The Chemical Composition of Biogas Digestates Determines Their Effect on Soil Microbial Activity


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiment
2.2. Dehydrogenase Activity (DHA)
2.3. Soil Respiration
2.4. Statistical Analysis
3. Results and Discussion
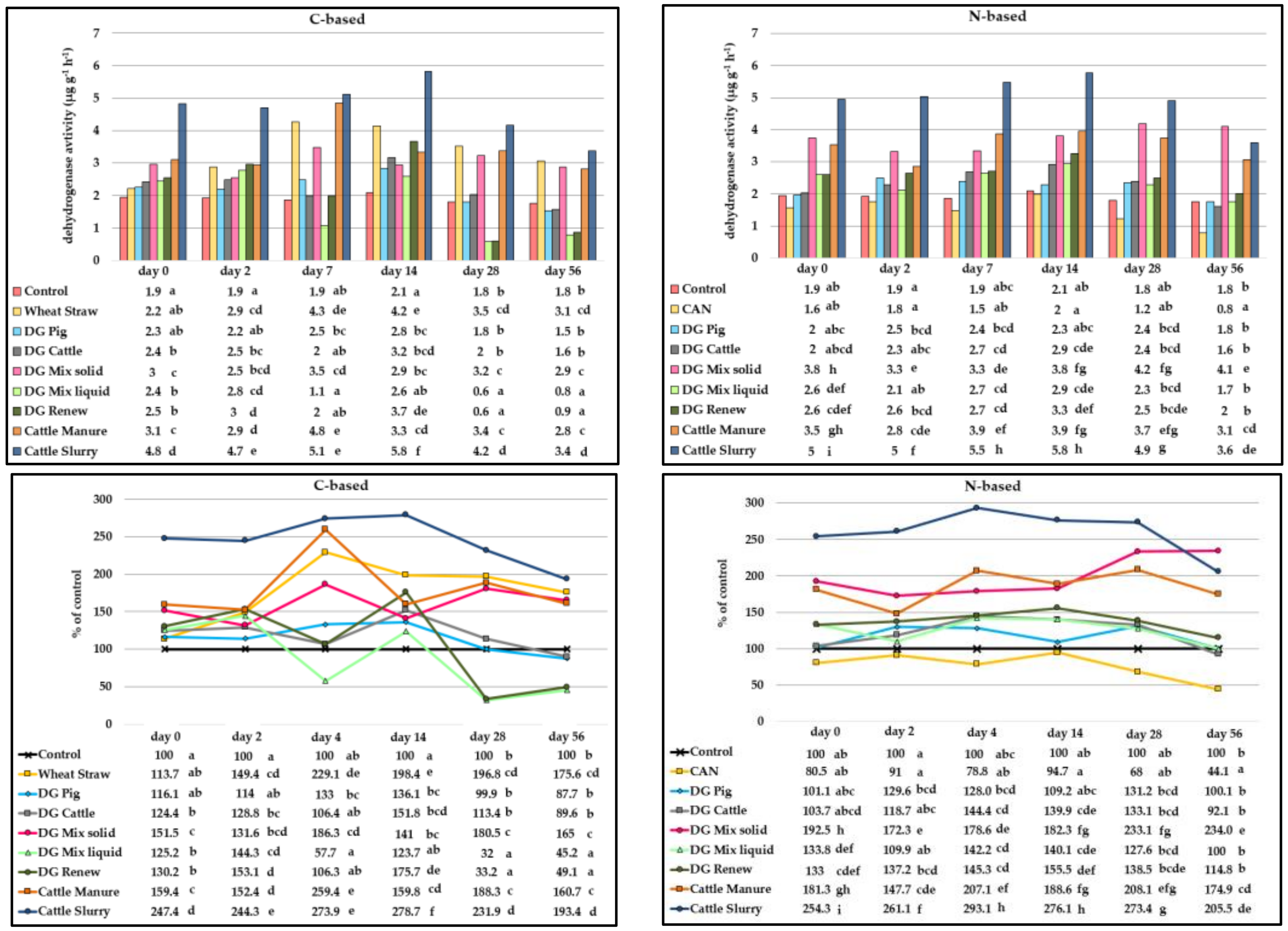
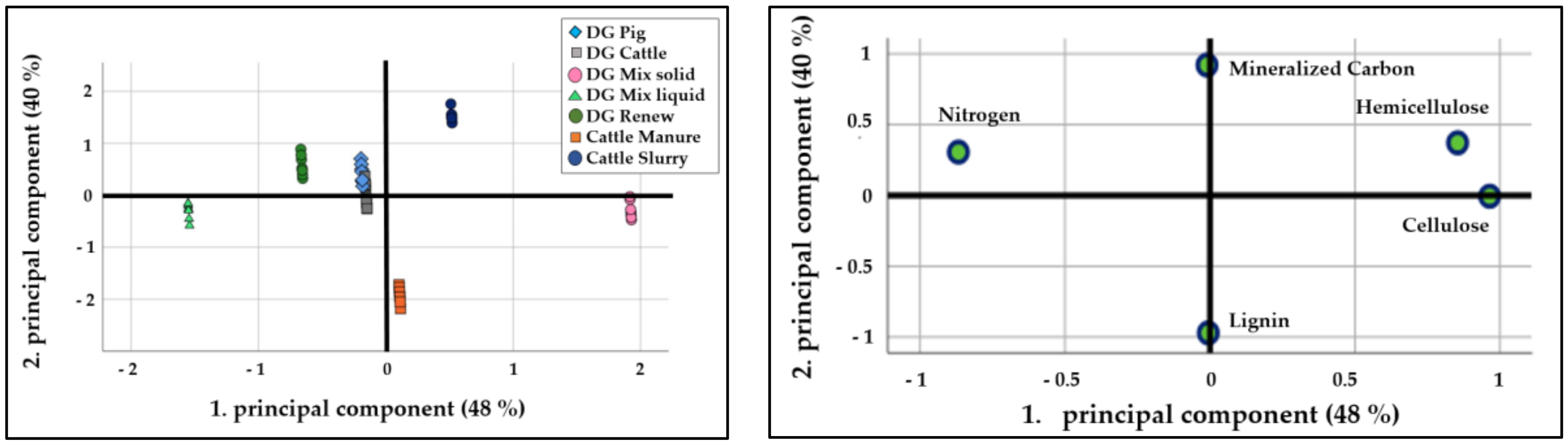
3.1. Dehydrogenase Activity
3.2. Soil Respiration
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Appels, L.; Lauwers, J.; Degrève, J.; Helsen, L.; Lievens, B.; Willems, K.; Van Impe, J.; Dewil, R. Anaerobic digestion in global bio-energy production: Potential and research challenges. Renew. Sustain. Energy Rev. 2011, 15, 4295–4301. [Google Scholar] [CrossRef]
- Tambone, F.; Scaglia, B.; D’Imporzano, G.; Schievano, A.; Orzi, V.; Salati, S.; Adani, F. Assessing amendment and fertilizing properties of digestates from anaerobic digestion through a comparative study with digested sludge and compost. Chemosphere 2010, 81, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Möller, K.; Müller, T. Effects of anaerobic digestion on digestate nutrient availability and crop growth: A review. Eng. Life Sci. 2012, 12, 242–257. [Google Scholar] [CrossRef]
- Maroušek, J.; Bartoš, P.; Filip, M.; Kolář, L.; Konvalina, P.; Maroušková, A.; Moudrý, J.; Peterka, J.; Šál, J.; Šoch, M.; et al. Advances in the agrochemical utilization of fermentation residues reduce the cost of purpose-grown phytomass for biogas production. Energy Sour. Part A Recovery Util. Environ. Eff. 2020, 1–11. [Google Scholar] [CrossRef]
- Kolář, L.; Kužel, S.; Peterka, J.; Borová-Batt, J. Utilisation of Waste from Digesters for Biogas Production. In Biofuel’s Engineering Process Technology; Bernardes, M.A.d.S., Ed.; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Stehel, V.; Horák, J.; Vochozka, M. Prediction of institutional sector development and analysis of enterprises active in agriculture. E+M Ekon. A Manag. 2019, 22, 103–118. [Google Scholar] [CrossRef]
- Abubaker, J.; Risberg, K.; Pell, M. Biogas residues as fertilisers—Effects on wheat growth and soil microbial activities. Appl. Energy 2012, 99, 126–134. [Google Scholar] [CrossRef]
- Odlare, M.; Pell, M.; Svensson, K. Changes in soil chemical and microbiological properties during 4 years of application of various organic residues. Waste Manag. 2008, 28, 1246–1253. [Google Scholar] [CrossRef]
- Abubaker, J.; Cederlund, H.; Arthurson, V.; Pell, M. Bacterial community structure and microbial activity in different soils amended with biogas residues and cattle slurry. Appl. Soil Ecol. 2013, 72, 171–180. [Google Scholar] [CrossRef]
- Hupfauf, S.; Bachmann, S.; Fernández-Delgado Juárez, M.; Insam, H.; Eichler-Löbermann, B. Biogas digestates affect crop P uptake and soil microbial community composition. Sci. Total Environ. 2016, 542, 1144–1154. [Google Scholar] [CrossRef]
- García-Sánchez, e.; Siles, J.A.; Cajthaml, T.; García-Romera, I.; Tlustoš, P.; Száková, J. Effect of digestate and fly ash applications on soil functional properties and microbial communities. Eur. J. Soil Biol. 2015, 71, 1–12. [Google Scholar] [CrossRef]
- Garcia-Sanchez, M.; Garcia-Romera, I.; Cajthaml, T.; Tlustos, P.; Szakova, J. Changes in soil microbial community functionality and structure in a metal-polluted site: The effect of digestate and fly ash applications. J. Env. Manag. 2015, 162, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Abubaker, J.; Risberg, K.; Jönsson, E.; Dahlin, A.S.; Cederlund, H.; Pell, M. Short-Term Effects of Biogas Digestates and Pig Slurry Application on Soil Microbial Activity. Appl. Environ. Soil Sci. 2015, 2015, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Risberg, K.; Cederlund, H.; Pell, M.; Arthurson, V.; Schnurer, A. Comparative characterization of digestate versus pig slurry and cow manure—Chemical composition and effects on soil microbial activity. Waste Manag. 2017, 61, 529–538. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, C.; Alburquerque, J.A.; Clemente, R.; Bernal, M.P. Soil C and N mineralisation and agricultural value of the products of an anaerobic digestion system. Biol. Fertil. Soils 2012, 49, 313–322. [Google Scholar] [CrossRef]
- Johansen, A.; Carter, M.S.; Jensen, E.S.; Hauggard-Nielsen, H.; Ambus, P. Effects of digestate from anaerobically digested cattle slurry and plant materials on soil microbial community and emission of CO2 and N2O. Appl. Soil Ecol. 2013, 63, 36–44. [Google Scholar] [CrossRef]
- Korschens, M.; Weigel, A.; Schulz, E. Turnover of soil organic matter (SOM) and long-term balances—Tools for evaluating sustainable productivity of soils. Z. Fuer Pflanz. Und Bodenkd. (Ger.) 1998, 161, 409–424. [Google Scholar] [CrossRef]
- Prays, N.; Dominik, P.; Sanger, A.; Franko, U. Biogas residue parameterization for soil organic matter modeling. PLoS ONE 2018, 13, e0204121. [Google Scholar] [CrossRef] [Green Version]
- Maucieri, C.; Barbera, A.C.; Borin, M. Effect of injection depth of digestate liquid fraction on soil carbon dioxide emission and maize biomass production. Ital. J. Agron. 2016, 10, 6–11. [Google Scholar] [CrossRef]
- Makádi, M.; Tomócsik, A.; Orosz, V. Digestate: A New Nutrient Source—Review. In Biogas; Kumar, S., Ed.; IntechOpen: London, UK, 2012; pp. 295–310. [Google Scholar]
- Cavalli, D.; Corti, M.; Baronchelli, D.; Bechini, L.; Marino Gallina, P. CO2 emissions and mineral nitrogen dynamics following application to soil of undigested liquid cattle manure and digestates. Geoderma 2017, 308, 26–35. [Google Scholar] [CrossRef]
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2013, 34, 473–492. [Google Scholar] [CrossRef] [Green Version]
- Alburquerque, J.A.; de la Fuente, C.; Bernal, M.P. Chemical properties of anaerobic digestates affecting C and N dynamics in amended soils. Agric. Ecosyst. Environ. 2012, 160, 15–22. [Google Scholar] [CrossRef]
- Kumar, S.; Chaudhuri, S.; Maiti, S.K. Soil Dehydrogenase Enzyme Activity in Natural and Mine Soil—A Review. Middle East. J. Sci. Res. 2013, 13, 898–906. [Google Scholar]
- Wolinska, A.; Stepniewska, Z. Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; Canuto, R.A., Ed.; IntechOpen: London, UK, 2012; pp. 183–210. [Google Scholar]
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant. Anal. 2008, 28, 123–134. [Google Scholar] [CrossRef]
- Huang, P.M.; Hardie, A.G. Formation Mechanisms of Humic Substances in the Environment. In Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental Systems; Senesi, N., Xing, B., Huang, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 41–109. [Google Scholar] [CrossRef]
- Ottow, J.C.G. Mikrobiologie von Böden—Biodiversität, Ökophysiologie und Metagenomik; Springer: Berlin, Germany, 2011. [Google Scholar] [CrossRef]
- Trumbore, S.E. Potential responses of soil organic carbon to global environmental change. Proc. Natl. Acad. Sci. USA 1997, 94, 8284–8291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume, H.-P.; Brümmer, G.W.; Horn, R.; Kandeler, E.; Kögel-Knabner, I.; Kretzschmar, R.; Stahr, K.; Wilke, B.-M. Scheffer/Schachtschabel Soil Science, 1st ed.; Springer: Berlin, Germany, 2015. [Google Scholar]
- Bermejo Domínguez, G. Agro-Ecological Aspects when Applying the Remaining Products from Agricultural Biogas Processes as Fertilizer in Crop Production. Ph.D. Thesis, Humboldt-Universität zu Berlin, Berlin, Germany, August 2012. [Google Scholar]
- Fangueiro, D.; Gusmão, M.; Grilo, J.; Porfírio, G.; Vasconcelos, E.; Cabral, F. Proportion, composition and potential N mineralisation of particle size fractions obtained by mechanical separation of animal slurry. Biosyst. Eng. 2010, 106, 333–337. [Google Scholar] [CrossRef]
- Bahri, H.; Rasse, D.P.; Rumpel, C.; Dignac, M.F.; Bardoux, G.; Mariotti, A. Lignin degradation during a laboratory incubation followed by 13C isotope analysis. Soil Biol. Biochem. 2008, 40, 1916–1922. [Google Scholar] [CrossRef]
- Kirk, T.K. Enzymatic “Combustion”: The Microbial Degradation of Lignin. Annu. Rev. Microbiol. 1987, 41, 465–505. [Google Scholar] [CrossRef]
- Chen, R.; Blagodatskaya, E.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Kuzyakov, Y. Decomposition of biogas residues in soil and their effects on microbial growth kinetics and enzyme activities. Biomass Bioenergy 2012, 45, 221–229. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Arshad, M.A. Particulate Organic Carbon Content and Potential Mineralization as Affected by Tillage and Texture. Soil Sci. Soc. Am. J. 1997, 61, 1382–1386. [Google Scholar] [CrossRef]
- Schulz, E. Einfluss von Bodenart und langjährig differenzierter düngung auf die Akkumulation und Stabilisierung Organischer Bodensubstanz (OBS) in unterschiedlichen Bodenfraktionen. Arch. Agron. Soil Sci. 2000, 45, 365–380. [Google Scholar] [CrossRef]
- Rigby, H.; Smith, S.R. Nitrogen availability and indirect measurements of greenhouse gas emissions from aerobic and anaerobic biowaste digestates applied to agricultural soils. Waste Manag. 2013, 33, 2641–2652. [Google Scholar] [CrossRef] [PubMed]
- DIN EN ISO 23753-1:2011. Bodenbeschaffenheit—Bestimmung der Dehydrogenaseaktivität in Böden—Teil 1: Verfahren mit Triphenyltetrazoliumchlorid (TTC); Beuth-Verlag: Berlin, Germany, 2011. [CrossRef]
- DIN ISO 10381-1:2003-08 Bodenbeschaffenheit—Probenahme—Teil 1: Anleitung zur Aufstellung von Probenahmeprogrammen (ISO 10381-1:2002); Beuth-Verlag: Berlin, Germany, 2003. [CrossRef]
- DIN EN ISO 16072:2011. German Title: Bodenbeschaffenheit—Laborverfahren zur Bestimmung der Mikrobiellen Bodenatmung (ISO 16072:2002); Beuth Verlag: Berlin, Germany, 2011. [CrossRef]
- Chapman, S.B. A Simple Conductimetric Soil Respirometer for Field Use. OIKOS 1971, 22, 348–353. [Google Scholar] [CrossRef]
- Makádi, M.; Tomócsik, A.; Orosz, V.; Bogdányi, Z.; Biró, B. Effect of a biogas-digestate and bentonite on some enzyme activities of the amended soils. Cereal Res. Commun. 2007, 35, 741–744. [Google Scholar] [CrossRef]
- Islam, M.R.; Singh Chauhan, P.; Kim, Y.; Kim, M.; Sa, T. Community level functional diversity and enzyme activities in paddy soils under different long-term fertilizer management practices. Biol. Fertil. Soils 2010, 47, 599–604. [Google Scholar] [CrossRef]
- Saha, S.; Prakash, V.; Kundu, S.; Kumar, N.; Lal Mina, B. Soil enzymatic activity as affected by long term application of farm yard manure and mineral fertilizer under a rainfed soybean–wheat system in N-W Himalaya. Eur. J. Soil Biol. 2008, 44, 309–315. [Google Scholar] [CrossRef]
- Trevors, J.T. Effect of substrate concentration, inorganic nitrogen, O2 concentration, temperature and pH on dehydrogenase activity in soil. Plant. Soil 1984, 77, 285–293. [Google Scholar] [CrossRef]
- Goyal, S.; Chander, K.; Mundra, M.C.; Kapoor, K.K. Influence of inorganic fertilizers and organic amendments on soil organic matter and soil microbial properties under tropical conditions. Biol. Fertil. Soils 1999, 29, 196–200. [Google Scholar] [CrossRef]
- Dhull, S.; Goyal, S.; Kapoor, K.; Mundra, M. Microbial biomass carbon and microbial activities of soils receiving chemical fertilizers and organic amendments. Arch. Agron. Soil Sci. 2004, 50, 641–647. [Google Scholar] [CrossRef]
- Chu, H.; Lin, X.; Fujii, T.; Morimoto, S.; Yagi, K.; Hu, J.; Zhang, J. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 2007, 39, 2971–2976. [Google Scholar] [CrossRef]
- Kautz, T.; Wirth, S.; Ellmer, F. Microbial activity in a sandy arable soil is governed by the fertilization regime. Eur. J. Soil Biol. 2004, 40, 87–94. [Google Scholar] [CrossRef]
- Rao, D.; Ghai, S. Urease und dehydrogenase activity of alkali and reclaimed soils. Aust. J. Soil Res. 1985, 23, 661–665. [Google Scholar] [CrossRef]
- Liu, L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Grabber, J.H. How Do Lignin Composition, Structure, and Cross-Linking Affect Degradability? A Review of Cell Wall Model Studies. Crop. Sci. 2005, 45, 820–831. [Google Scholar] [CrossRef] [Green Version]
- Vikman, M.; Karjomaa, S.; Kapanen, A.; Wallenius, K.; Itavaara, M. The influence of lignin content and temperature on the biodegradation of lignocellulose in composting conditions. Appl. Microbiol. Biotechnol. 2002, 59, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Austin, A.T.; Ballare, C.L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2010, 107, 4618–4622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annabi, M.; Houot, S.; Francou, C.; Poitrenaud, M.; Bissonnais, Y.L. Soil Aggregate Stability Improvement with Urban Composts of Different Maturities. Soil Sci. Soc. Am. J. 2007, 71, 413–423. [Google Scholar] [CrossRef]
- Ernst, G.; Müller, A.; Göhler, H.; Emmerling, C. C and N turnover of fermented residues from biogas plants in soil in the presence of three different earthworm species (Lumbricus terrestris, Aporrectodea longa, Aporrectodea caliginosa). Soil Biol. Biochem. 2008, 40, 1413–1420. [Google Scholar] [CrossRef]
- Wentzel, S.; Joergensen, R.G. Effects of biogas and raw slurries on grass growth and soil microbial indices. J. Plant. Nutr. Soil Sci. 2016, 179, 215–222. [Google Scholar] [CrossRef]
- Madritch, M.D.; Hunter, M.D. Intraspecific litter diversity and nitrogen deposition affect nutrient dynamics and soil respiration. Oecologia 2003, 136, 124–128. [Google Scholar] [CrossRef]
- Tewary, C.K.; Pandey, U.; Singh, J.S. Soil and litter respiration rates in different microhabitats of a mixed oak-conifer forest and their control by edaphic conditions and substrate quality. Plant. Soil 1982, 65, 233–238. [Google Scholar] [CrossRef]
- Yanni, S.F.; Whalen, J.K.; Simpson, M.J.; Janzen, H.H. Plant lignin and nitrogen contents control carbon dioxide production and nitrogen mineralization in soils incubated with Bt and non-Bt corn residues. Soil Biol. Biochem. 2011, 43, 63–69. [Google Scholar] [CrossRef]
- Stewart, C.E.; Moturi, P.; Follett, R.F.; Halvorson, A.D. Lignin biochemistry and soil N determine crop residue decomposition and soil priming. Biogeochemistry 2015, 124, 335–351. [Google Scholar] [CrossRef] [Green Version]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Berg, B.; Matzner, E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environ. Rev. 1997, 5, 1–25. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Sinsabaugh, R.L.; Repert, D.A.; Parkhurst, D.F. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 2000, 81, 2359–2365. [Google Scholar] [CrossRef]
- Frey, S.D.; Knorr, M.; Parrent, J.L.; Simpson, R.T. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. For. Ecol. Manag. 2004, 196, 159–171. [Google Scholar] [CrossRef]
- Agren, G.I.; Bosatta, E.; Magill, A.H. Combining theory and experiment to understand effects of inorganic nitrogen on litter decomposition. Oecologia 2001, 128, 94–98. [Google Scholar] [CrossRef]
- Arthurson, V. Closing the Global Energy and Nutrient Cycles through Application of Biogas Residue to Agricultural Land—Potential Benefits and Drawback. Energies 2009, 2, 226–242. [Google Scholar] [CrossRef] [Green Version]
- Marcato-Romain, C.-E.; Mohtar, R.; Revel, J.-C.; Pouech, P.; Hafidi, M.; Guiresse, M. Impact of anaerobic digestion on organic matter quality in pig slurry. Int. Biodeterior. Biodegrad. 2009, 63, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Wiesler, F.; Hund-Rinke, K.; Gäth, S.; George, E.; Greef, J.M.; Hölzle, L.E.; Holz, F.; Hülsbergen, K.-J.; Pfeil, R.; Severin, K.; et al. Application of Organic Fertilisers and Organic Residual Materials in Agriculture; Federal Ministry of Food and Agriculture: Bonn, Germany, 2015. [Google Scholar]
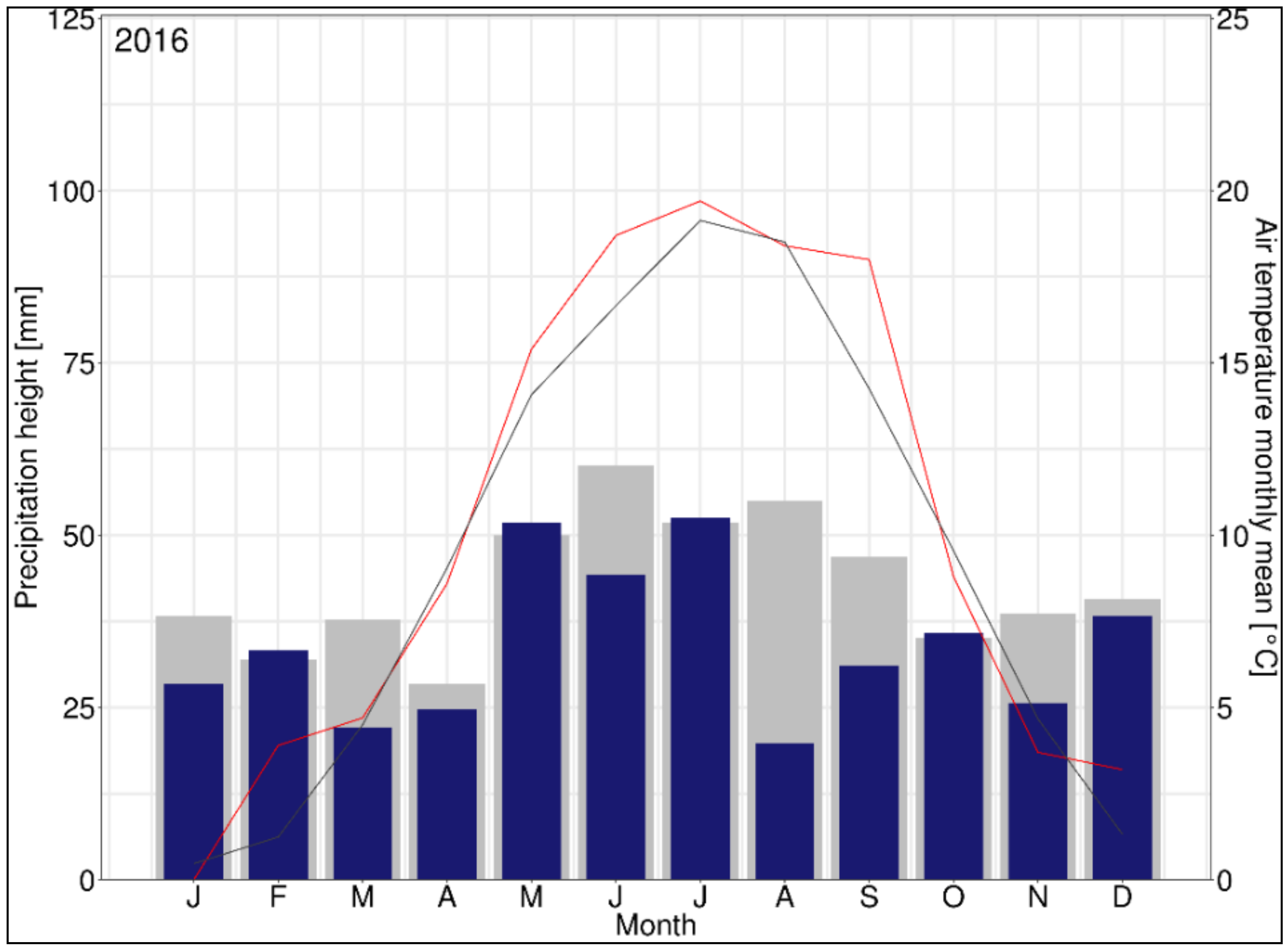







{kind=link}
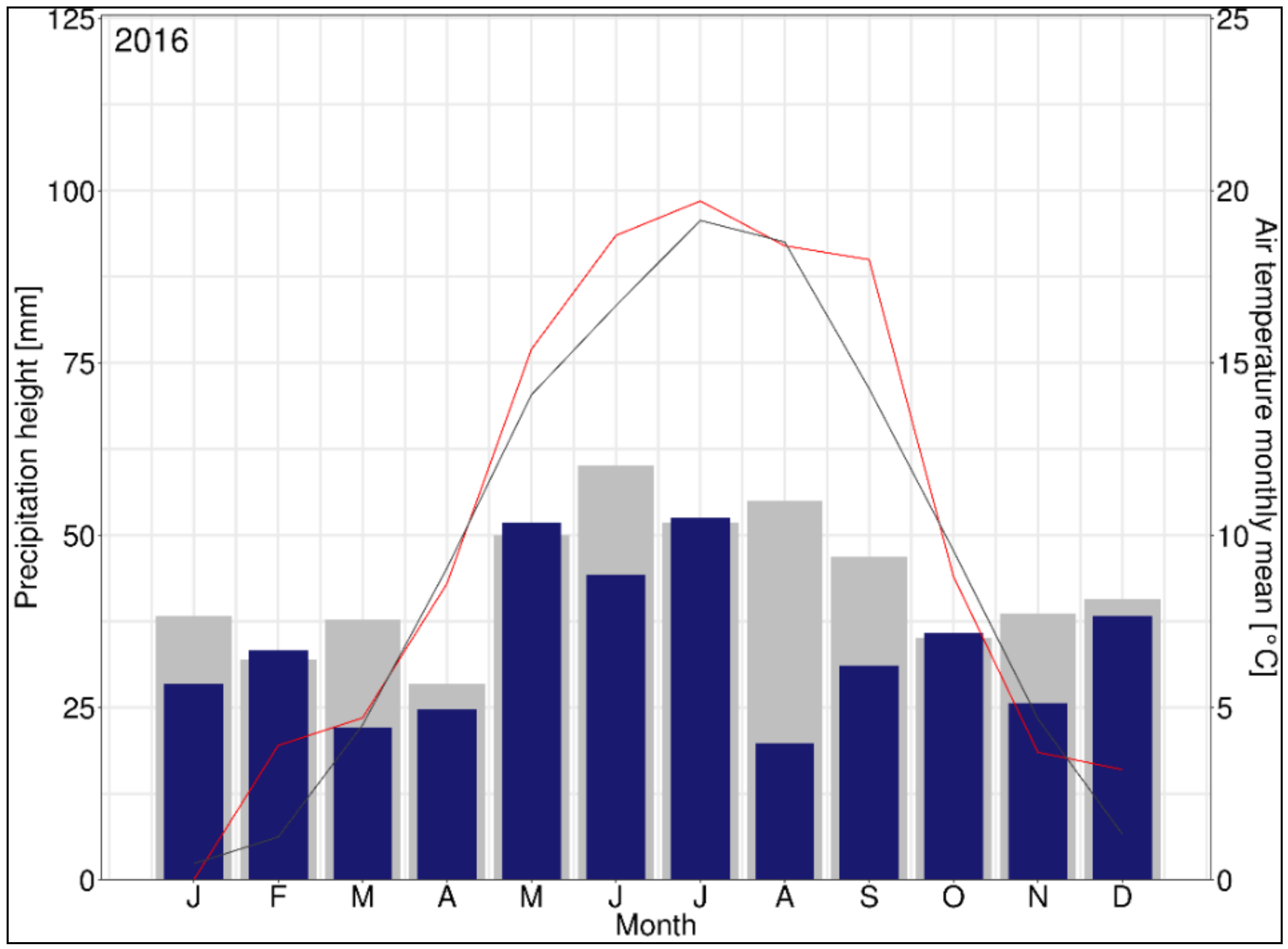
{kind=link}
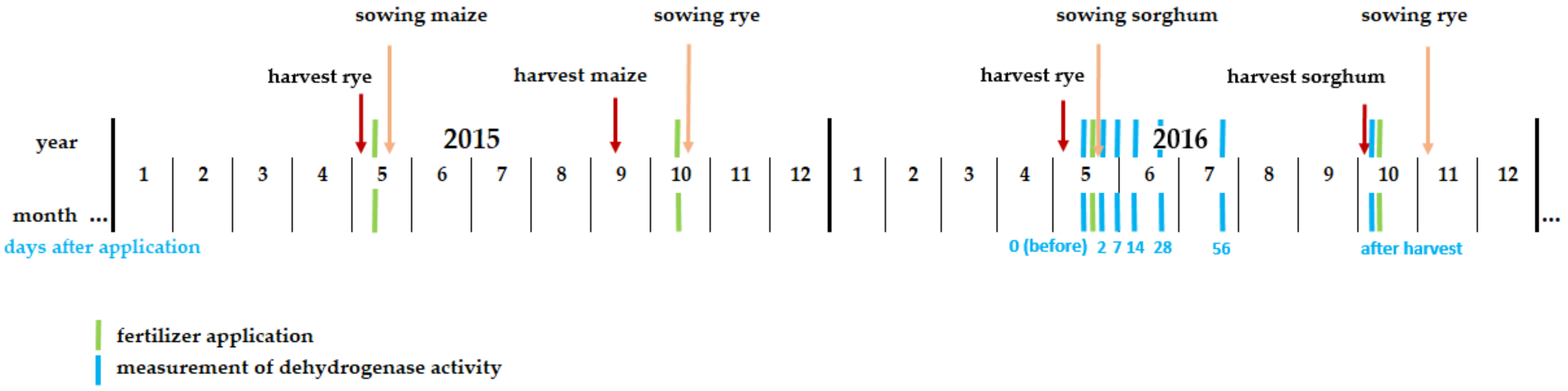
{kind=link}
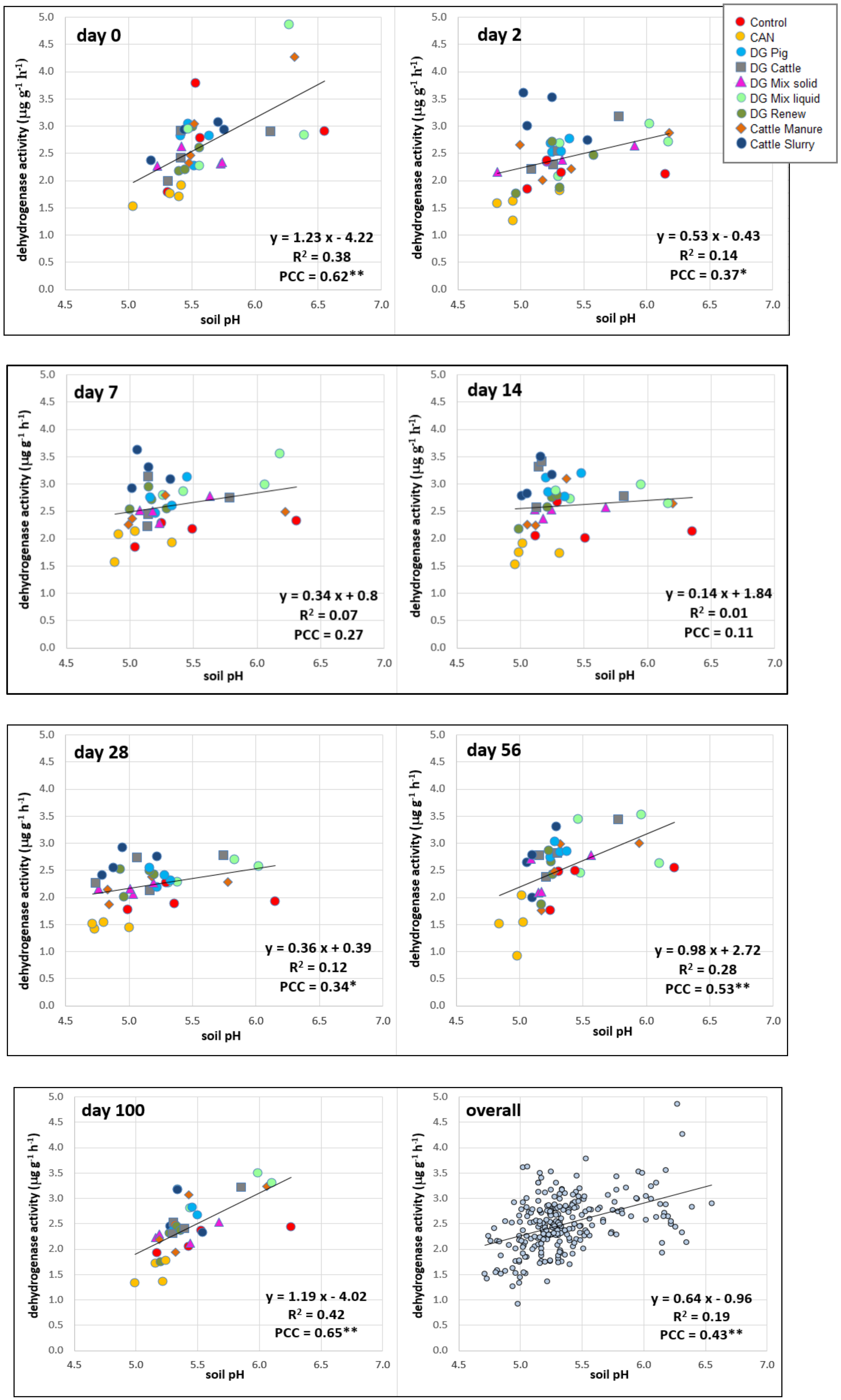{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | A | B | C | D | ||||
|---|---|---|---|---|---|---|---|---|
| Average Input | 49% | cattle slurry | 43% | pig slurry | 86% 14% | corn silage rye and rye silage | 31% | cattle slurry |
| 24% | grass silage | |||||||
| 34% | corn silage | 46% | corn silage | 20% | corn silage | |||
| 11% | grass silage | 6% | millet silage | 16% | cattle manure | |||
| 3% | triticale silage | 4% | grass silage | 9% | rye silage | |||
| 3% | fodder remains | 1% | fodder remains | |||||
| Operating temperature | mesophile | mesophile | thermophile | mesophile | ||||
| Retention time | 70 days | 60 days | 50 days | 80 days | ||||
| Related products | “DG cattle” and “Cattle Slurry” | “DG Pig” | “DG Renew” | “DG Mix liquid” and “DG Mix solid” | ||||
| Applied Amount of | Application Date | DG Cattle | DG Pig | DG Renew | DG Mix Solid | DG Mix Liquid | Cattle Slurry | Cattle Manure |
|---|---|---|---|---|---|---|---|---|
| Organic fertilizer (t ha−1 FM) | Average Spring | 24.9 | 37.6 | 15.7 | 6.8 | 26.8 | 19.3 | 7.5 |
| Average Autumn | 17.2 | 23.8 | 13 | 4.9 | 23.9 | 15 | 5 | |
| 10/2015 (to rye) | 14.3 | 15.1 | 12.5 | 4.2 | 20.2 | 13.3 | 5 | |
| 5/2016 (to sorghum) | 28.8 | 23.9 | 15.3 | 6.8 | 32.5 | 23.6 | 7.5 | |
| Corg (t ha−1) | Average Spring | 0.6; same amount for all | ||||||
| Average Autumn | 0.5; same amount for all | |||||||
| 10/2015 (to rye) | 0.4; same amount for all | |||||||
| 5/2016 (to sorghum) | 0.7; same amount for all | |||||||
| N from organic fertilization (kg ha−1) | Average Spring | 109 | 117.9 | 90.6 | 36.8 | 118.1 | 62.8 | 48.7 |
| Average Autumn | 72.8 | 79 | 75.4 | 26.8 | 108.9 | 46.4 | 40.5 | |
| 10/2015 (to rye) | 57.6 | 61.2 | 68 | 22.2 | 68.4 | 37.8 | 30.8 | |
| 5/2016 (to sorghum) | 124.5 | 108.0 | 88.4 | 36.6 | 124 | 68.4 | 62.3 | |
| N from balancing mineral fertilization (kg ha−1) | Average Spring | 21.5 | 12.6 | 39.9 | 94 | 4.1 | 67.7 | 81.9 |
| Average Autumn | 36.1 | 29.9 | 33.6 | 82.2 | 0 | 62.5 | 68.5 | |
| 10/2015 (to rye) | 10.8 | 7.2 | 0.4 | 46.2 | 0 | 30.6 | 37.6 | |
| 5/2016 (to sorghum) | 0 | 16.4 | 36.1 | 87.9 | 0.5 | 56 | 62.2 | |
| Ntotal (kg ha−1) | Average Spring | 130.5; same amount for all | ||||||
| Average Autumn | 109; same amount for all | |||||||
| 10/2015 (to rye) | 68.4; same amount for all | |||||||
| 5/2016 (to sorghum) | 124.5; same amount for all | |||||||
| K (kg ha−1) | Average Spring | 74.7 | 71.7 | 78.3 | 29.3 | 104.5 | 47 | 46.4 |
| Average Autumn | 42.9 | 59 | 52.9 | 31.1 | 81.3 | 45.3 | 43.1 | |
| 10/2015 (to rye) | 44 | 44.2 | 74.6 | 14.9 | 74 | 28.3 | 23.1 | |
| 5/2016 (to sorghum) | 107.2 | 81 | 82.3 | 31.8 | 118 | 58 | 56.1 | |
| P (kg ha−1) | Average Spring | 17.8 | 18.0 | 15.2 | 10.7 | 19.2 | 12.3 | 9.5 |
| Average Autumn | 11.2 | 17.6 | 9.8 | 8.2 | 12.7 | 10.9 | 12.3 | |
| 10/2015 (to rye) | 11.7 | 16.3 | 9.7 | 5.2 | 12.9 | 9.6 | 7.4 | |
| 5/2016 (to sorghum) | 22.4 | 36.0 | 17.8 | 12.6 | 21.0 | 15.2 | 12.0 | |
| Parameter | Unit | DG Cattle | DG Pig | DG Renew | DG Mix Solid | DG Mix Liquid | Cattle Slurry | Cattle Manure | CAN | Straw | Glucose |
|---|---|---|---|---|---|---|---|---|---|---|---|
| DM | % | 6.7 | 6.2 | 6.9 | 21.2 | 5.2 | 6.3 | 26.3 | 100 | 89.8 | 98.0 |
| Ntotal | % DM | 6 | 6.6 | 7.8 | 2.5 | 6.5 | 4.5 | 2.3 | 27 | 0.5 | n.d. |
| Corg | % DM | 38.2 | 39.5 | 42.2 | 41.6 | 34.9 | 44 | 28.1 | 0 | 51.9 | 42.1 |
| C:N-Ratio | 6:1 | 6:1 | 5:1 | 17:1 | 5:1 | 10:1 | 12:1 | - | 103:1 | n.d. | |
| P | g kg−1 DM | 12.2 | 17.4 | 11.1 | 5.9 | 12.2 | 11.5 | 5.6 | 0 | 1.2 | n.d. |
| K | g kg−1 DM | 45.7 | 47.4 | 85.9 | 16.8 | 70.1 | 33.9 | 17.6 | 0 | 10.6 | n.d. |
| Mg | g kg−1 DM | 8.5 | 9.4 | 3.2 | 4.7 | 11 | 8.9 | 4.3 | 0 | 0.7 | n.d. |
| Na | g kg−1 DM | 6.7 | 5.9 | 3.5 | 2.5 | 8.7 | 9.4 | 2.4 | 0 | 0.3 | n.d. |
| Cellulose | % DM | 11 | 9.9 | 12.3 | 23.9 | 4.1 | 14.6 | 8.4 | 0 | 43.8 | n.d. |
| Hemicellulose | % DM | 10 | 12.9 | 6 | 18.2 | 0 | 14.1 | 3.2 | 0 | 39.9 | n.d. |
| Lignin | % DM | 16.7 | 15.7 | 18.7 | 21.1 | 17 | 9.9 | 21.4 | 0 | 6.3 | n.d. |
| pH | 7.8 | 7.7 | 8.1 | 9 | 7.6 | 6.5 | 7.9 | n.d. | 7.1 | n.d. |
| Soil | Sand (%) | Silt (%) | Clay (%) | Corg (%) | Ntotal (%) | pH |
|---|---|---|---|---|---|---|
| Berge | 75.6 | 16.6 | 7.8 | 0.7 | 0.06 | 5.8 |
| Thyrow | 83.1 | 14.2 | 2.7 | 0.4 | 0.05 | 5.4 |
| Substrate | DG Cattle | DG Pig | DG Renew | DG Mix Solid | DG Mix Liquid | Cattle Slurry | Cattle Manure | Wheat Straw | Glucose |
|---|---|---|---|---|---|---|---|---|---|
| Berge | 20 b | 25 b | 44 c | 29 b | 25 b | 51 c | 11 a | 72 d | 95 e |
| Thyrow | 31 abc | 34 bcd | 40 cd | 24 ab | 29 abc | 49 de | 16 a | 61 f | 88 e |
| Component | Pearson’s Correlation Coefficient | Significance | ||||
|---|---|---|---|---|---|---|
| Overall | Berge | Thyrow | Overall | Berge | Thyrow | |
| N | 0.21 | 0.11 | 0.34 | - | - | - |
| Cellulose | 0.15 | 0.13 | −0.14 | - | - | - |
| Hemicellulose | 0.22 | 0.24 | −0.18 | - | - | - |
| Lignin | −0.82 | −0.78 | −0.90 | <0.01 | <0.01 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, K.; Roß, C.-L.; Hoffmann, M.; Muskolus, A.; Ellmer, F.; Kautz, T. The Chemical Composition of Biogas Digestates Determines Their Effect on Soil Microbial Activity. Agriculture 2020, 10, 244. https://doi.org/10.3390/agriculture10060244
Nielsen K, Roß C-L, Hoffmann M, Muskolus A, Ellmer F, Kautz T. The Chemical Composition of Biogas Digestates Determines Their Effect on Soil Microbial Activity. Agriculture. 2020; 10(6):244. https://doi.org/10.3390/agriculture10060244
Chicago/Turabian StyleNielsen, Kerstin, Christina-Luise Roß, Marieke Hoffmann, Andreas Muskolus, Frank Ellmer, and Timo Kautz. 2020. "The Chemical Composition of Biogas Digestates Determines Their Effect on Soil Microbial Activity" Agriculture 10, no. 6: 244. https://doi.org/10.3390/agriculture10060244
APA StyleNielsen, K., Roß, C.-L., Hoffmann, M., Muskolus, A., Ellmer, F., & Kautz, T. (2020). The Chemical Composition of Biogas Digestates Determines Their Effect on Soil Microbial Activity. Agriculture, 10(6), 244. https://doi.org/10.3390/agriculture10060244