Abstract
Water deficit is one of the most problematic stressors worldwide. In this context, the use of biostimulants represents an increasingly ecological practice aimed to improve crop tolerance and mitigate the negative effects on the productivity. Here, the effect derived from the foliar application of ERANTHIS®®, a biostimulant based on seaweed (Ascophyllum nodosum and Laminaria digitata) and yeast extracts, was tested on tomato plants grown under mild water-stress conditions. The potential stress mitigation action was evaluated by monitoring morphometric (fresh weight and dry matter content), physiological (stem water potential) and biochemical (ROS scavenger enzymes activity, proline, abscisic acid, hydrogen peroxide and photosynthetic pigment content) parameters closely related to the occurrence and response to stress at both flowering and fruit-set timing. In general, we observed that plants grown under drought conditions and treated with the biostimulant had a lower amount of ABA, and MDA and proline correlated to a lower activity of ROS scavenger enzymes compared to untreated plants. These data, together with the higher stem water potential and photosynthetic pigment levels recorded for the treated plants, suggest that ERANTHIS®® may mitigate water stress effects on tomato.
1. Introduction
Nowadays, the lack of water represents one of the most spread abiotic stressors worldwide [1,2]. The impact of climate change is leading to a progressive reduction in rainfall in several geographic areas of our planet, thus resulting in more severe, longer and frequent drought periods that may have negative consequences on plant productivity [3]. Indeed, since water-shortage phenomena slow plant growth and development, they have been estimated to cause not only the loss of more than 50% of the total production [4], but also a significant loss of crop quality [5]. Moreover, drought negatively affects the photosynthesis by damaging chloroplasts, limiting the electron transfer, and generating cellular oxidative stress [6].
In the past, in order to limit the negative effects of abiotic stresses, chemical fertilizers and other polluting formulations have been widely used as agriculture practices [7]. Despite their use positively affecting the growth and agricultural yields of plants grown under adverse environmental conditions, this approach has resulted in other problems, including the massive contamination of the soil and atmosphere [8]. Consequently, finding new eco-sustainable approaches aimed to help plants to counteract the negative effects deriving from the lack of water, without generating further environmental pollution, is one of the most important challenges for agricultural research [9]. One of the possible solutions is represented by biostimulants. These formulations belong to a new generation of products, and are able to positively regulate plant physiological processes and increase abiotic stress tolerance [10,11]. Commonly, these formulations are a mix of bioactive molecules working in synergy. Because of their complex composition, understanding their mechanisms of action is a hard challenge, but several scientific evidences demonstrated that these products are able to help plants to cope with adverse environmental conditions [12]. Among the different classes of biostimulants, seaweed extracts are one of the most widely used in agriculture [13]. These products can be applied to the soil, in hydroponic systems, or as foliar spray, and they are able to improve plant growth, plant tolerance to abiotic and biotic stress, and fruit quality [14]. The effects of seaweed extracts application may be ascribed to the synergistic activity of some compounds, such as hormones, betaines, polyamines, alginates, carrageenans, fucans and phlorotannins [14,15].
Tomato (Solanum lycopersicum L.) is one of the most largely cultivated vegetables belonging to the Solanaceae family around the world, together with potato (Solanum tuberosum L.). Currently, more than 180 million tons of fruits are produced per year and commercialized all over the world [16]. Similarly to the other species belonging to the genus Solanum, tomato is highly susceptible to drought. In particular, it has been shown that a decrease in the irrigation determines a strong loss of quality and production yields [17]. In the present study, the foliar application of ERANTHIS®®, a biostimulant based on brown seaweed (Ascophylum nodosum and Laminaria digitata) and yeast extracts, was tested for its mitigation action on water stress effects on tomato by measuring morphometric (fresh weight and dry matter content), physiological (stem water potential) and biochemical (reactive oxygen species (ROS) scavenger enzyme activity, proline, abscisic acid, hydrogen peroxide and photosynthetic pigment content) parameters.
2. Materials and Methods
2.1. Plant Material, Biostimulant and Experimental Conditions
2.1.1. Plant Material
Solanum lycopersicum (tomato) seeds, var. Micro Tom, were purchased by Pan America Seed (Chicago, IL, USA). Seeds were sown in rock wool cubes and transferred in pots containing 100% sand at the first true leaf appearance. The experiment was carried out in the greenhouse under controlled conditions (17–26 °C, 16 h/8 h day/night, 45–82% humidity). For two weeks, plants were irrigated on alternate days and macro and micronutrients were provided by Hoagland solution [18] once a week. Tomato plants were then divided into the following three different groups: (1) control (untreated) plants grown under well-watered conditions, (2) untreated stressed and (3) biostimulant-treated stressed (ERANTHIS®® stressed) plants, both groups grown under mild water stress conditions.
2.1.2. Biostimulant
The biostimulant ERANTHIS®® was provided by Green Has Italia S.p.A. (Canale, Italy). The formulation contained extracts of seaweed (Ascophyllum nodosum and Laminaria digitata) and yeasts. The label of the products claimed to contain 2.5% (w/w) of organic nitrogen and 14% (w/w) of organic carbon. It had a density of 1.2 g mL−1, a pH (in 1% w/w water solution) of 5.0 ± 0.5 (measured on three different replicates and expressed as mean ± standard deviation) and an electric conductivity (water solution 1 g L−1) of 250 μS cm−1. The quantification of both bioactive compounds and antioxidant properties contained in ERANTHIS®® was experimentally determined via spectrophotometric assays using Cary60®® Agilent spectrophotometer (Agilent Technologies, Santa Clara, CA, USA). For these determinations, the biostimulant was previously diluted 1:1000 (v/v) in water, as previously reported [19,20]. Briefly, Folin–Ciocalteu assay was employed for the evaluation of the total polyphenol content (TPC); aluminum chloride assay was employed for the measurement of both total flavonol (TFlC) and flavonoid (TFdC) content; pH differential method for the total anthocyanin content (TAC); carotenoid assay for the quantification of the total carotenoid content (TCC); BL-DMAC assay for the measurement of the total flavan-3-ol content (TF3C). Moreover, the potential antioxidant property of the biostimulant was also measured in terms of both radical scavenging (ABTS and DPPH assay) and reducing activity (FRAP assay).
2.1.3. Experimental Conditions
For the experiments, a solution containing the biostimulant (2 mL L−1) or tap water only was sprayed on leaves the following four times: (i) at the fourth true leaf appearance; (ii) 7 days after the first treatment; (iii) at flowering time; (iv) at fruit-set time. After the first biostimulant application, stressed plants were watered with 50% less water in comparison to the untreated ones grown under optimal conditions (100% hydration), until the end of the experiment. Plant material was collected 5 h after the third (flowering, time 1) and the fourth (fruit-set, time 2) treatment and used for morphometric (shoots and roots), biochemical (leaves) and physiological (leaves) analyses (Figure 1).
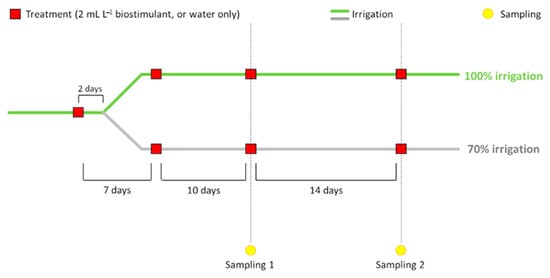
Figure 1.
Schematic representation of the experimental design. At the beginning, both plant groups were irrigated with full water supply. At the fourth true leaf appearance, after the first biostimulant treatment, water stress was induced in half of the plants by reducing the water intake by 50%, resulting in 30% less water at the end of the trial (untreated stressed and treated stressed plants). The other half continued to be well-irrigated (untreated plants). The third treatment was done at flowering time, while the fourth one at fruit set. Plant samples were collected 5 h after the last two treatments. Untreated and untreated stressed plant groups were sprayed with tap water, instead of the biostimulant, at each treatment.
2.2. Morphometric Measurements
Shoots and roots fresh weight (FW) was measured by using a technical scale. The sand was washed away from the roots by using distilled water and these were then dried and weighed. The dry matter content % was measured drying the material at 75 °C for 48 h to obtain the dry weight (DW). The DMC% was measured applying the following formula: (DW/FW) × 100 [21].
2.3. Hydrogen Peroxide (H2O2)
The hydrogen peroxide content was detected as previously reported [22]. Fresh leaves (0.3 g) were powdered and homogenized with 1.5 mL of 0.1% (w/v) trichloroacetic acid (TCA). The samples were centrifuged at 12,000× g for 15 min and 0.5 mL of supernatant was added to 0.5 mL of 10 mM potassium phosphate buffer (pH 7.0) and 1.0 mL of 1 M potassium iodide (KI). After reading the absorbance at 390 nm, the H2O2 content was calculated based on a standard curve.
2.4. Total Soluble Protein Content
The total soluble proteins were extracted as reported according to Contartese and colleagues [23]. The total protein content was evaluated using the Bradford assay, with bovine serum albumin as a standard [24]. Briefly, 100 μL of reaction mix (Bio-Rad Laboratories, Inc., Louisville, KY, USA) was added to equal volume of sample opportunely diluted in water. The absorbance was read at 595 nm after 30 min of incubation against a blank containing 100 μL of extraction buffer and reaction mix.
2.5. SOD—Superoxide Dismutase (EC 1.15.1.1)
Superoxide dismutase activity evaluation was based on the nitro blue tetrazolium (NBT) degradation performed by the enzyme thanks to a superoxide anion, generated photochemically [25]. The reaction was prepared in 1.0 mL final volume, containing 50 mM sodium phosphate buffer pH 7.8, 13 mM methionine, 0.1 mM EDTA (ethylenediaminetetraacetic acid), 75 µM NBT, 2 µM riboflavin and enzyme extract. To avoid degradation, riboflavin was added at last. The reaction was started by placing the samples under a light source (4000 lux) for 15 min. The following two blanks were prepared: one without enzyme extract placed under the light in order to completely develop the reaction and another one containing the enzyme extract placed in the dark to avoid the reaction. The absorbance was detected at 560 nm.
2.6. POX—Peroxidase (EC 1.11.1.7)
Guaiacol was used as reaction substrate. The enzyme activity was measured by monitoring the absorbance variation at 470 nm for 15 min. One mL final volume contained 50 mM sodium phosphate buffer pH 7.0, 0.27 mM hydrogen peroxide, enzyme extract and 0.33 mM guaiacol. The reaction was started by adding the guaiacol, which leads to a color change towards orange [22].
2.7. GST—Glutathione-S-Transferase (EC 2.5.1.18)
The reaction was performed using 1-Chloro-2,4-dinitrobenzene (CDNB) as a substrate and to evaluate the enzyme activity the absorbance variation at 340 nm was monitored for 15 min. One mL reaction mixture was composed by 100 mM potassium phosphate buffer (pH 7.0), 1 mM reduced glutathione (GSH), 1 mM 1-chloro-2,4 dinitrobenzene (CDNB) (10 mM CDNB dissolved in 50% acetone stock solution), and enzyme extract. The reaction was started by adding CDNB [26].
2.8. ABA—Abscisic Acid
Abscisic acid (ABA) extraction and quantification was performed as previously reported [27]. Briefly, ABA was extracted using a mix of 80% (v/v) methanol, acidified with 1% (v/v) acetic acid. The extracts were filtered through 0.45 μm membrane filters, and analyzed by UHPLC-MS/MS. A standard curve was prepared by dissolving different concentrations of standard phytohormone in the same mix, from 60 ppt to 30 ppb. Abscisic acid profiles were carried out by using a Shimadzu Nexera UHPLC system coupled with a Shimadzu LCMS-8040 triple quadrupole (QqQ) mass spectrometer (Shimadzu Corporation, Kyoto, Japan). A Restek Raptor Biphenyl LC column (2.1 mm × 100 mm; 2.7 μm particle size) (Restek Corporation, Bellefonte, PA, USA) was used. The optimized chromatographic conditions were achieved using ultra-pure water with 0.1% (v/v) of HCOOH as solvent A and CH3CN as solvent B. The column temperature was set at 25 °C under 200 µL min−1 flow. The elution profile was performed isocratically at 60% (v/v) of solvent B from 0.1–5 min, followed by a 2 min of cleaning at 98% (v/v) of solvent B, and finally, 5 min to return to initial conditions for the re-equilibration of the column. The injection volume was 5 μL. The QqQ mass spectrometer was equipped with an electrospray ionization source (ESI) operating in negative mode using argon (Ar) as collision gas, the source temperature was 250 °C, and the desolvation temperature was 400 °C. The nebulizing and drying gas flow (N2) was set to 3 L min−1. Ionized samples were analyzed in multiple reaction monitoring (MRM) using following abscisic acid quantification transitions: m/z 263.30/153.00, CE = 10.0 V. Lab Solutions software (Shimadzu Corporation, Kyoto, Japan) was used for control and data processing.
2.9. Malondialdehyde
The extent of lipid peroxidation was estimated by determining the concentration of thiobarbituric acid-reactive substances (TBARS). Malondialdehyde extraction was performed in 0.1% (w/v) trichloroacetic acid (TCA). The extract was centrifuged at 15,000× g for 10 min and 0.5 mL of the supernatant obtained was then added to 1.5 mL 0.5% (w/v) TBA in 20% (w/v) TCA. The mixture was incubated at 90 °C in a shaking water bath for 20 min, and the reaction was stopped in ice-water bath. Then, the samples were centrifuged at 10,000× g for 5 min, and the absorbance was read at 532 nm. The value for non-specific absorption at 600 nm was subtracted. The amount of TBARS (red pigment) was calculated from the extinction coefficient 155 mM−1 cm−1 [28].
2.10. Proline
Proline extraction was performed with a mix containing 1% (w/v) ninhydrin dissolved in 60% acetic acid (v/v) and 20% ethanol (v/v). After two days at 4 °C, the extract was heated at 95 °C for 20 min and then centrifuged at 11,200× g for 1 min. The supernatant was transferred in a cuvette and the absorbance was read at 520 nm [27].
2.11. Photosynthetic Pigments
Chlorophyll and carotenoid extraction was performed in ethanol 96% (v/v) in a 1:10 (w/v) proportion. The absorbance was read at 664 nm (chlorophyll a), 649 nm (chlorophyll b) and 470 nm (carotenoids). The pigment content was calculated based on [29].
2.12. Stem Water Potential Measurement
Stem water potential (ψstem) was measured for each timing (flowering and fruit set) using a Scholander-type pressure chamber (Soil Moisture Equipment Corp., Santa Barbara, CA, USA). One leaf for plant was placed in a humidified plastic bag covered with aluminum foil to stop transpiration. After 30 min the leaves were cut and allowed to equilibrate in dark conditions before the measurement.
2.13. Statistical Analysis
All statistical analyses were performed by using SYSTAT 10.0 (Systat Software Inc., San Jose, CA, USA). The obtained results were analyzed by t-test comparing untreated/stressed with untreated/unstressed plants (graphs related to water stress effect) or treated/unstressed with untreated/stressed plants (graphs related to biostimulant effects). The significant threshold was imposed under 0.05. For stem water potential data, one way ANOVA followed by Tukey’s post hoc test was performed. Moreover, the correlation between ABA content and water potential data was verified dividing the covariance of the two variables by the product of their standard deviations.
3. Results
3.1. Bioactive Compounds and Antioxidant Properties of the Biostimulant ERANTHIS®®
The spectrophotometric characterization of the biostimulant revealed that the formulation contained bioactive molecules having an oxygenated heterocyclic ring functionalized with one or more hydroxyl groups (Table 1). In particular, phenolic compounds represented about 1.5% of the product fresh weight. Our analyses also showed that many of these compounds belonged to flavonoid, flavan-3-ol and flavanol families. On the other hand, as measured by the pH differential method and carotenoid assay, anthocyanins and carotenoids were not present. Finally, the biostimulant recorded high values for ABTS, DPPH and FRAP assays. In particular, DPPH and ABTS were higher with respect to FRAP, suggesting that ERANTHIS®® had a stronger radical scavenging activity compared to the reducing capacity.

Table 1.
UV/Vis spectrophotometric determination of bioactive compounds and antioxidant properties of ERANTHIS®®. Values are expressed as a mean ± SD of three different replicates.
3.2. Water Stress and ERANTHIS®® Application Effects on Morphometric Parameters
Tomato plants grown with a reduced water supply (30% less water provided compared to the optimal conditions) showed a significant decrease in their shoot and root fresh weight (FW) in comparison to control tomato plants. However, an increase in the percentage of shoot and root dry matter content was observed for the same comparison (Figure 2). Significant differences were observed at flowering (Figure 2A) for both fresh weight and dry matter content measurements, while they were less evident at fruit-set time (Figure 2B). In particular, the shoot FW of the stressed plants was −48.3% and −46.2%, respectively, at flowering and fruit set, meanwhile the root FW was −38.1% and −37.2% at the same sampling time, respectively. The most significant differences were observed for the shoot and root dry matter contents; an increment of about +50% and +15% was observed in the shoot dry matter content, respectively, at flowering and fruit-set timing. On the other hand, the root dry matter content was +59.5% and +39.2%, respectively.
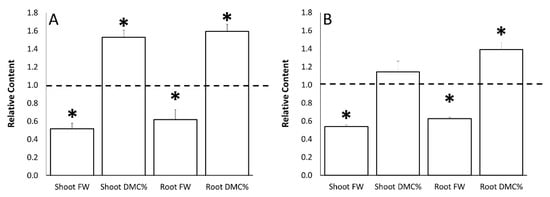
Figure 2.
Water stress effects on the fresh weight (FW) and dry matter content percentage (DMC%) of tomato shoot and roots at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measurements obtained by untreated/stressed and untreated/unstressed (bars). The dotted line indicates the relative value of each parameter related to untreated/unstressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between untreated/stressed and untreated/unstressed, as measured by t-test.
After evaluating the water stress effects on the untreated plants, the influence of ERANTHIS®® on the plant morphometric parameters was evaluated under stress conditions. Figure 3 shows the comparison between biostimulant-treated stressed and untreated stressed plants at flowering (Figure 3A) and fruit-set timing (Figure 3B). In this case, significant differences were observed at the first sampling time only (Figure 3A), while at the second one, the treated stressed plants showed values very similar to the untreated stressed plants (Figure 3B). At flowering, root FW did not differ from untreated stressed plants, while the shoot FW was −17.9%, the shoot dry matter content was −20.6%, and the root dry matter content was −7.2%.
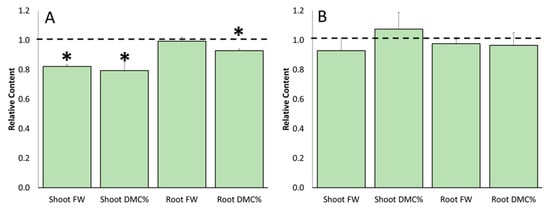
Figure 3.
Effects derived from the biostimulant application on the fresh weight (FW) and dry matter content percentage (DMC%) of tomato shoot and roots at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measurements obtained by treated/stressed and untreated/stressed (bars). The dotted line indicates the relative value of each parameter related to untreated/stressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between treated/stressed and untreated/stressed, as measured by t-test.
3.3. Stem Water Potential and Its Correlation with Abscisic Acid Content
Stem water potential (ψstem) is considered one of the most important physiological parameters to evaluate plant water status [30,31].
Figure 4 displays the changes in stem water potential, recorded in tomato plants under our experimental conditions. When stressed, both the treated and untreated plants showed a reduction in the stem water potential values. On the one hand, the ψstem of the untreated/stressed plants strongly decreased in comparison to well-irrigated plants at both flowering time (+152.9%) and fruit setting (+148.4%), (Figure 4A,B). On the other hand, the treated/stressed plants showed less reduced stem water potential values, −95.6% and −48.5%, respectively, at flowering at fruit setting with respect to well-irrigated plants (Figure 4A,B).
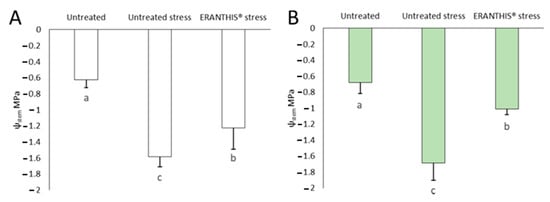
Figure 4.
Stem water potential of untreated/unstressed, untreated/stressed and biostimulant-treated/stressed plants at flowering (A) and fruit-set (B) sampling time. Bars with different lowercase letters indicate significant different values at p ≤ 0.05 as measured by one-way ANOVA followed by Tukey’s HSD post hoc test.
Abscisic acid (ABA) is one of the hormones involved in plant signaling in response to water stress [32], and its rising content is correlated to more negative ψstem values, as shown in Figure 5. The highest stress level, namely, the lowest ψstem and the highest ABA content, was observed in the untreated/stressed plants, while ERANTHIS®®-treated/stressed plants showed less ABA content in the leaves correlated to higher stem water potential values. The stress mitigation effect was more evident at fruit set (Figure 5).
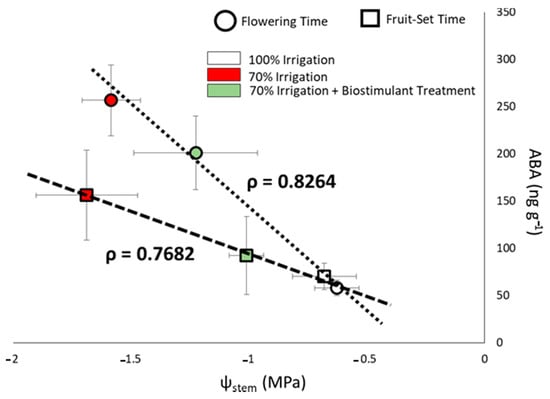
Figure 5.
Stem water potential (ψstem) and abscisic acid (ABA) correlation at flowering (circles) and fruit-set (squares) sampling time. The white shapes represent untreated plants with 100% full water supply, the red ones untreated/stressed plants and the green ones treated/stressed plants. The dotted lines show the correlation between ABA content and ψstem at the two different sampling times. The ρ coefficient was calculated dividing the covariance of the two variables by the product of their standard deviations.
3.4. Water Stress Effects on Stress-Related Biochemical Parameters
Stress occurrence was also verified by monitoring different non-enzymatic and enzymatic parameters notoriously related to stress and plant responses to stress. Figure 6 summarizes the results regarding the selected non-enzymatic parameters, namely, photosynthetic pigment content, together with leaf ABA, H2O2, MDA, and proline accumulation. Water stress led to a decrease in photosynthetic pigment accumulation, while the values of all the other leaf parameters were significantly higher with respect to plants grown under well-watered conditions (Figure 6A). Upon water stress, the total chlorophyll and carotenoid content was 39.7% and 35.3% lower (Figure 6A), respectively, at the flowering stage, while its decrease was not significant at fruit setting (Figure 6B). In general, the increase in all the other parameters was enhanced at flowering with respect to fruit-setting time. In detail, the ABA content was around four times and two times higher (Figure 6A,B) compared to well-irrigated plants at flowering and fruit setting, respectively, whereas hydrogen peroxide increased by 110.9% and 67.2%, MDA 79.1% and 33.6%, and finally proline 234.4% and 109.7%, at the two time points, respectively.
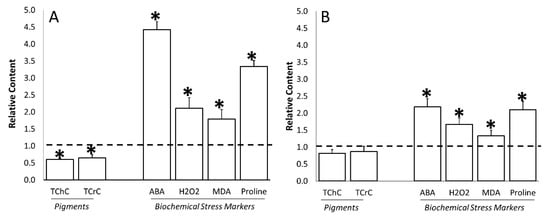
Figure 6.
Water stress effects on the total content of chlorophyll (TChC), carotenoids (TCrC), abscisic acid (ABA), hydrogen peroxide (H2O2), malondialdehyde (MDA), and proline of tomato leaves at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measurements obtained by untreated/stressed and untreated/unstressed (bars). The dotted line indicates the relative value of each parameter related to untreated/unstressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between untreated/stressed and untreated/unstressed, as measured by t-test.
Figure 7 showed the increase in the activity of ROS scavenger enzymes, namely, SOD, POX and GST, which were induced when the tomatoes were exposed to water stress. In particular, GST activity was 1.5 times higher at both the flowering and fruit setting (Figure 7A,B). Differently, as already observed for non-enzymatic parameters, POX and SOD enzyme activities resulted to be less induced at the fruit setting with respect to flowering time. Indeed, POX was 4.5 times and 2.5 times higher at the flowering and fruit setting, respectively, while SOD was two times and 1.5 times higher at the same sampling times.
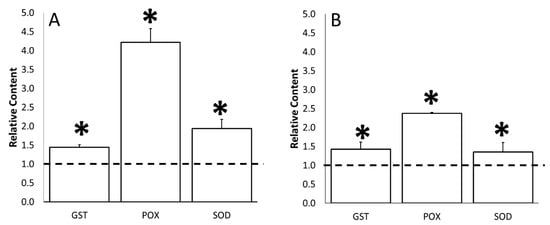
Figure 7.
Water stress effects on the content of glutathione-S-transferase (GST), peroxidase (POX), and superoxide dismutase (SOD) of tomato leaves at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measurements obtained by untreated/stressed and untreated/unstressed (bars). The dotted line indicates the relative value of each parameter related to untreated/unstressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between untreated/stressed and untreated/unstressed, as measured by t-test.
3.5. ERANTHIS®® Influence on Stress-Related Biochemical Parameters under Stress Conditions
After observing the effects of water stress and its related plant responses, we evaluated whether ERANTHIS®® treatments were able to modulate the level of the analyzed enzymatic and non-enzymatic parameters under stress conditions. As shown in Figure 8, the biostimulant applications affected the content of photosynthetic pigments that significantly increased at both the flowering and fruit-set timing (Figure 8A,B), while the levels of biochemical stress markers, such as ABA, proline and MDA, decreased. The total chlorophyll and carotenoids were, respectively, 1.3 and 1.5 times higher at flowering, and 1.6 and 1.3 times higher at fruit setting. On the contrary, ABA accumulation decreased by about 22% at the first sampling time and 42.6% at the second one. The malondialdehyde level decreased by 40.4% and 24.5% with the treatment, while the proline content was reduced by 14.7% and 41.1% at both the time points, respectively. The ERANTHIS®® applications did not affect the content of hydrogen peroxide.
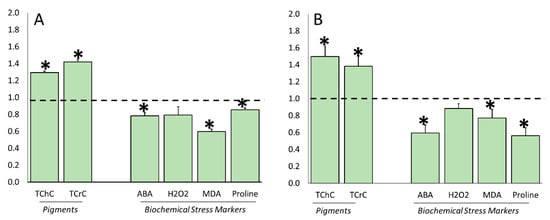
Figure 8.
Effects derived from the biostimulant application on the total content of chlorophyll (TChC), carotenoids (TCrC), abscisic acid (ABA), hydrogen peroxide (H2O2), malondialdehyde (MDA), and proline of tomato leaf at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measurements obtained by treated/stressed and untreated/stressed (bars). The dotted line indicates the relative value of each parameter related to untreated/stressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between treated/stressed and untreated/stressed, as measured by t-test.
As for enzymatic scavenging (Figure 9), only SOD showed a significant decreased activity at flowering, its values being 3.6 times lower compared to the stressed plants (Figure 9A). Differently, the other ROS enzymatic scavenger activity was lower in the stressed/treated compared to the untreated/stressed plants, even if the result was not significantly different. Except for SOD, the enzymatic activity values were very similar at both the sampling times.
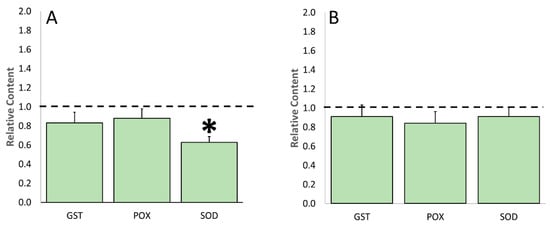
Figure 9.
Effects derived from the biostimulant application on the content of glutathione-S-transferase (GST), peroxidase (POX), and superoxide dismutase (SOD) of tomato leaves at flowering (Panel A) and fruit-set (Panel B) sampling time. Data of each evaluation were expressed as relative content, comparing the measures obtained by treated/stressed and untreated/stressed (bars). The dotted line indicates the relative value of each parameter related to untreated/stressed plants. The symbol “*”, when present, indicates significant differences (p ≤ 0.05) between treated/stressed and untreated/stressed, as measured by t-test.
4. Discussion
Biostimulants are complex matrices containing substances of natural origin with different useful active compounds, as products or plant extracts, often derived from processing waste. In particular, ERANTHIS®® contains seaweed extracts (Ascophyllum nodosum and Laminaria digitata) and yeast extract substances. Seaweed extracts are known to be rich in bioactive compounds, such as phenolic compounds, with antioxidant properties [33]. In this study, a preliminary chemical characterization of the biostimulant was carried out via UV/Vis spectrophotometry. Our analyses confirmed that ERANTHIS®® is a rich source of phenolic compounds, mainly belonging to the flavonoid and flavanol family, while a small amount of flavan-3-ols was also detected. However, anthocyanins (LOD: 0.05 mmol; LOQ: 0.16 mmol) and carotenoids (LOD: 0.12 mmol; LOQ: 0.36 mmol) were absent. Surprisingly, the evaluation of the antioxidant capacity of the formulation revealed that ERANTHIS®® had a strong radical scavenging activity and a low reducing metal activity. As previously reported in the literature, the metal-reducing activity has been commonly correlated to polyphenolic molecules that have hydroxyl groups orto- or meta-oriented on their own chemical scaffold. This peculiar orientation of the hydroxyl groups allows the chelation of organic metals by polyphenol compounds, thus exerting a strong reducing activity [34]. Consequently, taking into account the values recorded for ABTS, DPPH and FRAP, it is reasonable to think that the bioactive molecules present in the ERANTHIS®® formulation have either a single hydroxyl group or two positioned in para.
Because of their specific formulation, biostimulants represent a category of products known to improve the mitigation and tolerance to abiotic stresses [10]. On the one hand, seaweed extracts are among the most common substances used as biostimulants and they might play a role in counteracting drought stress [35]. Indeed, Ascophyllum nodosum extracts have been shown to enhance drought tolerance in tomato [36], soybean [37], bean [38], and Salvia officinalis [39], by modulating the osmotic and antioxidant response to stress. On the other hand, yeast extracts are substances known to increase the yield and quality of different crops, including tomato [40], orange [41] and wheat [42], under both control and stress conditions. In particular, Hammad and colleagues observed a reduced induction of stress-related responses in biostimulant-treated wheat, thus suggesting a mitigation of the drought stress effect instead of a tolerance increase in this crop [42].
In our experiment, we evaluated the influence of ERANTHIS®® application on tomato morphometric, physiological and biochemical responses to mild water stress at flowering and fruit-set timing. One of the most evident effects of the reduction in water supply is the reduction in plant biomass and production yield in different vegetable crops, including tomato [43,44]. With regard to biomass, the same trend was observed in our study, with the strong decrease in shoot and root fresh weight in the stressed plants compared to those grown under optimal conditions at both the flowering and fruit-set sampling times. Moreover, the dry matter content was increased by stress at flowering, while the difference was less evident at fruit set [45]. ERANTHIS®® application changed tomato morphometric response to stress at the flowering time only. Indeed, the decrease in shoot FW, and the increase in shoot and root dry content were less evident than those of untreated plants. Similarly, field applications of two algal derivatives cause a 26% reduction in shoot biomass and a remodulation of the root-to-shoot ratio under osmotic stress [13].
Although ERANTHIS®® slightly changes tomato morphometric responses to stress, its influence on tomato physiological and biochemical responses to stress was more evident. This observation could be related to the plant material collection timing and to the crop variability. Indeed morphometric, physiological and biochemical parameters did not show changes at the same time, since before observing modifications at the phenotypic level, variations in gene expression, protein synthesis and physiological processes are necessary. Moreover, crop plant variability may negatively affect some parameters more than others [46]. From a physiological point of view, one of the most evident effects of progressive soil dehydration is stem water potential decrease [47]. The same result was observed in our experiment in tomato plants irrigated with 30% less water than those grown under the optimal conditions. However, the stem water potential in stressed plants treated with the biostimulant was less reduced than those of untreated stressed plants. Biostimulants are known to help plants in counteracting water shortage [10]. Plants of mint grown under three watering levels (50%, 70% or 100% of substrate capacity) and treated with different kinds of biostimulants, showed a significant increase in stem water potential compared to plants without the biostimulant treatment at all the watering conditions [48]. Stem water potential level, a sensitive indicator of water stress, could be correlated to that of ABA [32], since it is involved in the plant systemic response to stress, and the raising of its level corresponds to a raising of the defense barriers [49,50]. Tomato plants grown under water stress conditions showed a significant increase in ABA content, such as observed in maize plants stressed by using polyethylene glycol (PEG) to induce dehydration [51]. However, when treated with the biostimulant, water stressed tomato plants showed a reduction in ABA accumulation.
Together with a decrease in ABA level under stress conditions, ERANTHIS®® application also led to a significant decrease in oxidative stress level. Water stress is indeed known to lead the increase in non-enzymatic oxidative stress-related molecules, such as MDA and H2O2 [52]. Hydrogen peroxide is a reactive oxygen species produced at the basal level during plant life, but strongly increased after stress induction. Whereas, malondialdehyde is a marker of lipid peroxidation and it normally increases in stress conditions [53]. After the biostimulant treatments, MDA levels were much lower compared to untreated plants under water stress, such as observed by Goni [36]. Together with lipid peroxidation, a negative consequence of oxidative stress is the decrease in photosynthetic pigments with a consequent photosynthetic efficiency reduction [54,55]. Parida and colleagues [56] observed a decrease in both chlorophyll and carotenoid content under water stress conditions. Accordingly, water stress decreased the tomato content of both the photosynthetic pigments, but only at the flowering time. This effect could be due to an acclimatization to the stress [57,58], already observed by Munns in plants grown under salt stress, a condition similar to drought [59]. Moreover, the application of ERANTHIS®® reduced the affection of water stress on both chlorophyll and carotenoid leaf levels at the flowering time. Similarly, the use of other Ascophyllum nodosum-based biostimulant limits the damage, caused by water deficit, to the photosynthetic structures of tomato plants under water stress at the flowering time [60].
The increase in ABA and H2O2 during water stress can lead to the increase in plant antioxidant enzymatic [51,61,62] and non-enzymatic [63] scavenging machinery, such as that observed in the untreated/stressed plants under our experimental conditions. Concerning the enzymatic scavenging, POX, SOD and GST activity levels increased when the plants were stressed. However, ERANTHIS®®-treated tomatoes exhibited a lower increase in SOD activity with respect to untreated plants. Similarly, cherry tomato stressed seedlings showed a reduction in SOD activity when treated with different seaweed biostimulants [64]. As for the non-enzymatic antioxidant scavenging, our experiment confirmed the accumulation of proline under stress conditions [65]; however, ERANTHIS®® treatment reduced the level of this osmolyte. Differently, other seaweed-based biostimulants induced an increase in proline accumulation under stressed conditions [36]. Since higher SOD activity is one of the top stress indicators in plants, our biostimulant could possibly act by postponing drought stress. Moreover, reduced proline accumulation in plants treated with the biostimulant might be also related to a basal lower stress level.
5. Conclusions
Biostimulants are considered good allies in countering abiotic stresses, such as water shortage. Tomato is a crop largely cultivated all around the world, and the use of these kinds of products can be useful to avoid damages and production losses [66]. Taken together, our results suggest a stress mitigation in tomato plants, due to the biostimulant application. Indeed, ERANTHIS®® application reduced ψstem decrease and ABA accumulation under mild water stress conditions [5]. In comparison with untreated/stressed plants, the general decrease in oxidative stress level (lower MDA level and higher photosynthetic pigment content), and the reduction in tomato antioxidant enzymatic and non-enzymatic responses (reduced SOD and proline level) could also support the hypothesized stress mitigation effect of ERANTHIS®®, especially at the tomato flowering time. This effect could be related to the biostimulant formulation itself. Indeed, as demonstrated through spectrophotometric analysis, ERANTHIS®® contains antioxidant molecules, such as flavonoids and flavanols, which are able to contribute to ROS scavenging [42]. These molecules are probably derived from seaweed extracts, known to be rich in antioxidant compounds [67]. Further studies and insights are ongoing to better understand the mode of action of ERANTHIS®® at the molecular level. Indeed, Systems and colleagues described the possible mode of action of seaweed extracts related to the modulation of genes involved in the stress response, such as those responsible for pigment production and plant antioxidant response [68]. Anyway, a biostimulant’s mode of action is complex to be studied because of the presence of different raw materials acting in synergy, and for this reason a multidisciplinary approach is needed [69]. Moreover, biostimulants are not normally species-specific, so the hope is that this type of effect will also be observed on other crops [1]. For this reason, it will be important to continue with further studies, in order to deepen ERANTHIS®® mode of action and verify its effects on different crops.
Author Contributions
Conceptualization, V.C., F.S. and C.M.B.; data curation, C.C., C.A., G.M. and I.V.; formal analysis, C.C., C.A., G.M. and I.V.; funding acquisition, F.S. and C.M.B.; investigation, C.C. and F.S.; methodology, C.C., C.A., I.V. and C.M.B.; project administration, F.S. and C.M.B.; resources, F.S. and C.M.B.; software, G.M.; supervision, F.S.; visualization, G.M.; writing—original draft, C.C., C.A., G.M. and C.M.B.; writing—review and editing, I.V., V.C. and F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the local research grants (ex 60%, 2019–2020) from the University of Turin, Italy, assigned to C.M.B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available because a patent related to a part of this work is pending.
Conflicts of Interest
C.C., V.C. and I.V. were employed by Green Has Italia S.p.A. C.C. was also a PhD student of the PhD program in Pharmaceutical and Biomolecular Sciences of the University of Turin, and in apprenticeship in Green Has Italia S.p.A. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Green Has Italia S.p.A. provided the biostimulant used for this experimentation free of charge.
References
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoB Plants 2018, 10, ply016. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Antle, J.; Garrett, K.A.; Izaurralde, R.C.; Mader, T.; Marshall, E.; Nearing, M.; Robertson, G.P.; Ziska, L. Indicators of climate change in agricultural systems. Clim. Chang. 2020, 163, 1719–1732. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P.; Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hasan, S.A.; Fariduddin, Q.; Ahmad, A. Growth of tomato (Lycopersicon esculentum) in response to salicylic acid under water stress. J. Plant Interact. 2008, 3, 297–304. [Google Scholar] [CrossRef]
- Arora, N.K. Agricultural sustainability and food security. Environ. Sustain. 2018, 1, 217–219. [Google Scholar] [CrossRef]
- Liliane, T.N.; Charles, M.S. Factors Affecting Yield of Crops. Factors Affecting Yield of Crops. Agron. Clim. Chang. Food Secur. 2020, 1–16. [Google Scholar] [CrossRef]
- Hagenlocher, M.; Meza, I.; Anderson, C.C.; Min, A.; Renaud, F.G.; Walz, Y.; Siebert, S.; Sebesvari, Z. Drought vulnerability and risk assessments: State of the art, persistent gaps, and research agenda. Environ. Res. Lett. 2019, 14, 083002. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Agliassa, C.; Acquadro, A.; Contartese, V.; Garabello, C.; Bertea, C.M. Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants 2020, 9, 1308. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.; Yakhin, I.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef]
- Di Stasio, E.; Cirillo, V.; Raimondi, G.; Giordano, M.; Esposito, M.; Maggio, A. Osmo-Priming with Seaweed Extracts Enhances Yield of Salt-Stressed Tomato Plants. Agronomy 2020, 10, 1559. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Brassica napus Growth is Promoted by Ascophyllum nodosum (L.) Le Jol. Seaweed Extract: Microarray Analysis and Physiological Characterization of N, C, and S Metabolisms. J. Plant Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- United Nations. The Future of Food and Agriculture: Trends and Challenges. Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. Available online: http://www.fao.org/3/a-i6583e.pdf (accessed on 17 June 2021).
- FAO. The Future of Food and Agriculture: Trends and Challenges. Available online: http://www.fao.org/3/i6583e/i6583e.pdf (accessed on 17 June 2021).
- Duan, J.-X.; Duan, Q.-X.; Zhang, S.-F.; Cao, Y.-M.; Yang, C.-D.; Cai, X.-D. Morphological, physiological, anatomical and histochemical responses of micropropagated plants of Trichosanthes kirilowii to hydroponic and soil conditions during acclimatization. Plant Cell Tissue Organ Cult. 2020, 142, 177–186. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Grange, E.; Mannino, G.; Van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance During Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Perrone, A.; Campobenedetto, C.; Schittone, A.; Bertea, C.M.; Gentile, C. Phytochemical profile and antioxidative properties of Plinia trunciflora fruits: A new source of nutraceuticals. Food Chem. 2020, 307, 125515. [Google Scholar] [CrossRef] [PubMed]
- Birouste, M.; Zamora-Ledezma, E.; Bossard, C.; Pérez-Ramos, I.M.; Roumet, C. Measurement of fine root tissue density: A comparison of three methods reveals the potential of root dry matter content. Plant Soil 2014, 374, 299–313. [Google Scholar] [CrossRef]
- Cantarello, C.; Volpe, V.; Azzolin, C.; Bertea, C. Modulation of enzyme activities and expression of genes related to primary and secondary metabolism in response to UV-B stress in cucumber (Cucumis sativus L.). J. Plant Interact. 2005, 1, 151–161. [Google Scholar] [CrossRef]
- Contartese, V.; Garabello, C.; Occhipinti, A.; Barbero, F.; Bertea, C.M. Effects of a new biostimulant on gene expression and metabolic responses of tomato plants. Acta Hortic. 2016, 1148, 35–42. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Krishnan, N.; Chattopadhyay, S.; Kundu, J.K.; Chaudhuri, A. Superoxide dismutase activity in haemocytes and haemolymph of Bombyx mori following bacterial infection. Curr. Sci. 2002, 83, 321–325. [Google Scholar]
- Jain, M.; Bhalla-Sarin, N. Glyphosate-Induced Increase in Glutathione S-Transferase Activity and Glutathione Content in Groundnut (Arachis hypogaea L.). Pestic. Biochem. Physiol. 2001, 69, 143–152. [Google Scholar] [CrossRef]
- Mannino, G.; Nerva, L.; Gritli, T.; Novero, M.; Fiorilli, V.; Bacem, M.; Bertea, C.M.; Lumini, E.; Chitarra, W.; Balestrini, R. Effects of Different Microbial Inocula on Tomato Tolerance to Water Deficit. Agronomy 2020, 10, 170. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophyll Fluorescence Signatures of Leaves during the Autumnal Chlorophyll Breakdown. J. Plant Physiol. 1987, 131, 101–110. [Google Scholar] [CrossRef]
- Choné, X.; Van Leeuwen, C.; Chéry, P.; Ribéreau-Gayon, P. Terroir influence on water status and nitrogen status of non-irrigated Cabernet Sauvignon (Vitis vinifera). Vegetative development, must and wine compositios. S. Afr. J. Enol. Vitic. 2001, 22, 8–15. [Google Scholar]
- Naor, A. Midday Stem Water Potential as a Plant Water Stress Indicator for Irrigation Scheduling in Fruit Trees. Acta Hortic. 2000, 537, 447–454. [Google Scholar] [CrossRef]
- Tuteja, N. Abscisic Acid and Abiotic Stress Signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef]
- Rolt, A.; Cox, L.S. Structural basis of the anti-ageing effects of polyphenolics: Mitigation of oxidative stress. BMC Chem. 2020, 14, 1–13. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants 2018, 10, plx051. [Google Scholar] [CrossRef] [PubMed]
- Mansori, M.; Chernane, H.; Latique, S.; Benaliat, A.; Hsissou, D.; El Kaoua, M. Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris L.). Environ. Boil. Fishes 2014, 27, 1689–1698. [Google Scholar] [CrossRef]
- Mansori, M.; Chernane, H.; Latique, S.; Benaliat, A.; Hsissou, D.; El Kaoua, M. Effect of seaweed extract (Ulva rigida) on the water deficit tolerance of Salvia officinalis L. Environ. Boil. Fishes 2016, 28, 1363–1370. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef]
- El-Boray, M.S.; Mostafa, M.F.M.; Salem, S.E.; Sawwah, O.E. Improving Yield and Fruit Quality of Washington Navel Orange Using Foliar Applications of Some Natural Biostimulants. J. Plant Prod. 2015, 6, 1317–1332. [Google Scholar] [CrossRef][Green Version]
- Hammad, S.A.; Ali, O.A. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agric. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A Review on Potential Plant-Based Water Stress Indicators for Vegetable Crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Beekwilder, J.; Contartese, V.; Karlova, R.; Bertea, C.M. The application of a biostimulant based on tannins affects root architecture and improves tolerance to salinity in tomato plants. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vats, S. Biotic and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Bhanu, A.N. Assessment of Genetic Diversity in Crop Plants—An Overview. Adv. Plants Agric. Res. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Liu, F.; Andersen, M.N.; Jacobsen, S.-E.; Jensen, C.R. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ. Exp. Bot. 2005, 54, 33–40. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, E.A.; El-Ansary, D.O.; Mattar, M.A. Effects of Water Stress and Modern Biostimulants on Growth and Quality Characteristics of Mint. Agronomy 2019, 10, 6. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Jiang, M. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef]
- Amjad, M.; Akhtar, S.S.; Yang, A.; Akhtar, J.; Jacobsen, S.-E. Antioxidative Response of Quinoa Exposed to Iso-Osmotic, Ionic and Non-Ionic Salt Stress. J. Agron. Crop. Sci. 2015, 201, 452–460. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Effect of water stress on antioxidant systems and oxidative parameters in fruits of tomato (Solanum lycopersicon L., cv. Micro-tom). Physiol. Mol. Biol. Plants 2013, 19, 363–378. [Google Scholar] [CrossRef]
- Zgallaï, H.; Steppe, K.; LeMeur, R. Effects of Different Levels of Water Stress on Leaf Water Potential, Stomatal Resistance, Protein and Chlorophyll Content and Certain Anti-oxidative Enzymes in Tomato Plants. J. Integr. Plant Biol. 2006, 48, 679–685. [Google Scholar] [CrossRef]
- Khaleghi, E.; Arzani, K.; Moallemi, N.; Barzegar, M. Evaluation of Chlorophyll Content and Chlorophyll Fluorescence Parameters and Relationships between Chlorophyll a, b and Chlorophyll Content Index under Water Stress in Olea europaea cv. Dezful. Int. Sch. Sci. Res. Innov. 2012, 6, 2108–2111. [Google Scholar]
- Parida, A.K.; Dagaonkar, V.S.; Phalak, M.S.; Umalkar, G.V.; Aurangabadkar, L.P. Alterations in photosynthetic pigments, protein and osmotic components in cotton genotypes subjected to short-term drought stress followed by recovery. Plant Biotechnol. Rep. 2007, 1, 37–48. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef]
- Hossain, M.A.; Wani, S.H.; Bhattacharjee, S.; Burritl, D.J.; Tran, L.S.P. Drought stress tolerance in plants. Physiol. Biochem. 2016, 1, 20–31. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Peripolli, M.; Dornelles, S.H.B.; Lopes, S.J.; Tabaldi, L.A. Application of biostimulants in tomato subjected to water deficit: Physiological, enzymatic and production responses. Rev. Bras. Eng. Agric. Ambient. 2021, 25, 90–95. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Giannakoula, A.; Ilias, I. The effect of water stress and salinity on growth and physiology of tomato (Lycopersicon esculentum Mil.). Arch. Biol. Sci. 2013, 65, 611–620. [Google Scholar] [CrossRef]
- Murtic, S.; Oljaca, R.; Murtic, M.S.; Koleska, I.; Muhic, A. Enzymatic Antioxidant Responses to Biostimulants in Cherry. J. Anim. Plant Sci. 2019, 29, 1664–1672. [Google Scholar]
- Amini, A.; Mirkhani, N.; Pourfard, P.P.; Ashjaee, M.; Khodkar, M.A. Thermo-economic optimization of low-grade waste heat recovery in Yazd combined-cycle power plant (Iran) by a CO2 transcritical Rankine cycle. Energy 2015, 86, 74–84. [Google Scholar] [CrossRef]
- Libutti, A.; Cammerino, A.R.B.; Monteleone, M. Risk Assessment of Soil Salinization Due to Tomato Cultivation in Mediterranean Climate Conditions. Water 2018, 10, 1503. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of Antioxidant Potential of Seaweed Extracts for Enrichment of Convenience Food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef]
- EL Boukhari, M.E.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in Seaweed Extract Based Biostimulants: Manufacturing Process and Beneficial Effect on Soil-Plant Systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).