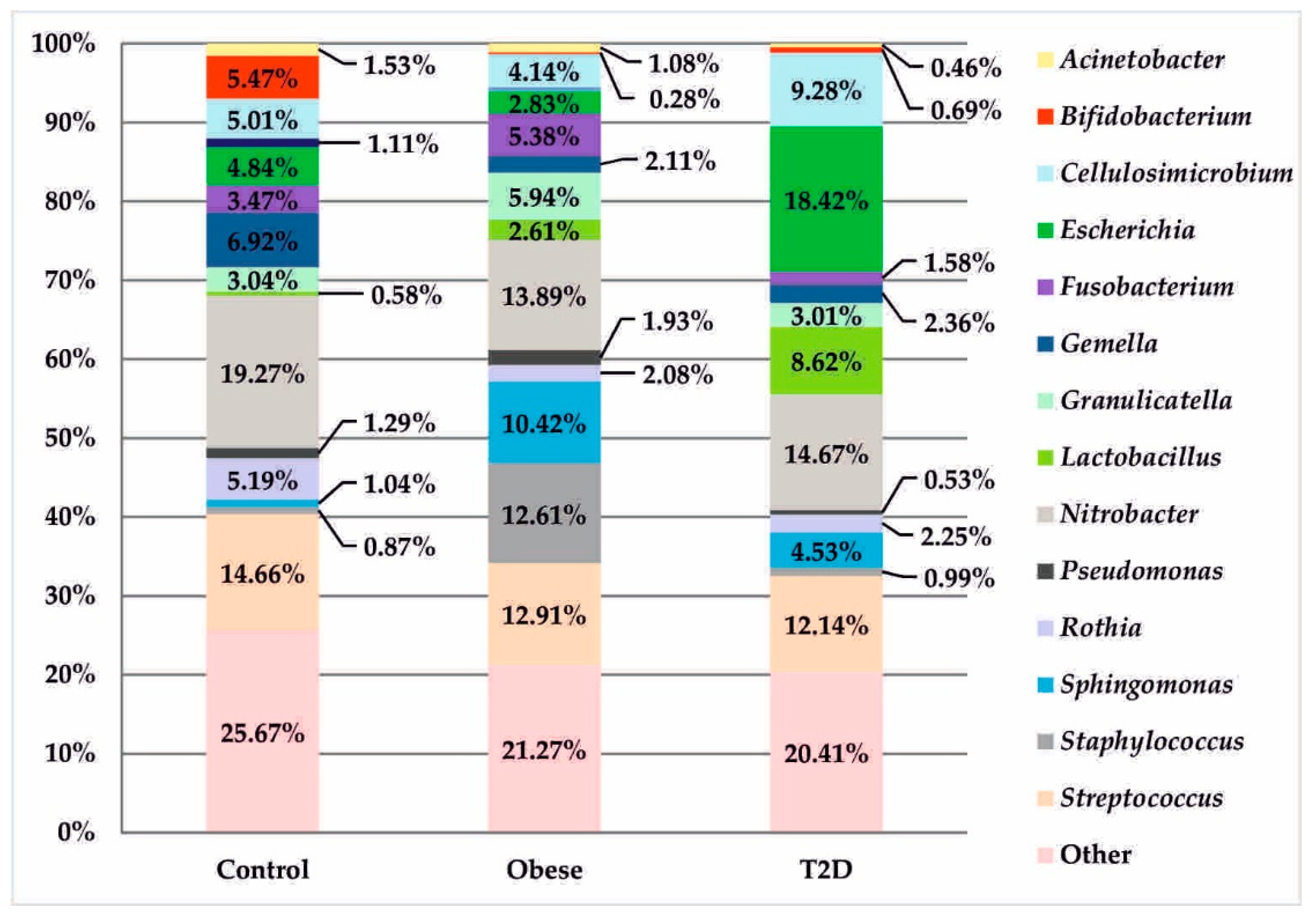
3.2. Metagenomic Sequencing
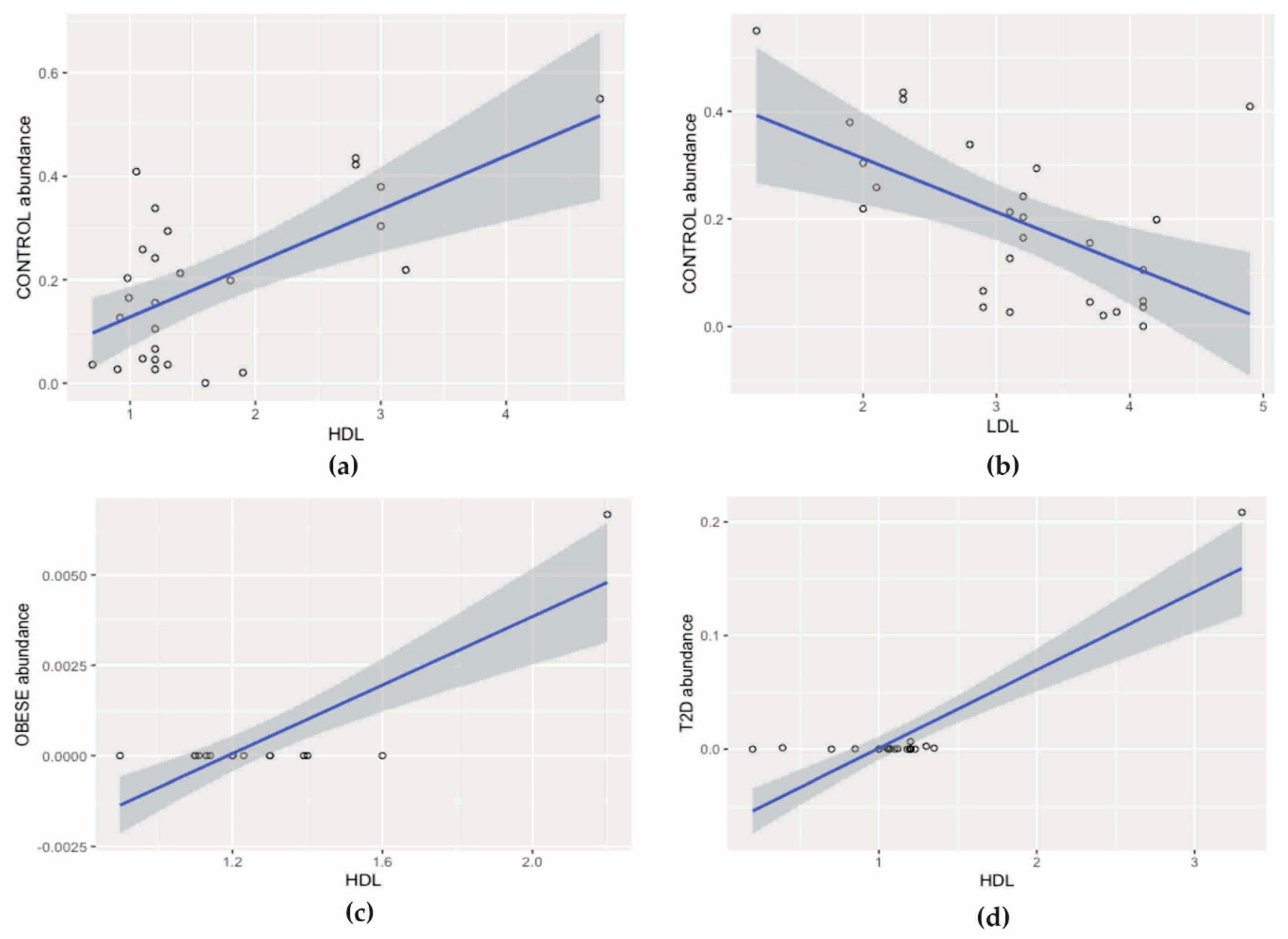
3.3. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization Home Page. Available online: https://apps.who.int/iris/bitstream /handle/10665/204871/9789241565257_eng.pdf;jsessionid=DF43C80C9B15F5093C9F74ABF9574E35?sequence=1 (accessed on 18 December 2019).
- International Diabetes Federation Home Page. Available online: https://www.idf.org/e-library/epidemiology-research/diabetes-atlas/13-diabetes-atlas-seventh-edition.html (accessed on 18 December 2019).
- Wang, Z.-K.; Yang, Y.-S. Upper gastrointestinal microbiota and digestive diseases. World J. Gastroenterol. 2013, 19, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yang, M.; Zhou, K.; Zhang, L.; Tian, L.; Lv, S.; Jin, Y.; Qian, W.; Xiong, H.; Lin, R.; et al. Diversity of Duodenal and Rectal Microbiota in Biopsy Tissues and Luminal Contents in Healthy Volunteers. J. Microbiol. Biotechnol. 2015, 25, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLOS Biology 2016, 14, 036103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, E.; Donat, E.; Ribes-Koninckx, C.; Fernández-Murga, M.L.; Sanz, Y. Duodenal-Mucosal Bacteria Associated with Celiac Disease in Children. Appl. Environ. Microbiol. 2013, 79, 5472–5479. [Google Scholar] [CrossRef] [Green Version]
- Schippa, S.; Iebba, V.; Barbato, M.; Di Nardo, G.; Totino, V.; Checchi, M.P.; Longhi, C.; Maiella, G.; Cucchiara, S.; Conte, M.P. A distinctive ’microbial signature’ in celiac pediatric patients. BMC Microbiol. 2010, 10, 175. [Google Scholar] [CrossRef]
- Angelakis, E.; Armougom, F.; Carrière, F.; Bachar, D.; Laugier, R.; Lagier, J.-C.; Robert, C.; Michelle, C.; Henrissat, B.; Raoult, D. A Metagenomic Investigation of the Duodenal Microbiota Reveals Links with Obesity. PLoS ONE 2015, 10, e0137784. [Google Scholar] [CrossRef]
- Gosiewski, T.; Szała, L.; Pietrzyk, A.; Brzychczy-Włoch, M.; Heczko, P.B.; Bulanda, M. Comparison of Methods for Isolation of Bacterial and Fungal DNA from Human Blood. Curr. Microbiol. 2014, 68, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bik, E.M.; Eckburg, P.B.; Gill, S.R.; Nelson, K.E.; Purdom, E.A.; Francois, F.; Perez, G.P.; Blaser, M.J.; Relman, D.A. Molecular analysis of the bacterial microbiota in the human stomach. Proc. Natl. Acad. Sci. USA 2006, 103, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Pussinen, P.J.; Havulinna, A.S.; Lehto, M.; Sundvall, J.; Salomaa, V. Endotoxemia Is Associated With an Increased Risk of Incident Diabetes. Diabetes Care 2011, 34, 392–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrozinska, S.; Radkowski, P.; Gosiewski, T.; Szopa, M.; Bulanda, M.; Ludwig-Galezowska, A.H.; Morawska, I.; Sroka-Oleksiak, A.; Matejko, B.; Kapusta, P.; et al. Qualitative Parameters of the Colonic Flora in Patients with HNF1A-MODY Are Different from Those Observed in Type 2 Diabetes Mellitus. J. Diabetes Res. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Le Bastard, Q.; Al-Ghalith, G.A.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic review: Human gut dysbiosis induced by non-antibiotic prescription medications. Aliment. Pharmacol. Ther. 2018, 47, 332–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amar, J.; Chabo, C.; Waget, A.; Klopp, P.; Vachoux, C.; Bermúdez-Humarán, L.G.; Smirnova, N.; Bergé, M.; Sulpice, T.; Lahtinen, S.; et al. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: Molecular mechanisms and probiotic treatment: Bacterial translocation during diabetes. EMBO Mol. Med. 2011, 3, 559–572. [Google Scholar] [CrossRef]
- Stenman, L.K.; Waget, A.; Garret, C.; Klopp, P.; Burcelin, R.; Lehinen, M.J. Potential probiotic Bifidobacterium animalis ssp. lactis 420 prevents weight gain and glucose intolerance in diet-induced obese mice. Benef. Microbes 2014, 5, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Stenman, L.K.; Lehtinen, M.J.; Meland, N.; Christensen, J.E.; Yeung, N.; Saarinen, M.T.; Courtney, M.; Burcelin, R.; Lähdeaho, M.-L.; Linros, J.; et al. Probiotic with or without Fiber Controls Body Fat Mass, Associated With Serum Zonulin, in Overweight and Obese Adults—Randomized Controlled Trial. EBioMedicine 2016, 13, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Stenman, L.K.; Waget, A.; Garret, C.; Briand, F.; Burcelin, R.; Sulpice, T.; Lahtinen, S. Probiotic B420 and prebiotic polydextrose improve efficacy of antidiabetic drugs in mice. Diabetol. Metab. Syndr. 2015, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in Lean and Overweight Healthy Subjects. Obes. 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Salamon, D.; Sroka, A.; Kapusta, P.; Szopa, M.; Mrozińska, S.; Ludwig, A.H.; Wołkow, P.P.; Bulanda, M.; Klupa, T.; Małecki, M.T.; et al. Characteristics of gut microbiota in adult patients with type 1 and type 2 diabetes based on next-generation sequencing of the 16S rRNA gene fragment. Pol. Arch. Intern. Med. 2018, 128, 6. [Google Scholar] [CrossRef]
- An, H.M.; Park, S.Y.; Lee, D.K.; Kim, J.R.; Cha, M.K.; Lee, S.W.; Lim, H.T.; Kim, K.J.; Ha, N.J. Antiobesity and lipid-lowering effects of Bifidobacterium spp. in high fat diet-induced obese rats. Lipids Heal. Dis. 2011, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Ichim, T.E.; Patel, A.N.; Shafer, K.A. Experimental support for the effects of a probiotic/digestive enzyme supplement on serum cholesterol concentrations and the intestinal microbiome. J. Transl. Med. 2016, 14, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kießling, G.; Schneider, J.; Jahreis, G. Long-term consumption of fermented dairy products over 6 months increases HDL cholesterol. Eur. J. Clin. Nutr. 2002, 56, 843–849. [Google Scholar] [CrossRef] [Green Version]
- Hassanein, W.A.; Awny, N.M.; Ibraheim, S.M. Cholesterol reduction by Lactococcus lactis KF147. Afr J. Microbiol Res. 2013, 7, 4338–4349. [Google Scholar]
- Lee, W.-K.; Lim, H.-J.; Kim, S.-Y.; Kimoto, H.; Ohmomo, S.; Tashiro, Y.; Takebe, H. Hypocholesterolemic Effect of Lactococcus lactis subsp. lactis biovar diacetylactis N7 and Lactococcus lactis subsp. lactis 527 Strains in SD Rats. Biosci. Microflora 2005, 24, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Yadav, K.; Bhardwaj, A.; Kaur, G.; Iyer, R.; De, S.; Malik, R.K. Potential of Lactococcus lactis as a probiotic and functional lactic acid bacteria in dairy industry. Int. J. Probiotics Prebiotics. 2009, 4, 219–228. [Google Scholar]
- Angelakis, E.; Merhej, V.; Raoult, D. Related actions of probiotics and antibiotics on gut microbiota and weight modification. Lancet Infect. Dis. 2013, 13, 889–899. [Google Scholar] [CrossRef]
- Murphy, E.F.; Cotter, P.D.; Hogan, A.; O’Sullivan, O.; Joyce, A.; Fouhy, F.; Clarke, S.F.; Marques, T.M.; O’Toole, P.W.; Stanton, C.; et al. Divergent metabolic outcomes arising from targeted manipulation of the gut microbiota in diet-induced obesity. Gut. 2013, 62, 220–226. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Marlicz, W.; Ostrowska, L.; Łoniewski, I. Intestinal microbiota and its potential relationship with obesity. Endokrynol. Otył. Zab. Przem. Mat. 2013, 9, 20–28. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Sequence (5′ -> 3′) | Reaction Mixture | Amplification Program | |
|---|---|---|---|
| F: ACGGCCNNRACTCCTAC 1 R: TTACGGNNTGGACTACHV | Water | 2.6 μl |  |
| Kappa | 5.0 μl | ||
| Primer F (10 μM) | 0.2 μl | ||
| Primer R (10 μM) | 0.2 μl | ||
| DNA | 2.0 μl | ||
| F: CCTACGGGNGGCWGCAG 2 R: GACTACHVGGGTATCTAATCC | Water | 10.5 μl |  |
| Kappa | 12.5 μl | ||
| Primer F (10 μM) | 0.5 μl | ||
| Primer F (10 μM) | 0.5 μl | ||
| DNA | 1.0 μl | ||
| Parameter | Control (n = 27) | Obese (n = 17) | T2D (n = 22) | p-Value |
|---|---|---|---|---|
| Age [years] | 42 (36.0–48.5) | 40 (26–42) | 45.5 (37.0–55.25) | p = 0.179 |
| BMI [kg/m2] | 23.2 (22.9–23.7) | 45 (42.2–5.2) | 44 (40.1–47.1) | p < 0.001 |
| HbA1c [%] | 5.2 (5.1–5.3) | 5.3 (5.2–5.5) | 6.25 (6.1–6.5) | p < 0.001 |
| Total cholesterol [mmol/l] | 5.1 (4.9–5.2) | 4.5 (3.6–5.0) | 3.9 (3.4–5.4) | p = 0.003 |
| HDL [mmol/l] | 0.98 (0.91–3.0) | 1.14 (1.13–1.23) | 1 (0.7–1.18) | p = 0.040 |
| LDL [mmol/l] | 3.16 (0.88) | 2.75 (0.65) | 2.73 (0.98) | p = 0.160 |
| TGs [mmol/l] | 0.9 (0.9–1.2) | 1.26 (0.9–1.7) | 1.6 (1.5–1.9) | p = 0.005 |
| ALT [U/l] | 20 (18.0–25.6) | 44 (28–91) | 47 (22.0–178.5) | p < 0.001 |
| Taxonomic Level | Abundance 1 | Number of Reads | Percent of Reads 2 |
|---|---|---|---|
| kingdom | 1 | 4,844,701 | 98.50% |
| phylum | 7 | 4,844,701 | 98.50% |
| class | 22 | 4,844,701 | 98.50% |
| order | 43 | 4,844,701 | 98.50% |
| family | 100 | 4,840,921 | 98.40% |
| genus | 175 | 4,720,767 | 95.96% |
| species | 149 | 1,948,899 | 39.61% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sroka-Oleksiak, A.; Młodzińska, A.; Bulanda, M.; Salamon, D.; Major, P.; Stanek, M.; Gosiewski, T. Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. J. Clin. Med. 2020, 9, 369. https://doi.org/10.3390/jcm9020369
Sroka-Oleksiak A, Młodzińska A, Bulanda M, Salamon D, Major P, Stanek M, Gosiewski T. Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. Journal of Clinical Medicine. 2020; 9(2):369. https://doi.org/10.3390/jcm9020369
Chicago/Turabian StyleSroka-Oleksiak, Agnieszka, Agata Młodzińska, Małgorzata Bulanda, Dominika Salamon, Piotr Major, Maciej Stanek, and Tomasz Gosiewski. 2020. "Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study" Journal of Clinical Medicine 9, no. 2: 369. https://doi.org/10.3390/jcm9020369
APA StyleSroka-Oleksiak, A., Młodzińska, A., Bulanda, M., Salamon, D., Major, P., Stanek, M., & Gosiewski, T. (2020). Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. Journal of Clinical Medicine, 9(2), 369. https://doi.org/10.3390/jcm9020369