Protein Profiling of Serum Extracellular Vesicles Reveals Qualitative and Quantitative Differences after Differential Ultracentrifugation and ExoQuick™ Isolation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of EVs
2.2. EVs Protein Enrichment and Purification
2.3. Two-Dimensional Multiplex Fluorescence Gel Electrophoresis (2D-DIGE)
2.4. Multiplex Fluorescence-Based Western Blot Analysis
3. Results
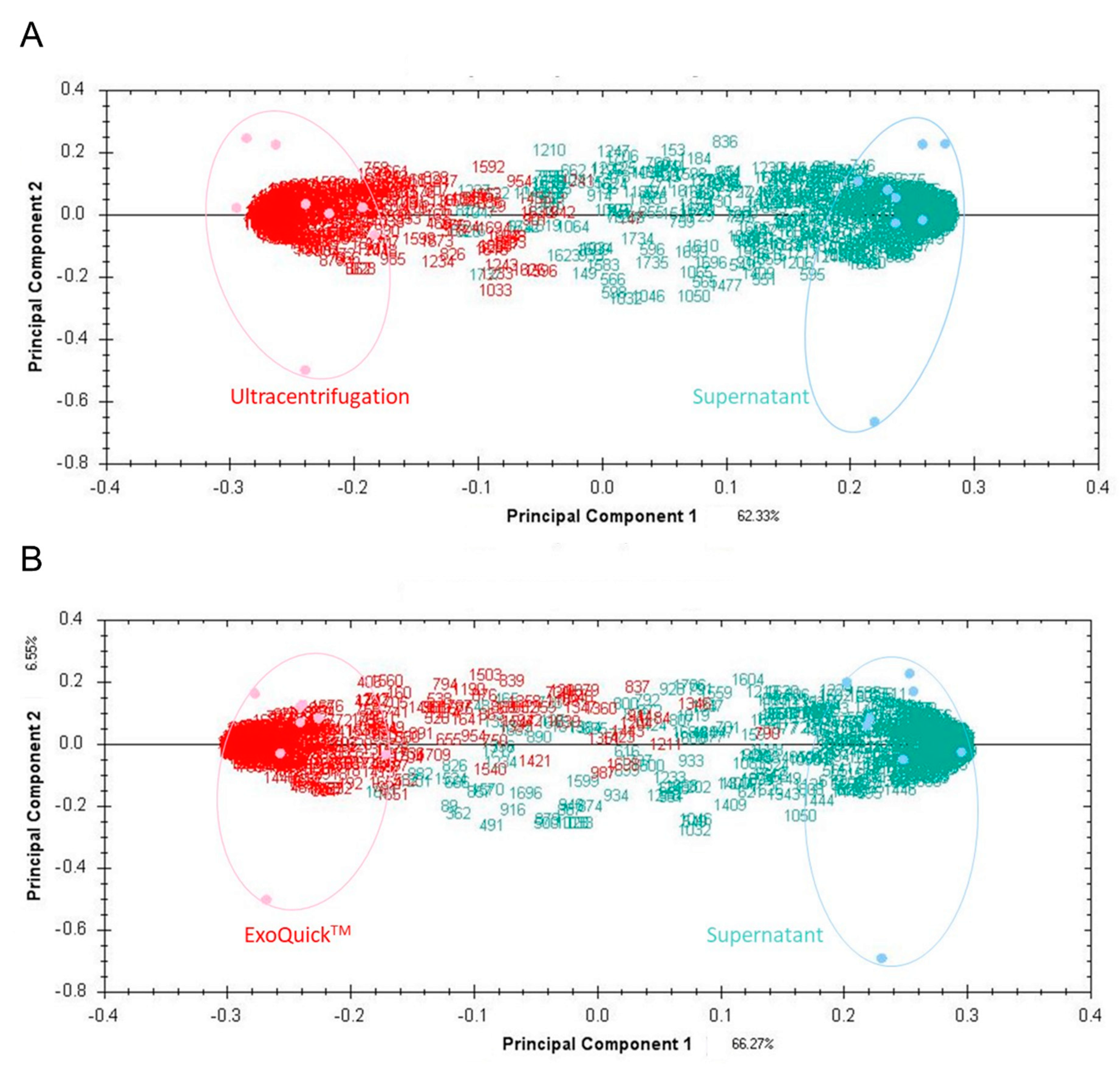
3.1. EV Protein Profile Depending on Isolation Method
3.2. Mass Spectrometry & Pathway Analyses
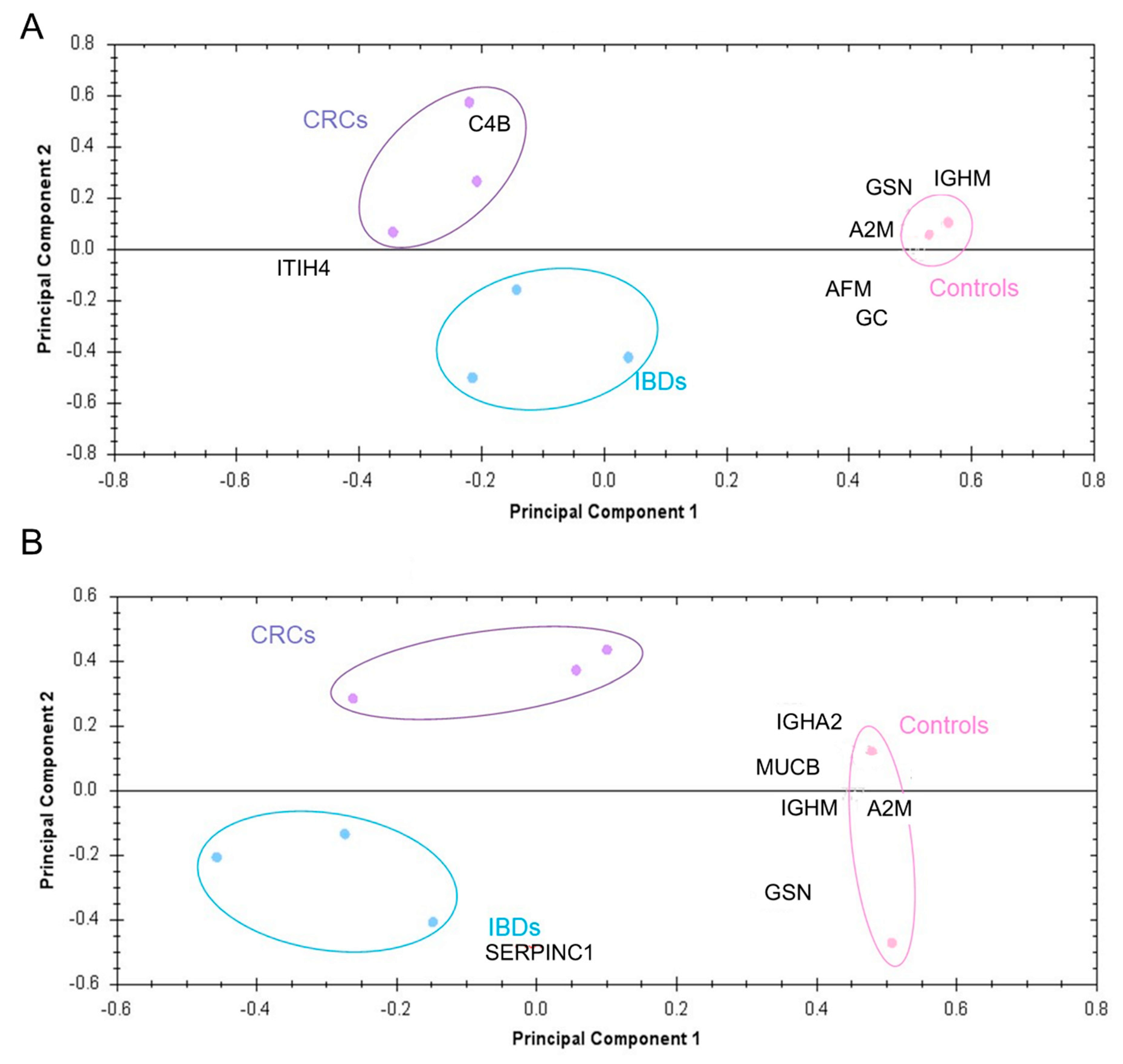
3.3. Potential Diagnostic Power of EV Proteins
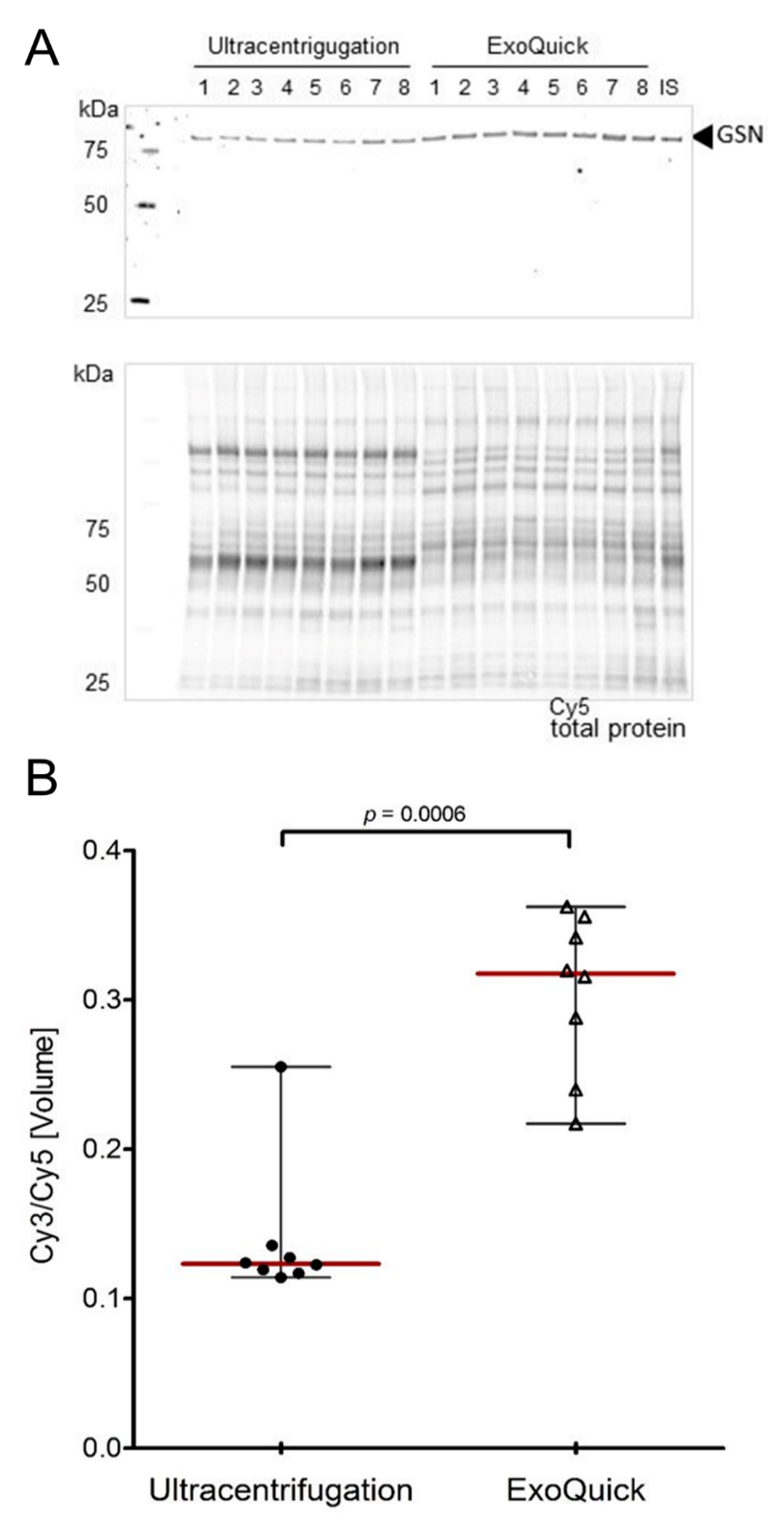
3.4. Validation of Target Proteins by Fluorescence-Based Western Blot
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability Statement
Novelty and Impact Statements
Abbreviations
| ICB-L | Interdisciplinary Center for Biobanking-Lübeck |
| EVs | Extracellular vesicles |
| IBD | Inflammatory bowel disease |
| CRC | Colorectal cancer |
| 2D DIGE | Multiplex-fluorescence two-dimensional difference gel electrophoresis |
| MALDI-TOF MS | Matrix-assisted laser desorption/ionization time of flight mass spectrometry |
| GSN | Gelsolin |
| PCA | Principal component analyses |
| UICC | Union Internationale Contre le Cancer |
References
- Xu, P.; Zhu, Y.; Sun, B.; Xiao, Z. Colorectal cancer characterization and therapeutic target prediction based on microRNA expression profile. Sci. Rep. 2016, 6, 20616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odze, R.D.; Goldblum, J.; Noffsinger, A.; Alsaigh, N.; Rybicki, L.A.; Fogt, F. Interobserver Variability in the Diagnosis of Ulcerative Colitis-Associated Dysplasia by Telepathology. Mod. Pathol. 2002, 15, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Althumairi, A.; Lazarev, M.G.; Gearhart, S.L. Inflammatory bowel disease associated neoplasia: A surgeon’s perspective. World J. Gastroenterol. 2016, 22, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xia, W.; Lv, Z.; Ni, C.; Xin, Y.; Yang, L. Liquid Biopsy for Cancer: Circulating Tumor Cells, Circulating Free DNA or Exosomes? Cellular physiology and biochemistry. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 755–768. [Google Scholar] [CrossRef]
- Whiteside, T.L. Tumor-Derived Exosomes and Their Role in Cancer Progression. Adv. Clin. Chem. 2016, 74, 103–141. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, S.; Palanisamy, V. Horizontal transfer of RNAs: Exosomes as mediators of intercellular communication. Wiley Interdiscip. Rev. RNA 2012, 3, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.J.; Kim, O.Y.; Gho, Y.S. Extracellular vesicles as emerging intercellular communicasomes. BMB Rep. 2014, 47, 531–539. [Google Scholar] [CrossRef]
- Xiao, H.; Lässer, C.; Shelke, G.V.; Wang, J.; Rådinger, M.; Lunavat, T.R.; Malmhäll, C.; Lin, L.H.; Li, J.; Li, L.; et al. Mast cell exosomes promote lung adenocarcinoma cell proliferation – role of KIT-stem cell factor signaling. Cell Commun. Signal. 2014, 12, 64. [Google Scholar]
- Plebanek, M.P.; Angeloni, N.; Vinokour, E.; Li, J.; Henkin, A.; Martinez-Marin, D.; Filleur, S.; Bhowmick, R.; Henkin, J.; Miller, S.D.; et al. Pre-metastatic cancer exosomes induce immune surveillance by patrolling monocytes at the metastatic niche. Nat. Commun. 2017, 8, 1319. [Google Scholar] [CrossRef]
- Wahlund, C.; Güclüler, G.; Hiltbrunner, S.; Veerman, R.E.; Näslund, T.I.; Gabrielsson, S. Exosomes from antigen-pulsed dendritic cells induce stronger antigen-specific immune responses than microvesicles in vivo. Sci. Rep. 2017, 7, 17095. [Google Scholar] [CrossRef] [Green Version]
- Greening, D.; Gopal, S.K.; Mathias, R.; Liu, L.; Sheng, J.; Zhu, H.-J.; Simpson, R.; Sheng, J. Emerging roles of exosomes during epithelial–mesenchymal transition and cancer progression. Semin. Cell Dev. Boil. 2015, 40, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.T.; Lu, J.T.; Ran, Z.H.; Zheng, Q. Exosome in intestinal mucosal immunity. J. Gastroenterol. Hepatol. 2016, 31, 1694–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Li, C.; Zhou, T.; Liu, X.; Liu, X.; Li, X.; Chen, D. Role of exosomal proteins in cancer diagnosis. Mol. Cancer 2017, 16, 145. [Google Scholar] [CrossRef] [PubMed]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, E.F.; Ham, A.-J.L.; Tabb, D.L.; Billheimer, D.; Roth, B.J.; Chang, S.S.; Cookson, M.S.; Hinton, T.J.; Cheek, K.L.; Hill, S.; et al. Prostate cancer serum biomarker discovery through proteomic analysis of alpha-2 macroglobulin protein complexes. Proteom. Clin. Appl. 2008, 2, 1223–1233. [Google Scholar] [CrossRef] [Green Version]
- Dieplinger, H.; Ankerst, D.; Burges, A.; Lenhard, M.; Lingenhel, A.; Fineder, L.; Buchner, H.; Stieber, P. Afamin and Apolipoprotein A-IV: Novel Protein Markers for Ovarian Cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1127–1133. [Google Scholar] [CrossRef] [Green Version]
- Sogawa, K.; Takano, S.; Iida, F.; Satoh, M.; Tsuchida, S.; Kawashima, Y.; Yoshitomi, H.; Sanda, A.; Kodera, Y.; Takizawa, H.; et al. Identification of a novel serum biomarker for pancreatic cancer, C4b-binding protein α-chain (C4BPA) by quantitative proteomic analysis using tandem mass tags. Br. J. Cancer 2016, 115, 949–956. [Google Scholar] [CrossRef]
- Szatanek, R.; Baran, J.; Siedlar, M.; Baj-Krzyworzeka, M. Isolation of extracellular vesicles: Determining the correct approach (Review). Int. J. Mol. Med. 2015, 36, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Helwa, I.; Cai, J.; Drewry, M.D.; Zimmerman, A.; Dinkins, M.B.; Khaled, M.L.; Seremwe, M.; Dismuke, W.M.; Bieberich, E.; Stamer, W.D.; et al. A Comparative Study of Serum Exosome Isolation Using Differential Ultracentrifugation and Three Commercial Reagents. PLoS ONE 2017, 12, e0170628. [Google Scholar] [CrossRef]
- Sáenz-Cuesta, M.; Arbelaiz, A.; Oregi, A.; Irizar, H.; Osorio-Querejeta, I.; Muñoz-Culla, M.; Bañales, J.; Falcón-Pérez, J.M.; Olascoaga, J.; Otaegui, D. Methods for Extracellular Vesicles Isolation in a Hospital Setting. Front. Immunol. 2015, 6, 50. [Google Scholar] [CrossRef]
- Gemoll, T.; Kollbeck, S.L.; Karstens, K.F.; Hò, G.G.; Hartwig, S.; Strohkamp, S.; Schillo, K.; Thorns, C.; Oberländer, M.; Kalies, K.; et al. EB1 protein alteration characterizes sporadic but not ulcerative colitis associated colorectal cancer. Oncotarget 2017, 8, 54939–54950. [Google Scholar] [CrossRef] [PubMed]
- Hagner-McWhirter, A.; Laurin, Y.; Larsson, A.; Bjerneld, E.J.; Rönn, O. Cy5 total protein normalization in Western blot analysis. Anal. Biochem. 2015, 486, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-T.; Huang, Y.-Y.; Zheng, L.; Qin, S.-H.; Xu, X.-P.; An, T.-X.; Xu, Y.; Wu, Y.-S.; Hu, X.-M.; Ping, B.-H.; et al. Comparison of isolation methods of exosomes and exosomal RNA from cell culture medium and serum. Int. J. Mol. Med. 2017, 40, 834–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webber, J.; Clayton, A. How pure are your vesicles? J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M.; Livshts, M.A. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef] [PubMed]
- Linares, R.; Tan, S.; Gounou, C.; Arraud, N.; Brisson, A.R. High-speed centrifugation induces aggregation of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29509. [Google Scholar] [CrossRef]
- Rosa-Fernandes, L.; Rocha, V.; Carregari, V.C.; Urbani, A.; Palmisano, G. A Perspective on Extracellular Vesicles Proteomics. Front. Chem. 2017, 5, 102. [Google Scholar] [CrossRef] [Green Version]
- Lobb, R.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Moller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Zarovni, N.; Corrado, A.; Guazzi, P.; Zocco, D.; Lari, E.; Radano, G.; Muhhina, J.; Fondelli, C.; Gavrilova, J.; Chiesi, A. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches. Methods 2015, 87, 46–58. [Google Scholar] [CrossRef]
- Kurz, S.; Thieme, R.; Amberg, R.; Groth, M.; Jahnke, H.-G.; Pieroh, P.; Horn, L.-C.; Kolb, M.; Huse, K.; Platzer, M.; et al. The anti-tumorigenic activity of A2M—A lesson from the naked mole-rat. PLoS ONE 2017, 12, e0189514. [Google Scholar] [CrossRef]
- Blom, A.M.; Villoutreix, B.O.; Dahlbäck, B. Functions of human complement C4b-binding protein in realtion to its structure. Arch. Immunol. Ther. Exp. 2004, 52, 83–95. [Google Scholar]
- Hsu, H.-M.; Chu, C.-M.; Chang, Y.-J.; Yu, J.-C.; Chen, C.-T.; Jian, C.-E.; Lee, C.-Y.; Chiang, Y.-T.; Chang, C.-W.; Chang, Y.-T. Six novel immunoglobulin genes as biomarkers for better prognosis in triple-negative breast cancer by gene co-expression network analysis. Sci. Rep. 2019, 9, 4484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noske, A.; Denkert, C.; Schober, H.; Sers, C.; Zhumabayeva, B.; Weichert, W.; Dietel, M.; Wiechen, K. Loss of Gelsolin expression in human ovarian carcinomas. Eur. J. Cancer 2005, 41, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.-M.; Klee, F.; Edlund, K.; Grinberg, M.; Hammad, S.; Marchan, R.; Cadenas, C.; Niggemann, B.; Zänker, K.S.; Rahnenführer, J.; et al. Gelsolin Is Associated with Longer Metastasis-free Survival and Reduced Cell Migration in Estrogen Receptor-positive Breast Cancer. Anticancer. Res. 2015, 35, 5277–5285. [Google Scholar]
- Fan, N.-J.; Gao, C.-F.; Wang, C.-S.; Lv, J.-J.; Zhao, G.; Sheng, X.-H.; Li, D.-H.; Liu, Q.-Y.; Yin, J. Discovery and Verification of Gelsolin as a Potential Biomarker of Colorectal Adenocarcinoma in a Chinese Population: Examining Differential Protein Expression using an Itraq Labelling-Based Proteomics Approach. Can. J. Gastroenterol. 2012, 26, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Tucher, C.; Bode, K.; Schiller, P.; Claßen, L.; Birr, C.; Souto-Carneiro, M.; Blank, N.; Lorenz, H.-M.; Schiller, M. Extracellular Vesicle Subtypes Released From Activated or Apoptotic T-Lymphocytes Carry a Specific and Stimulus-Dependent Protein Cargo. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Abramowicz, A.; Widlak, P.; Pietrowska, M. Proteomic analysis of exosomal cargo: The challenge of high purity vesicle isolation. Mol. BioSyst. 2016, 12, 1407–1419. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ultracentrifugation (UC) | ExoQuick™ | Overlapped Protein Spots | ||||
|---|---|---|---|---|---|---|
| Differentially Expressed (p < 0.05) Spots | Identified Spots | Differentially Expressed (p < 0.05) Spots | Identified Spots | Total Number | Identified Spots | |
| Controls vs IBDs | 31 | 9 | 18 | 9 | 7 | 5 |
| Controls vs. CRC | 62 | 16 | 21 | 4 | 8 | 3 |
| IBDs vs. CRCs | 14 | 5 | 17 | 5 | 7 | 3 |
| Controls vs. IBDs vs. CRCs | 32 | 10 | 26 | 11 | 8 | 6 |
| Spot No. | Protein Identity | Gene Symbol | Accession No. | MW [kDa]a | pIa | MW [kDa]b | pIb | Mascot Score (MS/MS) | Sequence Coverage [%] | UC vs. Exo (Pellets) | Ctrl. vs. IBD (UC) | Ctrl. vs. CRC (UC) | IBD. vs. CRC (UC) | Ctrl. vs. IBD vs. CRC (UC) | Ctrl. vs. IBD (Exo) | Ctrl. vs. CRC (Exo) | IBD. vs. CRC (Exo) | Ctrl. vs. IBD vs. CRC (Exo) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Afamin | AFM | P43652 | 139 | 5.19 | 69.0 | 5.6 | 61.9 | 13.7 | ↓ | ↓ | ↓ | ||||||
| 2 | Alpha-2-macroglobulin | A2M | P01023 | 186 | 6.24 | 163.2 | 6.0 | 145.4 | 3.5 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | ||
| 3 | A2M | 186 | 6.19 | 86.2 | 24.2 | ↓ | ||||||||||||
| 4 | A2M | 186 | 6.28 | 131.0 | 22.7 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||||
| 5 | A2M | 186 | 6.32 | 89.4 | 1.9 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||||
| 6 | A2M | 186 | 6.14 | 86.4 | 1.9 | ↓ | ↓ | |||||||||||
| 7 | A2M | 185 | 6.41 | 85.6 | 19.8 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||||
| 8 | Antithrombin-III | SERPINC1 | P01008 | 118 | 5.32 | 52.6 | 6.3 | 72.4 | 3.0 | ↑ | ↓ | ↓ | ||||||
| 9 | C4b-binding protein alpha chain | C4BPA | P04003 | 124 | 6.55 | 67.0 | 7.9 | 97.0 | 34.2 | ↓ | ||||||||
| 10 | C4BPA | 71 | 5.22 | 127.0 | 35.3 | ↓ | ↓ | ↓ | ||||||||||
| 11 | CD5 antigen-like | CD5L | O43866 | 65 | 5.60 | 38.1 | 5.2 | 70.8 | 25.1 | ↑ | ||||||||
| 12 | Complement C1r subcomponent | C1R | P00736 | 65 | 5.36 | 80.1 | 5.8 | 79.9 | 3.7 | ↑ | ||||||||
| 13 | Complement C4-B | C4B | P0C0L5 | 57 | 4.29 | 192.6 | 6.9 | 72.0 | 0.7 | ↑ | ↑ | ↑ | ↑ | |||||
| 14 | C4B | 56 | 4.73 | 72.9 | 1.5 | ↓ | ↑ | |||||||||||
| 15 | Complement component C6 | C6 | P13671 | 170 | 6.46 | 104.7 | 6.4 | 210 | 37.4 | ↑ | ↓ | |||||||
| 16 | C6 | 169 | 6.53 | 138.0 | 33.4 | ↑ | ↓ | |||||||||||
| 3 | Complement factor H | CFH | P08603 | 186 | 6.19 | 139.0 | 6.2 | 56.1 | 30.8 | ↓ | ||||||||
| 17 | CFH | 186 | 6.01 | 176.0 | 40.1 | ↓ | ||||||||||||
| 18 | Gelsolin | GSN | P06396 | 141 | 6.32 | 85.6 | 5.9 | 70.2 | 2.8 | ↑ | ↓ | ↓ | ↓ | |||||
| 19 | GSN | 140 | 6.34 | 59.1 | 22.0 | ↑ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||||
| 20 | GSN | 141 | 6.23 | 92.0 | 19.6 | ↑ | ↓ | ↓ | ||||||||||
| 14 | Haptoglobin | HP | P00738 | 56 | 4.73 | 45.2 | 6.1 | 90.6 | 30.5 | ↓ | ↑ | |||||||
| 21 | Ig alpha-1 chain C region | IGHA1 | P01876 | 80 | 6.29 | 37.6 | 6.1 | 118.0 | 47.9 | ↓ | ||||||||
| 22 | IGHA1 | 107 | 5.61 | 97.0 | 40.8 | ↓ | ||||||||||||
| 23 | IGHA1 | 116 | 5.30 | 64.8 | 33.7 | ↑ | ↓ | ↓ | ||||||||||
| 24 | IGHA1 | 108 | 6.62 | 61.9 | 46.5 | ↑ | ||||||||||||
| 25 | IGHA1 | 105 | 6.02 | 37.6 | 6.1 | 184.0 | 56.9 | ↓ | ||||||||||
| 26 | IGHA1 | 105 | 6.47 | 60.6 | 18.7 | ↓ | ||||||||||||
| 27 | Ig alpha-2 chain C region | IGHA2 | P01877 | 107 | 5.74 | 36.5 | 5.7 | 244.0 | 8.5 | ↓ | ↓ | ↑ | ↓ | |||||
| 25 | IGHA2 | 105 | 6.02 | 118.0 | 40.3 | ↓ | ||||||||||||
| 28 | IGHA2 | 104 | 6.20 | 92.1 | 5.0 | ↓ | ||||||||||||
| 21 | IGHA2 | 80 | 6.29 | 61.9 | 36.2 | ↓ | ||||||||||||
| 29 | IGHA2 | 106 | 5.80 | 80.9 | 22.4 | ↓ | ↓ | |||||||||||
| 22 | IGHA2 | 107 | 5.61 | 66.2 | 31.5 | ↓ | ||||||||||||
| 23 | IGHA2 | 116 | 5.30 | 71.4 | 5.0 | ↑ | ↓ | ↓ | ||||||||||
| 8 | IGHA2 | 118 | 5.32 | 87.3 | 5.0 | ↑ | ||||||||||||
| 30 | Ig kappa chain C region | IGKC | P01834 | 24 | 6.39 | 11.6 | 5.5 | 89.8 | 32.1 | ↓ | ||||||||
| 31 | IGKC | 24 | 6.60 | 11.6 | 5.5 | 92.6 | 34.9 | ↓ | ||||||||||
| 32 | Ig mu chain C region | IGHM | P01871 | 60 | 6.26 | 49.3 | 6.4 | 57.7 | 13.3 | ↑ | ↓ | ↓ | ||||||
| 33 | IGHM | 129 | 6.31 | 105.5 | 9.7 | ↑ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||||
| 34 | Ig mu heavy chain disease protein* | MUCB* | P04220* | 67 | 6.28 | 43.0 | 5.0 | 157.4 | 3.3 | ↓ | ↓ | ↓ | ||||||
| 35 | MUCB* | 141 | 6.55 | 81.0 | 2.8 | ↓ | ↓ | ↓ | ||||||||||
| 36 | Inter-alpha-trypsin inhibitor heavy chain H4 | ITIH4 | Q14624 | 168 | 5.11 | 103.3 | 6.5 | 76.1 | 17.8 | ↓ | ↑ | ↑ | ↑ | |||||
| 37 | Keratin, type II cytoskeletal 2 | KRT2 | P35908 | 27 | 5.15 | 65.4 | 8.9 | 68.8 | 35.7 | ↓ | ||||||||
| 14 | Serum paraoxonase/arylesterase 1 | PON1 | P27169 | 56 | 4.73 | 39.7 | 5.0 | 94.9 | 4.5 | ↓ | ↑ | |||||||
| 38 | Vitamin D-binding protein | GC | P02774 | 90 | 5.29 | 52.9 | 5.3 | 203.0 | 62.2 | ↑ | ↓ | ↓ | ↓ | |||||
| 39 | Zinc finger protein 705A | ZNF705A | Q6ZN79 | 41 | 6.43 | 34.7 | 10.3 | 57.3 | 26.3 | ↓ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gemoll, T.; Strohkamp, S.; Rozanova, S.; Röder, C.; Hartwig, S.; Kalthoff, H.; Lehr, S.; ElSharawy, A.; Habermann, J.K. Protein Profiling of Serum Extracellular Vesicles Reveals Qualitative and Quantitative Differences after Differential Ultracentrifugation and ExoQuick™ Isolation. J. Clin. Med. 2020, 9, 1429. https://doi.org/10.3390/jcm9051429
Gemoll T, Strohkamp S, Rozanova S, Röder C, Hartwig S, Kalthoff H, Lehr S, ElSharawy A, Habermann JK. Protein Profiling of Serum Extracellular Vesicles Reveals Qualitative and Quantitative Differences after Differential Ultracentrifugation and ExoQuick™ Isolation. Journal of Clinical Medicine. 2020; 9(5):1429. https://doi.org/10.3390/jcm9051429
Chicago/Turabian StyleGemoll, Timo, Sarah Strohkamp, Svitlana Rozanova, Christian Röder, Sonja Hartwig, Holger Kalthoff, Stefan Lehr, Abdou ElSharawy, and Jens K. Habermann. 2020. "Protein Profiling of Serum Extracellular Vesicles Reveals Qualitative and Quantitative Differences after Differential Ultracentrifugation and ExoQuick™ Isolation" Journal of Clinical Medicine 9, no. 5: 1429. https://doi.org/10.3390/jcm9051429