Change in Maize Final Leaf Numbers and Its Effects on Biomass and Grain Yield across China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
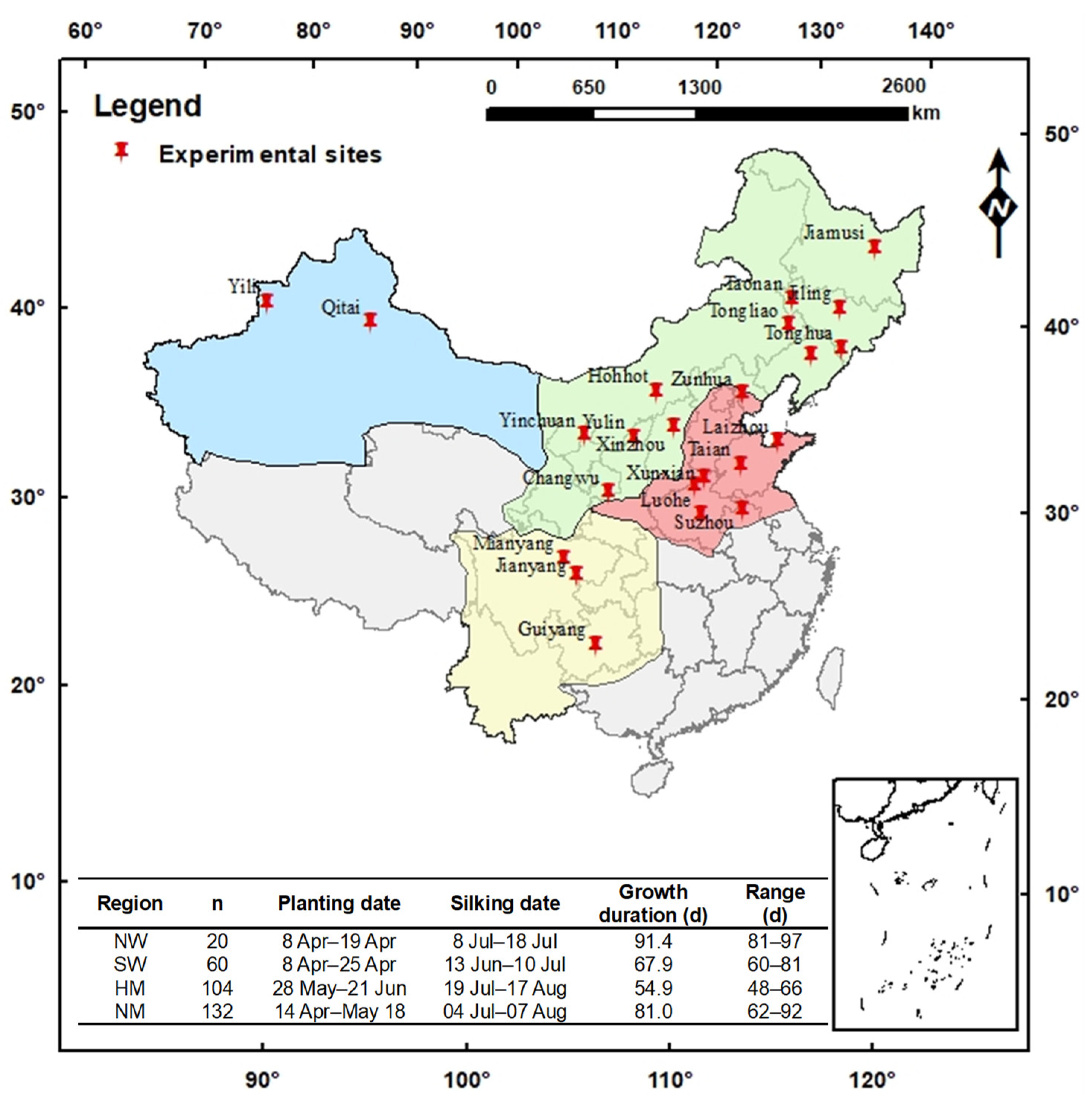
2.1. Site Description and Experimental Design
2.2. Sampling and Measurements
2.3. Statistical Analysis
3. Results
3.1. Difference in Final Leaf Number between Cultivars across All Locations
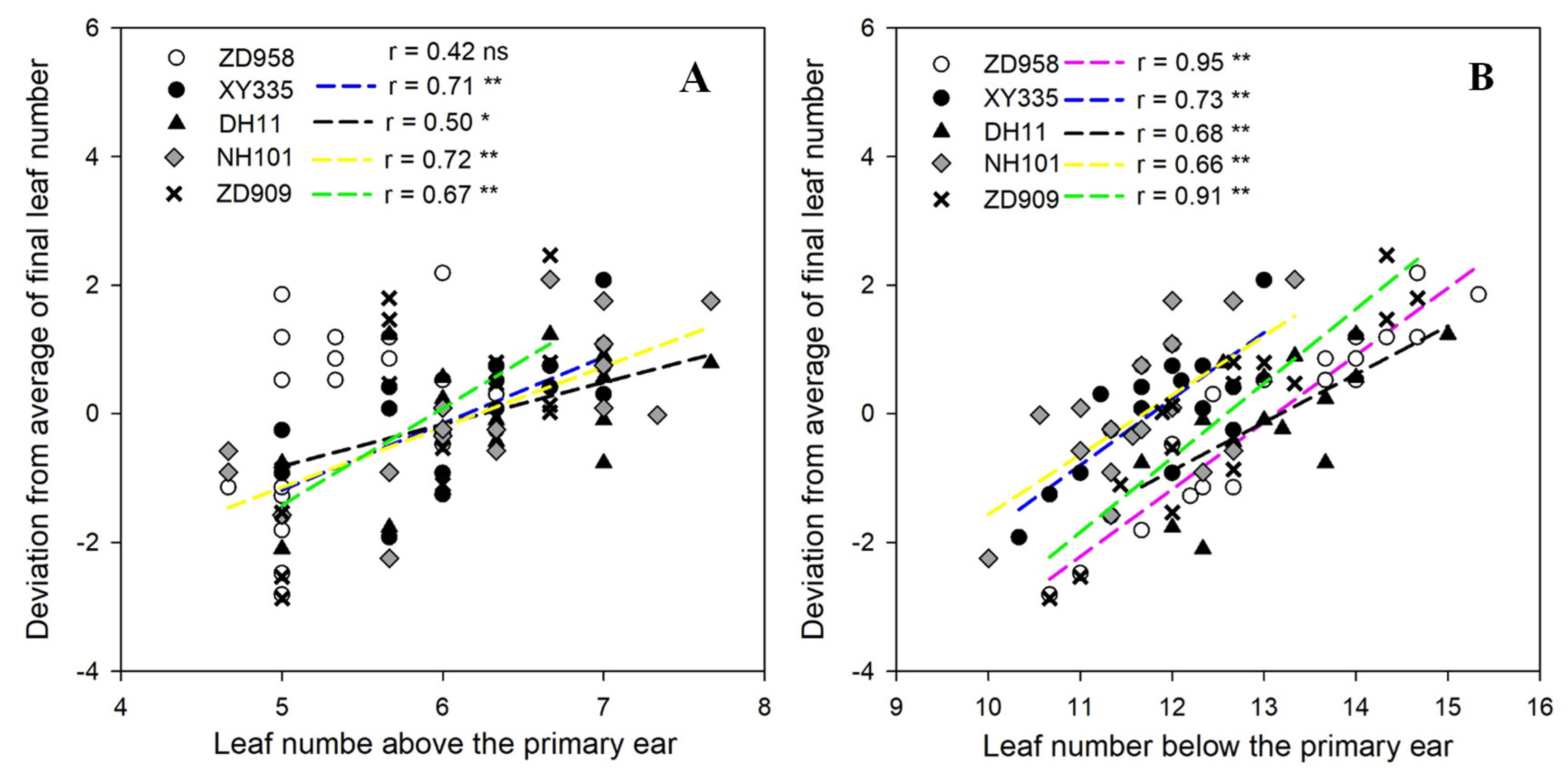
3.2. Relationships between Final Leaf Numbers and Leaf Numbers Both Above and Below the Primary Ear
3.3. Effects of Climatic Variables on Final Leaf Number
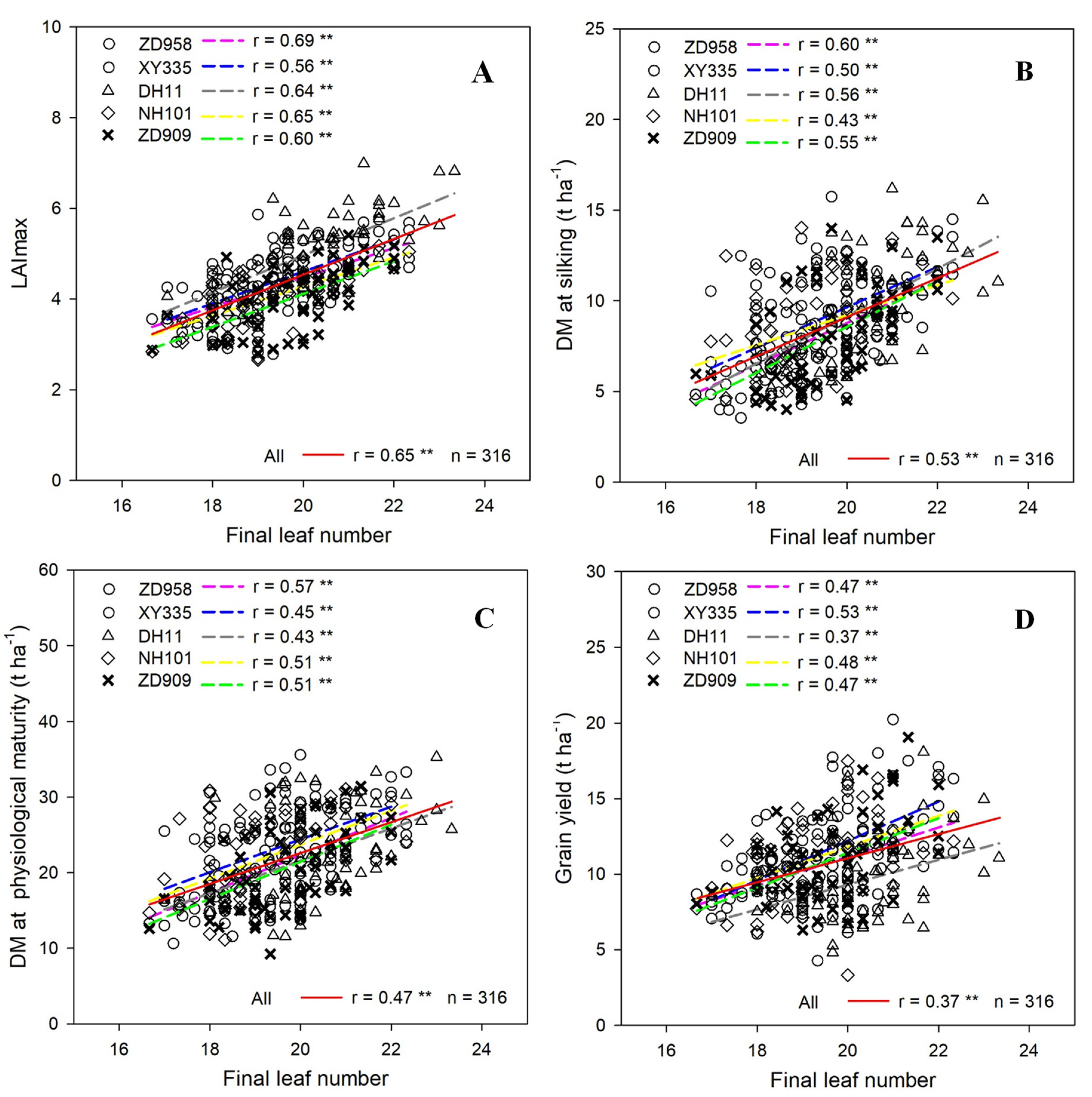
3.4. Relationships between Final Leaf Number and LAImax, DM, and Grain Yield
4. Discussion
4.1. Maize Final Leaf Numbers and Leaf Numbers Above and Below the Primary Ear
4.2. Relationships between Final Leaf Number and Climatic Factors and Vegetative Growth Days
4.3. Relationship between Final Leaf Numbers and DM and Grain Yield
4.4. Maize Final Leaf Numbers and Crop Models
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ort, D.R.; Long, S.P. Botany. Limits on yields in the Corn Belt. Science 2014, 344, 484–485. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT—Agriculture Database. Available online: http://faostat.fao.org/ (accessed on 15 December 2016).
- Leff, B.; Ramankutty, N.; Foley, J.A. Geographic distribution of major crops across the world. Glob. Biogeochem. Cycles 2004, 18, 1–27. [Google Scholar] [CrossRef]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Shabani, F. Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Sci. Rep. 2017, 7, 5910–5913. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Liu, Y.; Xie, R.; Ming, B.; Ma, D.; Li, S.; Mei, X. Temporal and spatial variation in accumulated temperature requirements of maize. Field Crop Res. 2014, 158, 55–64. [Google Scholar] [CrossRef]
- Liu, B.; Chen, X.; Meng, Q.; Yang, H.; Wart, J.V. Estimating maize yield potential and yield gap with agro-climatic zones in China—Distinguish irrigated and rainfed conditions. Agric. Meteorol. 2017, 239, 108–117. [Google Scholar] [CrossRef]
- Bonhomme, R.; Derieux, M.; Kiniry, J.R.; Edmeades, G.O.; Ozier-Lafontaine, H. Maize leaf number sensitivity in relation to photoperiod in multilocation field trials. Agron. J. 1991, 83, 153–157. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, R.; Hou, P.; Li, S.; Liang, S. Phenological responses of maize to changes in environment when grown at different latitudes in China. Field Crop Res. 2013, 144, 192–199. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Xia, L.; Xia, F.; Zhao, B. Effects of waterlogging on the yield and growth of summer maize under field conditions. Can. J. Plant Sci. 2014, 94, 23–31. [Google Scholar] [CrossRef]
- Cavero, J.; Farre, I.; Debaeke, P.; Faci, J.M. Simulation of maize yield under water stress with the EPICphase and CROPWAT Models. Agron. J. 2000, 92, 679–689. [Google Scholar] [CrossRef]
- Elawad, H.E. Growth and Production of Maize (Zea Mays L.) as Affected by Water Treatment, Organic and Inorganic Fertilizers. Ph.D. Thesis, University of Khartoum, Khartoum, Sudan, 2007. [Google Scholar]
- Ellis, R.H.; Summerfield, R.J.; Edmeades, G.O.; Roberts, E.H. Photoperiod, leaf number, and interval from tassel initiation to emergence in diverse cultivars of maize. Crop Sci. 1992, 32, 398–403. [Google Scholar] [CrossRef]
- Tollenaar, M.; Hunter, R.B. A photoperiod and temperature sensitive period for leaf number of maize1. Crop Sci. 1983, 23, 457–460. [Google Scholar] [CrossRef]
- Warrington, I.J.; Kanemasu, E.T. Corn growth response to temperature and photoperiod. III. leaf number1. Agron. J. 1983, 75, 762–766. [Google Scholar] [CrossRef]
- Zhang, F.; Mugo, S. Primary study on the effect of long photo-period on different maize Germplasm. J. Maize Sci. 2001, 9, 54–56. [Google Scholar]
- Birch, C.J.; Hammer, G.L.; Rickert, K.G. Temperature and photoperiod sensitivity of development in five cultivars of maize (Zea mays L.) from emergence to tassel initiation. Field Crop Res. 1998, 55, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Hunter, R.B.; Hunt, L.A.; Kannenberg, L.W. Photoperiod and temperature effects on corn. Can. J. Plant Sci. 1974, 54, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Birch, C.J.; Vos, J.; Putten, P.E.L. Plant development and leaf area production in contrasting cultivars of maize grown in a cool temperate environment in the field. Eur. J. Agron. 2003, 19, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Tollenaar, M. Duration of the grain-filling period in maize is not affected by photoperiod and incident PPFD during the vegetative phase. Field Crop Res. 1999, 62, 15–21. [Google Scholar] [CrossRef]
- Allen, J.R.; Mckee, G.W.; Mcgahen, J.H. Leaf number and maturity in hybrid corn1. Agron. J. 1973, 65, 233–235. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Rácz, C.; Dobos, A.C.; Helyes, L. Effect of water supply on spectral traits and their relationship with the productivity of sweet corns. Agronomy 2019, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, X.; Zhang, X.; Chen, Q.; Xu, G.; Xu, D.; Wang, C.; Liang, Y.; Wu, L.; Huang, C.; et al. The genetic architecture of leaf number and its genetic relationship to flowering time in maize. New Phytol. 2016, 210, 256–268. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Han, Y.; Li, Y.; Wang, G.; Feng, L.; Fan, Z.; Du, W.; Yang, B.; Cao, C.; Mao, S. Spatial distribution of light interception by different plant population densities and its relationship with yield. Field Crop Res. 2015, 184, 17–27. [Google Scholar] [CrossRef]
- Liu, G.; Hou, P.; Xie, R.; Ming, B.; Wang, K.; Xu, W.; Liu, W.; Yang, Y.; Li, S. Canopy characteristics of high-yield maize with yield potential of 22.5 Mg ha−1. Field Crop Res. 2017, 213, 221–230. [Google Scholar] [CrossRef]
- Shaver, D. Genetics and breeding of maize with extra leaves above the ear. In Proceedings of the 38th Annual Corn and Sorghum Industry Research Conference, Washington, DC, USA, 7–8 December 1983; pp. 161–180. [Google Scholar]
- Begna, S.H.; Hamilton, R.I.; Dwyer, L.M.; Stewart, D.W.; Smith, D.L. Variability among maize hybrids differing in canopy architecture for above-ground dry matter and grain yield. Maydica 2000, 45, 135–141. [Google Scholar]
- Subedi, K.D.; Ma, B.L. Ear position, leaf area, and contribution of individual leaves to grain yield in conventional and leafy maize hybrids. Crop Sci. 2005, 45, 2246–2257. [Google Scholar] [CrossRef]
- Begna, S.H.; Hamilton, R.I.; Dwyer, L.M.; Stewart, D.W.; Cloutier, D.; Assemat, L.; Foroutan-pour, K.; Smith, D.L. Morphology and yield response to weed pressure by corn hybrids differing in canopy architecture. Eur. J. Agron. 2001, 14, 293–302. [Google Scholar] [CrossRef]
- Dwyer, L.M.; Andrews, C.J.; Stewart, D.W.; Ma, B.L.; Dugas, J.A. Carbohydrate levels in field-grown leafy and normal maize genotypes. Crop Sci. 1995, 35, 1020–1027. [Google Scholar] [CrossRef]
- Andrews, C.J.; Dwyer, L.M.; Stewart, D.W.; Dugas, J.A.; Bonn, P. Distribution of carbohydrate during grainfill in Leafy and normal maize hybrids. Can. J. Plant Sci. 2000, 80, 87–95. [Google Scholar] [CrossRef]
- Birch, C.J.; Rickert, K.G.; Hammer, G.L. Modelling leaf production and crop development in maize (Zea mays L.) after tassel initiation under diverse conditions of temperature and photoperiod. Field Crop Res. 1998, 58, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Lizaso, J.I.; Batchelor, W.D.; Westgate, M.E. A leaf area model to simulate cultivar-specific expansion and senescence of maize leaves. Field Crop Res. 2003, 80, 1–17. [Google Scholar] [CrossRef]
- Pohlert, T. Use of empirical global radiation models for maize growth simulation. Agric. Meteorol. 2004, 126, 47–58. [Google Scholar] [CrossRef]
- Van Wart, J.; Kersebaum, K.C.; Peng, S.; Milner, M.; Cassman, K.G. Estimating crop yield potential at regional to national scales. Field Crop Res. 2013, 143, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Li, S.K.; Wang, C.T. Potential and Ways to High Yield in Maize; Science Press: Beijing, China, 2010; pp. 15–85. [Google Scholar]
- Li, J. Production, breeding and process of maize in China. In Handbook of Maize: Its Biology; Bennetzen, J.L., Hake, S.C., Eds.; Springer: New York, NY, USA, 2009; pp. 563–576. [Google Scholar]
- Hou, P.; Liu, Y.E.; Liu, W.M.; Liu, G.Z.; Xie, R.Z.; Wang, K.R.; Ming, B.; Wang, Y.H.; Zhao, R.L.; Zhang, W.J.; et al. How to increase maize production without extra nitrogen input. Resour. Conserv. Recy. 2020, 160, 104913. [Google Scholar] [CrossRef]
- Ming, B.; Xie, R.; Hou, P.; Li, L.; Wang, K.; Li, S. Changes of maize planting density in China. Sci. Agric. Sin. 2017, 50, 1960–1972. [Google Scholar]
- Hou, P.; Gao, Q.; Xie, R.; Li, S.; Meng, Q.; Kirkby, E.A.; Römheld, V.; Müller, T.; Zhang, F.; Cui, Z.; et al. Grain yields in relation to N requirement: Optimizing nitrogen management for spring maize grown in China. Field Crop Res. 2012, 129, 1–6. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, P.; Xie, R.; Hao, W.; Li, S.; Mei, X. Spatial variation and improving measures of the utilization efficiency of accumulated temperature. Crop Sci. 2015, 55, 1806–1817. [Google Scholar] [CrossRef]
- Xu, W.; Liu, C.; Wang, K.; Xie, R.; Ming, B.; Wang, Y.; Zhang, G.; Liu, G.; Zhao, R.; Fan, P.; et al. Adjusting maize plant density to different climatic conditions across a large longitudinal distance in China. Field Crop Res. 2017, 212, 126–134. [Google Scholar] [CrossRef]
- Zhen, X.; Shao, H.; Zhang, W.; Huo, W.; Batchelor, W.D.; Hou, P.; Wang, E.; Mi, G.; Miao, Y.; Li, H.; et al. Testing a bell-shaped function for estimation of fully expanded leaf area in modern maize under potential production conditions. Crop J. 2018, 6, 527–537. [Google Scholar] [CrossRef]
- Liu, G.; Yang, Y.; Liu, W.; Guo, X.; Xue, J.; Xie, R.; Ming, B.; Wang, K.; Hou, P.; Li, S. Leaf removal affects maize morphology and grain yield. Agronomy 2020, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- The National Meteorological Information Center of the China Meteorological Administration (CMA). Available online: http://data.cma.cn/ (accessed on 6 November 2017).
- Yan, M.; Liu, X.; Zhang, W.; Li, X.; Liu, S. Spatio-temporal changes of ≥10°C accumulated temperature in northeastern China since 1961. Chin. Geogr. Sci. 2011, 21, 17–26. [Google Scholar] [CrossRef]
- Jones, C.A.; Kiniry, J.R. CERES-Maize: A Simulation Model of Maize Growth and Development; Texas A&M University Press: College Station, TX, USA, 1986; p. 194. [Google Scholar]
- Hocking, R.R. The analysis and selection of variables in linear regression. Biometrics 1976, 32, 1–49. [Google Scholar] [CrossRef]
- Pecina-Martínez, J.A.; Mendoza-Castillo, M.C.; López-Santillán, J.A.; Castillo-González, F.; Mendoza-Rodríguez, M. Phenological and morphological response of tamaulipas maize to contrasting environments in México. Agrociencia 2009, 43, 681–694. [Google Scholar]
- Cui, M.; Jia, B.; Liu, H.; Kan, X.; Zhang, Y.; Zhou, R.; Li, Z.; Yang, L.; Deng, D.; Yin, Z. Genetic mapping of the leaf number above the primary ear and its relationship with plant height and flowering time in maize. Front. Plant Sci. 2017, 8, 1437. [Google Scholar] [CrossRef] [Green Version]
- Freire, A.I.; Dias, K.O.G.; Oliveira, L.B.V.; Nalin, R.S.; Guedes, F.L.; Souza, J.C. Genetic control of the number of leaves above the ear in maize. Genet. Mol. Res. 2015, 14, 1318–1323. [Google Scholar] [CrossRef] [PubMed]
- Tsimba, R.; Edmeades, G.O.; Millner, J.P.; Kemp, P.D. The effect of planting date on maize grain yields and yield components. Field Crop Res. 2013, 150, 135–144. [Google Scholar] [CrossRef]
- Keating, B.A.; Wafula, B.M. Modelling the fully expanded area of maize leaves. Field Crop Res. 1992, 29, 163–176. [Google Scholar] [CrossRef]
- Yakoub, A.; Lloveras, J.; Biau, A.; Lindquist, J.L.; Lizaso, J.I. Testing and improving the maize models in DSSAT: Development, growth, yield, and N uptake. Field Crop Res. 2017, 212, 95–106. [Google Scholar] [CrossRef]
- Muchow, R.C.; Carberry, P.S. Environmental control of phenology and leaf growth in a tropically adapted maize. Field Crop Res. 1989, 20, 221–236. [Google Scholar] [CrossRef]
- Yang, H.S.; Dobermann, A.; Cassman, K.G.; Walters, D.T. Hybrid-Maize—A Simulation Model for Corn Growth and Yield; University of Nebraska-Lincoln: Lincoln, NE, USA, 2006; p. 67. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Tmean (°C) | Tmax (°C) | Tmin (°C) | Tr (°C) | Rd (MJ m−2) | At (°C·day) | Ra (MJ m−2) | Pd (h) | Pre (mm) | A-Ir (mm) |
|---|---|---|---|---|---|---|---|---|---|---|
| NW | 18.8 | 26.4 | 11.6 | 14.9 | 10.1 | 1708.1 | 946.2 | 14.8 | 84.7 | 260 |
| SW | 21.5 | 26.4 | 18.0 | 8.4 | 6.8 | 1473.3 | 467.8 | 13.6 | 308.3 | 45 |
| HM | 26.7 | 31.4 | 22.7 | 8.6 | 8.4 | 1489.6 | 467.6 | 14.3 | 285.4 | 98 |
| NM | 20.4 | 26.5 | 14.6 | 11.8 | 9.2 | 1667.7 | 759.8 | 14.7 | 219.3 | 145 |
| Cultivar | n | Mean | Minimum | Maximum | Range | Coefficient of Variation (%) |
|---|---|---|---|---|---|---|
| DH11 | 58 | 20.4 a | 17.0 | 23.3 | 6.3 | 6.1 |
| ZD958 | 69 | 19.7 b | 16.7 | 22.3 | 5.6 | 7.1 |
| ZD909 | 58 | 19.7 b | 16.7 | 22.0 | 5.3 | 6.0 |
| NH101 | 69 | 19.2 c | 16.7 | 22.3 | 5.6 | 6.5 |
| XY335 | 62 | 19.2 c | 17.0 | 22.0 | 5.0 | 6.0 |
| ALL | 316 | 19.6 | 16.7 | 23.3 | 6.6 | 6.7 |
| Cultivar | n | Leaf Number Above Ear | Leaf Number Below Ear | Final Leaf Number | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | CV (%) | Mean | Range | CV (%) | Mean | Range | CV (%) | ||
| DH11 | 17 | 6.3 a | 5.0–7.7 | 11.7 | 14.2 a | 12.7–16.0 | 6.1 | 20.4 a | 18.3–21.7 | 4.7 |
| ZD958 | 18 | 5.4 b | 4.7–6.3 | 8.9 | 14.1 a | 11.7–16.3 | 9.4 | 19.5 b | 16.7–21.7 | 7.5 |
| ZD909 | 18 | 5.9 a | 5.0–6.7 | 10.4 | 13.6 a | 11.7–15.7 | 8.1 | 19.5 b | 16.7–22.0 | 7.2 |
| NH101 | 18 | 6.2 a | 4.7–7.7 | 14.1 | 12.7 b | 11.0–14.3 | 6.3 | 18.9 b | 16.7–21.0 | 6.1 |
| XY335 | 18 | 6.1 a | 5.0–7.0 | 11.3 | 12.8 b | 11.3–14.0 | 5.6 | 18.9 b | 17.0–21.0 | 5.4 |
| ALL | 89 | 6.0 | 4.7–7.7 | 12.6 | 12.5 | 10.0–15.3 | 9.3 | 19.5 | 16.7–22.0 | 6.8 |
| Cultivar | Tmean (°C) | Tmax (°C) | Tmin (°C) | Tr (°C) | Ra (MJ m−2) | At (°C·day) | Acc-Ra (MJ m−2) | Pd (h) | Growth Duration (d) |
|---|---|---|---|---|---|---|---|---|---|
| DH11 | −0.30 * | −0.26 * | −0.37 ** | 0.36 ** | 0.03 * | 0.42 ** | 0.45 ** | 0.53 ** | 0.40 ** |
| ZD958 | −0.07 ns | 0.01 ns | −0.20 ns | 0.39 ** | 0.51 ** | 0.52 ** | 0.49 ** | 0.76 ** | 0.36 ** |
| ZD909 | −0.14 ns | −0.04 ns | −0.29 * | 0.47 ** | 0.57 ** | 0.53 ** | 0.57 ** | 0.70 ** | 0.40 ** |
| NH101 | −0.41 ** | −0.36 ** | −0.47 ** | 0.43 ** | 0.44 ** | 0.40 ** | 0.54 ** | 0.66 ** | 0.49 ** |
| XY335 | −0.50 ** | −0.44 ** | −0.55 ** | 0.47 ** | 0.32 * | 0.37 ** | 0.57 ** | 0.54 ** | 0.57 ** |
| ALL | −0.25 ** | −0.18 ** | −0.33 ** | 0.39 ** | 0.39 ** | 0.46 ** | 0.49 ** | 0.59 ** | 0.42 ** |
| Cultivar | Partial Regression Equation | R2 | Pr > P |
|---|---|---|---|
| DH11 | y = 1.323x1 + 1.48 | 0.29 | <0.01 |
| ZD958 | y = 2.058x1 − 3.93 | 0.58 | <0.01 |
| ZD909 | y = 0.383x1 − 3.13 | 0.49 | <0.01 |
| NH101 | y = 1.499x1 − 0.123x3 + 1.04 | 0.50 | <0.01 |
| XY335 | y = 0.978x1 − 0.136x2 + 8.19 | 0.43 | <0.01 |
| ALL | y = 1.411x1 − 0.039x4 + 0.059 | 0.36 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Ming, B.; Xie, R.; Liu, G.; Wang, K.; Yang, Y.; Guo, X.; Hou, P.; Li, S. Change in Maize Final Leaf Numbers and Its Effects on Biomass and Grain Yield across China. Agriculture 2020, 10, 411. https://doi.org/10.3390/agriculture10090411
Liu W, Ming B, Xie R, Liu G, Wang K, Yang Y, Guo X, Hou P, Li S. Change in Maize Final Leaf Numbers and Its Effects on Biomass and Grain Yield across China. Agriculture. 2020; 10(9):411. https://doi.org/10.3390/agriculture10090411
Chicago/Turabian StyleLiu, Wanmao, Bo Ming, Ruizhi Xie, Guangzhou Liu, Keru Wang, Yunshan Yang, Xiaoxia Guo, Peng Hou, and Shaokun Li. 2020. "Change in Maize Final Leaf Numbers and Its Effects on Biomass and Grain Yield across China" Agriculture 10, no. 9: 411. https://doi.org/10.3390/agriculture10090411