Abstract
Understanding the resistance spectrum and underlying genetic mechanisms is critical for managing herbicide-resistant populations. In this study, resistance to acetyl CoA carboxylase (ACCase) and acetolactate synthase (ALS) inhibitors was investigated in four suspected resistant populations of Alopecurus myosuroides (ALOMY-001 to ALOMY-004) and Lolium multiflorum (LOLMU-001 to LOLMU-004), collected from cereal production fields in Ireland. Glasshouse assays with three ALOMY-active herbicides [propaquizafop, cycloxydim (ACCase) and mesosulfuron + iodosulfuron (ALS)] or five LOLMU-active herbicides [pinoxaden, propaquizafop, cycloxydim (ACCase) and mesosulfuron + iodosulfuron, pyroxsulam (ALS)], and target-site resistance mechanism studies, based on pyrosequencing, were carried out in each of those populations. For A. myosuroides, Ile-1781-Leu ACCase mutation contributed to propaquizafop and cycloxydim resistance (shoot dry weight GR50 resistance factor (RF) = 7.5–35.5) in all ALOMY populations, and the independent Pro-197-Thr or Pro-197-Ser ALS mutation contributed to mesosulfuron + iodosulfuron resistance (RF = 3.6–6.6), in ALOMY-002 to ALOMY-004. Most of the analyzed plants for these mutations were homo/heterozygous combinations or only heterozygous. For L. multiflorum, phenotypic resistance to mesosulfuron + iodosulfuron (RF = 11.9–14.6) and pyroxsulam (RF = 2.3–3.1) was noted in all LOLMU populations, but the Pro-197-Gln or Pro-197-Leu ALS mutation (mostly in homozygous status) was identified in LOLMU-001, LOLMU-002 and LOLMU-004 only. Additionally, despite no known ACCase mutations in any LOLMU populations, LOLMU-002 survived pinoxaden and propaquizafop application (RF = 3.4 or 1.3), and LOLMU-003 survived pinoxaden (RF = 2.3), suggesting the possibility of non-target-site resistance mechanisms for ACCase and/or ALS resistance in these populations. Different resistance levels, as evidenced by a reduction in growth as dose increased above field rates in ALOMY and LOLMU, were due to variations in mutation rate and the level of heterozygosity, resulting in an overall resistance rating of low to moderate. This is the first study confirming cross- and multiple resistance to ACCase- and ALS-inhibiting herbicides, highlighting that resistance monitoring in A. myosuroides and L. multiflorum in Ireland is critical, and the adoption of integrated weed management strategies (chemical and non-chemical/cultural strategies) is essential.
1. Introduction
Black-grass (Alopecurus myosuroides) and Italian ryegrass (Lolium multiflorum) are the most problematic grass weeds threatening the sustainability of crop rotations dominated by autumn-sown cereals across north-western Europe [1]. They are particularly difficult to control in early-sown winter cereals established after non-inversion tillage, due to the high seed retention of freshly shed populations, and as peak seed germination between September and November coincides with the sowing and early development of winter cereals [2,3]. In the United Kingdom (UK), the economic threshold (i.e., weed density that causes a 5% yield loss) for A. myosuroides is just 12 plants m−2, and for L. multiflorum it is 5 plants m−2, suggesting that even low plant populations may reduce crop yields [4,5].
As A. myosuroides and L. multiflorum are genetically variable obligate outcrossers, with high fecundity and rapid seedbank turnover [6,7], they are at high-risk of evolving resistance from continuous herbicide use [8,9]. To date, A. myosuroides has evolved resistance to seven different herbicide modes of action (MOA) and L. multiflorum to eight different herbicide MOA, worldwide [10]. In the UK, A. myosuroides resistant to Herbicide Resistance Action Committee (HRAC) MOA groups 1 (legacy A) [acetyl-CoA carboxylase (ACCase) inhibitors], 2 (B) [acetolactate synthase (ALS) inhibitors], 3 (K1) (e.g., residual herbicide, pendimethalin) and 5 (C2) (e.g., isoproturon, which was banned in the European Union in 2017) has been reported [10]. Consequently, resistant A. myosuroides occurs on most UK crop farms that use herbicides as a single weed control method [11]. While herbicide-resistant L. multiflorum is not as widespread as resistant A. myosuroides in the UK or in Europe [1], cases are now increasing [12] and populations resistant to herbicide MOA groups 1, 2, 5, 9 (G) (e.g., non-selective herbicide, glyphosate) and 15 (K3) (e.g., residual herbicide, flufenacet) have been reported [10]. Importantly, more than 75% of resistance cases within the UK and Europe are associated with two MOA groups: ACCase/ALS inhibitors.
The ACCase inhibitors control grass weeds by inhibiting acetyl CoA carboxylase, the enzyme required for fatty acid biosynthesis, and include herbicide chemistries of phenylpyrazoline (DEN), aryloxyphenoxypropionate (FOP), and cyclohexanedione (DIM) [13]. The ALS inhibitors control grass weeds by inhibiting acetolactate synthase, a key enzyme leading to the pathway to the branched-chain amino acids valine, leucine and isoleucine, and include herbicide chemistries of sulfonylurea (SU), triazolopyrimidine (TP), imidazolinone, pyrimidinyl-thiobenzoate and sulfonyl-aminocarbonyl-triazolinone [14]. Resistance to ACCase- and ALS-inhibiting herbicides, based on, both target-site resistance (TSR) due to mutations in the target genes, and non-target-site resistance (NTSR) usually through enhanced metabolism, has been reported in field populations of A. myosuroides and Lolium spp. in various regions [11,12,15,16,17,18].
In Ireland, agriculture area accounts for 65% (approximately 4.5 million ha) of the total land area [19]. Of this agricultural area, approximately 92% is devoted to grassland and 8% to arable cereal and crop production. Crop production in Ireland has evolved over the last 50 years from a mixed enterprise farming base, where spring cereals were grown in rotation as a part of grassland system, to current specialized cereal monoculture or rotation with all annual crops [20], which has resulted in much less proliferation of A. myosuroides. While the first occurrence of A. myosuroides was reported in 1987 [21], it was only recorded as a problem in isolated cases, in contrast to the UK or parts of Europe. Lolium multiflorum can occur as a weed of cereal crops in mixed rotations, which include grass, however, it has not been reported as a serious weed in cereal rotations on farms in Ireland, until recently. Non-selective and post-emergent herbicides have been the mainstay for weed control in arable rotations [22]. Growers exclusively rely on post-emergent ACCase- and ALS-inhibiting herbicides, which are the only remaining MOA available for selective grass weed control within cereal and broad-leaved crops (with the exception of herbicide MOA group 3, propyzamide, which provides largely residual control of a range of grass weeds in winter oilseed rape) [23]. The main registered commercial ACCase- and ALS-inhibiting herbicides are targeted for the control of Avena fatua (wild oats) or Anisantha sterilis (sterile brome), which are, the most competitive grass weeds in cereal crops in Ireland. This use may have inadvertently influenced potential A. myosuroides and L. multiflorum control and resistance evolution.
In Ireland, more than 139,000 ha of cereal, oilseed rape and bean crops were treated with ACCase- and ALS-inhibiting herbicides in 2016, with ACCase-DEN-pinoxaden being the most widely used herbicide to control A. fatua, on at least 114,927 ha of wheat and barley (spring and winter) area, which represents an 11% increase in pinoxaden-treated area, from 2012 [22]. The ACCase-FOP-propaquizafop and ACCase-DIM-cycloxydim were used on 7792 ha (3% decrease) of broad-leaved crops, including oilseed rape and beans to control A. fatua and A. sterilis, while ALS-SU-mesosulfuron + iodosulfuron was used on 13,664 ha of winter wheat (0.4% increase) and ALS-TP-pyroxsulam was used on 3367 ha (29% increase) of winter wheat. Many of these A. fatua herbicides are also active on L multiflorum while these herbicides other than the ACCase-pinoxaden and ALS-pyroxsulam are active on A. myosuroides. It is logical to expect that recent practices may have allowed some degree of resistance to develop in isolated A. myosuroides and L. multiflorum populations, to ACCase and/or ALS inhibitors, even though they may not have been the intended target.
Monitoring of herbicide-resistant grass weeds in Ireland’s climate and cropping system is only recently being addressed unlike in the UK, as these challenges developed later [23]. A recent grass weed survey in Ireland found suspect A. myosuroides and L. multiflorum strains that were difficult to control during the growing season. To date, no reports of herbicide resistance have been reported for control failures of A. myosuroides and/or L. multiflorum in Ireland. The aims of the present study were to: (a) determine the resistance patterns and the resistance levels to ACCase- and ALS-inhibiting herbicides in the suspected resistant populations of A. myosuroides and L. multiflorum, collected from cereal production fields in Ireland, and (b) to verify whether a TSR mechanism is involved in order to identify the mutation(s) that confer resistance. This knowledge is critical to develop integrated weed management (IWM) strategies to eliminate or control these resistant populations at a critical point in the development of weed challenges in this region.
2. Materials and Methods
2.1. Seed Materials and Growing Conditions
A nationwide observational survey of 136 farms was conducted from June to September 2020, which included a questionnaire, grass weed assessments, and the collection of seed samples for glasshouse resistance testing (Vijaya Bhaskar et al. unpublished). The survey farms were selected using a stratified pattern, to ensure different types of crop establishment systems were represented, using grower lists provided by Teagasc (the state-funded research and advisory service) and commercial crop advisors. The study fields within farms were identified on the basis of specific crop establishment systems practiced. A total of 157 field samples (approx. one sample per farm) of the most problematic grass weed species specified by each grower: A. fatua (73 populations), Anisantha spp. and Bromus spp., (52 populations), Phalaris minor (12 populations), A. myosuroides (12 populations) and L. multiflorum (8 populations) were collected. From this survey, we found four A. myosuroides (ALOMY-001 to ALOMY-004) and four L. multiflorum (LOLMU-001 to LOLMU-004) populations, with a high herbicide survival rate in the fields, and growers had reported control failures or difficult-to-control with ACCase- and/or ALS-inhibiting herbicides at recommended field rate, that were used in this study.
Information regarding the position and field history of each of the suspected resistant populations is given in Table 1 and Table 2. In addition, a known ACCase/ALS-susceptible A. myosuroides population S-006 (Table 1), and a susceptible L. multiflorum population (SORG-002) from an organic seed supplier (Fruit Hill Farm, Co. Cork, Ireland) were used as susceptible controls. The fields within Co. Meath for ALOMY-002 and ALOMY-003, and LOLMU-003 and LOLMU-004, were located c. 25.6 km and c. 31.2 km apart, respectively. Seeds from suspect resistant and susceptible populations were collected, air-dried and stored in paper bags at 15 °C.

Table 1.
Location and field history of the suspected resistant (ALOMY-001 to ALOMY-004) and susceptible (S-006) populations of A. myosuroides collected from cereal production fields prior to 2020-harvest for resistance testing.
Table 2.
Location and field history of the suspected resistant (LOLMU-001 to LOLMU-004) populations of L. multiflorum collected from cereal production fields prior to 2020-harvest for resistance testing.
Prior to the resistance testing experiments, seed populations of A. myosuroides were placed in an incubator at 30 °C for six weeks to break seed dormancy. Subsequently, seeds from A. myosuroides and L. multiflorum populations were placed in a petri-dish with dense filter paper (VWR ® Grade 413), imbibed overnight in a combination of gibberellic acid (GA3) and potassium nitrate (KNO3) solution to assist seed germination [23]. Seeds were then planted in 35.5 × 21.5 × 5 cm plastic trays containing Kettering loam and lime-free grit mix in a 4:1 ratio, with the addition of Osmocote Mini™ (1 kg t−1). Each population had 35 seeds planted per tray for each replicate. Trays were watered as required throughout the experiment. The plants of A. myosuroides and L. multiflorum were grown in two separate glasshouse compartments with an 18/12 °C (day/night) temperature regime at a photoperiod of 16 h, supplemented with artificial lighting to maintain a minimum light intensity of 250 μmol quanta m−2 s−1 at the Teagasc Crops Research Centre, Oak Park, Carlow/Ireland.
2.2. Single-Dose Assays
A range of registered ACCase- and ALS-inhibiting herbicides for A. myosuroides and/or L. multiflorum control were tested at the recommended application rates (for field application in Ireland) to determine the sensitivity and resistance patterns of the populations of A. myosuroides and L. multiflorum.
For A. myosuroides, two ACCase inhibitors propaquizafop (Falcon ®, Adama) at 100 g active substance (a.s) ha−1; and cycloxydim (Stratos Ultra ®, BASF) at 200 g a.s ha−1 and one ALS inhibitor mesosulfuron + iodosulfuron (Pacifica Plus ®, includes amidosulfuron, Bayer CropScience Ltd., Cambridge, UK) at 15 + 5 g a.s ha−1 plus 1% v/v Biopower adjuvant (6.7% w/w 3, 6-dioxaeicosylsulphate sodium salt (EAC1) and 20.1% w/w 3, 6-dioxaoctadecylsulphate sodium salt, Bayer), were applied. For L. multiflorum, three ACCase inhibitors pinoxaden (Axial Pro ®, includes cloquintocet-mexyl, Syngenta) at 30.3 g a.s ha−1 plus 1% v/v Adigor adjuvant (47% w/w methylated rapeseed oil, Syngenta); propaquizafop at 150 g a.s ha−1 and cycloxydim at 150 g a.s ha−1 and two ALS inhibitors mesosulfuron + iodosulfuron at 15 + 5 g a.s ha−1 plus 1% v/v Biopower adjuvant and pyroxsulam (Broadway star ®, includes florasulam, Corteva Agrisciences) at 18.8 g a.s ha−1 plus 1% v/v Kantor adjuvant (EC 790 g L−1 alkoxylated triglycerides, Interagro Ltd., Bishop’s Stortford, UK) were applied.
Herbicides were applied to the plants at growth stage (GS) 12–13 (BBCH) [24], using a Generation III Research Track Sprayer (DeVries Manufacturing, Hollandale, MN, USA) with a Teejet 8002-EVS flat fan nozzle, at a pressure of 2.5 bar and a water volume equivalent of 200 L ha−1. The experiment was a randomized design with two replicates, a minimum of 27 plants per population for each replicate of each herbicide treatment and was repeated twice. In addition to untreated controls, susceptible populations S-006 for A. myosuroides and SORG-002 for L. multiflorum were included. At 28 days after spraying, plants were rated as dead (susceptible) or alive (resistant). Above-ground shoots were harvested, dried at 70° C for 72 h and weighed. Plant survival was assessed for each population at each herbicide treatment by expressing the percentage of surviving seedlings. The above ground shoot dry weight for each replicate was expressed as the percentage of the mean dry weight of the untreated control replicates of the corresponding population.
2.3. Dose-Response Assays
Dose-response assays on all populations were carried out to quantify the level of resistance or sensitivity to ACCase- and ALS-inhibiting herbicides. When seedlings reached the 2–3 leaf growth stage, three independent dose-response experiments for A. myosuroides and five independent dose-response experiments for L. multiflorum were established (Table 3). The selected herbicide dose rates represent 0.25 to 8 times the recommended field rate in Ireland. Each experiment was randomized with three replicates, a minimum of 26 plants per population per replicate for each dose. Above-ground shoot dry weights were harvested and measured 28 days after spraying using methodology above.
Table 3.
ACCase and ALS inhibitor dose-response herbicide treatments.
2.4. Analysis of Target-Site Resistance Mutations
Prior to collecting leaf samples to detect TSR mutations within the ACCase and ALS genes, suspected resistant plants at the 2–3 leaf growth stage (15 plants per population) were sprayed with ALS-inhibiting herbicide mesosulfuron + iodosulfuron (15 + 5 g a.s. ha−1) and additionally, A. myosuroides plants were sprayed with ACCase-inhibiting herbicide cycloxydim (200 g a.s. ha−1), all applied at recommended field rates. Eight leaf blades from eight individual plants per resistant population that had survived herbicide treatments, and from untreated control plants in susceptible populations, were harvested 21 days after spraying and kept at −20 °C until used. Genomic DNA was extracted from a total of 80 leaf samples with the MagMAX TM Plant DNA Kit using the Thermo Scientific TM Pharma Kingfisher TM flex 96 Deep-well Head Magnetic Particle Processor. The DNA concentration of each sample was determined using the Quanti-iT TM PicoGreen TM dsDNA Assay kit (Invitrogen TM). Then dilutions were made at 40 ng µL−1. A sealed elusion plate, containing frozen DNA, was placed upright in a Styrofoam cooler surrounded by cold ice packs, and sent to IDENTXX, Stuttgart, Germany for TSR genotyping using pyrosequencing. Known homozygous and heterozygous point mutations in the ACCase gene at position Ile-1781, Trp-2027, lle-2041, Asp-2078 and Gly-2096, and in the ALS gene at position Pro-197 and Trp-574 were analyzed Sequencing and data analysis were performed as described by Keshtkar et al. [25].
2.5. Statistical Analysis
Survival data and shoot dry weight data for single-dose assays of A. myosuroides and L. multiflorum populations were analyzed separately using ANOVA in R (v.3.6.3), to determine treatment-by-experiment interactions (two replicates per experiment). The interactions for percent survival or percent shoot dry weight was not significant (p < 0.05). Therefore, the data was averaged over the four replicates in each of A. myosuroides and L. multiflorum populations.
Dose-response models were fitted to the shoot dry weight data using the drc package [26] in R (v.3.6.3). Lack-of-fit F-tests were performed to assess model fit (p > 0.05). For A. myosuroides, a four-parameter log-logistic model was used to model the propaquizafop and cycloxydim shoot dry weight data, and a four-parameter Weibull-1 model to model the mesosulfuron + iodosulfuron shoot dry weight data. As residuals were normally distributed, data were not transformed. For L. multiflorum, a four-parameter log-logistic model with a Box-Cox transformation was used to model the pinoxaden, propaquizafop and cycloxydim shoot dry weight data, and a four-parameter Weibull-1 model with a Box-Cox transformation to model the mesosulfuron + iodosulfuron and pyroxsulam shoot dry weight data. As virtually complete control of a susceptible population S-006 for A. myosuroides or SORG-002 for L. multiflorum was achieved at the lowest dose (0.25x field rate) used for most herbicides, a model could not be fitted to these populations. Fitted models estimated the growth rate GR50 (i.e., the effective dose required to obtain a growth reduction of 50% relative to untreated plants) and resistance factor (RF) calculated as the ratio between the GR50 values of each resistant population by the GR50 values of the susceptible control.
3. Results
3.1. Single-Dose Assays
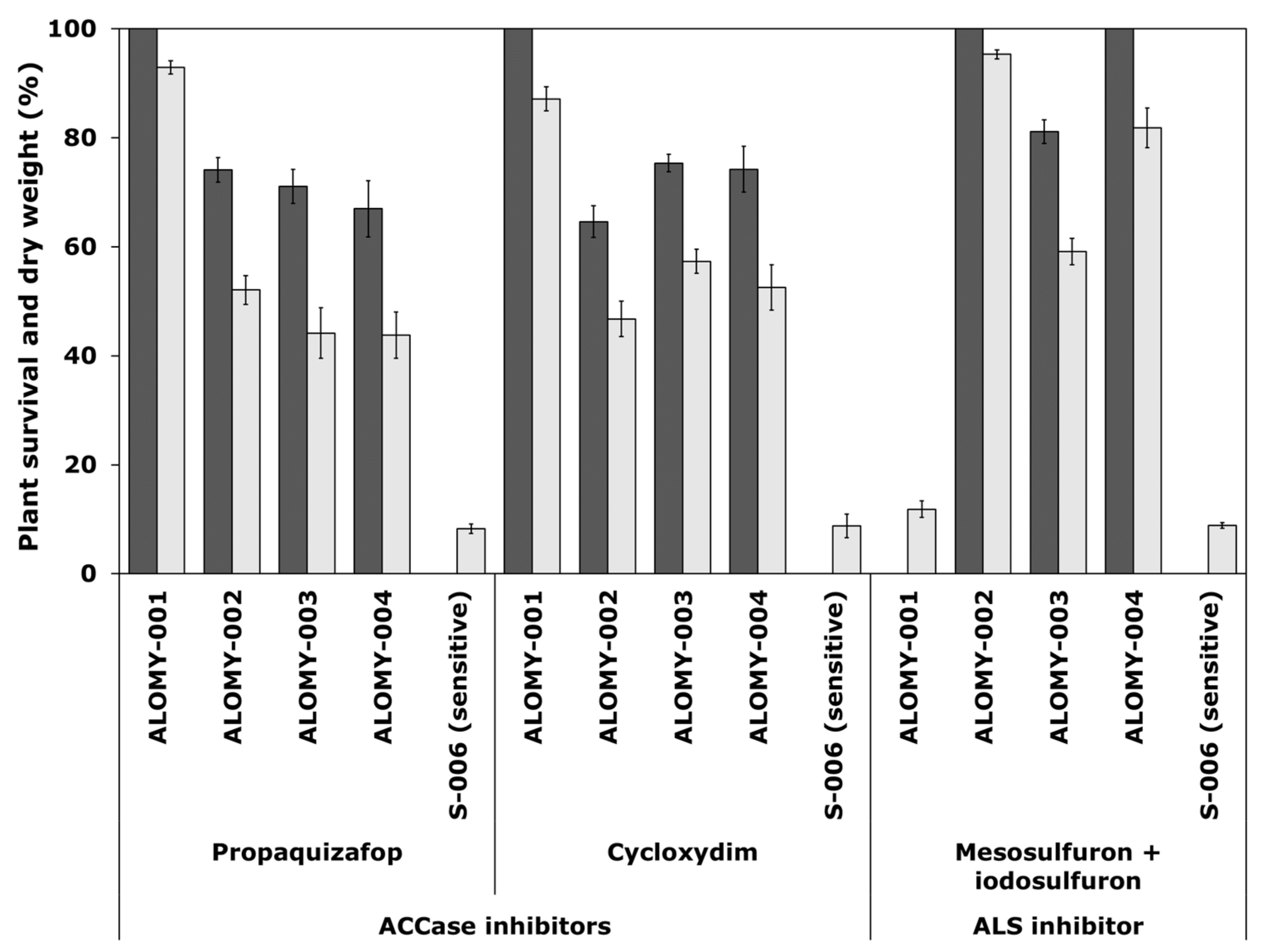
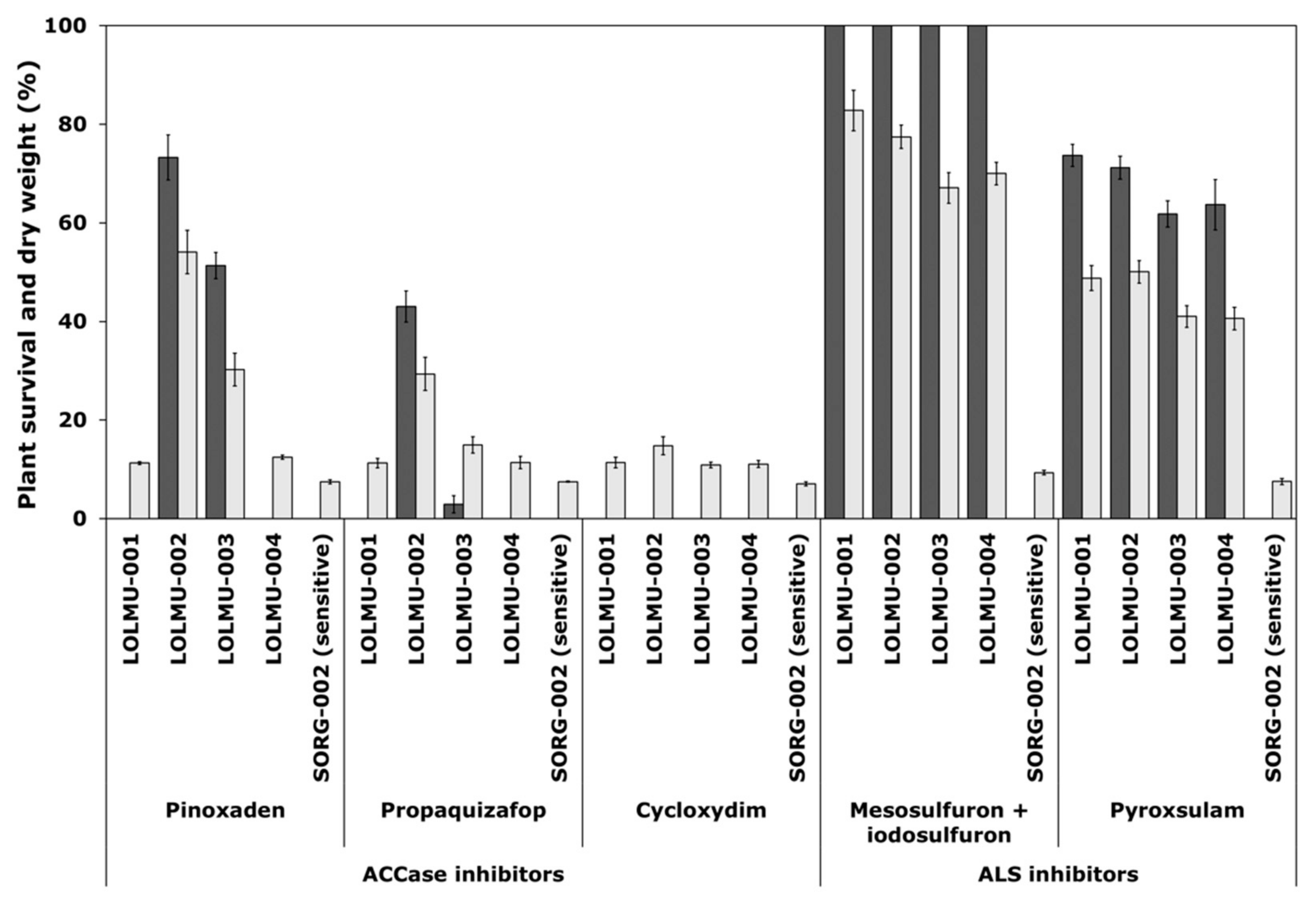
The susceptible A. myosuroides population S-006 and the susceptible L. multiflorum population SORG-002 were totally controlled by all herbicides tested at recommended field rate (Figure 1 and Figure 2).
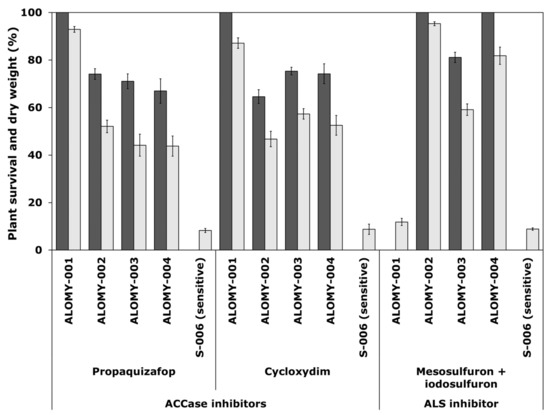
Figure 1.
Survival (black) and shoot dry weight (grey), expressed as a percentage of untreated controls of A. myosuroides populations. These were treated with two ACCase-inhibiting herbicides propaquizafop (100 g a.s ha−1) and cycloxydim (200 g a.s ha−1) and one ALS-inhibiting herbicide mesosulfuron + iodosulfuron (15 + 5 g a.s ha−1), all applied at recommended Ireland field rate. Error bars are standard error of the mean.
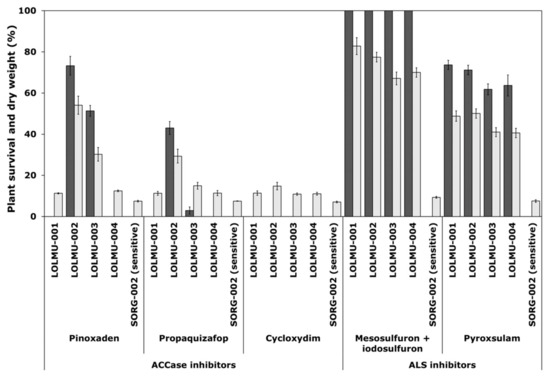
Figure 2.
Survival (black) and shoot dry weight (grey), expressed as a percentage of untreated controls of L. multiflorum populations. These were treated with three ACCase-inhibiting herbicides pinoxaden (30.3 g a.s ha−1) propaquizafop (150 g a.s ha−1) and cycloxydim (150 g a.s ha−1) and two ALS-inhibiting herbicides mesosulfuron + iodosulfuron (15 + 5 g a.s ha−1) and pyroxsulam (18.8 g a.s ha−1), all applied at recommended Ireland field rate. Error bars are standard error of the mean.
For A. myosuroides, resistance was recorded in suspected resistant population ALOMY-001 to both ACCase-inhibiting herbicides propaquizafop (100% survival) and cycloxydim (100% survival), but was totally controlled by the ALS-inhibiting herbicide mesosulfuron + iodosulfuron (Figure 1). Resistance was recorded in the other three suspect populations ALOMY-002, ALOMY-003 and ALOMY-004 to both MOA, with a plant survival rate of 65% to 75% to ACCase inhibitors, respectively, and 81% or 100% to an ALS inhibitor.
For L. multiflorum, resistance was recorded in all four suspect populations (LOLMU-001 to LOLMU-004) to both ALS-inhibiting herbicides mesosulfuron + iodosulfuron (100% survival) and pyroxsulam (62–74% survival) (Figure 2). LOLMU-001 and LOLMU-004 were totally controlled by all three ACCase-inhibiting herbicides pinoxaden, propaquizafop and cycloxydim. Whereas, with LOLMU-002 resistance or variation in sensitivity to pinoxaden and propaquizafop (73% or 43% survival) was recorded, and similarly in LOLMU-003 to pinoxaden (51% survival) and propaquizafop (3% survival only, which is still susceptible), but cycloxydim was found to be effective on both. The results from shoot dry weight data were similar to that of plant survival for the two species (Figure 1 and Figure 2).
3.2. Dose-Response Assays
Shoot dry weight dose-response models confirmed that there were significant differences in response (p < 0.01) amongst the A. myosuroides or L. multiflorum populations in GR50 when treated with ACCase- and ALS-inhibiting herbicides. Model fitting confirmed the extent of resistance, depending on the population and the herbicide, which had been identified in the single-dose assays.
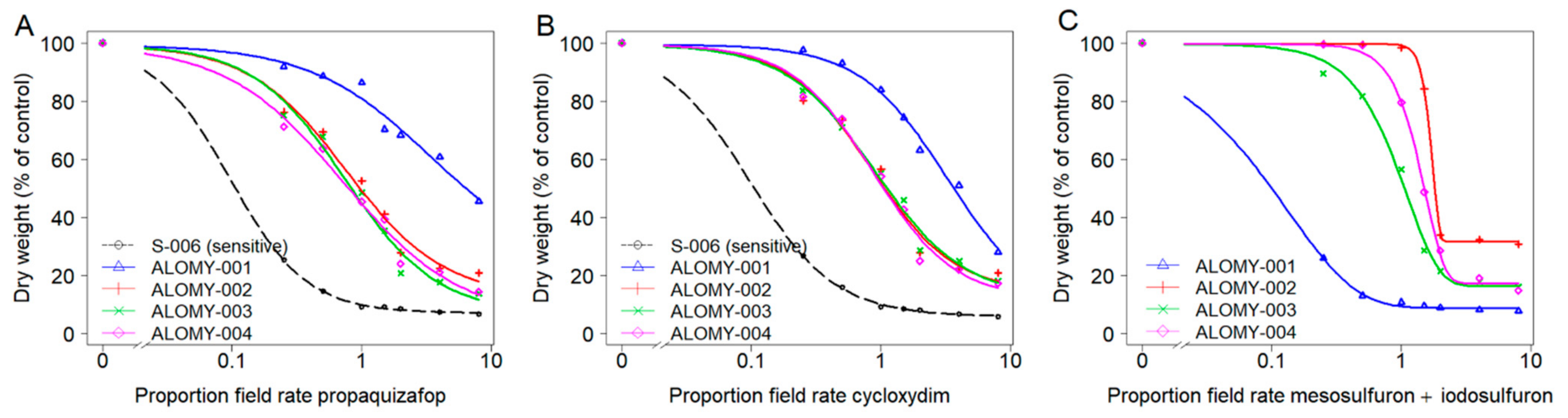
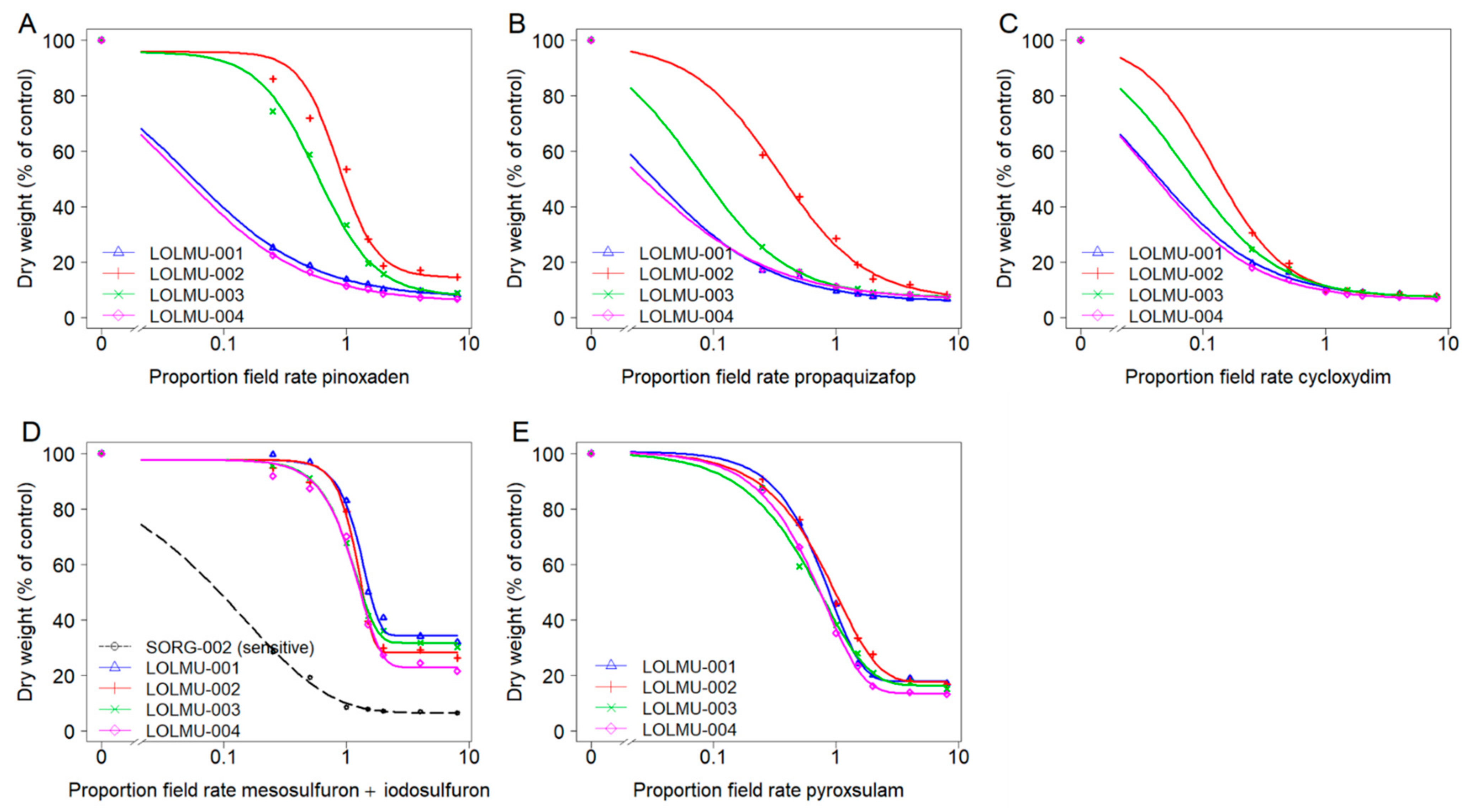
The susceptible populations S-006 for A. myosuroides, and SORG-002 for L. multiflorum were well controlled, with shoot dry weight reduced by >93% at the recommended field rate of both ACCase and ALS-inhibiting herbicides. In contrast, even though the highest dose rate (i.e., 8x field rate) did not cause complete mortality, shoot dry weight data revealed that the growth of the suspect resistant populations (ALOMY or LOLMU) was clearly affected as the dose rate increased, with more pronounced effects observed from herbicide dose rate equal to or above 1.5× field rate (Figure 3 and Figure 4).
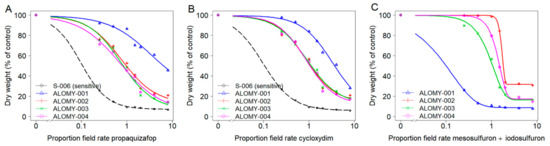
Figure 3.
Dose-response analysis of A. myosuroides populations for two ACCase-inhibiting herbicides propaquizafop (A) and cycloxydim (B) and one ALS-inhibiting herbicide mesosulfuron + iodosulfuron (C). Note: The control of susceptible population S-006 at the lowest dose was virtually complete, therefore a model (C) could not be fitted with S-006.
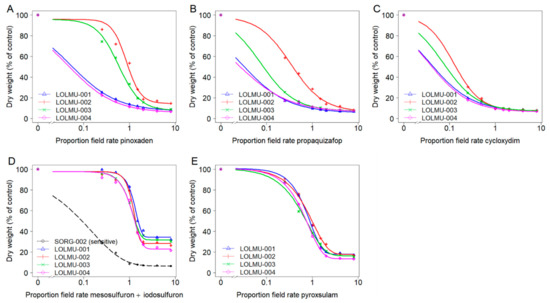
Figure 4.
Dose-response analysis of L. multiflorum populations for three ACCase-inhibiting herbicides pinoxaden (A), propaquizafop (B) and cycloxydim (C) and two ALS-inhibiting herbicides mesosulfuron + iodosulfuron (D) and pyroxsulam (E). Note: With the exception of model (D), the control of susceptible population SORG-002 at the lowest dose was virtually complete and hence other models could not be fitted with SORG-002.
For A. myosuroides, suspect resistant population ALOMY-001 displayed the highest resistance, with the GR50 herbicide application rate value estimates being more than 3.2 times the recommended field rate of both ACCase-inhibiting herbicides (Figure 3A,B), and RF was >30 for propaquizafop or cycloxydim (Table 4). In the other three suspect populations ALOMY-002 to ALOMY-004, while their GR50 values being slightly below recommended field rate (0.7–0.9), their RF of between 7.5 and 7.9 for propaquizafop, and between 8.7 and 9.3 for cycloxydim, indicated ineffective control with both ACCase-inhibiting herbicides. For the ALS-inhibiting herbicide, GR50 values of ALOMY-002 and ALOMY-004 populations were 1.7 or 1.4 times the recommended field rate of mesosulfuron + iodosulfuron, and the GR50 values of ALOMY-003 were close to recommended field rate (0.9) (Figure 3C). The RF ranged between 3.6 and 6.6 for mesosulfuron + iodosulfuron (Table 4), indicating that ALOMY-002, ALOMY-003 and ALOMY-004 were also ALS-resistant. GR50 values of ALOMY-001 was similar to S-006, confirming that ALOMY-001 was ALS-sensitive.
Table 4.
GR50 values (standard errors in parenthesis) from shoot dry weight response to two ACCase inhibitors propaquizafop and cycloxydim and one ALS inhibitor mesosulfuron + iodosulfuron for susceptible (S-006) and resistant (ALOMY-001 to ALOMY-004) populations of A. myosuroides. Resistance factor (RF) was calculated as ratio between the GR50 of resistant population by the GR50 of the susceptible population. ACCase and ALS mutations identified at Ile-1781 or Pro-197 position only. Eight plants were analyzed per population. Number of homozygous and heterozygous A. myosuroides plants for specific mutations are reported in the bracket.
For L. multiflorum, suspect resistant populations LOLMU-002 and LOLMU-003 were statistically different (p < 0.05), compared with the remaining two resistant populations with each of the three ACCase-inhibiting herbicides pinoxaden, propaquizafop and cycloxydim (Figure 4A–C). GR50 values of LOLMU-001 and LOLMU-004 were consistent with SORG-002 across ACCase-inhibiting herbicides (Table 5). Whereas, GR50 values of LOLMU-002 and LOLMU-003 were slightly below recommended field rate of pinoxaden (0.9 or 0.6), their RF was 3.4 or 2.3, indicating reduced sensitivity. In addition, while GR50 values of LOLMU-002 remained well below recommended field rate of propaquizafop (0.3), and RF was also low (1.3), there were some surviving plants, even at the 1.5× field rate, strongly indicating that it is in the process of evolving resistance. Conversely, although LOLMU-003 had a few survivors following propaquizafop application, and both LOLMU-002 and LOLMU-003 had a few survivors to cycloxydim, to dose rates below field rate, they were still well controlled at recommended field rate, with shoot dry weight reduced by >88%. For ALS-inhibiting herbicides, GR50 values of LOLMU-001 to LOLMU-004 ranged between 1.0 and 1.3 times the recommended field rate of mesosulfuron + iodosulfuron, whereas\their GR50 values were slightly below recommended field rate of pyroxsulam (0.6–0.8) (Figure 4D,E). The RF ranged between 11.9 and 14.6 for mesosulfuron + iodosulfuron, and between 2.3 and 3.1 for pyroxsulam (Table 5) indicating ALS resistance.
Table 5.
GR50 values (standard errors in parenthesis) from shoot dry weight response to three ACCase inhibitors pinoxaden, propaquizafop and cycloxydim and two ALS inhibitors mesosulfuron + iodosulfuron and pyroxsulam for susceptible (SORG-002) and resistant (LOLMU-001 to LOLMU-004) populations of L. multiflorum. Resistance factor (RF) was calculated as ratio between the GR50 of resistant population by the GR50 of the susceptible population. No known ACCase mutations detected. ALS mutations identified at Pro-197 position only. Eight plants were analyzed per population. Number of homozygous and heterozygous L. multiflorum plants for ALS mutation is reported in the bracket.
3.3. Analysis of Target-Site Resistance Mutations
Pyrosequencing analysis discovered one of the five mutated positions, i.e., Ile-1781 in the ACCase gene, in all four suspect resistant (ALOMY-001 to ALOMY-004) populations of A. myosuroides (Table 4). In ALOMY-001, six individuals were homozygous for the Ile-1781-Leu substitutions, and two were heterozygous Ile-1781-Ile/Leu. More homozygous mutant plants might have conferred stronger resistance to recommended field rate of propaquizafop and cycloxydim in this population. In ALOMY-002 to ALOMY-004, variability of the ratio between homo/heterozygous, or only heterozygous mutant individuals might have conferred a relatively lower level of resistance to recommended field rate of both ACCase-inhibiting herbicides. Additionally, ALOMY-002 to ALOMY-004 also had the Pro-197 mutation in the ALS gene (Table 4). Heterozygous mutant plants (Pro-197-Pro/Thr in ALOMY-002 and ALOMY-004, or combination of Pro-197-Pro/Thr and Pro-197-Pro/Ser in ALOMY-003) were predominant and only one from ALOMY-002 was homozygous (Pro-197-Thr), all of which were sufficient to confer mesosulfuron + iodosulfuron failure at recommended field rate. ALOMY-001 had no mutations at Pro-197 or Trp-574 and herbicide screening also confirmed similar sensitivity as S-006 to mesosulfuron + iodosulfuron.
For L. multiflorum, different substitutions for the Pro-197 position in the ALS gene were identified in suspect populations LOLMU-001, LOLMU-002 and LOLMU-004 only (Table 5). Predominantly homozygous mutant plants (Pro-197-Gln in LOLMU-001 and LOLMU-002 or Pro-197-Leu in LOLMU-004) conferred resistance to recommended field rate of mesosulfuron + iodosulfuron and pyroxsulam. However, the resistance levels were stronger with mesosulfuron + iodosulfuron, compared with pyroxsulam. Despite no individuals possessing any of the known ACCase gene mutations in LOLMU-001 to LOLMU-004, it is interesting that LOLMU-002 was able to survive pinoxaden and propaquizafop, and LOLMU-003 to pinoxaden only, among these populations. In addition, phenotypic resistance to both ALS-inhibiting herbicides was noted in LOLMU-003, but without any known ALS gene mutations. These results indicate the likely presence of NTSR mechanisms for ACCase and/or ALS resistance in LOLMU-002 and LOLMU-003. The susceptible A. myosuroides (S-006) and L. multiflorum (SORG-006) populations had no mutations in any of the known positions in the ACCase and ALS genes.
4. Discussion
This study is the first to identify and characterize resistance to different ACCase- and ALS-inhibiting herbicides in populations of four A. myosuroides (ALOMY-001 to ALOMY-004) and four L. multiflorum (LOLMU-001 to LOLMU-004) from Ireland. For A. myosuroides, the homo/heterozygous combinations or only heterozygous Ile-1781 mutation was associated with resistance to ACCase-FOP-propaquizafop as well as to ACCase-DIM-cycloxydim in all four ALOMY populations, and the independent Pro-197 mutation (mostly in heterozygous status) was associated with resistance to the ALS-SU-mesosulfuron + iodosulfuron, additionally in ALOMY-002, ALOMY-003, and ALOMY-004. For L. multiflorum, all four LOLMU populations were resistant to ALS-SU-mesosulfuron + iodosulfuron as well as to ALS-TP-pyroxsulam, but the Pro-197 mutation (mostly in homozygous status) was only identified in LOLMU-001, LOLMU-002 and LOLMU-004. Despite no known ACCase mutations in these LOLMU populations, LOLMU-002 was able to survive pinoxaden and propaquizafop application and LOLMU-003 was able to survive pinoxaden, but both were cycloxydim-susceptible. These results suggest the presence of NTSR mechanisms for the difference between the genetic and phenotypic data in these two populations. In general, as both homo/heterozygous mutant plants carrying the Ile-1781 or Pro-197 had their growth impacted by higher herbicide dose rate (Figure 3 and Figure 4), and as not all the ACCase/ALS survivors especially in ALOMY-003 and ALOMY-004, or in LOLMU-002 to LOLMU-004 had substitutions (Table 4 and Table 5), the resistance recorded was classified as low to moderate in ALOMY and LOLMU.
Diverse mutated positions of ACCase (Ile-1781, Trp-1999, Trp-2027, Ile-2041, Asp-2078, Cys-2088, Gly-2096) and ALS (Pro-197, Asp-376, Trp-574, Ala-122, Ala-205 and Ser-653) conferring TSR have been identified in grass weed species [13,27,28,29]. Nevertheless, the Ile-1781 and Pro-197 mutations documented in this study are by far the most common mutation in the UK and in Europe to confer distinct resistance spectrum to different ACCase- and ALS-inhibiting herbicides in A. myosuroides and/or L. multiflorum [6,11,30,31,32,33]. Multiple resistance mechanisms via, TSR and/or NTSR to both MOA within a single field population of A. myosuroides and Lolium spp., and in predominantly self-pollinating Avena spp., and Anisantha and Bromus spp. are quite frequent worldwide [11,12,17,34,35,36,37,38,39]. Especially in the UK, NTSR to ACCase-inhibiting herbicides has been consistently more frequent than TSR mechanisms in L. multiflorum [11,40]. This may also be the case with LOLMU-002 and LOLMU-003 in this study, as supported by our preliminary results with pendimethalin assays and glutathione transferase phi (F) class 1 (GSTF1) protein, a reliable biomarker of NTSR in A. myosuroides (Onkokesung et al. unpublished).
For A. myosuroides, with the exception of ALOMY-001 to ALOMY-004, the remaining eight populations (including sensitive S-006) collected from this nationwide survey, were found to be completely sensitive to all three A. myosuroides-active herbicides applied at recommended field rate (0% survival and shoot dry weight reduction of >90%). Moreover, our records suggest that most surveyed growers had A. myosuroides incidence only in recent years, and strongly believed that the seeds have entered into their farm through different routes of contamination (e.g., via, second generation (C-2) seed imports from the UK, machinery moving from farm to farm, organic manure imports from other farms, or screenings from local mills) (Vijaya Bhaskar et al. unpublished). The source of A. myosuroides infestations (either indigenous or imported from the UK) will have implications for the adoption of weed spread prevention strategies. In general, for TSR to be noticed, such resistance types must have already been present at relatively high frequency [31], and growers have recorded weed control difficulties in the field. Herbicide cross-resistance to the same MOA within a single field such as in ALOMY-001, which was ACCase-resistant only, but was well controlled by mesosulfuron + iodosulfuron; or in LOLMU-001 and LOLMU-004, which were ALS-resistant only, but were well controlled by pinoxaden, propaquizafop and cycloxydim, demonstrates that growers can use alternative herbicide MOA as part of a strategy to eliminate or control these resistant populations. The current advice in the UK is to combine pre-emergence herbicides (and, if possible, use non-ACCase and non-ALS herbicides e.g., propyzamide in winter oilseed rape), with cultural and non-chemical IWM such as delayed autumn drilling, spring cropping, higher seed rates, ploughing, diverse rotations and hand rogueing to manage the soil seedbank of populations that were resistant to single site post-emergent herbicides [31].
However, future A. myosuroides control is threatened, where herbicide cross-resistance to the same, and different, MOA or multiple resistance mechanisms occur together, such as ALOMY-002 to ALOMY-004 which exhibit TSR to both ACCase- and ALS-inhibiting herbicides. Similar observations were noted in LOLMU-002 and LOLMU-003, which exhibited combined ALS TSR and ACCase NTSR resistance or ALS/ACCase NTSR resistance alone. It is interesting that ACCase-DIM applied at the recommended field rate was highly effective on both these populations. A similar observation was also reported by Cocker et al. [40], that the occurrence of resistance to FOP herbicides in L. multiflorum does not automatically confer cross-resistance to DIM even though both herbicides are from the ACCase group, confirming the complexity in predicting resistance types. Nevertheless, the use of cycloxydim should not be considered as a sustainable solution, taking into account the resistance mechanisms and the high potential of L. multiflorum for spreading herbicide-resistance traits [7,9,41]. It is interesting that clethodim (rarely used in Ireland to date [22]), which is also an ACCase-DIM herbicide, may still offer effective control of resistant populations carrying complex multiple resistance mechanisms [42], and may provide an opportunity to growers to use this post-emergent herbicide in sequence with propyzamide. However, Yu et al. [43] showed that, with Lolium strains, a number of different ACCase TSR mutations, including Ile-1781, confer clethodim resistance. In practical terms, rotational restriction of winter oilseed rape (recommended one crop of oilseed rape in five years), restricts the effectiveness of these alternative herbicides.
In general, long-term field experiments in the UK have shown that herbicide mixture, rotation or even non-chemical/cultural IWM will have only very little impact on populations with evolved resistance via multiple resistance mechanisms [17,31]. This is because evidence indicates that, at least in the case of TSR, once it is present in a population it does not disappear, even if the selection pressure is removed [44]. Economic models have estimated annual costs of around €464 million for managing herbicide-resistant A. myosuroides in the UK [45]. Therefore, it is critical to prevent the development of resistance, both TSR and NTSR, rather than trying to manage resistance once it has appeared in a population [46]. As Ireland is at a different point in resistance evolution to neighboring countries, due to its evolving cropping systems from a mixed farming base, lessons must be learned from other countries’ experience. Immediate adoption of a zero-tolerance approach and extreme IWM measures, such as whole cropping (cutting, baling and removing the affected straw), use of grass leys or fallows to prevent a significant soil weed seed bank should be considered for challenging weeds.
5. Conclusions
Using glasshouse experiments and molecular diagnostics to detect TSR mechanism, we confirmed the first cases of cross- and multiple resistance to ACCase- and ALS-inhibiting herbicides in four suspected resistant populations of A. myosuroides (ALOMY-001 to ALOMY-004) and L. multiflorum (LOLMU-001 to LOLMU-004) from Ireland. In A. myosuroides, the homo/heterozygous or only heterozygous Ile-1781 ACCase and Pro-197 ALS TSR mutations was found to be associated with ACCase/ALS resistance. While in L. multiflorum, even though mostly homozygous Pro-197 ALS TSR mutation was found to be associated with ALS resistance, there were ACCase/ALS-resistant populations without TSR, suggesting the possibility of NTSR. Mutation rate, level of heterozygosity and herbicide rates were critical for resistance in ALOMY and LOLMU, being classified as low to moderate. Overall, our findings, by definitively determining the resistance status of A. myosuroides and L. multiflorum, will increase awareness among researchers, agronomists and growers about the need to reduce the reliance on ACCase and ALS herbicides. These results will stress the importance of integrating cultural/non-chemical IWM with farm biosecurity measures, such as machinery hygiene, avoiding straw and manure imports, using C-1 certified seeds, as prevention and control strategies. The current study confirms the need for resistance monitoring in A. myosuroides and L. multiflorum to ensure that resistance problems do not increase in this region. Knowledge of resistance levels and cross-resistance among the main herbicide types is a valuable information tool to inform intervention strategies to slow or prevent TSR and NTSR resistance evolution in important grass weeds.
Author Contributions
Conceptualization, V.B.A.V., P.D.F., S.B. and M.H.; methodology and investigation, V.B.A.V., D.S., J.S. and S.K.C.; Data curation, V.B.A.V.; formal analysis, V.B.A.V.; writing—original draft, V.B.A.V.; supervision, S.B., P.D.F., S.K.C. and M.H.; writing—reviewing and editing, V.B.A.V., S.K.C., P.D.F., S.B.; funding acquisition, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded through the project ECT (Enable Conservation Tillage, Grant No: LLOC1079), which is a European Innovation Partnership (EIP) funded by the Department of Agriculture, Food, and the Marine (DAFM) under the Rural Development Programme 2014–2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Any further inquiries regarding data generated as part of this publication can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Moss, S.R.; Perryman, S.A.M.; Tatnell, L.V. Managing herbicide resistant black-grass (Alopecurus myosuroides): Theory and practice. Weed Techn. 2007, 21, 300–309. [Google Scholar] [CrossRef]
- Lutman, P.J.W.; Moss, S.R.; Cook, S.K.; Welham, S.J. A review of the effects of crop agronomy on the management of Alopecurus myosuroides. Weed Res. 2013, 53, 299–313. [Google Scholar] [CrossRef]
- Moss, S.R. Managing Herbicide-Resistant Rye-Grass: 42nd Annual Review of Weed Control; British Crop Protection Council (BCPC): Peterborough, UK, 2005; pp. 40–47. [Google Scholar]
- Moss, S.R. Black-grass (Alopecurus myosuroides): Why has this weed become such a problem in Western Europe and what are the solutions? Outlooks Pest Manag. 2017, 10, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Reverte, R. Understanding and Combating the Threat Posed by Lolium multiflorum as a Weed of Arable Crops. Ph.D. Thesis, School of Plant Sciences, University of Reading, Reading, UK, 2010. [Google Scholar]
- Délye, C.; Michel, S.; Bérard, A.; Chauvel, B.; Brunel, D.; Guillemin, J.P.; Dessaint, F.; Le Corre, V. Geographical variation in resistance to acetyl-coenzyme A carboxylase-inhibiting herbicides across the range of the arable weed Alopecurus myosuroides (black-grass). New Phytol. 2010, 186, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.S.; Welles, S.R.; Silvera, K.; Heap, I.M.; Heredia, S.M.; Martinez-Berdeja, A.; Palenscar, K.T.; Sweet, L.C.; Ellstrand, N.C. Taxonomic and life history bias in herbicide resistant weeds: Implications for deployment of resistant crops. PLoS ONE 2013, 8, e71916. [Google Scholar] [CrossRef] [Green Version]
- Moss, S.R. Herbicide cross-resistance in slender foxtail (Alopecurus myosuroides). Weed Sci. 1990, 38, 492–496. [Google Scholar] [CrossRef]
- Liu, M.; Hulting, A.G.; Mallory-Smith, C.A. Characterisation of multiple-herbicide-resistant Italian ryegrass (Lolium perenne spp. multiflorum). Pest Manag. Sci. 2014, 70, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Heap, I.M. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 15 May 2021).
- Hull, R.; Tatnell, L.V.; Cook, S.K.; Beffa, R.; Moss, S.R. Current status of herbicide-resistance weeds in the UK. Asp. Appl. Biol. 2014, 127, 261–271. [Google Scholar]
- Scarabel, L.; Panozzo, S.; Loddo, D.; Mathiassen, S.; Kristensen, M.; Kudsk, P.; Gitsopoulous, T.; Travlos, I.; Tani, E.; Chachalis, D.; et al. Diversified resistance mechanisms in multi-resistant Lolium spp. in three European countries. Front. Plant Sci. 2020, 11, 608845. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Délye, C.; Menchari, Y.; Guillemin, J.P.; Matéjicek, A.; Michel, S.; Camilleri, C.; Chauvel, B. Status of black-grass (Alopecurus myosuroides) resistance to acetyl-coenzyme A carboxylase inhibitors in France. Weed Res. 2007, 47, 95–105. [Google Scholar] [CrossRef]
- Marshall, R.; Moss, S.R. Characterisation and molecular basis of ALS inhibitor resistance in the grass weed Alopecurus myosuroides. Weed Res. 2008, 48, 439–447. [Google Scholar] [CrossRef]
- Hicks, H.L.; Comont, D.; Coutts, S.R.; Crook, L.; Hull, R.; Norris, K.; Neve, P.; Childs, D.Z.; Freckleton, R.P. The factors driving evolved herbicide resistance at a national scale. Nat. Ecol. Evol. 2018, 2, 520–536. [Google Scholar] [CrossRef] [Green Version]
- Scarabel, L.; Panozzo, S.; Varotto, S.; Sattin, M. Allelic variation of the ACCase gene and response to ACCase-inhibiting herbicides in pinoxaden-resistant Lolium spp. Pest Manag. Sci. 2011, 67, 932–941. [Google Scholar] [CrossRef]
- CSO. Structure of Farming in Ireland 2016, Central Statistics Office (CSO). Available online: https://www.cso.ie/en/releasesandpublications/ep/p-fss/farmstructuresurvey2016/ (accessed on 18 June 2021).
- Forristal, D.; Grant, J. The impact of break-crop and cereal rotations on crop performance and profit margin: Making Crop rotations fit for the future. Asp. Appl. Biol. 2011, 113, 29–36. Available online: https://www.aab.org.uk/aspects-of-applied-biology/ (accessed on 21 June 2021).
- BSBI. Botanical Society of Britain and Ireland (BSBI); BSBI: Durham, UK, 2021. [Google Scholar]
- DAFM. Pesticide Usage in Ireland, Arable Crops Survey Report 2016, Pesticide Registration and Control Divisions (PRCD) of the Department of Agriculture; Food and the Marine (DAFM), Co.: Kildare, Ireland, 2016. Available online: https://www.pcs.agriculture.gov.ie/media/pesticides/content/sud/pesticidestatistics/ArableReport2016Final100620.pdf (accessed on 21 June 2021).
- Alwarnaidu Vijayarajan, V.B.; Forristal, D.P.; Cook, S.K.; Staples, J.; Schilder, D.; Hennessy, M.; Barth, S. First report on assessing the severity of herbicide resistance to ACCase inhibitors pinoxaden, propaquizafop and cycloxydim in six Avena fatua populations in Ireland. Agronomy 2020, 10, 1362. [Google Scholar] [CrossRef]
- Hess, M.; Barralis, G.; Bleiholder, H.; Buhrs, L.; Eggers, T.H.; Hack, H.; Strauss, R. Use of the extended BBCH scale-general for the descriptions of the growth stages of mono- and dicotyledonous weed species. Weed Res. 1997, 37, 433–441. [Google Scholar] [CrossRef]
- Keshtkar, E.; Mathiassen, S.K.; Moss, S.R.; Kudsk, P. Resistance profile of herbicide-resistant Alopecurus myosuroides (black-grass) populations in Denmark. Crop Prot. 2015, 69, 83–89. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Tranel, P.J.; Wright, T.R.; Heap, I.M. Mutations in Herbicide-Resistant Weeds to ALS Inhibitors. Available online: http://www.weedscience.com (accessed on 18 May 2021).
- Yu, Q.; Han, H.; Powles, S.B. Mutations of the ALS gene endowing resistance to ALS-inhibiting herbicides in Lolium rigidum populations. Pest Manag. Sci. 2008, 64, 1229–1236. [Google Scholar] [CrossRef]
- Henriet, F.; Marechal, P.Y. Black-grass resistance to herbicides: Three years of monitoring in Belgium. Commun. Agric. Appl. Biol. Sci. 2009, 74, 471–478. [Google Scholar]
- Marshall, R.; Hanley, S.J.; Hull, R.; Moss, S.R. The presence of two different target-site resistance mechanisms in individual plants of Alopecurus myosuroides Huds, identified using a quick molecular test for the characterization of six and seven ACCase SNPs. Pest Manag. Sci. 2013, 69, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Torra, J.; Montull, J.M.; Taberner, A.; Onkokesung, N.; Boonham, N.; Edwards, R. Target-site and non-target-site resistance mechanisms confer multiple and cross- resistance to ALS and ACCase inhibiting herbicides in Lolium rigidum From Spain. Front. Plant Sci. 2021, 12, 625138. [Google Scholar] [CrossRef] [PubMed]
- Collavo, A.; Sattin, M. First glyphosate-resistant Lolium spp. biotypes found in a European annual arable cropping systems also affected by ACCase and ALS resistance. Weed Res. 2014, 54, 325–334. [Google Scholar] [CrossRef]
- Tardif, F.J.; Powles, S.B. Herbicide multiple-resistance in a Lolium rigidum biotype is endowed by multiple mechanisms-Isolation of a subset with resistant acetyl-CoA carboxylase. Physiol. Plant 1994, 91, 488–494. [Google Scholar] [CrossRef]
- Baucom, R.S. Evolutionary and ecological insights from herbicide resistant weeds: What have we learned about plant adaptation, and what is left to uncover? New Phytol. 2019, 223, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.S.; Tranel, P.J.; Stewart, C.N. Non-target-site herbicide resistance: A family business. Trends Plant Sci. 2007, 12, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Cocker, K.M.; Coleman, J.O.D.; Blair, A.M.; Clarke, J.H.; Moss, S.R. Biochemical mechanisms of cross-resistance to aryloxyphenoxypropionate and cyclohexanediones herbicides in populations of Avena spp. (wild oats). Weed Res. 2000, 40, 323–334. [Google Scholar] [CrossRef]
- Owen, M.; Goggin, D.E.; Powles, S.B. Non-target-site-based resistance to ALS-inhibiting herbicides in six Bromus rigidus populations from Western Australia cropping fields. Pest Manag. Sci. 2012, 68, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.; Onkokesung, N.; Brazier-Hicks, M.; Edwards, R.; Moss, S. Detection and characterization of resistance to acetolactate synthase inhibiting herbicides in Anisantha and Bromus species in the United Kingdom. Pest Manag. Sci. 2020, 76, 2473–2482. [Google Scholar] [CrossRef] [PubMed]
- Cocker, K.M.; Northcroft, D.S.; Coleman, J.O.D.; Moss, S.R. Resistance to ACCase-inhibiting herbicides and isoproturon in UK populations of Lolium multiflorum: Mechanisms of resistance and implications for control. Pest Manag. Sci. 2001, 57, 587–597. [Google Scholar] [CrossRef]
- Petit, C.; Duhieu, B.; Boucansaud, K.; Délye, C. Complex genetic control of non-target-site based resistance to herbicides inhibiting acetyl-coenzyme A carboxylase and acetolactate synthase in Alopecurus myosuroides Huds. Plant Sci. 2010, 178, 501–509. [Google Scholar] [CrossRef]
- Moss, S.R.; Riches, C.; Stormonth, D. Clethodim: It’s potential to combat herbicide-resistant Alopecurus myosuroides (black-grass). Asp. Appl. Biol. 2012, 117, 39–45. [Google Scholar]
- Yu, Q.; Collavo, A.; Zheng, M.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of acetyl-coenzyme a carboxylase mutations in resistant Lolium populations: Evaluation using clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauvel, B.; Guillemin, J.P.; Colbach, N. Evolution of a herbicide-resistant population of Alopecurus myosuroides Huds in a long-term cropping system experiment. Crop Prot. 2009, 28, 343–349. [Google Scholar] [CrossRef]
- Varah, A.; Ahodo, K.; Coutts, S.R.; Hicks, H.L.; Comont, D.; Crook, L.; Hull, R.; Neve, P.; Childs, D.Z.; Freckleton, R.; et al. The cost of human-induced evolution in an agricultural system. Nat. Sustain. 2020, 3, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.R. Integrated weed management (IWM): Why are farmers reluctant to adopt non-chemical alternatives to herbicides? Pest Manag. Sci. 2019, 75, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).