Correlates of Normal and Abnormal General Movements in Infancy and Long-Term Neurodevelopment of Preterm Infants: Insights from Functional Connectivity Studies at Term Equivalence

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. General Movement Assessment
2.3. MR Imaging
2.4. Functional MRI Analysis
2.5. Assessment of Neurodevelopmental Outcomes
2.6. Analysis of Functional Connectivity
2.7. Statistical Analysis
3. Results
3.1. The Motor Optimality Score Accounts for a Greater Proportion of the Variance in Bayley III Scores than the Fidgety Movements Score Alone
3.2. Functional Connectivity Studies
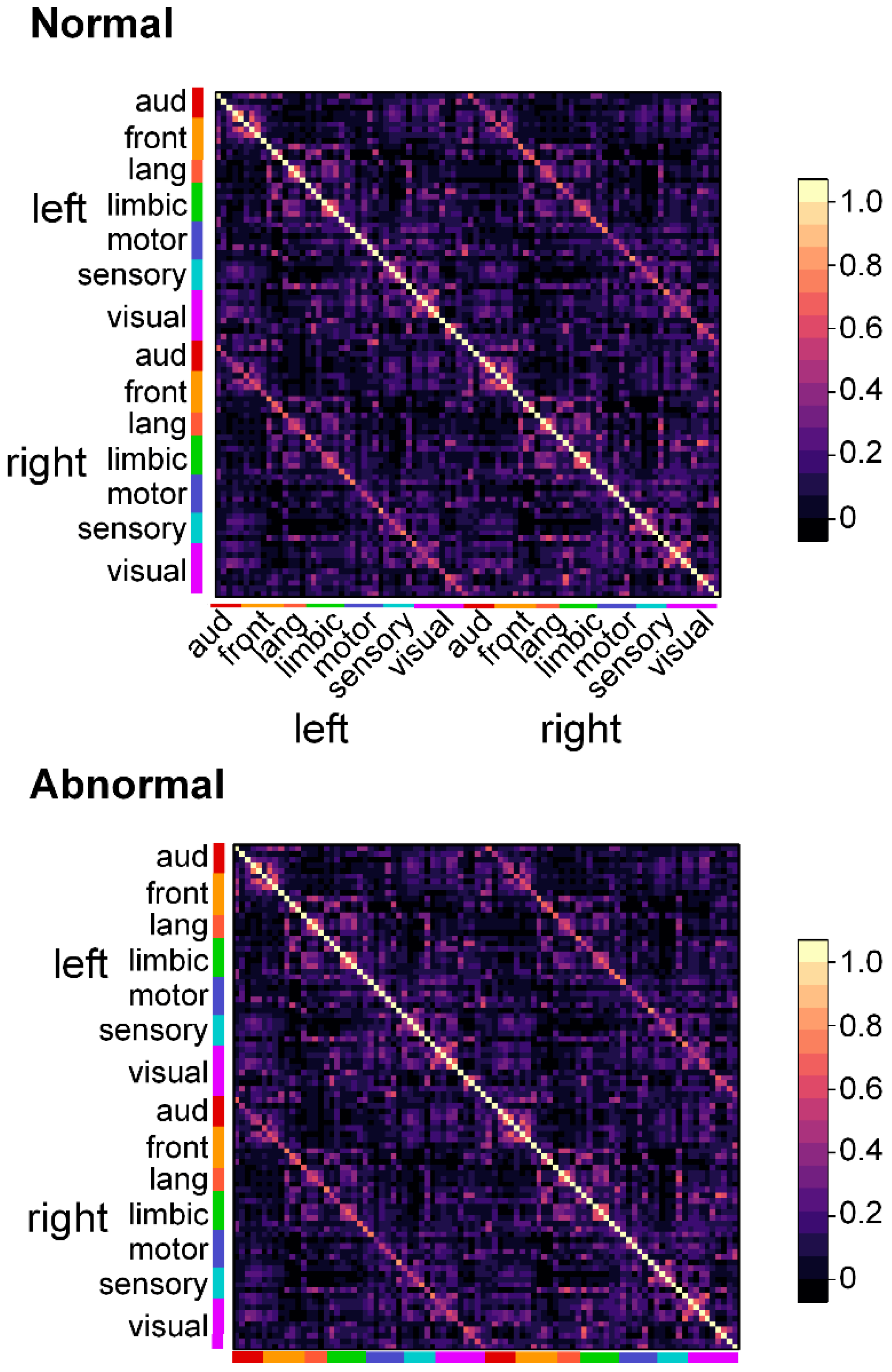
3.2.1. Infants with Normal FMs Demonstrate Greater Functional Connectivity than do Infants with Aberrant FMs
3.2.2. The Motor Optimality Score Reveals More Functional Connectivity Differences than does the Binary FM Score
3.3. Correlates of Movement Character and Functional Connectivity
3.3.1. Monotonous Movement Character is Associated with Altered Functional Connectivity between Primary Somatosensory Cortex and Visuospatial Regions
3.3.2. Stiff Movement Character is Associated with Decreased Connectivity between Visual Cortex and Cingulate Cortices
3.3.3. Jerky Movement Character is Associated with Increased Connectivity between Visual Cortex and Cingulate Cortices
3.3.4. Tremulous Movement Characters is not Associated with Differences in Regional Connectivity
3.4. Resting State Functional Connectivity Correlates of Normal and Adverse Long-Term Neurodevelopmental Outcomes
3.5. Functional Connectivity Increases as a Linear Function of Increasing Scores on Assessments of Long-Term Neurodevelopmental Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bosanquet, M.; Copeland, L.; Ware, R.; Boyd, R. A systematic review of tests to predict cerebral palsy in young children. Dev. Med. Child Neurol. 2013, 55, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, F.; Cioni, G.; Einspieler, C.; Roversi, M.F.; Bos, A.F.; Paolicelli, P.B.; Ranzi, A.; Prechtl, H.F. Cramped synchronized general movements in preterm infants as an early marker for cerebral palsy. Arch. Pediatr. Adolesc. Med. 2002, 156, 460–467. [Google Scholar] [CrossRef]
- Einspieler, C.; Prechtl, H.F.; Bos, A.F.; Ferrari, F.; Cioni, G. Prechtl’s Method on the Qualitative Assessment of General Movements in Preterm, Term and Young Infants; Cambridge University Press: London, UK, 2008. [Google Scholar]
- Prechtl, H.F.; Einspieler, C.; Cioni, G.; Bos, A.F.; Ferrari, F.; Sontheimer, D. An early marker for neurological deficits after perinatal brain lesions. Lancet 1997, 349, 1361–1363. [Google Scholar] [CrossRef]
- Peyton, C.; Yang, E.; Msall, M.E.; Adde, L.; Stoen, R.; Fjortoft, T.; Bos, A.F.; Einspieler, C.; Zhou, Y.; Schreiber, M.D.; et al. White Matter Injury and General Movements in High-Risk Preterm Infants. AJNR Am. J. Neuroradiol. 2017, 38, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyser, C.D.; Wheelock, M.D.; Limbrick, D.D., Jr.; Neil, J.J. Neonatal brain injury and aberrant connectivity. Neuroimage 2019, 185, 609–623. [Google Scholar] [CrossRef]
- Salavati, S.; Einspieler, C.; Vagelli, G.; Zhang, D.; Pansy, J.; Burgerhof, J.G.M.; Marschik, P.B.; Bos, A.F. The association between the early motor repertoire and language development in term children born after normal pregnancy. Early Hum. Dev. 2017, 111, 30–35. [Google Scholar] [CrossRef]
- Bruggink, J.L.; Einspieler, C.; Butcher, P.R.; Van Braeckel, K.N.; Prechtl, H.F.; Bos, A.F. The quality of the early motor repertoire in preterm infants predicts minor neurologic dysfunction at school age. J. Pediatr. 2008, 153, 32–39. [Google Scholar] [CrossRef]
- Bruggink, J.L.; Van Braeckel, K.N.; Bos, A.F. The early motor repertoire of children born preterm is associated with intelligence at school age. Pediatrics 2010, 125, e1356–e1363. [Google Scholar] [CrossRef] [Green Version]
- Bruggink, J.L.; Einspieler, C.; Butcher, P.R.; Stremmelaar, E.F.; Prechtl, H.F.; Bos, A.F. Quantitative aspects of the early motor repertoire in preterm infants: Do they predict minor neurological dysfunction at school age? Early Hum. Dev. 2009, 85, 25–36. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [Green Version]
- Doria, V.; Beckmann, C.F.; Arichi, T.; Merchant, N.; Groppo, M.; Turkheimer, F.E.; Counsell, S.J.; Murgasova, M.; Aljabar, P.; Nunes, R.G.; et al. Emergence of resting state networks in the preterm human brain. Proc. Natl. Acad. Sci. USA 2010, 107, 20015–20020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyser, C.D.; Inder, T.E.; Shimony, J.S.; Hill, J.E.; Degnan, A.J.; Snyder, A.Z.; Neil, J.J. Longitudinal analysis of neural network development in preterm infants. Cereb. Cortex 2010, 20, 2852–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyser, C.D.; Neil, J.J. Use of resting-state functional MRI to study brain development and injury in neonates. Semin Perinatol. 2015, 39, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyton, C.; Schreiber, M.D.; Msall, M.E. The Test of Infant Motor Performance at 3 months predicts language, cognitive, and motor outcomes in infants born preterm at 2 years of age. Dev. Med. Child Neurol. 2018, 60, 1239–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fjortoft, T.; Einspieler, C.; Adde, L.; Strand, L.I. Inter-observer reliability of the “Assessment of Motor Repertoire—3 to 5 Months” based on video recordings of infants. Early Hum. Dev. 2009, 85, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golan, A.; Marco, R.; Raz, H.; Shany, E. Imaging in the newborn: Infant immobilizer obviates the need for anesthesia. Isr. Med. Assoc. J. 2011, 13, 663–665. [Google Scholar]
- Shi, F.; Yap, P.T.; Wu, G.; Jia, H.; Gilmore, J.H.; Lin, W.; Shen, D. Infant brain atlases from neonates to 1- and 2-year-olds. PLoS ONE 2011, 6, e18746. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Bayley, N. Bayley Scales of Infant and Toddler Development, 3rd ed.; Harcourt Assessment, Inc.: San Antonio, TX, USA, 2006. [Google Scholar]
- Anderson, P.J.; De Luca, C.R.; Hutchinson, E.; Roberts, G.; Doyle, L.W.; Victorian Infant Collaborative, G. Underestimation of developmental delay by the new Bayley-III Scale. Arch. Pediatr. Adolesc. Med. 2010, 164, 352–356. [Google Scholar] [CrossRef] [Green Version]
- Vohr, B.R.; Stephens, B.E.; Higgins, R.D.; Bann, C.M.; Hintz, S.R.; Das, A.; Newman, J.E.; Peralta-Carcelen, M.; Yolton, K.; Dusick, A.M.; et al. Are outcomes of extremely preterm infants improving? Impact of Bayley assessment on outcomes. J. Pediatr. 2012, 161, 222–228.e3. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.; Johnson, S.; Haider, S.; Hennessy, E.; Marlow, N. Relationship between test scores using the second and third editions of the Bayley Scales in extremely preterm children. J. Pediatr. 2012, 160, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Smyser, C.D.; Snyder, A.Z.; Shimony, J.S.; Mitra, A.; Inder, T.E.; Neil, J.J. Resting-State Network Complexity and Magnitude Are Reduced in Prematurely Born Infants. Cereb. Cortex 2016, 26, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Ball, G.; Aljabar, P.; Arichi, T.; Tusor, N.; Cox, D.; Merchant, N.; Nongena, P.; Hajnal, J.V.; Edwards, A.D.; Counsell, S.J. Machine-learning to characterise neonatal functional connectivity in the preterm brain. Neuroimage 2016, 124, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Smyser, C.D.; Dosenbach, N.U.; Smyser, T.A.; Snyder, A.Z.; Rogers, C.E.; Inder, T.E.; Schlaggar, B.L.; Neil, J.J. Prediction of brain maturity in infants using machine-learning algorithms. Neuroimage 2016, 136, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Genovese, C.R.; Lazar, N.A.; Nichols, T. Thresholding of statistical maps in functional neuroimaging using the false discovery rate. Neuroimage 2002, 15, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.H.; Knowlton, B.J. The role of the basal ganglia in habit formation. Nat. Rev. Neurosci. 2006, 7, 464–476. [Google Scholar] [CrossRef]
- Prechtl, H.F.; Hopkins, B. Developmental transformations of spontaneous movements in early infancy. Early Hum. Dev. 1986, 14, 233–238. [Google Scholar] [CrossRef]
- Pugh, K.R.; Mencl, W.E.; Shaywitz, B.A.; Shaywitz, S.E.; Fulbright, R.K.; Constable, R.T.; Skudlarski, P.; Marchione, K.E.; Jenner, A.R.; Fletcher, J.M.; et al. The angular gyrus in developmental dyslexia: Task-specific differences in functional connectivity within posterior cortex. Psychol. Sci. 2000, 11, 51–56. [Google Scholar] [CrossRef]
- Miglioretti, D.L.; Boatman, D. Modeling variability in cortical representations of human complex sound perception. Exp. Brain Res. 2003, 153, 382–387. [Google Scholar] [CrossRef]
- Ball, G.; Pazderova, L.; Chew, A.; Tusor, N.; Merchant, N.; Arichi, T.; Allsop, J.M.; Cowan, F.M.; Edwards, A.D.; Counsell, S.J. Thalamocortical Connectivity Predicts Cognition in Children Born Preterm. Cereb. Cortex 2015, 25, 4310–4318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheelock, M.D.; Austin, N.C.; Bora, S.; Eggebrecht, A.T.; Melzer, T.R.; Woodward, L.J.; Smyser, C.D. Altered functional network connectivity relates to motor development in children born very preterm. Neuroimage 2018, 183, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Salzwedel, A.P.; Stephens, R.L.; Goldman, B.D.; Lin, W.; Gilmore, J.H.; Gao, W. Development of Amygdala Functional Connectivity During Infancy and Its Relationship With 4-Year Behavioral Outcomes. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2019, 4, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Hickok, G.; Poeppel, D. The cortical organization of speech processing. Nat. Rev. Neurosci. 2007, 8, 393. [Google Scholar] [CrossRef]
- Hertrich, I.; Dietrich, S.; Ackermann, H. The role of the supplementary motor area for speech and language processing. Neurosci. Biobehav. Rev. 2016, 68, 602–610. [Google Scholar] [CrossRef]
- Novak, I.; Morgan, C.; Adde, L.; Blackman, J.; Boyd, R.N.; Brunstrom-Hernandez, J.; Cioni, G.; Damiano, D.; Darrah, J.; Eliasson, A.C.; et al. Early, Accurate Diagnosis and Early Intervention in Cerebral Palsy: Advances in Diagnosis and Treatment. JAMA Pediatr. 2017, 171, 897–907. [Google Scholar] [CrossRef]
- Van Marter, L.J.; Kuban, K.C.; Allred, E.; Bose, C.; Dammann, O.; O’Shea, M.; Laughon, M.; Ehrenkranz, R.A.; Schreiber, M.D.; Karna, P.; et al. Does bronchopulmonary dysplasia contribute to the occurrence of cerebral palsy among infants born before 28 weeks of gestation? Arch. Dis. Child. Fetal Neonatal Ed. 2011, 96, F20–F29. [Google Scholar] [CrossRef]
- Natarajan, G.; Pappas, A.; Shankaran, S.; Kendrick, D.E.; Das, A.; Higgins, R.D.; Laptook, A.R.; Bell, E.F.; Stoll, B.J.; Newman, N.; et al. Outcomes of extremely low birth weight infants with bronchopulmonary dysplasia: Impact of the physiologic definition. Early Hum. Dev. 2012, 88, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.A.; Singer, L.T.; Fulton, S.; Salvator, A.; Short, E.J.; Klein, N.; Baley, J. Speech and language outcomes of children with bronchopulmonary dysplasia. J. Commun. Disord. 2002, 35, 393–406. [Google Scholar] [CrossRef]
- Potharst, E.S.; van Wassenaer-Leemhuis, A.G.; Houtzager, B.A.; Livesey, D.; Kok, J.H.; Last, B.F.; Oosterlaan, J. Perinatal risk factors for neurocognitive impairments in preschool children born very preterm. Dev. Med. Child Neurol. 2013, 55, 178–184. [Google Scholar] [CrossRef]
- Gough, A.; Linden, M.A.; Spence, D.; Halliday, H.L.; Patterson, C.C.; McGarvey, L. Executive functioning deficits in young adult survivors of bronchopulmonary dysplasia. Disabil. Rehabil. 2015, 37, 1940–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriram, S.; Schreiber, M.D.; Msall, M.E.; Kuban, K.C.; Joseph, R.M.; O’Shea, T.M.; Allred, E.N.; Leviton, A.; Investigators, E.S. Cognitive Development and Quality of Life Associated With BPD in 10-Year-Olds Born Preterm. Pediatrics 2018, 141, e20172719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, J.L.Y.; Doyle, L.W. An update on pulmonary and neurodevelopmental outcomes of bronchopulmonary dysplasia. Semin. Perinatol. 2018, 42, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Fjortoft, T.; Grunewaldt, K.H.; Lohaugen, G.C.; Morkved, S.; Skranes, J.; Evensen, K.A. Assessment of motor behaviour in high-risk-infants at 3 months predicts motor and cognitive outcomes in 10 years old children. Early Hum. Dev. 2013, 89, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Butcher, P.R.; van Braeckel, K.; Bouma, A.; Einspieler, C.; Stremmelaar, E.F.; Bos, A.F. The quality of preterm infants’ spontaneous movements: An early indicator of intelligence and behaviour at school age. J. Child Psychol. Psychiatry 2009, 50, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Leech, R.; Sharp, D.J. The role of the posterior cingulate cortex in cognition and disease. Brain 2014, 137, 12–32. [Google Scholar] [CrossRef] [Green Version]
- Grunewaldt, K.H.; Fjortoft, T.; Bjuland, K.J.; Brubakk, A.M.; Eikenes, L.; Haberg, A.K.; Lohaugen, G.C.; Skranes, J. Follow-up at age 10 years in ELBW children—Functional outcome, brain morphology and results from motor assessments in infancy. Early Hum. Dev. 2014, 90, 571–578. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | N = 123 |
|---|---|
| Male (n (%)) | 68 (55.3) |
| Gestational Age (weeks, mean ± SD) | 27.1 ± 2.0 |
| Gestational Age < 28 weeks (n (%)) | 67 (54.9) |
| Birth Weight (grams, mean ± SD)) | 973 ± 269 |
| Birth weight < 1000 g (n (%)) | 63 (51.6) |
| Oxygen support >36 weeks (n (%)) | 42 (34.7) |
| Necrotizing enterocolitis * (n (%)) | 10 (8.3) |
| Treated retinopathy of prematurity (n (%)) | 14 (11.6) |
| Severe intraventricular hemorrhage † (IVH; n (%)) | 9 (7.4) |
| Periventricular leukomalacia (PVL; n (%)) | 6 (5.0) |
| Severe IVH or PVL (n (%)) | 12 (9.8) |
| Normal FMs rating (n (%)) | 101 (82.8) |
| Motor Optimality Score (mean ± SD) | 21.3 ± 6.3 |
| Cerebral Palsy (n (%)) | 10 (8.3) |
| Bayley III Cognitive (mean ± SD) | 90.8 ± 17.3 |
| Bayley III Language (mean ± SD) | 88.4 ± 14.5 |
| Bayley III Motor (mean ± SD) | 92.5 ± 15.5 |
| Cognitive | Language | Motor | ||||
|---|---|---|---|---|---|---|
| Model 1 (FM only) | Model 2 (FM + MR) | Model 1 (FM only) | Model 2 (FM + MR) | Model 1 (FM only) | Model 2 (FM + MR) | |
| FM coefficient | 1.43 (0.63–2.24) p < 0.001 | 0.86 (0.04–1.68) p = 0.04 | 1.21 (0.55–1.87) p < 0.001 | 0.83 (0.14–1.51) p = 0.02 | 1.33 (0.63–2.04) p < 0.001 | 0.79 (0.07–1.50) p = 0.03 |
| MR coefficient | - | 2.04 (0.95–3.12) p < 0.001 | - | 1.37 (0.46–2.29) p = 0.004 | - | 1.96 (1.01–2.91) p < 0.001 |
| R-squared | 0.11 | 0.20 | 0.11 | 0.16 | 0.12 | 0.23 |
| Scanned (N = 47) | Not Scanned (N = 70) | P (Unadjusted) | |
|---|---|---|---|
| Male (N, %) | 27 (57.4) | 41 (54.0) | 0.71 |
| Mean Gestational age (weeks, ± SD) | 27.2 ± 0.3 | 27.0 ± 0.2 | 0.50 |
| Mean Birth Weight (g ± SD) | 994 ± 28 | 961 ± 32 | 0.63 |
| Bronchopulmonary dysplasia * (N, %) | 11 (24.4) | 31 (40.8) | 0.08 |
| Treated retinopathy of prematurity (N, %) | 3 (6.7) | 11 (14.5) | 0.25 |
| Severe IVH or PVL (N, %) | 4 (9.5) | 8 (10.5) | 1.00 |
| Normal FMs (N = 41) | Aberrant FMs (N = 12) | P | |
|---|---|---|---|
| Male | 26 (63.4) | 4 (33.3) | 0.10 |
| Mean Gestational age (weeks, ± SD) | 27.6 ± 1.6 | 26.6 ± 2.0 | 0.07 |
| Mean Birth Weight (g, ± SD) | 1050 ± 222 | 756 ± 217 | 0.001 |
| Mean Post-term age at GMA testing (weeks, ± SD) | 12.0 ± 1.3 | 11.92 ± 1.5 | 0.85 |
| Bronchopulmonary dysplasia * (N, %) | 12 (28.95) | 2 (66.7) | 0.48 |
| Treated retinopathy of prematurity (N, %) | 3 (5.26) | 2 (33.3) | 0.31 |
| Severe IVH or PVL (N, %) | 2 (4.9) | 3 (25) | 0.07 |
| Mean Post-conceptional age at MRI (weeks, ± SD) | 38.99 ± 2.52 | 37.61 ± 3.04 | 0.16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peyton, C.; Einspieler, C.; Fjørtoft, T.; Adde, L.; Schreiber, M.D.; Drobyshevsky, A.; Marks, J.D. Correlates of Normal and Abnormal General Movements in Infancy and Long-Term Neurodevelopment of Preterm Infants: Insights from Functional Connectivity Studies at Term Equivalence. J. Clin. Med. 2020, 9, 834. https://doi.org/10.3390/jcm9030834
Peyton C, Einspieler C, Fjørtoft T, Adde L, Schreiber MD, Drobyshevsky A, Marks JD. Correlates of Normal and Abnormal General Movements in Infancy and Long-Term Neurodevelopment of Preterm Infants: Insights from Functional Connectivity Studies at Term Equivalence. Journal of Clinical Medicine. 2020; 9(3):834. https://doi.org/10.3390/jcm9030834
Chicago/Turabian StylePeyton, Colleen, Christa Einspieler, Toril Fjørtoft, Lars Adde, Michael D. Schreiber, Alexander Drobyshevsky, and Jeremy D. Marks. 2020. "Correlates of Normal and Abnormal General Movements in Infancy and Long-Term Neurodevelopment of Preterm Infants: Insights from Functional Connectivity Studies at Term Equivalence" Journal of Clinical Medicine 9, no. 3: 834. https://doi.org/10.3390/jcm9030834
APA StylePeyton, C., Einspieler, C., Fjørtoft, T., Adde, L., Schreiber, M. D., Drobyshevsky, A., & Marks, J. D. (2020). Correlates of Normal and Abnormal General Movements in Infancy and Long-Term Neurodevelopment of Preterm Infants: Insights from Functional Connectivity Studies at Term Equivalence. Journal of Clinical Medicine, 9(3), 834. https://doi.org/10.3390/jcm9030834