Regulation of Angiogenesis Discriminates Tissue Resident MSCs from Effective and Defective Osteogenic Environments †


 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Recruitment
2.2. Tissue Collection and Processing
2.3. Colony Forming Potential
2.4. In Vitro Osteogenesis, Chondrogenesis, and Adipogenesis
2.5. Histology and Immunohistochemistry
2.6. Flow Cytometry
2.7. Quantitative Real-Time PCR (qPCR)
2.8. In Vitro Angiogenesis
2.9. Statistical Analysis
3. Results
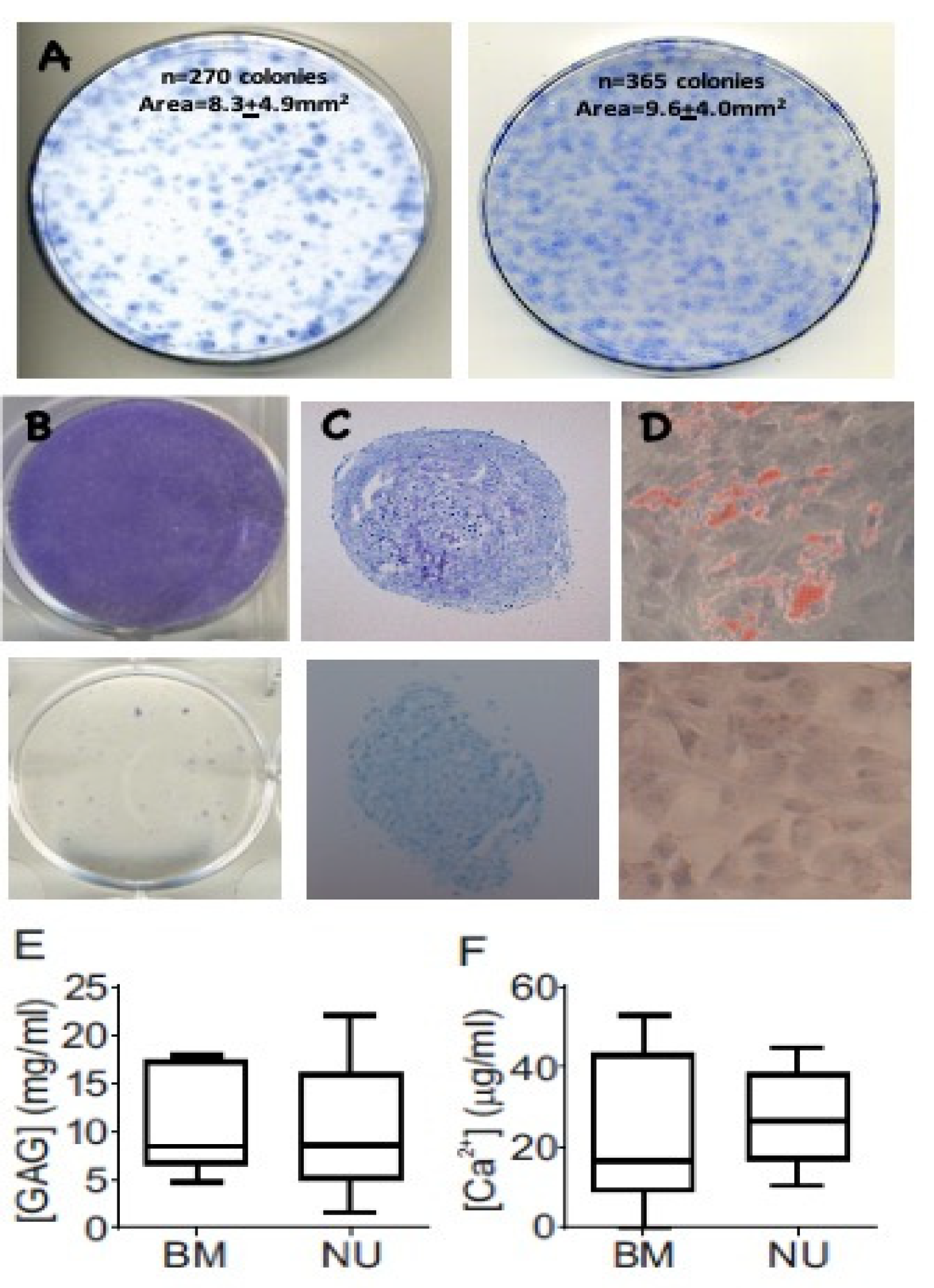
3.1. Colony Formation and Differentiation Capacity of Resident MSCs
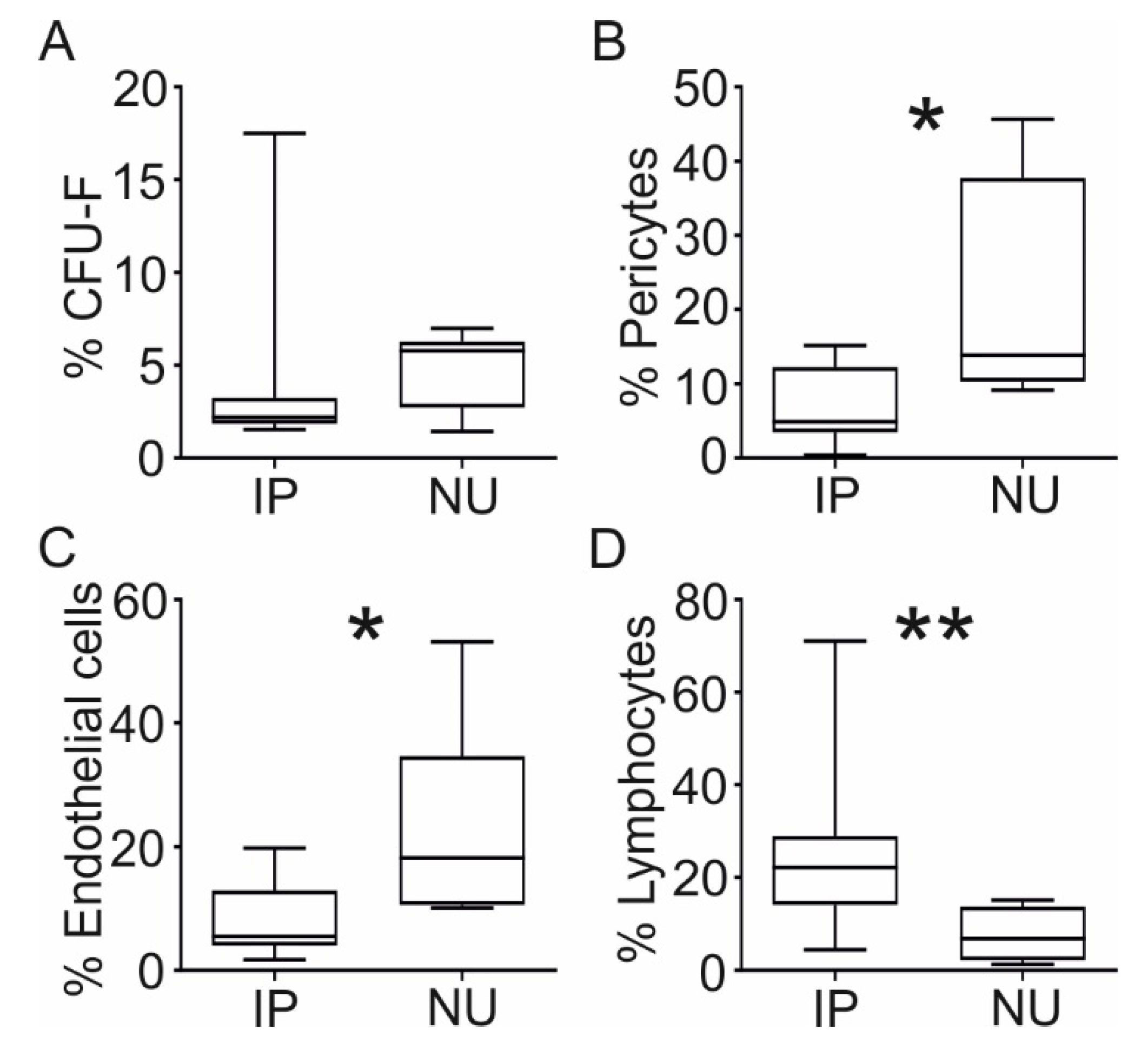
3.2. Cellular Composition of NU Tissue
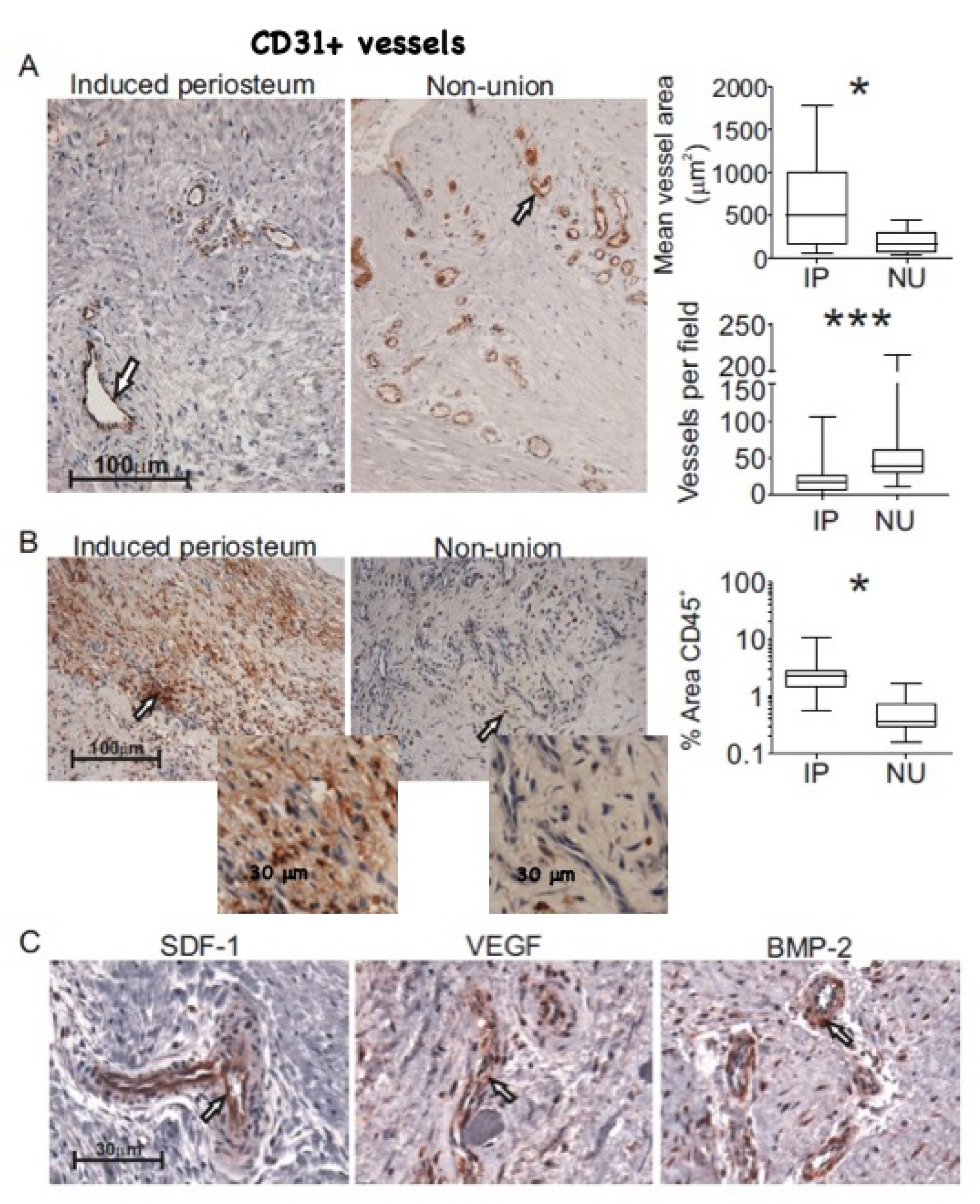
3.3. Histological Comparison of NU and IP Tissue
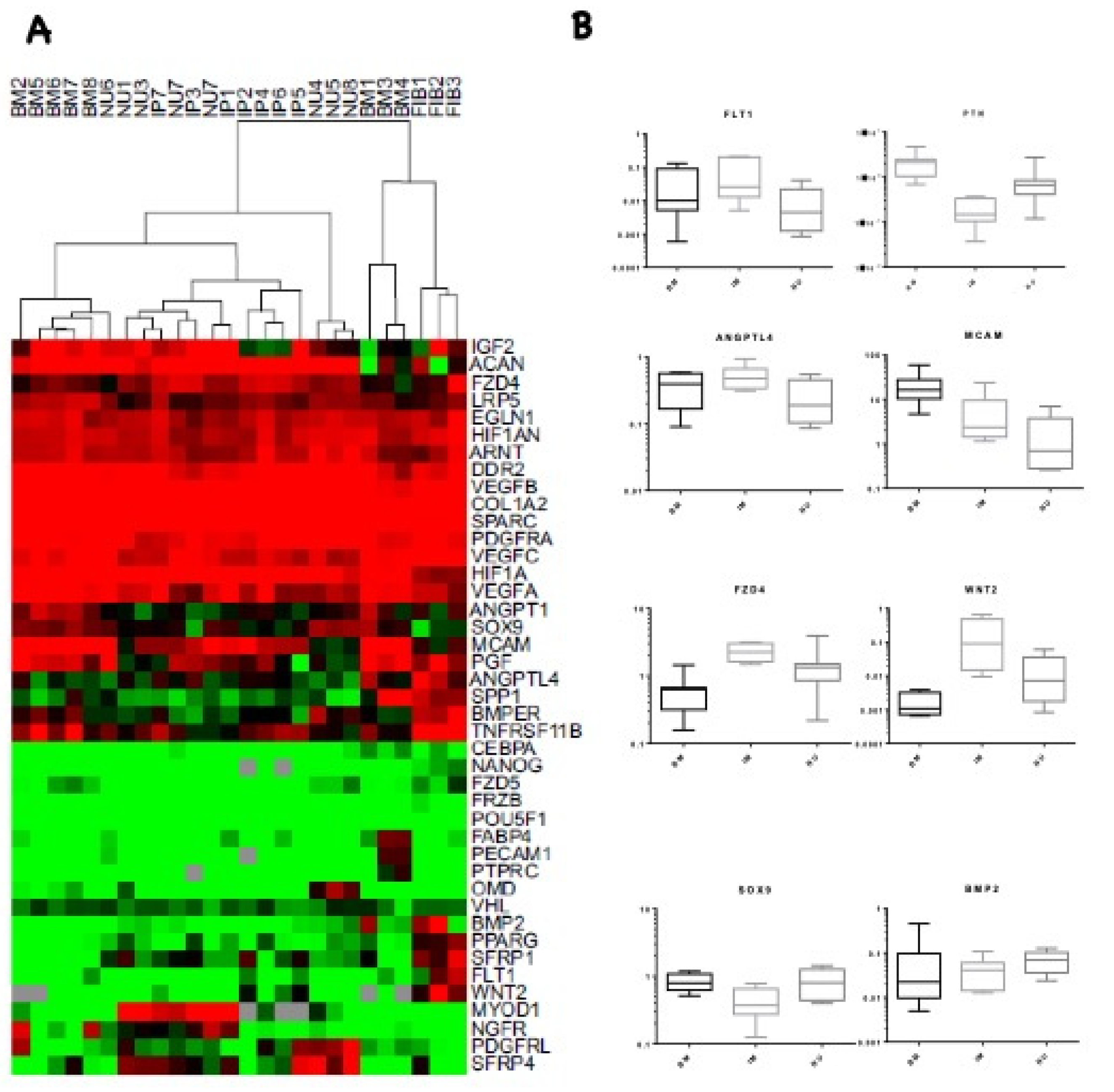
3.4. Transcriptional Profile Comparison of IP and NU Derived MSCs
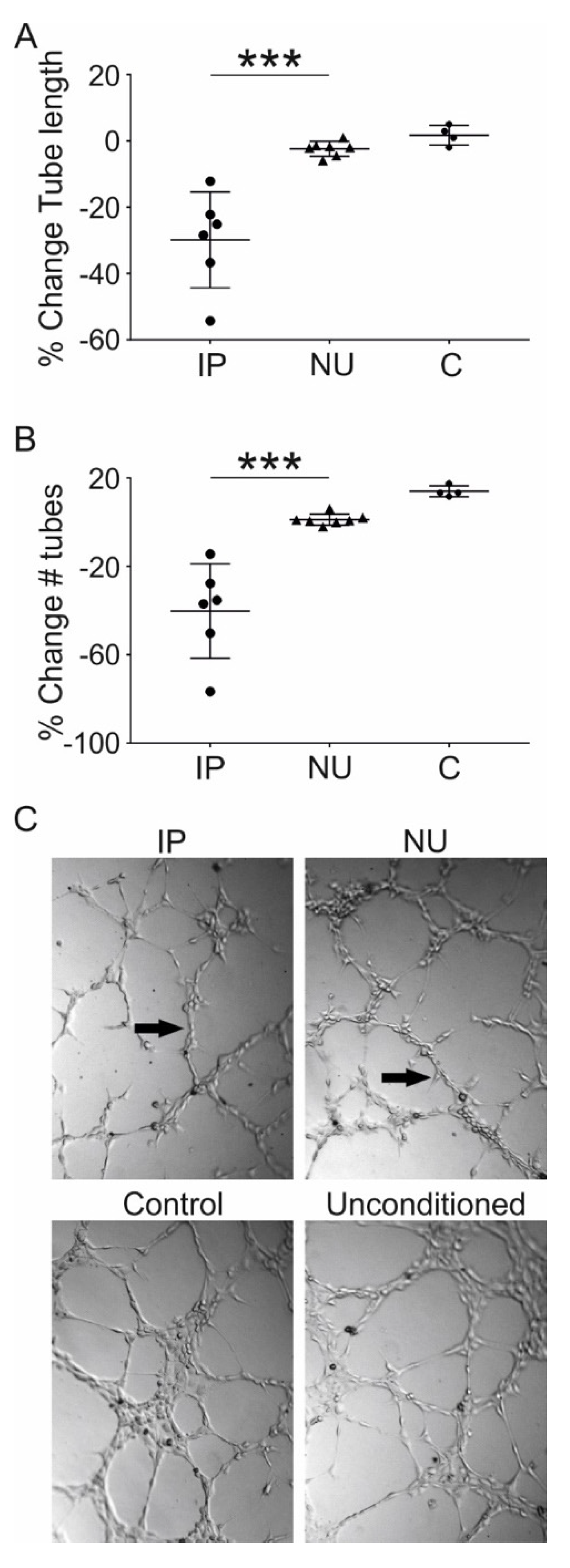
3.5. Assessment of MSC Mediated Angiogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mills, L.A.; Simpson, A.H. The relative incidence of fracture non-union in the Scottish population (5.17 million): A 5-year epidemiological study. BMJ Open 2013, 3, e002276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hak, D.J.; Fitzpatrick, D.; Bishop, J.A.; Marsh, J.L.; Tilp, S.; Schnettler, R.; Simpson, H.; Alt, V. Delayed union and nonunions: Epidemiology, clinical issues, and financial aspects. Injury 2014, 45, S3–S7. [Google Scholar] [CrossRef]
- Mills, L.; Tsang, J.; Hopper, G.; Keenan, G.; Simpson, A.H. The multifactorial aetiology of fracture nonunion and the importance of searching for latent infection. Bone Jt. Res. 2016, 5, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Megas, P. Classification of non-union. Injury 2005, 36 (Suppl. 4), S30–S37. [Google Scholar]
- Kostenuik, P.; Mirza, F.M. Fracture healing physiology and the quest for therapies for delayed healing and nonunion. J. Orthop. Res. 2016, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Panteli, M.; Pountos, I.; Jones, E.; Giannoudis, P.V. Biological and molecular profile of fracture non-union tissue: Current insights. J. Cell. Mol. Med. 2015, 19, 685–713. [Google Scholar] [CrossRef]
- Verdonk, R.; Goubau, Y.; Almqvist, F.K.; Verdonk, P. Biological methods to enhance bone healing and fracture repair. Arthroscopic 2015, 31, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Dimitriou, R.; Carr, I.M.; West, R.M.; Markham, A.F.; Giannoudis, P.V. Genetic predisposition to fracture non-union: A case control study of a preliminary single nucleotide polymorphisms analysis of the BMP pathway. BMC Musculoskelet. Disord. 2011, 12, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calori, G.M.; Albisetti, W.; Agus, A.; Iori, S.; Tagliabue, L. Risk factors contributing to fracture non-unions. Injury 2007, 38, S11–S18. [Google Scholar] [CrossRef]
- Hernigou, P.; Poignard, A.; Beaujean, F.; Rouard, H. Percutaneous autologous bone-marrow grafting for nonunions—Influence of the number and concentration of progenitor cells. J. Bone Jt. Surg. 2005, 87, 1430–1437. [Google Scholar]
- Granero-Moltó, F.; Weis, J.A.; Miga, M.I.; Landis, B.; Myers, T.J.; O’Rear, L.; Longobardi, L.; Jansen, E.D.; Mortlock, D.P.; Spagnoli, A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells 2009, 27, 1887–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calori, G.M.; Colombo, M.; Mazza, E.; Ripamonti, C.; Mazzola, S.; Marelli, N.; Mineo, G.V. Monotherapy vs. polytherapy in the treatment of forearm non-unions and bone defects. Injury 2013, 44 (Suppl. 1), S63–S69. [Google Scholar] [CrossRef]
- Giannoudis, P.V.; Ahmad, M.A.; Mineo, G.V.; Tosounidis, T.I.; Calori, G.M.; Kanakaris, N.K. Subtrochanteric fracture non-unions with implant failure managed with the “Diamond” concept. Injury 2013, 44 (Suppl. 1), S76–S81. [Google Scholar] [CrossRef]
- Goel, A.; Sangwan, S.S.; Siwach, R.C.; Ali, A.M. Percutaneous bone marrow grafting for the treatment of tibial non-union. Injury 2005, 36, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Hernigou, P.; Poignard, A.; Manicom, O.; Mathieu, G.; Rouard, H. The use of percutaneous autologous bone marrow transplantation in nonunion and avascular necrosis of bone. J. Bone Jt. Surg. Br. 2005, 87, 896–902. [Google Scholar] [CrossRef]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Fouraschen, S.M.; Hall, S.R.; de Jonge, J.; van der Laan, L.J. Support of hepatic regeneration by trophic factors from liver-derived mesenchymal stromal/stem cells. Methods. Mol. Biol. 2014, 1213, 89–104. [Google Scholar]
- Wang, M.; Rahnama, R.; Cheng, T.; Grotkopp, E.; Jacobs, L.; Limburg, S.; Kim, H.T.; Kuo, A.C. Trophic stimulation of articular chondrocytes by late-passage mesenchymal stem cells in coculture. J. Orthop. Res. 2013, 31, 1936–1942. [Google Scholar] [CrossRef]
- Qin, H.H.; Filippi, C.; Sun, S.; Lehec, S.; Dhawan, A.; Hughes, R.D. Hypoxic preconditioning potentiates the trophic effects of mesenchymal stem cells on co-cultured human primary hepatocytes. Stem Cell Res. Ther. 2015, 6, 237. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Huang, W.; Feng, Y.; Cai, W.; Wang, Y.; Wang, X.; Liang, J.; Wani, M.; Chen, J.; Zhu, P.; et al. MicroRNA-377 regulates mesenchymal stem cell-induced angiogenesis in ischemic hearts by targeting VEGF. PLoS ONE 2014, 9, e104666. [Google Scholar] [CrossRef]
- Premer, C.; Blum, A.; Bellio, M.A.; Schulman, I.H.; Hurwitz, B.E.; Parker, M.; Dermarkarian, C.R.; DiFede, D.L.; Balkan, W.; Khan, A.; et al. Allogeneic Mesenchymal Stem Cells Restore Endothelial Function in Heart Failure by Stimulating Endothelial Progenitor Cells. EBioMedicine 2015, 2, 467–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. All MSCs are pericytes? Cell Stem Cell 2008, 3, 229–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, I.R.; West, C.C.; Hardy, W.R.; James, A.W.; Park, T.S.; Nguyen, A.; Tawonsawatruk, T.; Lazzari, L.; Soo, C.; Peault, B. Natural history of mesenchymal stem cells, from vessel walls to culture vessels. Cell Mol. Life Sci. 2014, 71, 1353–1374. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Reed, A.A.; Joyner, C.J.; Brownlow, H.C.; Simpson, A.H. Human atrophic fracture non-unions are not avascular. J. Orthop. Res. 2002, 20, 593–599. [Google Scholar] [CrossRef]
- Bajada, S.; Marshall, M.J.; Wright, K.T.; Richardson, J.B.; Johnson, W.E. Decreased osteogenesis, increased cell senescence and elevated Dickkopf-1 secretion in human fracture non union stromal cells. Bone 2009, 45, 726–735. [Google Scholar] [CrossRef]
- Lawton, D.M.; Andrew, J.G.; Marsh, D.R.; Hoyland, J.A.; Freemont, A.J. Expression of the gene encoding the matrix gla protein by mature osteoblasts in human fracture non-unions. Mol. Pathol. 1999, 52, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Pontikoglou, C.; Deschaseaux, F.; Sensebé, L.; Papadaki, H.A. Bone marrow mesenchymal stem cells: Biological properties and their role in hematopoiesis and hematopoietic stem cell transplantation. Stem Cell Rev. Rep. 2011, 7, 569–589. [Google Scholar] [CrossRef]
- Ruf, W. Hemostasis keeps the stem cell niche in order. Blood 2016, 128, 1027–1029. [Google Scholar] [CrossRef] [Green Version]
- Masquelet, A.C. Muscle reconstruction in reconstructive surgery: Soft tissue repair and long bone reconstruction. Langenbeck’s archives of surgery. Dtsch. Ges. Chir. 2003, 388, 344–346. [Google Scholar]
- Retzepi, M.; Donos, N. Guided Bone Regeneration: Biological principle and therapeutic applications. Clin. Oral Implants Res. 2010, 21, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Giannoudis, P.V.; Faour, O.; Goff, T.; Kanakaris, N.; Dimitriou, R. Masquelet technique for the treatment of bone defects: Tips-tricks and future directions. Injury 2011, 42, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Karger, C.; Kishi, T.; Schneider, L.; Fitoussi, F.; Masquelet, A.C. Treatment of posttraumatic bone defects by the induced membrane technique. J. Orthop. Surg. Res. 2012, 98, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Masquelet, A.C.; Fitoussi, F.; Begue, T.; Muller, G.P. Reconstruction of the long bones by the induced membrane and spongy autograft. Ann. Chir. Plast. Esthet. 2000, 45, 346–353. [Google Scholar]
- Gruber, H.E.; Riley, F.E.; Hoelscher, G.L.; Bayoumi, E.M.; Ingram, J.A.; Ramp, W.K.; Bosse, M.J.; Kellam, J.F. Osteogenic and chondrogenic potential of biomembrane cells from the PMMA—Segmental defect rat model. J. Orthop. Res. 2012, 30, 1198–1212. [Google Scholar] [CrossRef]
- Masquelet, A.C.; Begue, T. The concept of induced membrane for reconstruction of long bone defects. Orthop. Clin. N. Am. 2010, 41, 27–37. [Google Scholar] [CrossRef]
- Cuthbert, R.J.; Churchman, S.M.; Tan, H.B.; McGonagle, D.; Jones, E.; Giannoudis, P.V. Induced periosteum a complex cellular scaffold for the treatment of large bone defects. Bone 2013, 57, 484–492. [Google Scholar] [CrossRef]
- Cuthbert, R.; Boxall, S.A.; Tan, H.B.; Giannoudis, P.V.; McGonagle, D.; Jones, E. Single-platform quality control assay to quantify multipotential stromal cells in bone marrow aspirates prior to bulk manufacture or direct therapeutic use. Cytotherapy 2012, 14, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.; Churchman, S.M.; English, A.; Buch, M.H.; Horner, E.A.; Burgoyne, C.H.; Reece, R.; Kinsey, S.; Emery, P.; McGonagle, D.; et al. Mesenchymal stem cells in rheumatoid synovium: Enumeration and functional assessment in relation to synovial inflammation level. Ann. Rheum. Dis. 2010, 69, 450–457. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, A.M.; Saim, A.B.; Tan, K.K.; Tan, G.H.; Mokhtar, S.A.; Rose, I.M.; Othman, F.; Idrus, R.B. Comparison of bioengineered human bone construct from four sources of osteogenic cells. J. Orthop. Sci. 2005, 10, 192–199. [Google Scholar] [CrossRef] [PubMed]
- De Bari, C.; Dell’Accio, F.; Vanlauwe, J.; Eyckmans, J.; Khan, I.; Archer, C.W.; Jones, E.A.; McGonagle, D.G.; Mitsiadis, C.; Pitzalis, C.; et al. Mesenchymal multipotency of adult human periosteal cells demonstrated by single-cell lineage analysis. Arthritis Rheum. 2006, 54, 1209–2121. [Google Scholar] [CrossRef] [Green Version]
- De Bari, C.; Dell’Accio, F.; Karystinou, A.; Guillot, P.V.; Fisk, N.M.; Jones, E.A.; McGonagle, D.; Khan, I.M.; Archer, C.W.; Mitsiadis, T.A.; et al. A biomarker-based mathematical model to predict bone-forming potency of human synovial and periosteal mesenchymal stem cells. Arthritis Rheum. 2008, 58, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnsdorf, E.J.; Jones, L.M.; Carter, D.R.; Jacobs, C.R. The Periosteum as a Cellular Source for Functional Tissue Engineering. Tissue Eng. Part A 2009, 15, 2637–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.; English, A.; Churchman, S.M.; Kouroupis, D.; Boxall, S.A.; Kinsey, S.; Giannoudis, P.G.; Emery, P.; McGonagle, D. Large-scale extraction and characterization of CD271+ multipotential stromal cells from trabecular bone in health and osteoarthritis: Implications for bone regeneration strategies based on uncultured or minimally cultured multipotential stromal cells. Arthritis Rheum. 2010, 62, 1944–1954. [Google Scholar]
- Jones, E.A.; English, A.; Kinsey, S.E.; Emery, P.; McGonagle, D.G.; Jones, R.A.; Straszynski, L.; Markham, A.F.; Meredith, D.M.; Jack, A. Isolation and characterization of bone marrow multipotential mesenchymal progenitor cells. Arthritis Rheum. 2002, 46, 3349–3360. [Google Scholar] [CrossRef]
- Parthymou, A.; Kardamakis, D.; Pavlopoulos, I.; Papadimitriou, E. Irradiated C6 glioma cells induce angiogenesis in vivo and activate endothelial cells in vitro. Int. J. Cancer 2004, 110, 807–814. [Google Scholar] [CrossRef]
- Chen, C.W.; Montelatici, E.; Crisan, M.; Corselli, M.; Huard, J.; Lazzari, L.; Peault, B. Perivascular multi-lineage progenitor cells in human organs: Regenerative units, cytokine sources or both? Cytokine Growth Factor Rev. 2009, 20, 429–434. [Google Scholar] [CrossRef]
- Churchman, S.M.; Ponchel, F.; Boxall, S.A.; Cuthbert, R.; Kouroupis, D.; Roshdy, T.; Giannoudis, P.V.; Emery, P.; McGonagle, D.; Jones, E.A. Transcriptional profile of native CD271+ multipotential stromal cells: Evidence for multiple fates, with prominent osteogenic and Wnt pathway signaling activity. Arthritis Rheum. 2012, 64, 2632–2643. [Google Scholar] [CrossRef]
- Bejar, M.T.; Ferrer-Lorente, R.; Pena, E.; Badimon, L. Inhibition of Notch rescues the angiogenic potential impaired by cardiovascular risk factors in epicardial adipose stem cells. FASEB J. 2016, 30, 2849–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, C.P.; Micklem, K.; Watt, S.M. A comparison of methods for quantifying angiogenesis in the Matrigel assay in vitro. Tissue Eng. Part C Methods 2011, 17, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappas, N.C.; Zeng, G.; Chappell, J.C.; Kearney, J.B.; Hazarika, S.; Kallianos, K.G.; Patterson, C.; Annex, B.H.; Bautch, V.L. The VEGF receptor Flt-1 spatially modulates Flk-1 signaling and blood vessel branching. J. Cell Biol. 2008, 181, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, D.M.; Kearney, J.B.; Johnson, J.H.; Rosenberg, M.P.; Kumar, R.; Bautch, V.L. The vascular endothelial growth factor (VEGF) receptor Flt-1 (VEGFR-1) modulates Flk-1 (VEGFR-2) signaling during blood vessel formation. Am. J. Pathol. 2004, 164, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, C.; Borsari, V.; Falconi, M.; Gigante, A.; Lazzarini, R.; Fini, M.; Di Primio, R.; Mattioli-Belmonte, M. Human periosteum-derived stem cells for tissue engineering applications: The role of VEGF. Stem Cell Rev. Rep. 2012, 8, 882–890. [Google Scholar] [CrossRef]
- Fong, G.-H.; Zhang, L.; Bryce, D.-M.; Peng, J. Increased hemangioblast commitment, not vascular disorganization, is the primary defect in flt-1 knock-out mice. Development 1999, 126, 3015–3025. [Google Scholar]
- Chappell, J.C.; Cluceru, J.G.; Nesmith, J.E.; Mouillesseaux, K.P.; Bradley, V.B.; Hartland, C.M.; Hashambhoy-Ramsay, Y.L.; Walpole, J.; Peirce, S.M.; Mac Gabhann, F.; et al. Flt-1 (VEGFR-1) coordinates discrete stages of blood vessel formation. Cardiovasc. Res. 2016, 111, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Lamprou, M.; Kaspiris, A.; Panagiotopoulos, E.; Giannoudis, P.V.; Papadimitriou, E. The role of pleiotrophin in bone repair. Injury 2014, 45, 1816–1823. [Google Scholar] [CrossRef]
- Zhang, L.; Kundu, S.; Feenstra, T.; Li, X.; Jin, C.; Laaniste, L.; El Hassan, T.E.A.; Ohlin, K.E.; Yu, D.; Olofsson, T. Pleiotrophin promotes vascular abnormalization in gliomas and correlates with poor survival in patients with astrocytomas. Sci. Signal. 2015, 8, ra125. [Google Scholar] [CrossRef]
- Li, G.; Bunn, J.; Mushipe, M.; He, Q.; Chen, X. Effects of pleiotrophin (PTN) over-expression on mouse long bone development, fracture healing and bone repair. Calcif. Tissue Int. 2005, 76, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Takita, H.; Ohata, N.; Tamura, M.; Kuboki, Y. Pleiotrophin regulates bone morphogenetic protein (BMP)-induced ectopic osteogenesis. J. Biochem. 2002, 131, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Iampietro, M.; Chuchana, P.; Guerit, D.; Djouad, F.; Noel, D.; Jorgensen, C. Involvement of angiopoietin-like 4 in matrix remodeling during chondrogenic differentiation of mesenchymal stem cells. J. Biol. Chem. 2014, 289, 8402–8412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.L.; Teo, Z.Q.; Chong, H.C.; Zhu, P.K.; Tan, M.J.; Tan, C.K.; Lam, C.R.I.; Sng, M.K.; Leong, D.T.W.; Tan, S.M.; et al. ANGPTL4 modulates vascular junction integrity by integrin signaling and disruption of intercellular VE-cadherin and claudin-5 clusters. Blood 2011, 118, 3990–4002. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yang, Y.H.; Basile, J.R. The Semaphorin 4D-Plexin-B1-RhoA signaling axis recruits pericytes and regulates vascular permeability through endothelial production of PDGF-B and ANGPTL4. Angiogenesis 2014, 17, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Gomez Perdiguero, E.; Liabotis-Fontugne, A.; Durand, M.; Faye, C.; Ricard-Blum, S.; Simonutti, M.; Augustin, S.; Robb, B.M.; Paques, M.; Valenzuela, D.M.; et al. ANGPTL4-alphavbeta3 interaction counteracts hypoxia-induced vascular permeability by modulating Src signalling downstream of vascular endothelial growth factor receptor 2. J. Pathol. 2016, 240, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Nurcombe, V.; Cool, S.M. Wnt signaling controls the fate of mesenchymal stem cells. Gene 2009, 433, 1–7. [Google Scholar] [CrossRef]
- Gong, Y.; Slee, R.B.; Fukai, N.; Rawadi, G.; Roman-Roman, S.; Reginato, A.M.; Wang, H.; Cundy, T.; Glorieux, F.H.; Lev, D.; et al. Osteoporosis-Pseudoglioma Syndrome Collaborative G. LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell 2001, 107, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Babij, P.; Zhao, W.; Small, C.; Kharode, Y.; Yaworsky, P.J.; Bouxsein, M.L.; Reddy, P.S.; Bodine, P.V.; Robinson, J.A.; Bhat, B.; et al. High bone mass in mice expressing a mutant LRP5 gene. J. Bone Miner Res. 2003, 18, 960–974. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.I.; Park, Y.S.; Im, G.I. Changes in the epigenetic status of the SOX-9 promoter in human osteoarthritic cartilage. J. Bone Miner. Res. 2013, 28, 1050–1060. [Google Scholar] [CrossRef]
- Chappell, J.C.; Darden, J.; Payne, L.B.; Fink, K.; Bautch, V.L. Blood Vessel Patterning on Retinal Astrocytes Requires Endothelial Flt-1 (VEGFR-1). J. Dev. Biol. 2019, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Chang, M.; Tsai, K.; Hung, M.; Chen, H.; Hung, S. Mesenchymal stem cells promote growth and angiogenesis of tumors in mice. Oncogene 2013, 32, 4343–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckermann, B.; Kallifatidis, G.; Groth, A.; Frommhold, D.; Apel, A.; Mattern, J.; Salnikov, A.; Moldenhauer, G.; Wagner, W.; Diehlmann, A. VEGF expression by mesenchymal stem cells contributes to angiogenesis in pancreatic carcinoma. Br. J. Cancer 2008, 99, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Han, Z.-P.; Zhang, S.-S.; Jing, Y.-Y.; Bu, X.-X.; Wang, C.-Y.; Sun, K.; Jiang, G.-C.; Zhao, X.; Li, R. Effects of inflammatory factors on mesenchymal stem cells and their role in the promotion of tumor angiogenesis in colon cancer. J. Biol. Chem. 2011, 286, 25007–25015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuthbert, R.J.; Jones, E.; Sanjurjo-Rodríguez, C.; Lotfy, A.; Ganguly, P.; Churchman, S.M.; Kastana, P.; Tan, H.B.; McGonagle, D.; Papadimitriou, E.; et al. Regulation of Angiogenesis Discriminates Tissue Resident MSCs from Effective and Defective Osteogenic Environments. J. Clin. Med. 2020, 9, 1628. https://doi.org/10.3390/jcm9061628
Cuthbert RJ, Jones E, Sanjurjo-Rodríguez C, Lotfy A, Ganguly P, Churchman SM, Kastana P, Tan HB, McGonagle D, Papadimitriou E, et al. Regulation of Angiogenesis Discriminates Tissue Resident MSCs from Effective and Defective Osteogenic Environments. Journal of Clinical Medicine. 2020; 9(6):1628. https://doi.org/10.3390/jcm9061628
Chicago/Turabian StyleCuthbert, R. J., E. Jones, C. Sanjurjo-Rodríguez, A. Lotfy, P. Ganguly, S. M. Churchman, P. Kastana, H. B. Tan, D. McGonagle, E. Papadimitriou, and et al. 2020. "Regulation of Angiogenesis Discriminates Tissue Resident MSCs from Effective and Defective Osteogenic Environments" Journal of Clinical Medicine 9, no. 6: 1628. https://doi.org/10.3390/jcm9061628