
Spatial and Temporal Variation in the Aphid–Parasitoid Interaction under Different Climates

 and
and
Abstract
:1. Introduction
2. Materials and Methods
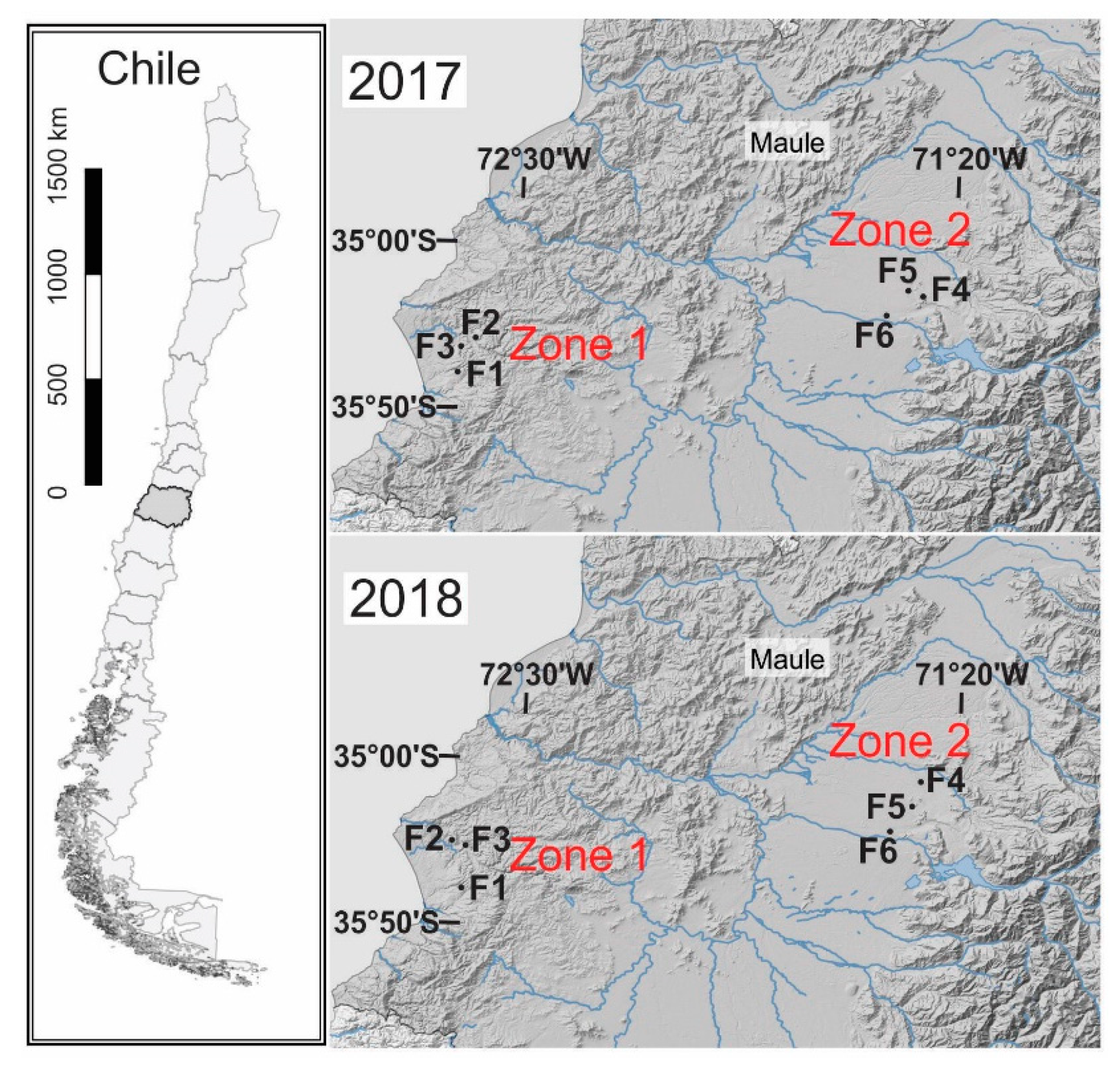
2.1. Sampling Sites and Climates
2.2. Aphids and Parasitoids Samplings
2.3. Aphid Bacterial Endosymbionts
2.4. Data Analysis
3. Results
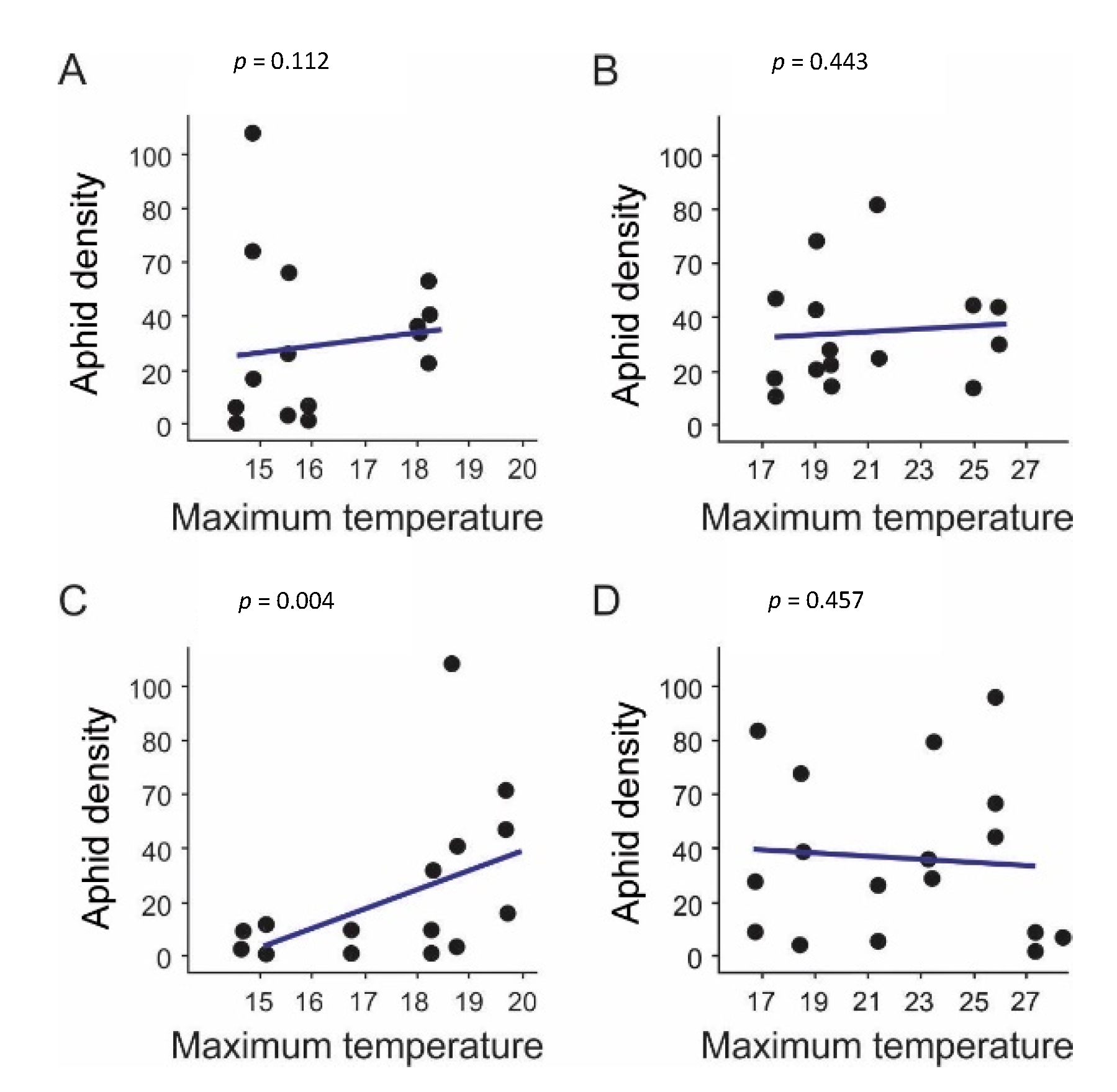
3.1. Registered Temperatures in the Field
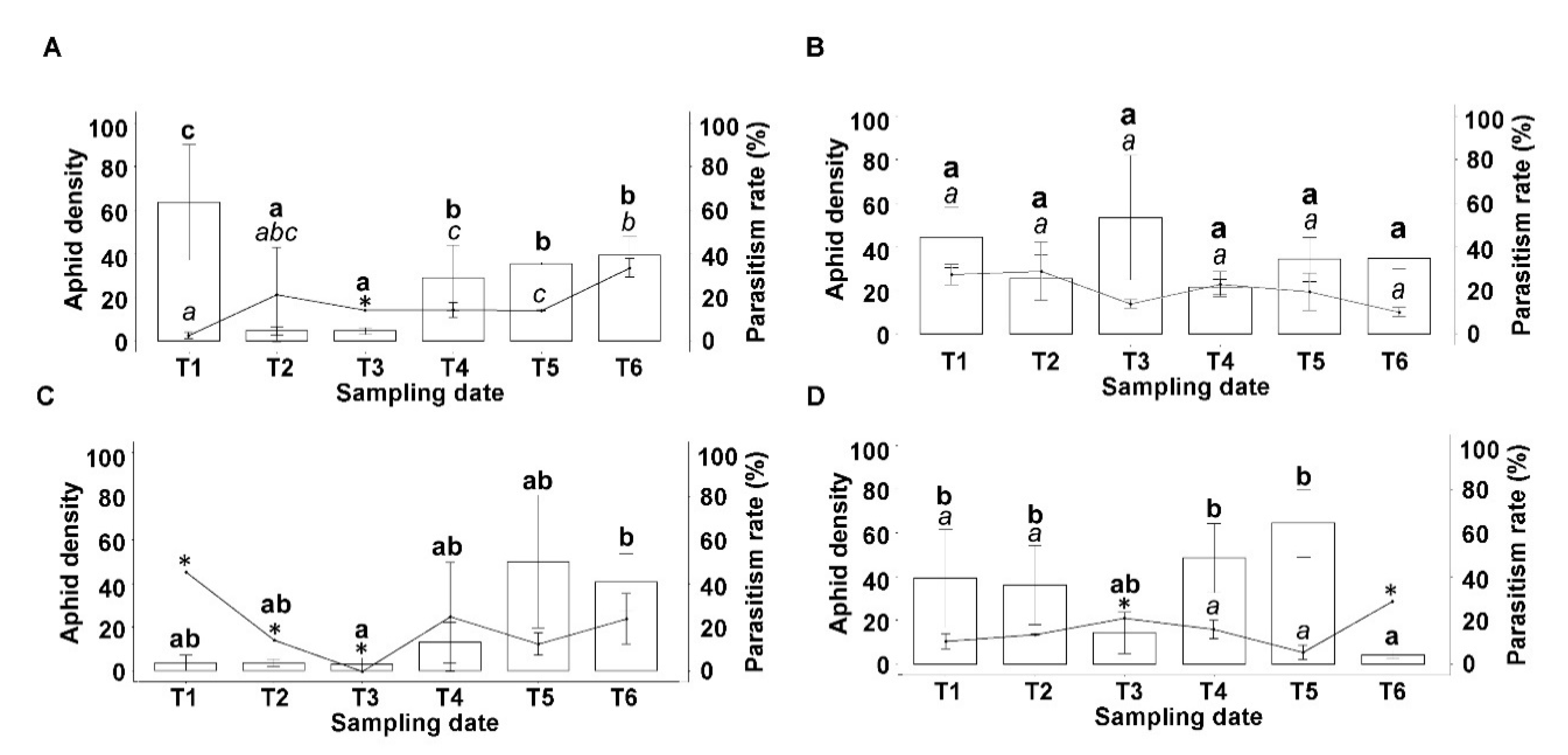
3.2. Sitobion avenae Density
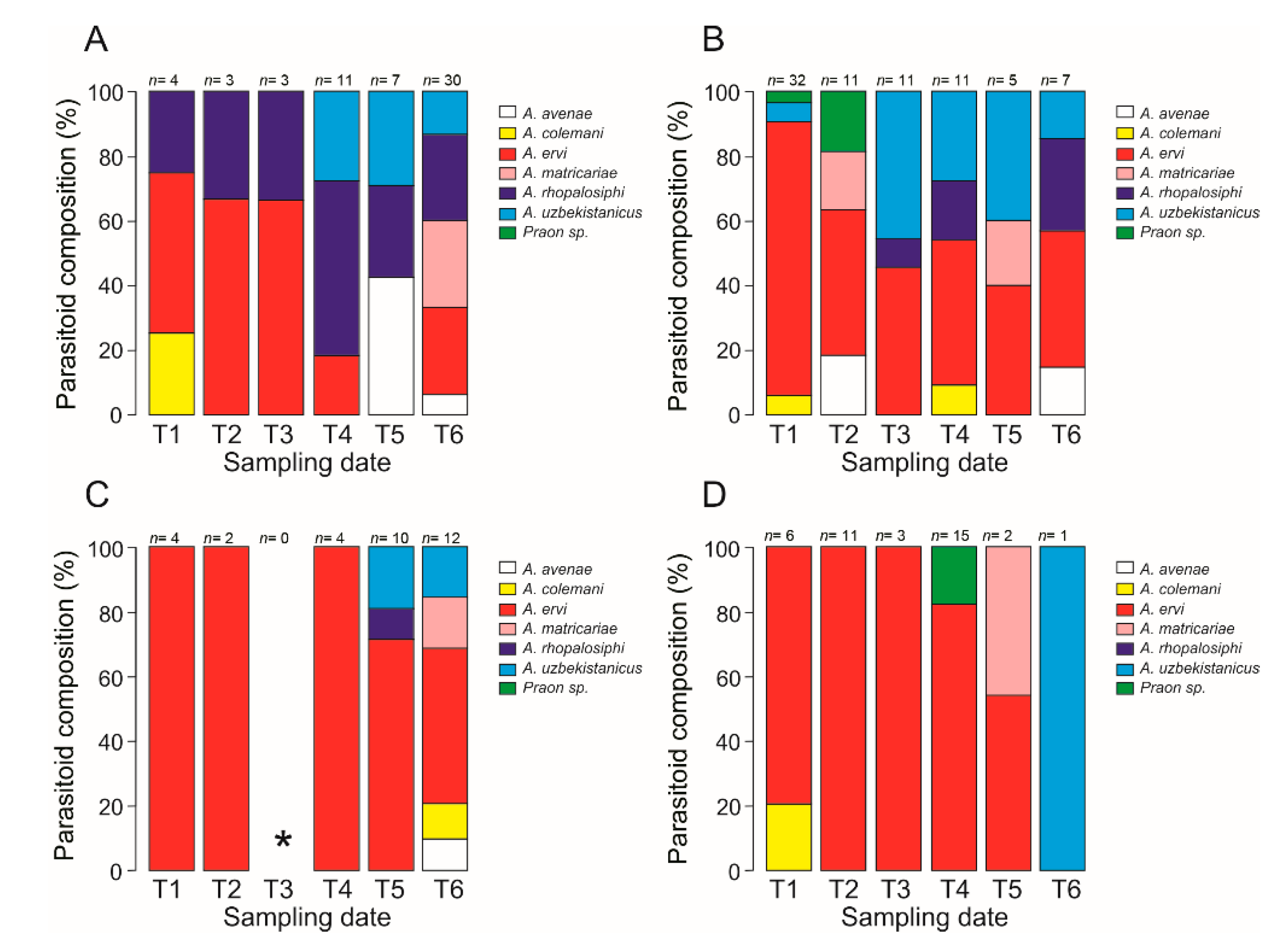
3.3. Parasitoid Composition
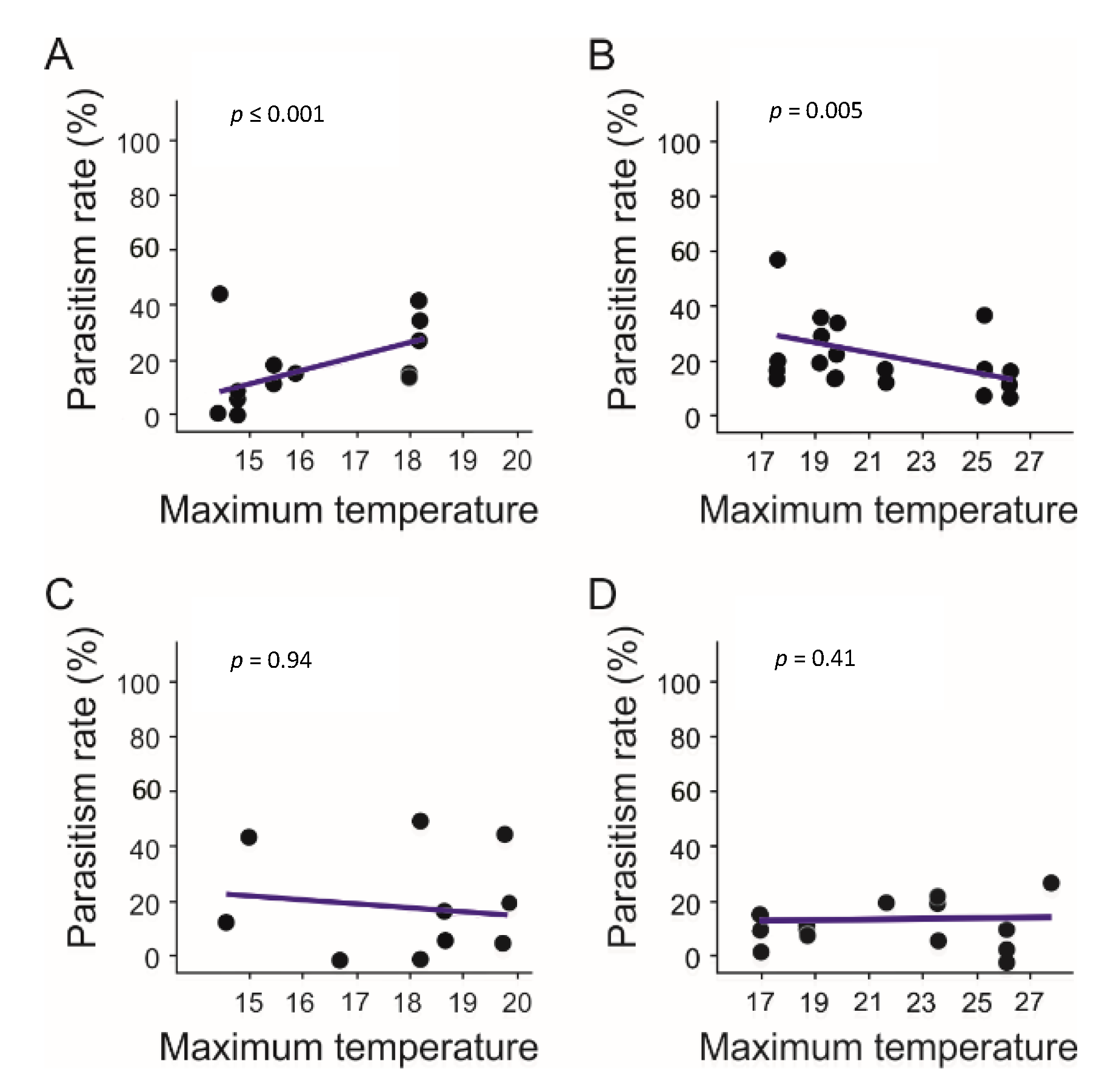
3.4. Parasitism Rate
3.5. Composition of Facultative Endosymbionts
4. Discussion
4.1. Factors Shaping Spatial and Temporal Sitobion avenae Populations
4.2. Parasitoid Assemblage
4.3. Spatial and Temporal Changes in Parasitism Rate
4.4. Facultative Endosymbionts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, G.C.; Valin, H.; Sands, R.D.; Havlík, P.; Ahammad, H.; Deryng, D.; Elliott, J.; Fujimori, S.; Hasegaw, T.; Heyhoe, E.; et al. Climate change effects on agriculture: Economic responses to biophysical shocks. Proc. Natl. Acad. Sci. USA 2014, 111, 3274–3279. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.M.; Vermeulen, S.A.; Aggarwal, P.K.; Corner-Dolloff, C.; Girvetz, E.; Loboguerrero, A.M.; Ramirez-Villegas, J.; Rosentock, T.; Sebastian, L.; Thornton, P.K.; et al. Reducing risks to food security from climate change. Glob. Food Sec. 2016, 11, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Blumenthal, D.M.; Runion, G.B.; Hunt, E.R.; Diaz-Soltero, H. Invasive species and climate change: An agronomic perspective. Clim. Chang. 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Bebber, D.P. Range-Expanding Pests and Pathogens in a Warming World. Annu. Rev. Phytopathol. 2015, 53, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Furlong, M.J.; Zalucki, M.P. Climate change and biological control: The consequences of increasing temperatures on host–parasitoid interactions. Curr. Opin. Insect Sci. 2017, 20, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.C.; Rispe, C.; Sunnucks, P. Ecology and evolution of sex in aphids. Trends Ecol. Evol. 2002, 17, 34–39. [Google Scholar] [CrossRef]
- Figueroa, C.C.; Fuentes-Contreras, E.; Molina-Montenegro, M.A.; Ramírez, C.C. Biological and genetic features of introduced aphid populations in agroecosystems. Curr. Opin. Insect Sci. 2018, 26, 63–68. [Google Scholar] [CrossRef]
- Simon, J.C.; Peccoud, J. Rapid evolution of aphid pests in agricultural environments. Curr. Opin. Insect Sci. 2018, 26, 17–24. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; John Wiley & Sons Ltd.: Chichester, UK, 2000. [Google Scholar]
- Dedryver, C.A.; Le Ralec, A.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Starý, P. The Aphidiidae of Chile (Hymenoptera, Ichneumonoidea, Aphidiidae). Dtsch. Entomol. Z. 1995, 42, 113–138. [Google Scholar] [CrossRef]
- Figueroa, C.C.; Simon, J.-C.; Le Gallic, J.-F.; Prunier-Leterme, N.; Briones, L.M.; Dedryver, C.; Niemeyer, H.M. Genetic structure and clonal diversity of an introduced pest in Chile, the cereal aphid Sitobion avenae. Heredity 2005, 95, 24–33. [Google Scholar] [CrossRef]
- Starý, P.; Gerding, M.; Norambuena, H.; Remaudière, G. Environmental-Research on aphid parasitoid biocontrol agents in Chile (Hym, Aphidiidae, Hom, Aphidoidea). J. Appl. Entomol. 1993, 115, 292–306. [Google Scholar] [CrossRef]
- Zepeda-Paulo, F.; Villegas, C.; Lavandero, B. Host genotype-endosymbiont associations and their relationship with aphid parasitism at the field level. Ecol. Entomol. 2017, 42, 86–95. [Google Scholar] [CrossRef]
- Zuñiga, E.; Van den Boch, R.; Drea, J.; Gruber, F. The biological control project against the cereal aphids (Hom., Aphididae) in Chile. II. Exploration, importation and quarantine of predator and parasitoid species. Agric. Téc. 1986, 46, 479–487. [Google Scholar]
- Welch, K.D.; Harwood, J.D. Temporal dynamics of natural enemy-pest interactions in a changing environment. Biol. Control 2014, 75, 18–27. [Google Scholar] [CrossRef]
- Heyworth, E.R.; Smee, M.R.; Ferrari, J. Aphid Facultative Symbionts Aid Recovery of Their Obligate Symbiont and Their Host After Heat Stress. Front. Ecol. Evol. 2020, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Montllor, C.B.; Maxmen, A.; Purcell, A.H. Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol. Entomol. 2002, 27, 189–195. [Google Scholar] [CrossRef]
- Heyworth, E.R.; Ferrari, J. Heat stress affects facultative symbiont-mediated protection from a parasitoid wasp. PLoS ONE 2016, 11, e0167180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doremus, M.R.; Smith, A.H.; Kim, K.L.; Holder, A.J.; Russell, J.A.; Oliver, K.M. Breakdown of a defensive symbiosis, but not endogenous defences, at elevated temperatures. Mol. Ecol. 2017, 27, 2138–2151. [Google Scholar] [CrossRef]
- Liu, X.D.; Lei, H.X.; Chen, F.F. Infection pattern and negative effects of a facultative endosymbiont on its insect host are environment-dependent. Sci. Rep. 2019, 9, 4013. [Google Scholar] [CrossRef]
- Bensadia, F.; Boudreault, S.; Guay, J.-F.; Michaud, D.; Cloutier, C. Aphid clonal resistance to a parasitoid fails under heat stress. J. Insect Physiol. 2006, 52, 146–157. [Google Scholar] [CrossRef]
- McLean, A.H.C.; Parker, B.J.; Hrcek, J.; Henry, L.M.; Godfray, H.C.J. Insect symbionts in food webs. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150325. [Google Scholar] [CrossRef] [Green Version]
- Thierry, M.; Hrček, J.; Lewis, O.T. Mechanisms structuring host–parasitoid networks in a global warming context: A review. Ecol. Entomol. 2019, 44, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Bannerman, J.A.; Roitberg, B.D. Impact of extreme and fluctuating temperatures on aphid-parasitoid dynamics. Oikos 2014, 123, 89–98. [Google Scholar] [CrossRef]
- Ma, G.; Hoffmann, A.A.; Ma, C.S. Daily temperature extremes play an important role in predicting thermal effects. J. Exp. Biol. 2015, 218, 2289–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Rudolf, V.H.W.; Ma, C.S. Extreme temperature events alter demographic rates, relative fitness, and community structure. Glob. Chang. Biol. 2015, 21, 1794–1808. [Google Scholar] [CrossRef]
- Falvey, M.; Garreaud, R.D. Regional cooling in a warming world: Recent temperature trends in the southeast Pacific and along the west coast of subtropical South America (1979–2006). J. Geophys. Res. Atmos. 2009, 114, D04102. [Google Scholar] [CrossRef]
- Piticar, A. Changes in agro-climatic indices related to temperature in Central Chile. Int. J. Biometeorol. 2019, 63, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: Knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric. Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef] [Green Version]
- INIA Agrometereología. Available online: https://agrometeorologia.cl/# (accessed on 3 March 2020).
- Araya-Osses, D.; Casanueva, A.; Román-Figueroa, C.; Uribe, J.M.; Paneque, M. Climate change projections of temperature and precipitation in Chile based on statistical downscaling. Clim. Dyn. 2020, 54, 4309–4330. [Google Scholar] [CrossRef]
- Gagic, V.; Hänke, S.; Thies, C.; Scherber, C.; Tomanovic, Z.; Tscharntke, T. Agricultural intensification and cereal aphid-parasitoid-hyperparasitoid food webs: Network complexity, temporal variability and parasitism rates. Oecologia 2012, 170, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xu, L.; Wu, Y.K.; Wang, Q.; Yao, Z.W.; Žikić, V.; Tomanović, Ž.; Ferrer-Suay, M.; Selfa, J.; Pujade-Villar, J.; et al. Species composition and seasonal dynamics of aphid parasitoids and hyperparasitoids in wheat fields in northern China. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sunnucks, P.; England, P.R.; Taylor, A.C.; Hales, D.F. Microsatellite and chromosome evolution of parthenogenetic sitobion aphids in Australia. Genetics 1996, 144, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Peccoud, J.; Bonhomme, J.; Mahéo, F.; de la Huerta, M.; Cosson, O.; Simon, J.-C. Inheritance patterns of secondary symbionts during sexual reproduction of pea aphid biotypes. Insect Sci. 2013, 21, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, D.A.; Zepeda-Paulo, F.; Ramírez, C.C.; Lavandero, B.; Figueroa, C.C. Diversity, frequency, and geographic distribution of facultative bacterial endosymbionts in introduced aphid pests. Insect Sci. 2017, 24, 511–521. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffs, C.T.; Lewis, O.T. Effects of climate warming on host-parasitoid interactions. Ecol. Entomol. 2013, 38, 209–218. [Google Scholar] [CrossRef]
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a Warming Climate: Insect Responses to Extreme High Temperatures. Annu. Rev. Entomol. 2021, 66, 163–184. [Google Scholar] [CrossRef]
- Jeffs, C.T.; Leather, S.R. Effects of extreme, fluctuating temperature events on life history traits of the grain aphid, Sitobion avenae. Entomol. Exp. Appl. 2014, 150, 240–249. [Google Scholar] [CrossRef]
- Han, P.; Becker, C.; Sentis, A.; Rostás, M.; Desneux, N.; Lavoir, A.V. Global change-driven modulation of bottom–up forces and cascading effects on biocontrol services. Curr. Opin. Insect Sci. 2019, 35, 27–33. [Google Scholar] [CrossRef]
- Asin, L.; Pons, X. Effect of high temperature on the growth and reproduction of corn aphids (homoptera: Aphididae) and implications for their population dynamics on the Northeastern Iberian Peninsula. Environ. Entomol. 2001, 30, 1127–1134. [Google Scholar] [CrossRef]
- Sentis, A.; Hemptinne, J.L.; Brodeur, J. Effects of simulated heat waves on an experimental plant-herbivore-predator food chain. Glob. Chang. Biol. 2013, 19, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Ciss, M.; Parisey, N.; Fournier, G.; Taupin, P.; Dedryver, C.A.; Pierre, J.S. Response of insect relative growth rate to temperature and host-plant phenology: Estimation and validation from field data. PLoS ONE 2014, 9, e86825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, H.E.; Pell, J.K. Interactions between entomopathogenic fungi and other natural enemies: Implications for biological control. Biocontrol Sci. Technol. 2000, 10, 737–752. [Google Scholar] [CrossRef]
- Ramsden, M.; Menendez, R.; Leather, S.; Wäckers, F. Do natural enemies really make a difference? Field scale impacts of parasitoid wasps and hoverfly larvae on cereal aphid populations. Agric. For. Entomol. 2017, 19, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Martínez, S.A.; Lavandero, B. The effect of landscape context on the biological control of Sitobion avenae: Temporal partitioning response of natural enemy guilds. J. Pest Sci. 2018, 91, 41–53. [Google Scholar] [CrossRef]
- Yang, L.; Liu, B.; Zhang, Q.; Zeng, Y.; Pan, Y.; Li, M.; Lu, Y. Landscape structure alters the abundance and species composition of early-season aphid populations in wheat fields. Agric. Ecosyst. Environ. 2019, 269, 167–173. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Carrao, H.; Barbosa, P.; Vogt, J. World drought frequency, duration, and severity for 1951–2010. Int. J. Climatol. 2014, 34, 2792–2804. [Google Scholar] [CrossRef] [Green Version]
- Aslam, T.J.; Johnson, S.N.; Karley, A.J. Plant-mediated effects of drought on aphid population structure and parasitoid attack. J. Appl. Entomol. 2013, 137, 136–145. [Google Scholar] [CrossRef]
- Romo, C.M.; Tylianakis, J.M. Elevated Temperature and Drought Interact to Reduce Parasitoid Effectiveness in Suppressing Hosts. PLoS ONE 2013, 8, e58136. [Google Scholar] [CrossRef] [Green Version]
- Beetge, L.; Krüger, K. Drought and heat waves associated with climate change affect performance of the potato aphid Macrosiphum euphorbiae. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mattson, W.J.; Haack, R.A. The role of drought in outbreaks of plant-eating insects. Bioscience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- Cui, H.; Wang, L.; Reddy, G.V.P.; Zhao, Z. Mild Drought Facilitates the Increase in Wheat Aphid Abundance by Changing Host Metabolism. Ann. Entomol. Soc. Am. 2020, 1–5. [Google Scholar]
- Tariq, M.; Wright, D.J.; Bruce, T.J.A.; Staley, J.T. Drought and Root Herbivory Interact to Alter the Response of Above-Ground Parasitoids to Aphid Infested Plants and Associated Plant Volatile Signals. PLoS ONE 2013, 8, e069013. [Google Scholar]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Sigsgaard, L. A survey of aphids and aphid parasitoids in cereal fields in Denmark, and the parasitoids’ role in biological control. J. Appl. Entomol. 2002, 126, 101–107. [Google Scholar] [CrossRef]
- Andrade, T.O.; Outreman, Y.; Krespi, L.; Plantegenest, M.; Vialatte, A.; Gauffre, B.; Van Baaren, J. Spatiotemporal variations in aphid-parasitoid relative abundance patterns and food webs in agricultural ecosystems. Ecosphere 2015, 6, Art113. [Google Scholar] [CrossRef] [Green Version]
- Eoche-Bosy, D.; Outreman, Y.; Oliveira Andrade, T.; Krespi, L.; van Baaren, J. Seasonal variations of host resources influence foraging strategy in parasitoids. Entomol. Exp. Appl. 2016, 161, 11–19. [Google Scholar] [CrossRef]
- Le Lann, C.; Roux, O.; Serain, N.; Van Alphen, J.J.M.; Vernon, P.; Van Baaren, J. Thermal tolerance of sympatric hymenopteran parasitoid species: Does it match seasonal activity? Physiol. Entomol. 2011, 36, 21–28. [Google Scholar] [CrossRef]
- Sigsgaard, L. The temperature-dependent duration of development and parasitism of three cereal aphid parasitoids, Aphidius ervi, A. rhopalosiphi, and Praon volucre. Entomol. Exp. Appl. 2000, 95, 173–184. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Kavallieratos, N.G.; Starý, P.; Petrović-Obradović, O.; Athanassiou, C.G.; Stanisa Vljević, L.Ž. Cereal aphids (Hemiptera: Aphidoidea) in Serbia: Seasonal dynamics and natural enemies. Eur. J. Entomol. 2008, 105, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.A.; Heinen, R.; Gols, R.; Thakur, M.P. Climate change-mediated temperature extremes and insects: From outbreaks to breakdowns. Glob. Chang. Biol. 2020, 26, 6685–6701. [Google Scholar] [CrossRef]
- Malina, R.; Praslička, J. Effect of temperature on the developmental rate, longevity and parasitism of Aphidius ervi haliday (Hymenoptera: Aphidiidae). Plant Prot. Sci. 2008, 44, 19–24. [Google Scholar] [CrossRef]
- Flores-Mejia, S.; Guay, J.F.; Fournier, V.; Cloutier, C. The influence of a parasitoid’s response to temperature on the performance of a tri-trophic food web. Ecol. Entomol. 2016, 41, 431–441. [Google Scholar] [CrossRef]
- Le Lann, C.; Lodi, M.; Ellers, J. Thermal change alters the outcome of behavioural interactions between antagonistic partners. Ecol. Entomol. 2014, 39, 578–588. [Google Scholar] [CrossRef]
- Ismail, M.; Van Baaren, J.; Hance, T.; Pierre, J.S.; Vernon, P. Stress intensity and fitness in the parasitoid Aphidius ervi (Hymenoptera: Braconidae): Temperature below the development threshold combined with a fluctuating thermal regime is a must. Ecol. Entomol. 2013, 38, 355–363. [Google Scholar] [CrossRef]
- Mackauer, A.M.; Völkl, W.; Url, S. Regulation of Aphid Populations by Aphidiid Wasps: Does Parasitoid Foraging Behaviour or Hyperparasitism Limit Impact? Oecologia 1993, 94, 339–350. [Google Scholar] [CrossRef]
- Traugott, M.; Bell, J.R.; Raso, L.; Sint, D.; Symondson, W.O.C. Generalist predators disrupt parasitoid aphid control by direct and coincidental intraguild predation. Bull. Entomol. Res. 2012, 102, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Baumann, P. Biology bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Koga, R.; Fukatsu, T. Host plant specialization governed by facultative symbiont. Science 2004, 303, 1989. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [Green Version]
- Vorburger, C.; Gehrer, L.; Rodriguez, P.; Douglas, A.E.; Ferrari, J.; Müller, C.B.; Kraaijeveld, A.R.; Godfray, H.C.J.; Ferrari, J.; Darby, A.C.; et al. A strain of the bacterial symbiont Regiella insecticola protects aphids against parasitoids. Biol. Lett. 2010, 6, 109–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Godfray, H.C.J. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecol. Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Degnan, P.H.; Hunter, M.S.; Moran, N.A. Bacteriophages encode factors required for protection in a symbiotic mutualism. Science 2009, 325, 992–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degnan, P.H.; Moran, N.A. Diverse phage-encoded toxins in a protective insect endosymbiont. Appl. Environ. Microbiol. 2008, 74, 6782–6791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.K.; Vorburger, C.; Moran, N.A. Genomic basis of endosymbiont-conferred protection against an insect parasitoid. Genome Res. 2012, 22, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.Q.; Purcell, A.H. Occurrence and transmission of facultative endosymbionts in aphids. Curr. Microbiol. 1997, 34, 220–225. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Zepeda-Paulo, F.; Dion, E.; Lavandero, B.; Mahéo, F.; Outreman, Y.; Simon, J.C.; Figueroa, C.C. Signatures of genetic bottleneck and differentiation after the introduction of an exotic parasitoid for classical biological control. Biol. Invasions 2016, 18, 565–581. [Google Scholar] [CrossRef]
- Harmon, J.P.; Moran, N.A.; Ives, A.R. Species Response to Environmental Change: Impacts of Food Web Interactions and Evolution. Science 2009, 323, 1347–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giller, P.S.; Ryan, B.; Kennedy, T.; Connery, J. Aphid-parasitoid interactions in a winter cereal crop: Field trials involving insecticide application. J. Appl. Entomol. 1995, 119, 233–239. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aphid | Parasitism Rate | R. insecticola Proportion | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Effect | χ2 | df | p | χ2 | df | p | χ2 | df | p |
| Year | 0.316 | 1 | 0.574 | 7.115 | 1 | 0.007 | 0.000 | 1 | 0.999 |
| Zone | 6.190 | 1 | 0.013 | 0.237 | 1 | 0.663 | 4.388 | 1 | 0.111 |
| Sampling date | 124.663 | 5 | <0.001 | 15.810 | 5 | 0.007 | 11.858 | 5 | 0.065 |
| Year × zone | 9.039 | 1 | 0.003 | 6.4722 | 1 | 0.010 | 0.000 | 1 | 0.999 |
| Year × sampling date | 13.126 | 5 | 0.022 | 5.309 | 5 | 0.379 | 0.000 | 5 | 1.000 |
| Zone × sampling date | 73.508 | 5 | <0.001 | 17.741 | 5 | 0.003 | 2.892 | 5 | 0.716 |
| Year × zone × sampling date | 72.122 | 5 | <0.001 | 22.391 | 5 | <0.001 | 0.000 | 5 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepúlveda, D.A.; Barrueto, G.; Correa, M.C.G.; Castañeda, L.E.; Figueroa, C.C. Spatial and Temporal Variation in the Aphid–Parasitoid Interaction under Different Climates. Agriculture 2021, 11, 344. https://doi.org/10.3390/agriculture11040344
Sepúlveda DA, Barrueto G, Correa MCG, Castañeda LE, Figueroa CC. Spatial and Temporal Variation in the Aphid–Parasitoid Interaction under Different Climates. Agriculture. 2021; 11(4):344. https://doi.org/10.3390/agriculture11040344
Chicago/Turabian StyleSepúlveda, Daniela A., Gonzalo Barrueto, Margarita C. G. Correa, Luis E. Castañeda, and Christian C. Figueroa. 2021. "Spatial and Temporal Variation in the Aphid–Parasitoid Interaction under Different Climates" Agriculture 11, no. 4: 344. https://doi.org/10.3390/agriculture11040344
APA StyleSepúlveda, D. A., Barrueto, G., Correa, M. C. G., Castañeda, L. E., & Figueroa, C. C. (2021). Spatial and Temporal Variation in the Aphid–Parasitoid Interaction under Different Climates. Agriculture, 11(4), 344. https://doi.org/10.3390/agriculture11040344