Abstract
Background: Intestinal infections remain a major public health burden in developing countries. Due to social, ecological, environmental, and cultural conditions, Indigenous peoples in Colombia are at particularly high risk. Materials: 137 stool samples were analyzed by microscopy and real-time-Polymerase Chain Reaction (RT-PCR), targeting protozoan parasites (Giardia intestinalis, Entamoeba histolytica, Cryptosporidium spp., and Cyclospora cayetanensis), bacteria (Campylobacter jejuni, Salmonella spp., Shigella ssp./enteroinvasive E. coli (EIEC), Yersinia spp., enterohemorrhagic E. coli (EHEC), enteropathogenic E. coli (EPEC), enterotoxin-producing E. coli (ETEC), enteroaggregative E. coli (EAEC), and Tropheryma whipplei), and helminths (Necator americanus, Strongyloides stercoralis, Ascaris lumbricoides, Ancylostoma spp., Trichuris. trichiura, Taenia spp., Hymenolepis nana, Enterobius vermicularis, and Schistosoma spp.). Microscopy found additional cases of helminth infections. Results: At least one pathogen was detected in 93% of the samples. The overall results revealed protozoa in 79%, helminths in 69%, and bacteria in 41%. G. intestinalis (48%), Necator/hookworm (27%), and EAEC (68%) were the most common in each group. Noteworthy, T. whipplei was positive in 7% and T. trichirua in 23% of the samples. A significant association of one infection promoting the other was determined for G. intestinalis and C. jejuni, helminth infections, and EIEC. Conclusions: The results illustrate the high burden of gastrointestinal pathogens among Indigenous peoples compared to other developing countries. Countermeasures are urgently required.
1. Introduction
The indigenous tribe called Wiwa lives in retracted areas in the Sierra Nevada de Santa Marta in the northeast of Colombia. Since Wiwa communities avoid contact with the outside world, little is known, e.g., about their burden of gastrointestinal infections. According to the Wiwas, gastrointestinal complaints are the leading problem in their communities, causing high morbidity and mortality rates. The focus of this study was on bacterial and protozoan infections, as the area’s climate and poor socio-economic conditions (simple housing, lack of sanitation, etc.) provided favorable conditions for these pathogens. Furthermore, the Wiwa’s traditional mode of agriculture (top dressing), living together with livestock, and poor drinking water access (rivers and unprotected wells) likely contribute to high infection rates.
These kind of infections cause approximately 2 billion diarrhetic episodes worldwide and kill about 5.8 million children (2015) [1]. A further major analysis factor for this study was soil transmitted helminths (STHs), as they belong to the “Big Five” of neglected tropical diseases [2].
The main aim was to find evidence for and determine the prevalence and underlying causes of gastrointestinal infections in Wiwa communities for effective countermeasures to be implemented.
2. Material and Methods
2.1. Ethical Approval
The study was performed in line with the Declaration of Helsinki. It was approved by the Ethics Committee of Valledupar, Cesar, Colombia (Acta no 0022013). Written informed consent was obtained from each participant or the parent/legal guardian of a child prior to participation. All participants were informed about their results and received treatment, if appropriate.
2.2. General Information
In July and November 2014, 137 indigenous volunteers from the villages of Tezhumake (Department Cesar) and Seminke (Department La Guajira) provided stool samples. This corresponds to 80% and 50% of the respective populations living in these communities. Additionally, 13 samples came from Valledupar (Department Cesar), a larger city in the area. In total, 102 participants provided complete anthropometric and clinical data, and a physical examination was performed by a specialist for internal medicine. In 34 cases, individual pieces of information were missing (e.g., age or weight). These values were statistically treated as missing, which means that an imputation procedure was not conducted.
2.3. Nucleic Acid Extractions and Polymerase Chain Reactions (PCRs)
DNA was extracted using a QIAmp DNA stool mini kit (QIAGEN, Hilden, Germany), following the manufacturer’s protocol. Extracts were thereafter stored at −20 °C. A part of the stool samples was preserved in 5% formalin to perform the microscopic analysis. PCR investigations were carried out on site. In addition, extracts and formalin samples were sent to the Bernhard–Nocht Institute for Tropical Medicine (BNITM, Hamburg, HH, Germany). During the BNITM microscopy, control examinations and further analysis were performed. Airfreight requirements were fulfilled, the cooling chain was not interrupted (based on the temperature control documentation), and transport was done by a specialized company (World Courier, Frankfurt, HE, Germany).
In-house real-time multiplex PCRs for protozoan and helminthic parasites targeting Entamoeba histolytica, Giardia intestinalis, Cryptosporidium spp., and Cyclospora cayetanensis, as well as Necator americanus, Strongyloides stercoralis, Ascaris lumbricoides, Ancylostoma spp., Trichuris trichiura, Schistosoma spp., Enterobius vermicularis, Taenia saginata, Taenia solium, and Hymenolepis nana were performed as described before [3]. Further, enteroinvasive bacterial pathogens like Campylobacter jejuni, Salmonella spp., Shigella ssp./enteroinvasive E. coli (EIEC), and Yersinia spp. were assessed by in-house real-time PCR [4]. To identify diarrheagenic E. coli infections, Rida Gene RT-PCR assays for enterohemorrhagic Escherichia coli (EHEC), enteropathogenic E. coli (EPEC), enteroinvasive E. coli (EIEC), enterotoxin-producing E. coli (ETEC), enteroinvasive E. coli (EIEC), and enteroaggregative E. coli (EAEC) (R-Biopharm AG, Darmstadt, HE, Germany) were used, as described before [5]. Furthermore, a species-specific PCR for Tropheryma whipplei [6] was applied [7]. The DNA of phocid herpesvirus was included as an internal control for the in-house PCRs [8,9].
In all RT-PCRs runs, positive and negative controls were included: As positive controls, synthetically designed target sequences linked by EcoR1 endonuclease restriction sites and inserted into pEX-A128 vector backbones were used (Eurofins Scientific SE). Negative controls included PCT water samples that had undergone the whole nucleic acid extraction process to exclude sample contamination. All primers and probes of the in-house PCRs were purchased from Eurofins, Hamburg, HH, Germany. The assays were performed on a multi-channel RotorGene Q Cycler (Qiagen, Hilden, HE, Germany).
In addition, light microscopy of all stool samples by direct saline and/or iodine mounts and following a formol-ethyl acetate concentration was performed.
2.4. Demographic Data
To evaluate the average parameters related to size, weight, and growth, the official tables provided by the Organización Mundial de Salud 2006–2007 were used, which are valid for the Colombian population (most currently available tables). Individuals aged 0–19 years were scored by their weight–size, weight–age, and size–age tables, each corresponding to the male and female sex. The Body Mass Index (BMI) was calculated according to World Health Organization (WHO) recommendations.
2.5. Statistical Methods
Statistical analyses were carried out using R 3.6.1 (2020 Version 131073, Boston, MA, USA, Open Source) [10]. To compare the strength of association for various groups, the Mantel–Haenszel odds ratio (OR) with a 95% confidence interval (CI) was computed. To compare more than two groups, an Analysis of Variance (ANOVA) was calculated. The relation of the predictor and outcomes was analyzed using the logit model. For categorical data, Chi-Square test was employed. Z-scores for weight-for-age (wfa), length/height for age (hfa), and BMI-for-age based on the WHO Child Growth Standards were calculated using the R-package “zscorer”.
3. Results
3.1. Demographic Results
Of the 137 stool samples, 50% (68) were from female and 50% (69) from male volunteers. In the overall results, there was no sex dependency detectable (OR 1.24; 0.95-CI [0.36, 4.72]).
The average age was 24.6 years (SD = 18.16). Twenty-one samples were from children aged 0–6, 42 from adolescents aged 6–18, and 74 from adults older than 18 years. Stool consistency was normal in 18% (25) of cases; all other stool samples were fluid, pappy, or had a mucous texture. In 4 cases, the stool was additionally covered with blood (PCR showed in all cases an infection with E. histolytica).
The 137 questionnaires and physical examinations revealed that 88% (119) had no complaints, 10% (13) complained about diarrhea, 2% (2) noted abdominal pain, and 2% (2) suffered from diarrhea and abdominal pain. In one case, information was missing.
Of the 119 individuals who stated to have no complaints, only 7% (8) were negative in all tests. Of those who stated complaints, only 3% (4) were negative in all tests (which may be due to pathogens not included in the test panel). Whether persons stated complaints or not, only 9% (12) were negative in all tests performed. When using the reported complaints as a predictor, we found a sensitivity of 10% and a specificity of 60%. There was no statistical association between the subjective reporting of symptoms and having an intestinal pathogen X2 (1, n = 137) = 3.3, p > 0.05.
For individuals without complaints, 51% carried helminths, 39% bacteria and 55% protozoa. Of individuals with complaints, 59% carried helminths, 38% bacteria, and 60% protozoa.
The majority of volunteers (38%) complaining about diarrhea and/or abdominal pain carried 3–4 pathogens. Among these, the most frequent combination was C. jejuni, G. intestinalis, and EPEC accompanied by either A. lumbricoides, T. trichirua, or E. vermicularis. Taking the PCR and microscopy results together, at least one pathogen was found in 93% of the samples, at least two in 69%, three in 52%, four in 37%, five in 19%, six in 12%, seven in 6%, eight in 3%, and nine in 1% (see Figure 1).
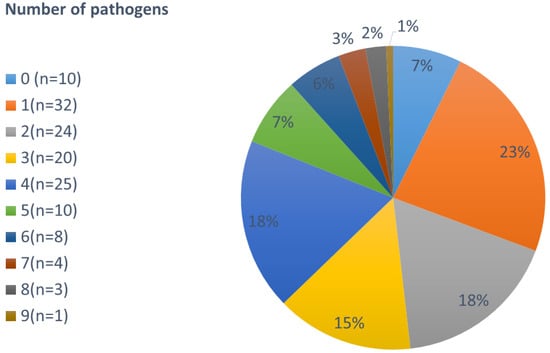
Figure 1.
Number of pathogens found per person (not stratified by age).
The maximum of 9 pathogens (C. jejuni, G. intestinalis, A. lumbricoides, T. trichiura, N. americanus, H. nana, E. vermicularis, EPEC and EIEC) was found in a seven-year old boy.
Children had significantly more infections (M = 3.81, SD = 2.34) than adults (M = 2.5, SD = 1.65), with adolescents ranking in the middle (M = 3.19, SD = 2.28). Figure 2 shows the number of infections in each age group. An ANOVA could be computed since Levene’s test for the homogeneity of variance was not significant (p = 0.24).
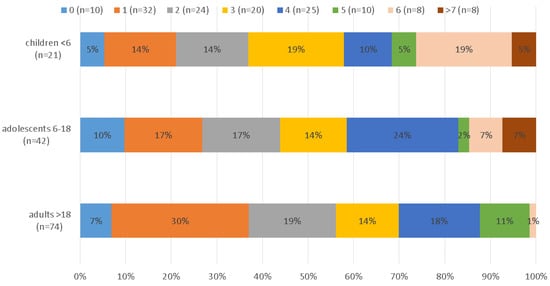
Figure 2.
Number of infecting pathogens stratified by age in years.
For protozoan parasites in particular, age was the most robust predictor: the older the individual, the less likely a positive result for protozoa. For more details, see Figure 3.
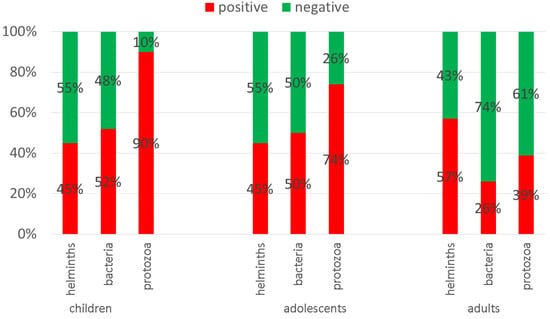
Figure 3.
Type of parasite per age group (n = type of parasite based on at least one positive detection).
3.2. Co-Infections
Co-infections were common: Bacterial and protozoan pathogens were present in 31%, protozoan and helminth infections in 34%, and bacterial and helminth pathogens in 25% of the cases. Infections with at least one pathogen from all three classes were seen in 21%.
Applying logistic regression, we found that the presence of bacteria significantly predicted protozoa and vice versa. When analyzing the predictive power of single parasites, some notable observations of interest can be explored:
Co-infection with C. jejuni and G. intestinalis was common, as C. jejuni showed a predictive value of 67% for G. intestinalis infections and a value of 43% for C. jejuni. Further symbiotic interactions were found for N. americanus/hookworm and EIEC/Shigella spp., A. lumbricoides, and T. trichiura.
S. stercoralis symbiotically interacts with T. trichiura and Necator/hookworm pathogens. Evidence suggests that S. stercoralis infections tend to support EIEC infections, rather than vice versa.
Notably, EPEC supports EAEC (90%) and ETEC infections (56%), EAEC supports EPEC (55%) and ETEC (46%), while ETEC only supports EPEC (63%). Interestingly, this observation is not true for various other pathogens, such as E. histolytica, E. nana, and Salmonella spp. For more detailed information, see Table 1.

Table 1.
Results of the logistic regression for single parasites. Significance: 0 ***, 0.001 **, 0.01 *. C. jejuni = Campylobacter jejuni, EAEC = enteroaggregative E. coli, ETEC = enterotoxin-producing E. coli, EHEC = enterohemorrhagic E. Coli, EPEC = enteropathogenic E. coli, EIEC/Shig. = Enteroinvasive E. coli/Shigella spp., Giardia = G. intestinalis, E. histo = Entamoeba histolytica, Necator = N. americanus, S. stercoralis = Strongyloides stercoralis, Ascaris = A. lumbricoides, Trichuris = T. trichiura, H. nana = Hymenolepis nana, Enterobius = E. vermicularis.
3.3. PCR Based Results
3.3.1. Protozoa
Overall, the protozoan PCRs indicated 79% positive results. G. intestinalis was the most common pathogen (48%), followed by C. cayetanensis (12%). Seventeen of the positive results for C. cayetanensis occurred in children (59%); 50% of these were in those 1–4 years of age and 50% were in those 5–12 years of age. E. histolytica was found in 7% of the samples. Cryptosporidium spp. (1%) was only detected in one sample from a 37-year old man with an unknown immune status. He did not report any symptoms but was also positive for EPEC, G. intestinalis, and C. jejuni.
3.3.2. Bacteria
The real-time-PCRs revealed positive results for bacteria in 41% of the samples. The leading bacteria within the E. coli group was EAEC (67%), followed by ETEC (53%), EPEC (42%), EHEC (20%), and Shigella/EIEC (18%). In the other bacterial panel, the most common pathogens were C. jejuni, Salmonella spp., and T. whipplei, with 31%, 5%, and 7%, respectively. Yersinia spp. did not occur.
3.3.3. Helminths
As a leading helminthic infectious agent, T. trichiura was found in 23% of the cases, followed by Hymenolepis nana (21%), N. americanus (18%), A. lumbricoides (13%), Taenia (4%), S. stercoralis (4%), and E. vermicularis (2%). Ancylostoma spp. and Schistosoma spp. were not found.
3.4. Microscopy Based Results
The microscopy results showed T. trichiura (20%), hookworms (18%), H. nana (13%), A. lumbricoides (11%), G. intestinalis (3%), and E. vermicularis (2%). N. americanus was found to be positive in 25 PCR runs and 24 times via microscopy (classified as Hookworm, with 92% agreement, kappa 0.73, p < 0.001). For A. lumbricoides, the results were positive under PCR 17 times and 15 times under microscopy (with a 96% agreement, kappa 0.79, p < 0.001). For G. intestinalis, PCR revealed 66 positive results, while microscopy revealed only 4 (54% agreement, kappa 0.06, p < 0.05). Microscopy missed infections compared to PCR in 40% of the cases (false negative); positive results not detected in PCR occurred in 11% of the cases.
3.5. Villages
Stratified by village, there were 172 positive results in Tezhumake, 202 in Seminke, and 18 in Valledupar. In Seminke, 100% of individuals tested carried at least one pathogen, in Tezhumake 90%, and in Valledupar 85%. See Table 2 for further results. Of all pathogens found within the age group of children <6 years of age, 64% were bacteria, 21% were protozoa, and 15% were helminths. Of all bacteria found in children <6 years of age, C. jejuni was found in 13% and Shigella spp. 0%.
Table 2.
Gastrointestinal organisms identified in stool samples by species groups stratified by village and age groups. C. jejuni = Campylobacer jejuni, EIEC = enteroinvasive E. coli, EHEC = enterohemorrhagic E. coli, ETEC = enterotoxin-producing E. coli, EAEC = enteroaggregative E. coli, EPEC = enteropathogenic E. coli, T. whipplei = Tropheryma whipplei, G. intestinalis = Giardia intestinalis, E. histolytica = Entamoeba histolytica, C. cayetanensis = Cyclospora cayetanensis, N. americanus = Necator americanus, S. stercoralis = Strongyloides stercoralis, T. trichiura = Trichuris trichirua, H. nana = Hymenolepis nana, E. vermicularis = Enterobius vermicularis.
In total, 70 of the 137 volunteers (51%) showed a positive result in the PCR targeting helminths. Eighty-six-percent of the cases (37) from Seminke carried at least one helminth infection, 37% (30) from Tezhumake, and 23% (3) from Valledupar.
Using logistic regression and referring to positive helminth cases as a subpopulation, we found that volunteers from Seminke were three times as likely to have an infection with helminths than volunteers from Tezhumake (X2 (2, n = 137) = 31.51, p > 0.001). Multiple infections with two or three pathogens were most common in Seminke (74%) (see also Table 2 and Figure 4).
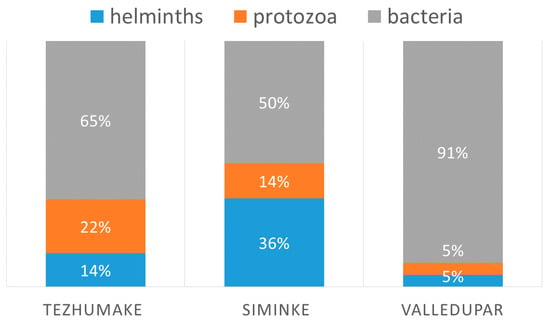
Figure 4.
Comparison of infections related to villages.
3.6. Demographic Data
Weight ranged between 11.5 and 77.0 kg, with a mean weight of 43.6 kg (Md = 47.5; SD = 16.2). Weight stratified by sex revealed a mean weight of 42.4 kg for men and 40.8 kg for women. Height varied between 63 and 180 cm, with a mean size of 138.7 cm (Md = 145.0; SD = 24.5). Overall, 50% of the volunteers were ≤144 cm, and 75% were ≤ 153 cm. Height revealed a mean value of 132.6 cm for women and 139.36 cm for men (normal range for Colombians: 158.65 cm for women and 170.64 cm for man). The reference curves for height and weight revealed critical age stage results (see Figure 5) Low CT values in the PCRs, indicating a high pathogen burden, were found in most cases. The majority of children had a BMI above average for their age.
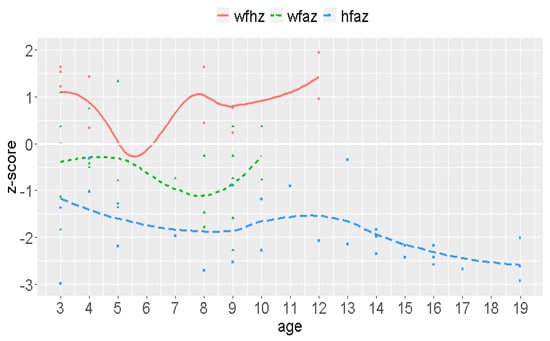
Figure 5.
Weight-for-height (wfhz), weight-for-age (wfaz), height for age (hfaz) z-scores.
4. Discussion
The Wiwas live in very retracted areas of the Sierra Nevada de Santa Marta, north-east of Colombia. The burden of pathogens is much higher in this area than in most other developing countries. In total, 93% of the Wiwa population showed one or more infection(s). All 137 samples contained a total of 538 pathogens.
G. intestinalis is the most frequent protozoan parasite in developing countries [11,12,13,14]. In contrast to the average infection rates of 20–30% in Guatemala [15] and 13–43% in Haiti [16], the prevalence in our communities was 48%. Interestingly, only 50% of the infected individuals stated complaints, which may point to asymptomatic G. intestinalis carriers [17,18]. The higher prevalence found in younger children (81% of the infections) is in accordance with previous observations [19]. The observed differences in positive findings for G. intestinalis by microscopy and real-time PCR can be explained by the higher sensitivity of the well-established PCR technique [3] and is a generally well-known phenomenon [20]. Furthermore, the low sensitivity of microscopy might suggest pathogen concentrations close to or below the microscopic detection threshold, making low-replicative colonization more likely than high-replicative infection.
E. histolytica was found in 7% of the cases and was diagnosed by highly specific PCR, which is able to distinguish between E. histolytica and the pathogen forms E. dispar/moshkovskii [21,22]. Compared to studies performed in rural areas of Venezuela [23] and Mexico [24], where a prevalence of 11% and 14% was respectively detected, the occurrence rate among the Wiwas seems to be lower. The different diagnostic techniques applied (microscopy and different PCRs) might have affected these findings.
The rate of C. cayetanensis was 12% in the communities. In comparison, the prevalence of C. cayetanensis was 3–7% in Haiti and 2.3% in Guatemala [25]. In accordance with previous findings [26], the majority of infections were found in children and adolescents (42.8%). Notably, 20% were found in indigenous adults.
Overall, one protective predictor was found for protozoal infections: With increasing age, the infection rates decreased. A similar effect was shown previously for enteric colonization with G. duodenalis in asymptomatic Madagascan children [27].
Within the bacterial panel, the E. coli family was dominant. EAEC is described as the second most common agent (24%) [28]. It is an emerging pathogen affecting children and adults worldwide, causing acute and persistent diarrhea [29]. We found EAEC in 15% of the children and 58% of the adults.
ETEC causes infections in up to 200 million people worldwide, accounting for approximately 100.000 deaths per year [30]. While ETEC is the most common cause for travelers’ diarrhea, with rates in Latin America of 30%, 53% of our samples tested positive.
In Brazil, EPEC is considered to be the cause for 5–10% of all pediatric diarrhea cases [31]. In our study, it was present in 25% of the children and in 42% of the overall samples. EPEC can cause secretory diarrhea with mucus, severe water and electrolyte losses, and even bloody diarrhea. Hu et al. [31] noted that the frequency of EPEC infections decreases with age. The authors suggested the development of immunity or the loss of receptors with specific adhesins as the reason for this result. In contrast to this result, the number of infected individuals in our study was 65% of adults and 23% of adolescents. In addition to the typical EPEC (tEPEC), atypical EPEC (aEPEC) was found in diarrhetic patients of all ages [29]. The aEPEC or highly infectious doses required in adults (108–1010 organisms) [29] could explain the high frequency of infections found in these age groups in our study. Furthermore, colonization needs to be considered [31].
Shigella spp. and EIEC have the same mechanism that causes diarrhea as they share virulence genes and are closely related [32]. In Brazil, the isolation of EIEC ranged from 0.5–15%, and in Bolivia, a prevalence of 2% was detected [29]. In our communities, the prevalence was 18.3%. As there is a high heterogenicity of EIEC isolates, the numbers of EIEC infections might be even higher than described [32]. Like many other pathogens, EIEC needs iron for bacterial colonization, so iron losses can be observed in infected patients [29].
EHEC/STEC belongs to a well-known group of foodborne pathogens distributed worldwide. Their ability to produce Shiga toxins makes them dangerous, and severe complications like Hemolytic Uremic Syndrome can occur [29]. While Gomes et al. [29] noted that the majority of patients are female (57%), with children below 5 years of age (54%) especially affected, we found children between 5–12 years of age (28% of the cases) to be most affected. Overall, we found positive results in 20% of the cases.
C. jejuni was found in 31% of the samples. Campylobacter infections are the main cause for foodborne bacterial illnesses and are even more common than Salmonella-associated foodborne diseases. Animals, especially chickens, as well as the environment, are promoting sources for such infections [33]. Chicken/poultry are the most common animals distributed within Wiwa communities. The consumption of raw milk might also contribute to the high number of infections [34].
In accordance with Young et al. [33], the prevalence for Salmonella spp. infections was lower than that for C. jejuni infections (5% and 31%).
T. whippelei was found in 7%. Since clinical manifestation could not be detected, colonization was assumed. In comparison to other developing countries, this rate is lower. For example, in Ghana, T. whipplei was found in 28% of children aged 2 months to 15 years [35]. Our results support the hypothesis of Vinnemeier et al. [35] that mainly children are colonized during the first years of life.
Thus far, helminth infections were described to be prevalent in Colombia, with rates of 0–9.9% [36]. In Wiwa communities, we found an overall prevalence of 69%. According to Pullan [37], about 1.5 billion people are infected with at least one soil-transmitted helminth [38], comprising 24% of the world´s population [39]. Therefore, STHs belong to the “Big Five” of neglected tropical diseases. The three leading STHs are A. lumbricoides, with 807–1.120 million infections per year; T. trichiura, with 604–795 million; and hookworm infections (A. duodenale and N. americanus), with 576–740 million [40]. These three were also the leading agents in our study, albeit in altered order (T. trichiura, hookworms, A. lumbricoides). The observations made by Moser [38] that A. lumbricoides tends to affect children under 12 years of age more frequently (20%) than older children (14%) and that T. trichiura infections are most common in patients older than 12 years (26%) compared to younger children (14%) could not be confirmed in our results. Infections were distributed equally across all age groups (see Table 2).
Ascaris and hookworm eggs mature in soil. Together with other worms (e.g., whipworms), the fecal–oral route is the most relevant one.
In contrast, hookworms release larvae that penetrate the skin, so the main route of transmission is walking barefoot on contaminated soil [40]. The threadworm is the most neglected tropical disease, although it is an exception among helminths, as it can cause endogenous autoinfections that lead to long lasting infections and systemic features with worm dissemination into various organs [41]. Brazil and Thailand are known to be hotspots for Strongyloides infections; the prevalence in Brazil is about 10–17%, while in Thailand, the overall prevalence is 24% [41]. We found 4% positive results, but these results did not come from multi-center studies using, e.g., Baerman’s method but instead from samples taken once on one day and analyzed with PCR only. In Venezuela, infection rates of 48% were reported in hospitalized patients, while only 3% rates were reported in community-based surveillance [41]. We found 4% positive results, which is high compared to 2%.
H. nana was found in 21% of the Wiwas. In a study performed in the rural zone of Colombia with children (5–15 years), 12% were infected [42]. Similar to our study, all age groups were affected.
The reasons for this high number of infections include bad water quality, low hygiene standards, and a lack of education. The consequences include gastrointestinal infections leading to permanent diarrhea, water and iron loss, electrolyte imbalances, anemia, protein deficits, malnutrition, stunted growth, intellectual impairment, cognitive and educational deficits, and complications in pregnancy (abortion, etc.) as documented (side) effects [2]. The crucial impact can be demonstrated by weight to age, weight to size, and size to age measurements. Children are in the percentile for age at −2/−3 below the average of 0, which is alarming. The average size of Wiwa women is 132.6 cm and 139.4 cm for men, while the normal sizes for Colombians are 158.7 cm for women and 170.6 cm for men. Although permanent and multiple gastrointestinal infections might be an underlying cause, other factors need to be considered, such as ethnic reasons.
The environmental impact becomes obvious when comparing the villages. While in Seminke 100% of the volunteers had at least one positive result, 89% had positive results in Tezhumake and 83% in Valledupar. Helminth infections were three times more common in Seminke than in the other villages. Although all communities have difficult living conditions, Seminke is the most distant from the next city. Furthermore, Seminke is surrounded by forest and Tezhumake by open grass-fields/steppes. Subsequently, the soil in Seminke stays moist longer making helminth transmission more likely.
The high number of co-infections is important to note. A significant association of one infection promoting the other was evident for G. intestinalis and C. jejuni (OR 3.00; 0.95-CI [1.40, 6.43]). Symbiotic interactions were determined for Necator/hookworm and EIEC/Shigella spp., A. lumbricoides, and T. trichiura. S. stercoralis seems to interact with T. trichiura and Necator/hookworm. Although the aforementioned interactions are symbiotic, this is not true for S. stercoralis and EIEC (see Table 2). Here, S. stercoralis supports EIEC unidirectionally. There are many potential reasons for the significant associations between infections. For example, G. intestinalis inducing dysbiosis in the intestinal microbiota might lead to an G. intestinalis expansion itself but might also produce favorable effects for other pathogens. In addition, immune-modulatory effects might contribute to co-infection interactions [43,44].
However, these findings only give tentative evidence due to the unequal sample sizes. Through the application of a propensity-score-approach for the identification of statistical twins [45], the results could be verified. Furthermore, logistic regression depends on varying sample sizes [46]. Subsequently, these findings need to be verified in larger scale studies.
Asymptomatic courses of intestinal infections are common in endemic regions [47], and many diseases have a broad range of clinical symptoms [48], but for such multiple and severe infections, it is difficult to believe that subjects do not experience complaints. Cultural socialization, however, supports the neglect of diseases like diarrhea. As most samples had a fluid or pappy texture, infected individuals might think that this is the normal consistency of stool. Furthermore, access to medical support is far away, with about a 6–12 h walking distance. If we consider (subjective) complaints to be a diagnostic test, the sensitivity would be 10% and the specificity 60%. However, when individuals stated complaints, 3–4 infections were present, with protozoal and/or helminth infections being the leading causes (60% and 58%).
In general, children were more infected than adults; in children, 2–3 infections most commonly occurred. Further, 90% of children below 6 years of age, 74% of the 6–18 year old volunteers, and 39% of the adults (above 18 years of age) suffered from protozoal pathogens. Among the co-infections, combinations of protozoa and helminths were the leading causes (34%), followed by protozoan and bacterial infections (31%) and bacterial and helminth pathogen combinations (25%). Infections with pathogens from all three groups were found in 21% of the cases.
Focusing on the limitations of the study, a further evaluation and classification of pathogens (stool culture, sequencing, etc.), as well as an analysis of water and soil, would be favorable. Due to financial limitations, these additional examinations could not be executed. Since not all inhabitants of the villages provided a stool sample, a selection bias cannot be excluded. However, due to the high compliance of volunteers (80% in Tezhumake and 50% in Seminke) and the results of the anamnesis and examination (feeling healthy and having no complaints), it is likely that no severe sampling error occurred. We also have to state that RT-PCR is not the optimum diagnostic procedure for the detection of E. vermicularis. The low proportion of detected E. vermicularis might be, in part, a consequence of insufficient sensitivity (56%) [3]. Thus, it is likely that infestations with low worm burdens may have gone undetected. From a technical point of view, the superiority of PCR compared to microscopy was confirmed, as microscopy missed infections (compared to PCR) in 40% of the samples [21,22,49,50,51].
The detected prevalence rates are alarming. The interplay of all the above factors accounts for the high burden of gastrointestinal diseases, confirming the observations made by the Wiwas themselves. The negative impact on indigenous communities is enormous. Death due to diarrhea is common, and other related diseases occur frequently (anemia, abortion, etc.). Counteractive interventions, such as drawing attention to public health and education, repetitive deworming programs, improvement of water quality, sanitation supply provisions, and permanent healthcare access are urgently needed.
Author Contributions
Conceptualization, S.K.; Data curation, S.K., M.H. and J.B.; Formal analysis, S.K., J.H., C.L.A.Q., M.H., R.M.H., J.B. and H.F.; Funding acquisition, S.K.; Investigation, S.K., J.H., G.A.C.M., J.J.C.G. and C.L.A.Q.; Methodology, S.K., D.B., J.H., C.L.A.Q., J.B. and H.F.; Project administration, S.K. and G.A.C.M.; Resources, R.M.H., J.B. and H.F.; Software, J.H., M.H. and J.B.; Supervision, S.K., D.B., G.A.C.M., J.J.C.G. and H.F.; Validation, D.B., G.A.C.M., M.H., J.B. and H.F.; Visualization, R.M.H. and J.B.; Writing—original draft, S.K.; Writing—review & editing, D.B., G.A.C.M., R.H., J.B. and H.F. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the European Funds of Regional Development (BWF/H/52228/2012/13. 10.10-1/3.4,6).
Acknowledgments
We gratefully thank Alba Luz-Luque Lommel for her support, Belkis Betin Arias and the health workers of the Dusakawi IPSI for their help in our study, Hernan José Maya Cadavid for providing us laboratory space in the Secretaria de Salud Departamental, and Christine Wegner, Iris Zielke, Doris von Schassen, and Birgit Raschdorff for performing the microscopy.
Conflicts of Interest
There are no conflict of interest.
References
- Hawash, Y.A.; Ismail, K.A.; Almehmadi, M. High Frequency of Enteric Protozoan, Viral, and Bacterial Potential Pathogens in Community-Acquired Acute Diarrheal Episodes: Evidence Based on Results of Luminex Gastrointestinal Pathogen Panel Assay. Korean J. Parasitol. 2017, 55, 513–521. [Google Scholar] [CrossRef] [PubMed]
- WHO. Soil-Transmitted Helminth Infections. Available online: https://www.who.int/news-room/fact-sheets/detail/soil-transmitted-helminth-infections (accessed on 19 June 2020).
- Köller, T.; Hahn, A.; Altangerel, E.; Verweij, J.J.; Landt, O.; Kann, S.; Dekker, D.; May, J.; Loderstädt, U.; Podbielski, A.; et al. Comparison of commercial and in-house real-time PCR platforms for 15 parasites and microsporidia in human stool samples without a gold standard. Acta Trop. 2020, 207, 105516. [Google Scholar] [CrossRef] [PubMed]
- Wiemer, D.; Loderstaedt, U.; von Wulffen, H.; Priesnitz, S.; Fischer, M.; Tannich, E.; Hagen, R.M. Real-time multiplex PCR for simultaneous detection of Campylobacter jejuni, Salmonella, Shigella and Yersinia species in fecal samples. Int. J. Med. Microbiol. 2011, 301, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Luetgehetmann, M.; Landt, O.; Schwarz, N.G.; Frickmann, H. Comparison of one commercial and two in-house TaqMan multiplex real-time PCR assays for detection of enteropathogenic, enterotoxigenic and enteroaggregative Escherichia coli. Trop. Med. Int. Health 2017, 22, 1371–1376. [Google Scholar] [CrossRef]
- Fenollar, F.; Laouira, S.; Lepidi, H.; Rolain, J.M.; Raoult, D. Value of Tropheryma whipplei quantitative polymerase chain reaction assay for the diagnosis of Whipple disease: Usefulness of saliva and stool specimens for first-line screening. Clin. Infect. Dis. 2008, 47, 659–667. [Google Scholar] [CrossRef]
- Frickmann, H.; Hanke, M.; Hahn, A.; Schwarz, N.G.; Landt, O.; Moter, A.; Kikhney, J.; Hinz, R.; Rojak, S.; Dekker, D.; et al. Detection of Tropheryma whipplei in stool samples by one commercial and two in-house real-time PCR assays. Trop. Med. Int. Health 2019, 24, 101–108. [Google Scholar] [CrossRef]
- Niesters, H.G. Quantitation of viral load using real-time amplification techniques. Methods 2001, 25, 419–429. [Google Scholar] [CrossRef]
- Frickmann, H.; Tenner-Racz, K.; Eggert, P.; Schwarz, N.G.; Poppert, S.; Tannich, E.; Hagen, R.M. Influence of parasite density and sample storage time on the reliability of Entamoeba histolytica-specific PCR from formalin-fixed and paraffin-embedded tissues. Diagn. Mol. Pathol. 2013, 22, 236–244. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Sabry, M.A.; Taher, E.S.; Meabed, E.M.H. Prevalence and genotyping of zoonotic Giardia from Fayoum Governate Egypt. Res. J. Parasitol. 2009, 4, 105–114. [Google Scholar] [CrossRef]
- Thompson, R.C.; Hopkins, R.M.; Homan, W.L. Nomenclature and genetic groupings of Giardia infecting mammals. Parasitol. Today 2000, 16, 210–213. [Google Scholar] [CrossRef]
- Foronda, P.; Bargues, M.D.; Abreu-Acosta, N.; Periago, M.V.; Valero, M.A.; Valladares, B.; Mas-Coma, S. Identification of genotypes of Giardia intestinalis of human isolates in Egypt. Parasitol. Res. 2008, 103, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, S.M.; el-Bahy, M.M.; Abd Elaziz, J.; El-Dardiry, M.A. Detection of protozoal parasites in the stools of diarrhoeic patients using different techniques. J. Egypt Soc. Parasitol. 2006, 36, 487–516. [Google Scholar] [PubMed]
- Jensen, L.A.; Marlin, J.W.; Dyck, D.D.; Laubach, H.E. Prevalence of multi-gastrointestinal infections with helminth, protozoan and Campylobacter spp. in Guatemalan children. J. Infect. Dev. Ctries. 2009, 3, 229–234. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lopez, A.S.; Bendik, J.M.; Alliance, J.Y.; Roberts, J.M.; da Silva, A.J.; Moura, I.N.; Arrowood, M.J.; Eberhard, M.L.; Herwaldt, B.L. Epidemiology of Cyclospora cayetanensis and other intestinal parasites in a community in Haiti. J. Clin. Microbiol. 2003, 41, 2047–2054. [Google Scholar] [CrossRef]
- Davies, A.P.; Campbell, B.; Evans, M.R.; Bone, A.; Roche, A.; Chalmers, R.M. Asymptomatic carriage of protozoan parasites in children in day care centers in the United kingdom. Pediatr. Infect. Dis. J. 2009, 28, 838–840. [Google Scholar] [CrossRef]
- Efunshile, M.A.; Ngwu, B.A.; Kurtzhals, J.A.; Sahar, S.; König, B.; Stensvold, C.R. Molecular Detection of the Carriage Rate of Four Intestinal Protozoa with Real-Time Polymerase Chain Reaction: Possible Overdiagnosis of Entamoeba histolytica in Nigeria. Am. J. Trop. Med. Hyg. 2015, 93, 257–262. [Google Scholar] [CrossRef]
- McElligott, J.T.; Naaktgeboren, C.; Makuma-Massa, H.; Summer, A.P.; Deal, J.L. Prevalence of intestinal protozoa in communities along the Lake Victoria region of Uganda. Int. J. Infect. Dis. 2013, 17, e658–e659. [Google Scholar] [CrossRef]
- Utzinger, J.; Botero-Kleiven, S.; Castelli, F.; Chiodini, P.L.; Edwards, H.; Köhler, N.; Gulletta, M.; Lebbad, M.; Manser, M.; Matthys, B.; et al. Microscopic diagnosis of sodium acetate-acetic acid-formalin-fixed stool samples for helminths and intestinal protozoa: A comparison among European reference laboratories. Clin. Microbiol. Infect. 2010, 16, 267–273. [Google Scholar] [CrossRef]
- Nazeer, J.T.; El Sayed Khalifa, K.; von Thien, H.; El-Sibaei, M.M.; Abdel-Hamid, M.Y.; Tawfik, R.A.; Tannich, E. Use of multiplex real-time PCR for detection of common diarrhea causing protozoan parasites in Egypt. Parasitol. Res. 2013, 112, 595–601. [Google Scholar] [CrossRef]
- Krumkamp, R.; Sarpong, N.; Schwarz, N.G.; Adlkofer, J.; Loag, W.; Eibach, D.; Hagen, R.M.; Adu-Sarkodie, Y.; Tannich, E.; May, J. Gastrointestinal infections and diarrheal disease in Ghanaian infants and children: An outpatient case-control study. PLoS Negl. Trop. Dis. 2015, 9, e0003568. [Google Scholar] [CrossRef]
- Rivero, Z.; Bracho, A.; Calchi, M.; Díaz, I.; Acurero, E.; Maldonado, A.; Chourio, G.; Arráiz, N.; Corzo, G. Detection and differentiation of Entamoeba histolytica and Entamoeba dispar by polymerase chain reaction in a community in Zulia State, Venezuela. Cad. Saude Publica 2009, 25, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.; Morán, P.; González, E.; García, G.; Ramiro, M.; Gómez, A.; de León Mdel, C.; Melendro, E.I.; Valadez, A.; Ximénez, C. Entamoeba histolytica and Entamoeba dispar: Prevalence infection in a rural Mexican community. Exp. Parasitol. 2005, 110, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Botero-Garcés, J.; Montoya-Palacio, M.N.; Barguil, J.I.; Castaño-González, A. An outbreak of Cyclospora cayetanensis in Medellín, Colombia. Rev. Salud Publica (Bogota) 2006, 8, 258–268. [Google Scholar] [CrossRef]
- Karanja, R.M.; Gatei, W.; Wamae, N. Cyclosporiasis: An emerging public health concern around the world and in Africa. Afr. Health Sci. 2007, 7, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Frickmann, H.; Schwarz, N.G.; Rakotozandrindrainy, R.; May, J.; Hagen, R.M. PCR for enteric pathogens in high-prevalence settings. What does a positive signal tell us? Infect. Dis. (Lond. Engl.) 2015, 47, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; DuPont, H.L.; Ramsey, D.J. Global etiology of travelers’ diarrhea: Systematic review from 1973 to the present. Am. J. Trop. Med. Hyg. 2009, 80, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.A.; Elias, W.P.; Scaletsky, I.C.; Guth, B.E.; Rodrigues, J.F.; Piazza, R.M.; Ferreira, L.C.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47 (Suppl. 1), 3–30. [Google Scholar] [CrossRef]
- Crofts, A.A.; Giovanetti, S.M.; Rubin, E.J.; Poly, F.M.; Gutiérrez, R.L.; Talaat, K.R.; Porter, C.K.; Riddle, M.S.; DeNearing, B.; Brubaker, J.; et al. Enterotoxigenic E. coli virulence gene regulation in human infections. Proc. Natl. Acad. Sci. USA 2018, 115, E8968–E8976. [Google Scholar] [CrossRef]
- Hu, J.; Torres, A.G. Enteropathogenic Escherichia coli: Foe or innocent bystander? Clin. Microbiol. Infect. 2015, 21, 729–734. [Google Scholar] [CrossRef]
- van den Beld, M.J.C.; Warmelink, E.; Friedrich, A.W.; Reubsaet, F.A.G.; Schipper, M.; de Boer, R.F.; Notermans, D.W.; Petrignani, M.W.F.; van Zanten, E.; Rossen, J.W.A.; et al. Incidence, clinical implications and impact on public health of infections with Shigella spp. and entero-invasive Escherichia coli (EIEC): Results of a multicenter cross-sectional study in the Netherlands during 2016–2017. BMC Infect. Dis. 2019, 19, 1037. [Google Scholar] [CrossRef]
- Young, K.T.; Davis, L.M.; Dirita, V.J. Campylobacter jejuni: Molecular biology and pathogenesis. Nat. Rev. Microbiol. 2007, 5, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Jaakkonen, A.; Kivistö, R.; Aarnio, M.; Kalekivi, J.; Hakkinen, M. Persistent contamination of raw milk by Campylobacter jejuni ST-883. PLoS ONE 2020, 15, e0231810. [Google Scholar] [CrossRef] [PubMed]
- Vinnemeier, C.D.; Klupp, E.M.; Krumkamp, R.; Rolling, T.; Fischer, N.; Owusu-Dabo, E.; Addo, M.M.; Adu-Sarkodie, Y.; Käsmaier, J.; Aepfelbacher, M.; et al. Tropheryma whipplei in children with diarrhoea in rural Ghana. Clin. Microbiol. Infect. 2016, 22, 65.e1–65.e3. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Bethony, J.; Bottazzi, M.E.; Brooker, S.; Buss, P. Hookworm: “The great infection of mankind”. PLoS Med. 2005, 2, e67. [Google Scholar] [CrossRef] [PubMed]
- Pullan, R.L.; Smith, J.L.; Jasrasaria, R.; Brooker, S.J. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasites Vectors 2014, 7, 37. [Google Scholar] [CrossRef]
- Moser, W.; Schindler, C.; Keiser, J. Efficacy of recommended drugs against soil transmitted helminths: Systematic review and network meta-analysis. BMJ 2017, 358, j4307. [Google Scholar] [CrossRef]
- Basuni, M.; Muhi, J.; Othman, N.; Verweij, J.J.; Ahmad, M.; Miswan, N.; Rahumatullah, A.; Aziz, F.A.; Zainudin, N.S.; Noordin, R. A pentaplex real-time polymerase chain reaction assay for detection of four species of soil-transmitted helminths. Am. J. Trop. Med. Hyg. 2011, 84, 338–343. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Parasites—Soil-Transmitted Helminths. Available online: https://www.cdc.gov/parasites/sth/index.html (accessed on 9 June 2020).
- Schär, F.; Trostdorf, U.; Giardina, F.; Khieu, V.; Muth, S.; Marti, H.; Vounatsou, P.; Odermatt, P. Strongyloides stercoralis: Global Distribution and Risk Factors. PLoS Negl. Trop. Dis. 2013, 7, e2288. [Google Scholar] [CrossRef]
- Hernández, P.C.; Morales, L.; Chaparro-Olaya, J.; Sarmiento, D.; Jaramillo, J.F.; Ordoñez, G.A.; Cortés, F.; Sánchez, L.K. Intestinal parasitic infections and associated factors in children of three rural schools in Colombia. A cross-sectional study. PLoS ONE 2019, 14, e0218681. [Google Scholar] [CrossRef]
- Buret, A.G.; Motta, J.P.; Allain, T.; Ferraz, J.; Wallace, J.L. Pathobiont release from dysbiotic gut microbiota biofilms in intestinal inflammatory diseases: A role for iron? J. Biomed. Sci. 2019, 26, 1. [Google Scholar] [CrossRef]
- Hemphill, A.; Müller, N.; Müller, J. Comparative Pathobiology of the Intestinal Protozoan Parasites Giardia lamblia, Entamoeba histolytica, and Cryptosporidium parvum. Pathogens(Basel Switzerland) 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Dettmann, E.; Becker, C.; Schmeißer, C. Distance functions for matching in small samples. Comput. Stat. Data Anal. 2011, 55, 1942–1960. [Google Scholar] [CrossRef]
- Hsieh, F.Y. Sample size tables for logistic regression. Stat. Med. 1989, 8, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Ortega, Y.R.; Sanchez, R. Update on Cyclospora cayetanensis, a food-borne and waterborne parasite. Clin. Microbiol. Rev. 2010, 23, 218–234. [Google Scholar] [CrossRef]
- Halliez, M.C.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. 2013, 19, 8974–8985. [Google Scholar] [CrossRef]
- Verweij, J.J.; Blangé, R.A.; Templeton, K.; Schinkel, J.; Brienen, E.A.; van Rooyen, M.A.; van Lieshout, L.; Polderman, A.M. Simultaneous detection of Entamoeba histolytica, Giardia lamblia, and Cryptosporidium parvum in fecal samples by using multiplex real-time PCR. J. Clin. Microbiol. 2004, 42, 1220–1223. [Google Scholar] [CrossRef]
- Haque, R.; Roy, S.; Siddique, A.; Mondal, U.; Rahman, S.M.; Mondal, D.; Houpt, E.; Petri, W.A., Jr. Multiplex real-time PCR assay for detection of Entamoeba histolytica, Giardia intestinalis, and Cryptosporidium spp. Am. J. Trop. Med. Hyg. 2007, 76, 713–717. [Google Scholar] [CrossRef]
- Stark, D.; Al-Qassab, S.E.; Barratt, J.L.; Stanley, K.; Roberts, T.; Marriott, D.; Harkness, J.; Ellis, J.T. Evaluation of multiplex tandem real-time PCR for detection of Cryptosporidium spp.; Dientamoeba fragilis, Entamoeba histolytica, and Giardia intestinalis in clinical stool samples. J. Clin. Microbiol. 2011, 49, 257–262. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).