Abstract
Packing density has a significant influence on the outcome of ensiling forage. In this study, we aim to investigate the effect of packing density on the ensiling properties, microbiome, and in vitro digestibility of barley silages. Barley was ensiled in polyethylene drum silos (30 L) with respective densities of 600, 650, 700, and 750 kg/m3 fresh matter (FM), and stored for 60 days. The bacterial communities, fermentation quality, and in vitro digestibility were analyzed. Fresh barley had a low count of lactic acid bacteria (LAB, 104 cfu/g of FM), and Lactobacillus was nearly undetectable (<1%). Increasing the packing density decreased the pH and the content of ammonia nitrogen (NH3-N), ethanol, neutral detergent fiber (NDF), and acid detergent fiber (ADF) of barley silage (p < 0.05), and increased in vitro digestibility of dry matter, NDF, ADF, and DM recovery (p < 0.05). A higher packing density decreased the abundances of Enterobacter (from 47.4% to 35.4%) and Clostridium (from 13.5% to 3.8%), and increased the abundance of Lactobacillus (from 1.8% to 17.0%). Thus, packing density positively correlated with Lactobacillus (p < 0.05) but negatively correlated with Enterobacter (p < 0.05). The pH and the content of ethanol were positively correlated with Enterobacter (p < 0.05) but negatively correlated with Lactobacillus (p < 0.05). In conclusion, the density of 750 kg/m3 FM resulted in the highest silage quality of the densities tested.
1. Introduction
Barley is suitable for making silage in China [1]. Whole-crop barley silage has attracted greater interest among growers, and may be an attractive substitute for maize, oat, or wheat silage since use of another crop can improve yield stability [2]. Animal performance varies with the quality of whole-crop cereal silage [3]. Therefore, improving barley silage quality may enhance animal productivity and subsequent farm profitability [4]. The maturity stage at harvest has a pronounced effect on the whole-crop barley quality and its subsequent silage quality because of an increase in DM content and the change of nutrient composition during plant development [5]. The maturity stage at harvest can affect packing density of the barley silage, as advanced maturity of barley results in stiffer stems which are harder to pack in silos [5]. In addition, barley stalks are hollow, similar to those of rice straw [6], which are not easy to compact. Because of the above limitations, maintaining anaerobic conditions within the silos through proper packing density is a key factor affecting silage quality [7,8,9]. Lower packing density during silage production may result in high residual oxygen in silage during early stages of fermentation, and then lead to the proliferation of harmful microorganisms [10], thereby, decreasing the dry matter (DM) recovery and nutrients [11]. These problems may be resolved through increasing compaction or density at ensiling, which could help to exclude air from the silo, thus, ensuring an anaerobic environment in which nutrients are preserved [12].
In recent years, the need to investigate the effects of density on silage quality has become clear. Storage losses have been shown to decrease by 50% by increasing the packing density from 160 to 320 kg/m3 DM [7]. A minimum packing dry weight density of 225 kg/m3 is recommended for corn silage [13]. Clearly, higher packing density can improve the quality of silage for animal performance, and most studies have shown this effect can be achieved with maize silage and forage paddy rice [14]. There are limited studies that have been conducted with barley silage. Therefore, it is essential to determine the effects of silage density on the microbial composition, fermentation, and nutritional composition of barley silage.
Recently, next-generation sequencing (NGS) has been extensively used to study microbial communities [1]. NGS in the silage ecosystem can provide specific microbial information at the genus level and opportunities for further evaluating the effects of different conditions on silage quality. Therefore, it is necessary to perform more studies on the effects of packing density on bacterial communities of barley silage.
It is hypothesized that low packing density has a negative effect on the quality of barley silage, and that density can influence the bacterial community. The objective of this study was to evaluate the effect of packing density on the ensiling properties, the bacterial microbiome, and in vitro digestibility of barley silages.
2. Materials and Methods
2.1. Barley Silage Preparation
The barley (new variety 18PJ-03 with low fiber content) was planted in the experimental field of the Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, located in Huhhot, Inner Mongolia, China (40°75′ N, 111°67′ E). The crop was sown on 30 March 2019 with a five-row planter using 0.15 m row spacing and a seeding rate of 225 kg/ha. At the tillering stage, the field was fertilized with 75 kg/ha of urea (N > 46%). No herbicides were applied to the crop. The fresh barley forage was harvested from 3 sites in the same field (200 × 20 m), and used as replicates. The sampling sites were set at the center line of the field and were as follows: 1 site was in the center, and 2 sites were 30 m away from the short side of the two sides. On 30 June 2019, the crop had reached the soft dough stage of maturity and was manually harvested at a height of 10 cm from ground level at three sites as replicates. Approximately 100 kg of fresh barley forage from each site was collected. Immediately, the collected materials were chopped into pieces of approximately 1–2 cm with a manual guillotine cutter (Type: 8012, Deli Group Co. Ltd., Ningbo, China). Fresh forage barley was sampled from each site, collected in sterilized bags in an icebox, and then immediately transported to the laboratory to determine the pH, microbial communities and populations, and chemical composition (Table 1).

Table 1.
pH, microbial population, and chemical composition of fresh barley (n = 3).
The chopped barley from each site was mixed well and divided into equal portions for four treatments. Twelve polyethylene drum silos (three for each treatment) with a volume of 30 L (diameter 310 mm, height 550 mm, Foshan Aosidun Packing Products Co., Ltd., Foshan, China) were used for the experiment, as they were used by Jungbluth et al. [15]. The treatments were 600 (S600), 650 (S650), 700 (S700), and 750 kg/m3 (S750) of fresh barley. The corresponding amount of forage barley for each treatment was weighed on a precision scale and the material compressed with a hydraulic press (Type: TF-32, Shandong Qufu Taifeng Machinery Equipment Co. Ltd., Qufu, China). After the filling was completed, the top of each silo was sealed using a sealing cover with a sealing rubber and a plunger ring and kept at an ambient temperature in a laboratory. After 60 days of ensiling, the polyethylene drum silos were weighed and the topmost 5 cm of silage discarded, as described by Anésio et al. [11]. The remainder of the silage in each silo was emptied, and placed in a plastic container sterilized with ethanol, and mixed thoroughly for homogenization. The homogenate was split into portions for analysis of microbial counts, chemical composition, fermentation profile, and bacterial community.
2.2. Silage Extract Preparation
Twenty-gram samples of either fresh barley or silages from each site were placed in 180 mL of double distilled sterile water, homogenized in a LW-09 sterile homogenizer (Shanghai Jingxin Industrial Development Co., Ltd., Shanghai, China) for 100 seconds, and then strained using 4 layers of cheesecloth. After the fresh barley and silage extracts were prepared, they were used immediately for pH analysis and sampled for microbiological analysis, and thereafter passed through a 0.22 μm filter and stored at −20 °C for fermentation pattern analysis.
2.3. Microbial Population
Fresh barley and silage extracts were used to assess the populations of lactic acid bacteria (LAB), aerobic bacteria (AB), coliform bacteria, and yeast using the method of plate count, as reported by Cai et al. [16]. The fresh barley and silage extracts constituted the first dilution, followed by 10-fold serial dilutions, diluted with a disinfected 8.50 g/L of NaCl solution. From three consecutive dilutions (10−3 to 10−6), triplicate 100 μL aliquots were plated on selective agar media for various microbes. The following culture media (Guangzhou Huankai Microbial Science and Technology Co., Ltd., Guangzhou, China) were used to isolate and enumerate various microorganisms: De Man Rogosa Sharpe agar for LAB, nutrient agar for AB, violet-red bile agar for coliform bacteria, and potato dextrose agar for yeast. The plates for enumerating LAB were placed in an anaerobic box (Anaerobic box; C-31, Mitsubishi Gas Chemical Co., INC., Tokyo, Japan) and incubated at 37 °C for 48 h in a general incubator (GP-01, Huangshi Hengfeng Medical Instrument Co., Ltd., Huangshi, China). The plates for coliform bacteria and aerobic bacteria were incubated at 37 °C for 48 h in aerobic conditions in the same general incubator [17,18]. The plates for yeasts were cultivated at 32 °C for 48 h in another general incubator [18]. Microorganism population numbers were expressed as colony forming units (cfu) per gram of fresh matter (FM).
2.4. Bacterial Diversity Analysis
2.4.1. DNA Isolation and PCR Amplification
DNA was extracted as reported by Guan et al. and Mu et al. [17,19] using a commercial DNA isolation kit (E.Z.N.A.® Omega Bio-tek, Norcross, GA, USA), in line with the manufacturer’s instructions. A NanoDrop2000 (Thermo Fisher Scientific Inc., Wilmington, DE, USA) was used after purification to measure the purity and concentration of DNA. Bacteria of the 16S rRNA (explicitly the V3–V4 regions) were amplified with primers 338F and 806R in an ABI GeneAmp® 9700 PCR thermocycler (98 °C for 2 min, followed by 25 cycles of 98 °C for 30 s, 50 °C for 30 s, 72 °C for 60 s, and a final extension at 72 °C for 5 min; ABI, Union City, CA, USA).
2.4.2. Sequencing and Data Analysis
The Illumina HiSeq 2500 platform (Illumina Inc., San Diego, CA, USA) was used to amplify PCR products. Raw reads were assembled using FLASH (version 1.2.7) after they were purified and quantified, and low-quality reads were excluded using Trimmomatic (Version 0.33). Chimeric sequences and final effective tags were removed and identified, respectively, using UCHIME software (version 4.2). An overall similarity of 97% of operational taxonomic units (OTUs) was generated using Usearch software. Mothur (version v.1.30) was used to determine the alpha diversity indices including Shannon, Chao1, Simpson, and Good’s coverage. The community structure (phylum and genus) was taxonomically classified using the Ribosome Database Project (RDP) and the Silva database (70% confidence threshold). A correlation network was drawn with OmicStudio tools. Pearson correlation analyses of the fermentation, bacterial composition, and density were performed with Vegan software.
2.5. Fermentation Quality, Chemical Composition, and In Vitro Digestibility
In the fresh barley and silage, the pH was determined using a glass electrode pH meter (PB-10, Sartorius, Gottingen, Germany). Lactic acid (LA), acetic acid (AA), propionic acid (PA), and butyric acid (BA) were analyzed with a high-performance liquid chromatography (HPLC) system with a pump (SIL-20AC, Shimadzu Co., Ltd., Kyoto, Japan), auto sampler (L-2200, Shimadzu Co., Ltd., Kyoto, Japan), SPD-20A diode array detector (DAD, 210 nm, Shimadzu Co., Ltd., Kyoto, Japan), and a column (Shodex RSpak KC-81, Kyoto, Japan). The analytical conditions were as reported by Adesogan et al. [20]. Ethanol was determined by the refractive index detector of the HPLC with a column (Shodex SUGAR Pak KS-801, Showa Denko K.K., Kawasaki, Japan) applying ultrapure water as the mobile phase with a 1.0 mL/min flow rate at 50 °C.
Approximately 100 g of each sample were dried in an oven at 65 °C for 48 h to measure the DM content, which was not corrected for losses of volatile compounds. The same dried samples that were used to determine the DM content were ground and passed through a 1 mm screen using a grinder (ZM-2248, Guangzhou zhenmai machinery equipment Co., Ltd., Guangzhou, China) for compositional analysis and in vitro digestibility assessment. Samples were combusted in a muffle furnace at 550 °C for 5 h to measure crude ash content. The weight lost after ashing was used as a measure of organic matter (OM) content. The rate of DM recovery was computed using the weight and DM content before and after ensiling. The ammonia nitrogen (NH3-N) and the total nitrogen (TN) concentrations were measured with a Kjeltec 8400 autoanalyzer (Foss Co. Ltd., Hillerød, Denmark), in line with the Kjeldahl method. The crude protein (CP) content was computed by multiplying TN content by 6.25. The content of acid detergent fiber (ADF) was sequentially analyzed after extraction of neutral detergent fiber (NDF), as stated by Van Soest et al. [21], using a ANKOM2000 fiber analyzer (Ankom Technology, Macedon, NY, USA). Sodium sulphite and heat-stable amylase were used for the NDF analytical procedure. The ether extract (EE) was determined as the Soxhlet method [22].
The method reported by Contreras-Govea et al. [23] and Kowalski et al. [24] was applied to determine in vitro digestibility of DM (IVDMD), ADF (IVADFD), and NDF (IVNDFD) of silages using the Daisy II System (ANKOM Technology Corp., Fairport, NY, USA). Rumen fluid from four Inner Mongolia semi-fine wool sheep (wethers) before morning feeding was collected by gastric tube. On a DM basis, the animals had been fed 400 g of Leymus chinensis hay, 40 g of alfalfa hay, and 240 g of a concentrate with 65% corn grain, 25% soybean meal, 10% wheat bran, and supplemental minerals and vitamins. Subsamples of each treatment (0.50g) were heat-sealed within F57 fiber bags (pore size = 38–45 µm, Beijing First Beef Cattle Info and Tech Research Center) using an impulse sealer (model #FS-200, Guangzhou Lingli Machinery Equipment Co., Ltd., Guangzhou, China). Rumen fluid was strained via 4 layers of cheesecloth and blended with buffer 1:2 (v/v). The buffer was prepared following the method reported by Menke et al. [25]. Fiber bags (F57) containing samples were incubated for 48 h, following the recommended Ankom procedures. After incubation, the fiber bags containing residues were gently rinsed with cold tap water, and then oven-dried for 48 h before DM, NDF, and ADF were measured. Then, the IVDMD, IVADFD, and IVNDFD were calculated according to the differences in their respective weight before and after incubation.
2.6. Statistical Analyses
Treatment effects were analyzed by one-factor analysis of variance with the general linear model (GLM) procedure of SAS (SAS System for Windows, version 9.1.3; SAS Inst. Inc., Cary, NC, USA) for all parameters. Analysis was performed for parts of the alpha diversity data and data of the microbial population, chemical composition, fermentation quality, and in vitro digestibility. The linear, quadratic, and cubic effects of the packing density of barley silage were tested by polynomial comparison method. Microbial population data were transformed to log10.
3. Results
3.1. Characteristics of Fresh Barley
The DM content of fresh barley was 337 g/kg of FM. The contents of CP, ADF, and NDF were 99, 236, and 443 g/kg DM, respectively. Low LAB counts (104 cfu/g of FM) and higher populations of agar-culturable aerobic bacteria, yeast, and coliform bacteria (105–107 cfu/g of FM) were observed in the fresh barley (Table 1).
3.2. Bacterial Community of Fresh Barley and Silages
A total of 70,103 reads were acquired by high-throughput sequencing in fresh barley (Table 2). The number of reads for silage treatments ranged from 68,226 to 78,074 with an average of 73,874. For all treatments, the average coverage was >99%. There was a tendency for fresh barley to have higher OTU and Simpson index values than silages (p < 0.10) (Table 2).
Table 2.
Number of reads and coverage of 16S rDNA and measures of bacterial diversity generated from the DNA data for fresh barley and silages of various densities (n = 3).
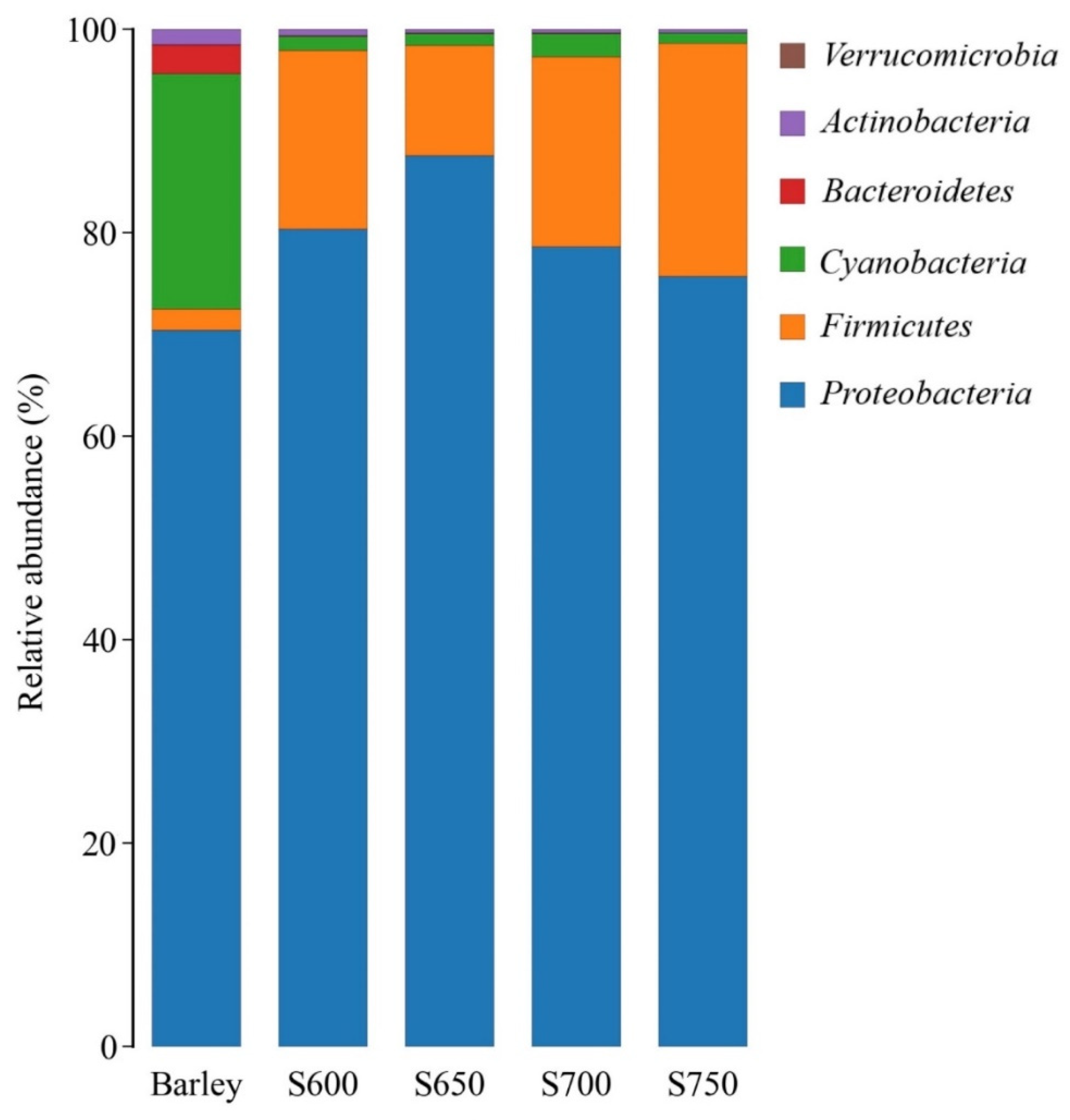
The prominent phyla of fresh barley were Proteobacteria (70.4%) and Cyanobacteria (23.1%) (Figure 1). Proteobacteria (ranging from 75.7% to 87.6%) and Firmicutes (ranged from 10.8% to 22.9%) dominated the bacterial community in barley silages after 60 days of ensiling.
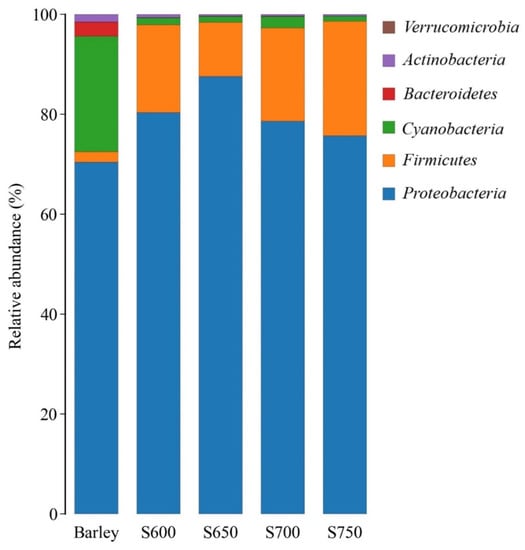
Figure 1.
Bacterial community found in fresh barley and silages at the phylum level (n = 3). S600, S650, S700, and S750 represent silages densities of 600, 650, 700, and 750 kg/m3, respectively.
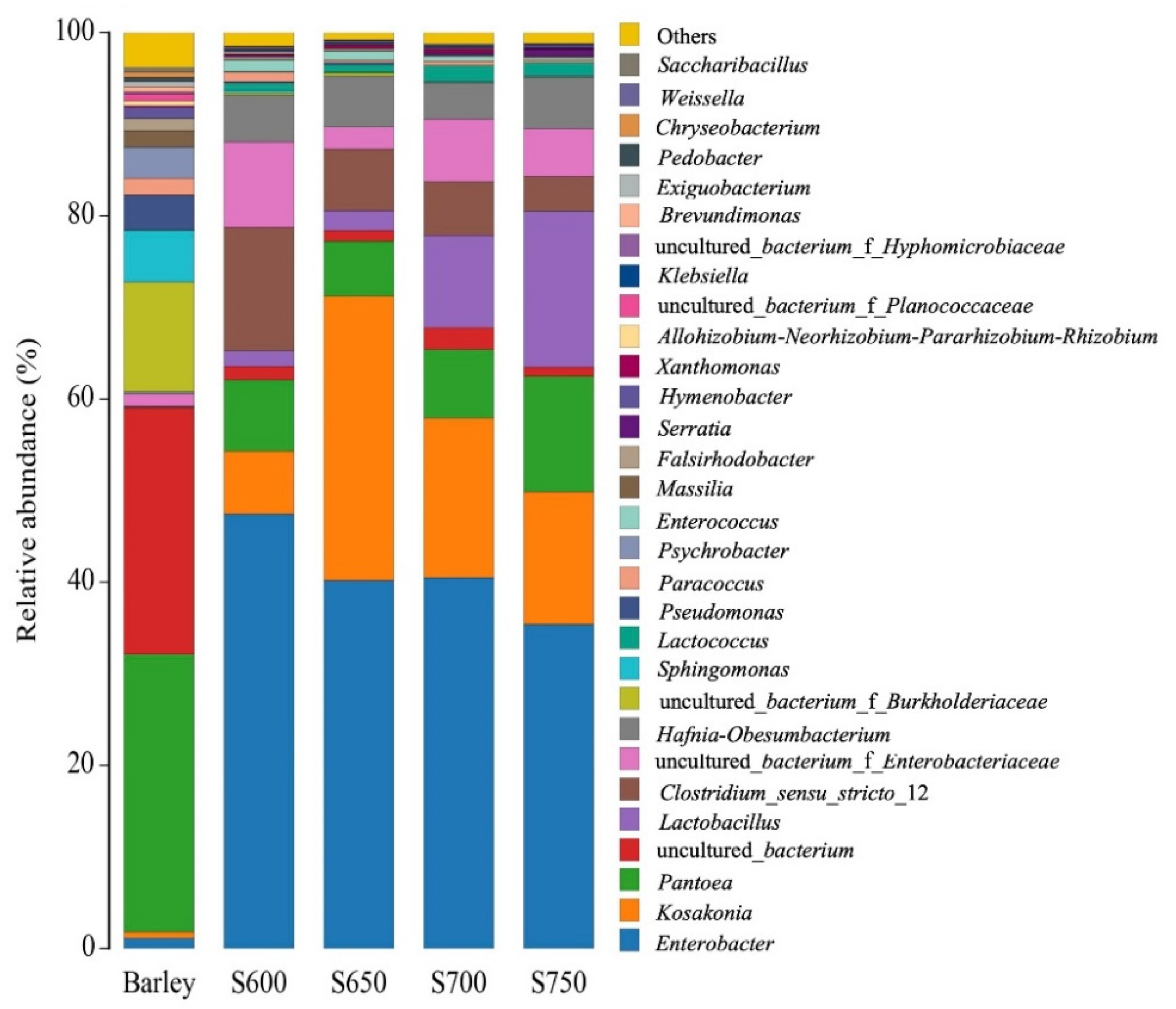
The bacterial composition in the fresh barley and silages at genus levels is displayed in Figure 2. In the fresh barley, the dominant bacterial genera were Pantoea (30.4%), followed by uncultured bacterium (26.9%), and uncultured bacterium_f_Burkholde (12.0%). However, Lactobacillus was almost undetectable in fresh barley (<1%). After fermentation, the genus Enterobacter predominated in all silages. As silo density increased, the relative abundance of Enterobacter decreased from 47.4% to 35.4% and that of Clostridium decreased from 13.5% to 3.8%. In contrast, Lactobacillus abundance increased to 17.0% in the S750 silage as compared with 1.8% in S600 silage. Kosakonia and Pantoea were found in high abundance (6.9–31.0% and 6.0–12.6%, respectively) in all silages. Uncultured bacterium from Enterobacteriaceae was found in S600, S700, and S750 silages, and the corresponding abundance was 9.3%, 6.7%, and 5.2%. Hafnia-Obesumbacterium was found only in S650 and S750 silages, and the corresponding abundance was 5.5% and 5.6%.
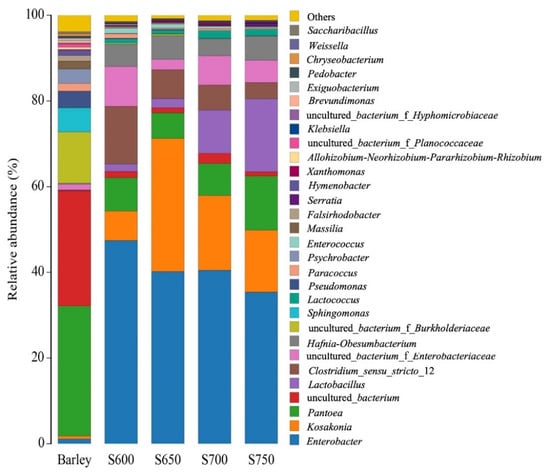
Figure 2.
Bacteria community analysis of the top 30 bacterial genera from fresh barley and silages (n = 3). S600, S650, S700, and S750 represent silages densities of 600, 650, 700, and 750 kg/m3, respectively. Y-axis shows the genus; X-axis shows sample name.
3.3. Microbial Population, Fermentation Quality, and Chemical Composition of Silage
After 60 days of ensiling, the LAB, AB, and yeast counts went from 106 to 107 cfu/g of FM in all treatments (Table 3). Packing density did not affect the counts of LAB and AB (p > 0.05). The numbers of yeast showed a quadratic change (p < 0.05). Furthermore, coliform bacteria were undetectable in all silages. Barley silages at a packing density of 750 kg/m3 had a lower (p < 0.05) pH and NH3-N concentration compared with S600 and S650. However, no differences in LA, AA, and PA contents were observed among all silages. No detectable propionic acid was present in silage from the 750 kg/m3 density treatment. There was no butyric acid detected in all silages (Table 3). The organic matter and DM contents showed no clear changes associated with silage densities. However, DM recovery increased linearly with an increase in packing density (p < 0.05). The contents of NDF and ADF in the S750 treatment were significantly lower than those in lower packing density (<650 kg/m3) treatments (p < 0.05). The concentration of ethanol was lower in the S700 and S750 treatments as compared with the lower packing density silages (p < 0.05). The IVDMD increased linearly with an increase in packing density (p < 0.05); with an increase in packing density, the IVNDFD and IVADFD showed linear and quadratic increasing trends (p < 0.05).
Table 3.
Microbial population, fermentation quality, chemical composition, IVDMD, IVNDFD and IVADFD of barley silage after 60 days of ensiling (n = 3).
3.4. Correlation Analysis Heat Map of the Bacterial Community, and Quality Indices, Correlation Networks among Main Bacterial Genera and Density
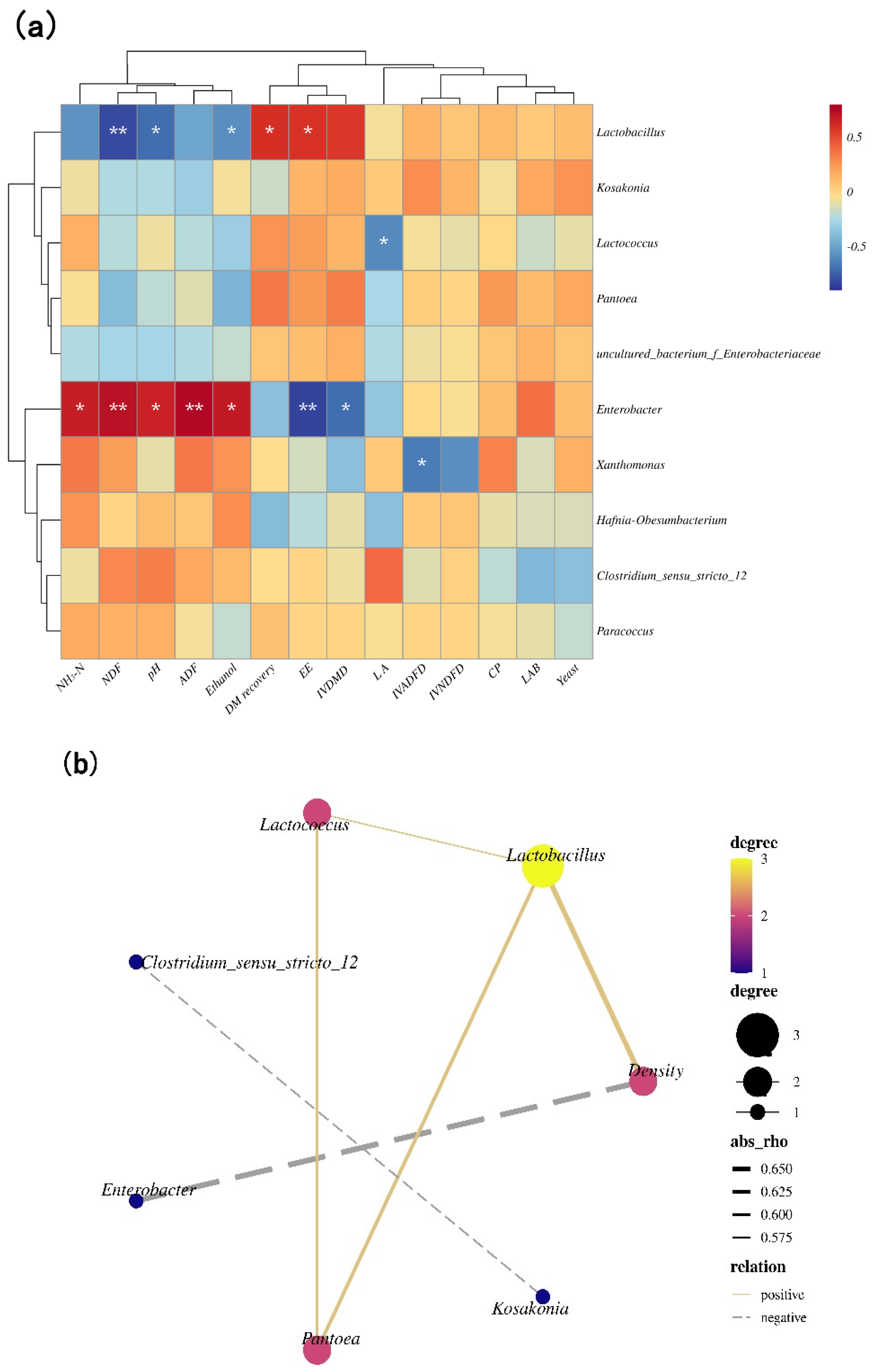
Enterobacter was positively associated with pH and the contents of NH3-N and ethanol (p < 0.05). Lactobacillus had a negative association with pH and ethanol concentrations (p < 0.05). Lactococcus and Xanthomonas had negative associations with LA concentrations and the IVADFD, respectively, (p < 0.05) (Figure 3a). The density had positive correlations with Lactobacillus and negative correlations with Enterobacter (p < 0.05) (Figure 3b).
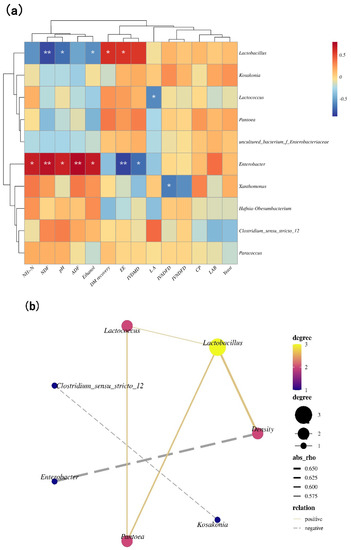
Figure 3.
(a) Correlation clustering and heat map analysis of top 10 genera with pH, LA, ethanol, NH3-N, LAB, yeast, CP, NDF, ADF, EE, DM recovery, IVDMD, IVNDFD, and IVADFD; (b) correlation network among main bacterial genera (top 10) and density (b) (n = 12). The values presented by colors in the heat map correspond to the Pearson correlation coefficient r, which ranged between −0.5 and 0.5, where r < 0 indicates a negative correlation (blue), r > 0 indicates a positive correlation (red), ‘*’ represents p < 0.05, and ‘**’ represents p < 0.01. In (b), absolute value of correlation coefficient > 0.5 and p-value < 0.05.
4. Discussion
4.1. Microbial Composition of Barley before and after Ensiling
In this experiment, the fresh materials have higher OTU and Simpson index values, which indicate that fresh barley has higher bacterial richness and bacterial diversity than the silage treatments, and this might be due to the acidic and anaerobic microenvironment [26]. Firmicutes are gram-positive bacteria of which a majority can degrade cellulose, protein, and starch [27]. Proteobacteria are a main phylum of gram-negative bacteria. Proteobacteria were the most abundant phyla in fresh barley, exceeding 70% in this study. In alfalfa silage, McGarvey et al. [28] stated that the most common bacteria present were Firmicutes (acid hydrolytic microorganisms) and Proteobacteria. In the present study, Proteobacteria and Firmicutes were the dominant phyla in all barley silage density treatments. These results contrast with those of Liu et al. [1], who reported that, after ensiling for 60 days, >99% of the bacteria present were in the phylum Firmicutes. LAB inoculation was used in their study, which likely accounted for the observed dominance of Firmicutes. Therefore, it is beneficial to apply exogenous LAB for making barley silage, especially inhibiting the growth of undesirable microorganisms, including the phylum Proteobacteria.
Lactobacillus is generally more abundant in silages treated with microbial inoculants [29], whereas the abundance of Lactobacillus of barley silage in the present study was only 1.8–17.0%. This result is possibly due to the limited occurrence (<1%) of Lactobacillus in fresh barley, similar to woody forage, which generally has a low abundance of Lactobacillus [30]. The abundance of Lactobacillus on fresh forage is also related to environmental factors such as weather, temperature, and sunlight. Du et al. [30] reported that Enterobacter sp. (34.69%) was the most abundant species in wilted paper mulberry silages without inoculant. Similarly, herein, Enterobacter was the most abundant genus in all silages, and higher density-treated silage had more Lactobacillus, but less Clostridium and Enterobacter. The probable reason is that increased packing density decreased oxygen penetration and increased the population of epiphytic aerobic bacteria on the surface of fresh barley, which led to a rapid depletion of oxygen in silo at the beginning of ensiling. Consequently, this was beneficial to the proliferation of Lactobacillus, since species of Lactobacillus are competitive and can thrive in anaerobic conditions and low pH and suppress the growth of undesirable microorganisms, including Clostridium and Enterobacter [19,31].
4.2. Fermentation Quality, Microbial Counts, DM Recovery, Chemical Composition, and In Vitro Digestibility of Barley Silages
The pH value is a basic value for evaluating silage fermentation. To obtain silage of an excellent quality, a large enough lactic acid-producing bacterial population is required to induce a rapid drop in pH [15]. In the present study, no additional lactic acid bacteria additives were used. Therefore, the differences in the pH values were mainly attributed to the differences in packing density. In the present study, the silages with a packing density of 750 kg/m3 had a lower pH (4.25) and NH3-N content than S600 and S650, which indicates lower proteolytic activity. This result substantiated previous results by Braman et al. [32] on maize silage (ensiling density of 174–344 kg of DM/m3), and by Sucu et al. [7] on maize silage (ensiling density of 168–216 kg of DM/m3) and sorghum silage (ensiling density of 132–178 kg of DM/m3). In addition, we found that lactic acid bacteria populations in all silages increased to 107 cfu/g FM, thus indicating that all silages enabled complete or almost complete fermentation. Zhang and Yu [33] evaluated how the ensiling density (500 and 600 kg/m3 FM) affected the Leymus chinensis silage quality. Their study indicated that lactic acid content was lower, while the coliform bacteria populations and the butyric acid contents were higher, in low-packing density silage (500 kg/m3 FM). However, in this present study, no significant differences were observed in lactic acid contents in different packing density silages. Moreover, the coliform bacteria and butyric acid were undetectable in our experimental silages. This was likely because different forages, ensiling methods, and densities were used [34]. The counts of agar-culturable aerobic bacteria in all silages decreased to 106 cfu/g FM as compared with the fresh barley, but their counts did not vary with packing density treatments. Yeasts can carry out both aerobic respiration and fermentation [31]. The fermentation process produces lactic acid, which may serve as an energy source for yeast survival [35]. The growth of yeasts under low pH can be inhibited by weak acids [35]. This combination of factors likely explains the differences in yeast counts of fresh barley and silages, and accounts for the quadratic changes in yeast population with increasing packing density in this study. Kung et al. [34] reported that the fermentation of silage dominated by Clostridium commonly had a high concentration of propionic acid (>0.3–0.5% DM). Indeed, Clostridium occurrence decreased with increasing silage density, and the propionic acid concentration in the present study tended to decrease with decreasing Clostridium abundance from 3.2 to 1.7 g/kg DM in higher density-treated silage, and was undetectable at the highest silage density.
Ruppel et al. [36] stated that packing density was negatively correlated with DM losses during storage in farm-scale silos. In the present study, packing density had a linear effect on the DM recovery, which was consistent with results of Sucu et al. [7], who stated that the DM recovery of tightly packed silage was higher than that of loosely packed silage (p = 0.06). The possible reason behind this result is probably that high ensiling density limits air osmosis and cuts down the oxidation losses during ensiling [7]. However, this result was contrary to the conclusions of Gallo et al. [37], who discovered that packing density (132 and 186 kg DM/m3) had no effect on DM loss. This is likely because different forages, ensiling methods, and densities were used. In the present study, the contents of DM and OM were unaffected by packing density, possibly because densities were so high that even higher packing density had no effect [38]. In this study, the contents of NDF and ADF in fresh whole-crop barley were very low compared with those reported by Kim et al. [39] at a similar maturity stage. Variety difference could account for the differences observed in NDF and ADF. In the present study, the contents of NDF and ADF in the high packing density (750 kg/m3) treatment were significantly lower than those in lower packing density (<650 kg/m3) treatments. This could be explained by the higher DM recovery in higher packing density-treated silages, which indirectly decreased relative amounts of structural carbohydrates. Moreover, the acid solubilization of hemicellulose displayed in an acid environment (low pH) [40] in higher packing density-treated silages should also be considered, and further investigation is needed. The quality of the S700- and S750- treated silage also benefited from a decreased ethanol concentration, which showed differences in yeast counts among the treatments [26]. Our ethanol results agree with those of Sucu et al. [7] and Shao et al. [9]. Recently, high DM and fiber digestibility have been considered to be indispensable for good forage quality [41]. In the present study, higher packing density (>700 kg/m3) increased the IVDMD of barley silage, which may be attributed to the high fermentation quality of barley silages with a higher packing density that contributes to conservation of digestible nutrients [42]. In addition, the higher IVDMD of silages might be due to the lower NDF and ADF contents [43,44]. According to Tian et al. [6], with an increase in DM losses, the in vitro digestibility of DM (r = −0.496, p < 0.01), NDF (r = −0.377, p < 0.01), and ADF (r = −0.544, p < 0.01) in rice straw silage all decreased. Similarly, in this experiment, the higher packing density silages showed consistently higher IVNDFD and IVADFD as compared with the lowest packing density silage (600 kg/m3), which is favored by the higher DM recovery in the higher packing density silages. Further investigation is needed.
4.3. The Relationship between the Microbial Community and Quality Indices
The relative abundances of bacteria have an important correlation with the final fermentation product [30]. It is well known that Lactobacillus can grow under low pH and completely anaerobic conditions during late stages of fermentation [19]. Thus, Lactobacillus was negatively associated with pH values in this study. Herein, pH values and concentrations of NH3-N and ethanol were positively correlated with Enterobacter (Figure 3a). This indicates that these factors are likely affected by Enterobacter populations, possibly due to the fact that Enterobacter competes with LAB in silage for available water-soluble carbohydrates and produces more ammonia nitrogen, AA, 2,3-butanediol, succinic acid, and endotoxins, thus decreasing silage nutritional value and reducing palatability [17,19]. Another reason is that ethanol may be produced by Enterobacter [45]. The above results were consistent with the result of Ogunade et al. [46]. The density had negative correlations with Enterobacter (Figure 3b), indicating that higher packing density decreased Enterobacter abundance due to the low pH and anaerobic conditions. The LA concentrations were negatively correlated with Lactococcus, indicating that increasing the content of LA resulted in a low pH (4.25–4.45) that potentially inhibited the growth of Lactococcus, which has low tolerance to acidity and grows poorly at pH values less than 4.8 [31]. Xanthomonas is a gram-negative, rod-shaped bacteria and a common plant pathogen. In this experiment, Xanthomonas showed a negative correlation with the IVADFD, and this result may be attributable to the degradation of cellulose fibers and hemicellulose of the cell wall by some species of Xanthomonas for the acquisition of plant nutrients [47], but further investigation is needed to test this hypothesis.
5. Conclusions
Increasing the packing density decreased NH3-N and ethanol production, while increasing DM recovery and in vitro digestibility of barley silage, which could be related to a change in the Lactobacillus community of the silage. Hence, 750 kg/m3 FM resulted in the highest silage quality of the densities tested.
Author Contributions
Conceptualization, L.S. and Y.X. (Yanlin Xue); Data curation, Y.X. (Yanlin Xue); Formal analysis, X.W.; Investigation, L.S., N.N., C.W., Y.X. (Yanzi Xiao), G.Y., S.L. and Y.X. (Yanlin Xue); Methodology, Z.L. (Ziqin Li); Project administration, F.Y.; Resources, Z.L. (Zhiping Liu); Software, X.L.; Validation, N.N. and C.W.; Writing—original draft, L.S. and Y.X. (Yanlin Xue); Writing—review and editing, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31772674), the Inner Mongolia Science and Technology Plan (2021GG0068) and the Inner Mongolia Program of High-end Foreign Experts Introduction.
Institutional Review Board Statement
The animal care and experimental procedures were handled strictly in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health of China. The protocols were approved by the China Agricultural University Animal Care and Use Committee (Permit No. AW20110202-6-2).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Bikel, D.; Ben-Mair, Y.A.; Yoav, S.; Ran, S.; Idan, R.; Yuri, P.; Shamay, J.; Joshua, M.; Roi, B.D. Nutritive value for high-yielding lactating cows of barley silage and hay as a substitute for wheat silage and hay in low-roughage diets. Anim. Feed Sci. Tech. 2020, 265, 114498. [Google Scholar] [CrossRef]
- Wallsten, J.; Hatfield, R. Cell wall chemical characteristics of whole-crop cereal silages harvested at three maturity stages. J. Sci. Food Agric. 2016, 96, 3604–3612. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Kim, D.H.; Amanullah, S.M.; Kim, S.C.; Song, Y.M.; Kim, H.Y. Effects of bacterial inoculants and cutting height on fermentation quality of barley silage. J. Korean Soc. Grassl. Forage Sci. 2014, 34, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, E. Effects of plant species, stage of maturity and additive on the feeding value of whole-crop cereal silage. J. Sci. Food Agric. 2007, 87, 789–801. [Google Scholar] [CrossRef]
- Tian, J.P.; Xu, N.X.; Liu, B.Y.; Huan, H.L.; Gu, H.R.; Dong, C.F.; Ding, C.L. Interaction effect of silo density and additives on the fermentation quality, microbial counts, chemical composition and in vitro degradability of rice straw silage. Bioresour. Technol. 2020, 297, 122412. [Google Scholar] [CrossRef]
- Sucu, E.; Kalkan, H.; Canbolat, O.; Filya, I. Effects of ensiling density on nutritive value of maize and sorghum silages. R. Bras. Zootec. 2016, 45, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Z.; Zhang, F.F.; Ma, C.H.; Li, F.F.; Wang, S.L. Effect of compaction on nutritional quality and aerobic stability of silage corn after opening silos. Trans. Chin. Soc. Agric. Eng. 2018, 34, 300–306. [Google Scholar] [CrossRef]
- Shao, T.; Wang, T.; Shimojo, M.; Masuda, Y. Effect of ensiling density on fermentation quality of Guineagrass (Panicum maximum Jacq.) silage during the early stage of ensiling. Asian Austral. J. Anim. 2005, 18, 1273–1278. [Google Scholar] [CrossRef]
- Driehuis, F.; Oude, E.S. The impact of the quality of silage on animal health and food safety: A review. Vet. Q. 2000, 22, 212–216. [Google Scholar] [CrossRef]
- Anésio, A.H.C.; Santos, M.V.; da Silva, L.D.; Silveira, R.R.; Braz, T.G.S.; Pereira, R.C. Effects of ensiling density on chemical and microbiological characteristics of sorghum silage. J. Anim. Feed Sci. 2017, 26, 65–69. [Google Scholar] [CrossRef]
- Johnson, L.M.; Harrison, J.H.; Davidson, D.; Mahanna, W.C.; Shinners, K.; Linder, D. Corn silage management: Effects of maturity, inoculation, and mechanical processing on pack density and aerobic stability. J. Dairy Sci. 2002, 85, 434–444. [Google Scholar] [CrossRef]
- Bruning, D.; Gerlach, K.; Wei, K.; Sudekum, K.H. Effect of compaction, delayed sealing and aerobic exposure on maize silage quality and on formation of volatile organic compounds. Grass Forage Sci. 2018, 73, 53–66. [Google Scholar] [CrossRef]
- Kawamoto, H.; Yamaguchi, H.; Komatsu, T.; Tanaka, O.; Oshibe, A. Effect of chopping and high-density ensiling on the silage fermentation of forage paddy rice (Oryza sativa L.). Jpn. J. Grassl. Sci. 2009, 54, 323–327. [Google Scholar] [CrossRef]
- Jungbluth, K.; Trimborn, M.; Maack, G.; Büscher, W.; Li, M.; Cheng, H.; Cheng, Q.; Sun, Y. Effects of three different additives and two different bulk densities on maize silage characteristics, temperature profiles, CO2 and O2–dynamics in small scale silos during aerobic exposure. Appl. Sci. 2017, 7, 545. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics and aerobic deterioration of silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- Xu, D.M.; Ding, W.R.; Ke, W.C.; Li, F.H.; Zhang, P.; Guo, X.S. Modulation of metabolome and bacterial community in whole crop corn silage by inoculating homofermentative Lactobacillus plantarum and heterofermentative Lactobacillus buchneri. Front. Microbiol. 2018, 9, 3299. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef]
- Adesogan, A.T.; Krueger, N.; Salawu, M.B.; Dean, D.B.; Staples, C.R. The influence of treatment with dual purpose bacterial inoculants or soluble carbohydrates on the fermentation and aerobic stability of bermudagrass. J. Dairy Sci. 2004, 87, 3407–3416. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Official Methods of Analysis; Association of Official Analytical Chemists: Arlington, VA, USA, 1990.
- Contreras-Govea, F.E.; Muck, R.E.; Mertens, D.R.; Weimer, P.J. Microbial inoculant effects on silage and in vitro ruminal fermentation, and microbial biomass estimation for alfalfa, bmr corn, and corn silages. Anim. Feed Sci. Tech. 2011, 163, 2–10. [Google Scholar] [CrossRef]
- Kowalski, Z.M.; Ludwin, J.; Górka, P.; Rinne, M.; Weisbjerg, M.R.; Jagusiak, W. The use of cellulase and filter bag technique to predict digestibility of forages. Anim. Feed Sci. Tech. 2014, 198, 49–56. [Google Scholar] [CrossRef]
- Menke, K.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.A.; Dong, Z.H.; Li, J.F.; Shao, T.A. Microbial community dynamics and their contributions to organic acid production during the early stage of the ensiling of Napier grass (Pennisetum purpureum). Grass Forage Sci. 2020, 75, 37–44. [Google Scholar] [CrossRef]
- Romero, J.J.; Zhao, Y.; Balseca-Paredes, M.A.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Laboratory silo type and inoculation effects on nutritional composition, fermentation, and bacterial and fungal communities of oat silage. J. Dairy Sci. 2017, 100, 1812–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol 2013, 114, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol 2011, 34, 235–241. [Google Scholar] [CrossRef]
- Du, Z.; Sun, L.; Chen, C.; Lin, J.; Yang, F.; Cai, Y. Exploring microbial community structure and metabolic gene clusters during silage fermentation of paper mulberry, a high-protein woody plant. Anim. Feed Sci. Tech. 2020, 114766. [Google Scholar] [CrossRef]
- Ávila, C.L.S.; Carvalho, B.F. Silage fermentation-updates focusing on the performance of micro-organisms. J. Appli. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braman, W.L.; Bryan, K.A.; Kurtz, J.E. 055 Effect of fermented corn silage density and bacterial inoculants on corn silage pH and fermentation end products. J. Anim. Sci. 2016, 94, 27. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.; Zhao, M.; Yu, Z. Lactic acid bacteria strains for enhancing the fermentation quality and aerobic stability of Leymus chinensis silage. Grass Forage Sci. 2016, 71, 472–481. [Google Scholar] [CrossRef]
- Kung, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.O.; Spoelstra, S.F. Microbiology of ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; Agron. Monogr. ASA, CSSA and SSSA: Madison, WI, USA, 2003; pp. 31–94. [Google Scholar]
- Ruppel, K.A.; Pitt, R.E.; Chase, L.E.; Galton, D.M. Bunker silo management and its relationship to forage preservation on dairy farms. J. Dairy Sci. 1995, 78, 141–153. [Google Scholar] [CrossRef]
- Gallo, A.; Bernardes, T.F.; Copani, G.; Fortunati, P.; Giuberti, G.; Bruschi, S.; Bryan, K.A.; Nielsen, N.G.; Witt, K.L.; Masoero, F. Effect of inoculation with Lactobacillus buchneri LB1819 and Lactococcus lactis O224 on fermentation and mycotoxin production in maize silage compacted at different densities. Anim. Feed Sci. Tech. 2018, 246, 36–45. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Kim, D.H.; Amanullah, S.M.; Lee, H.J.; Joo, Y.H.; Han, O.K.; Adesogan, A.T.; Kim, S.C. Effects of different cutting height on nutritional quality of whole crop barley silage and feed value on Hanwoo Heifers. Asian Austral. J. Anim. 2016, 29, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Dong, Z.H.; Li, J.F.; Chen, L.; Bai, Y.F.; Jia, Y.S.; Shao, T. Effects of lactic acid bacteria and molasses on fermentation dynamics, structural and nonstructural carbohydrate composition and in vitro ruminal fermentation of rice straw silage. Asian Austral. J. Anim. 2019, 32, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Baah, J.; Beauchemin, K.A. Conservation, fiber digestibility, and nutritive value of corn harvested at 2 cutting heights and ensiled with fibrolytic enzymes, either alone or with a ferulic acid esterase-producing inoculant. J. Dairy Sci. 2015, 98, 1214–1224. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Jiang, Y.; Ling, Q.; Na, N.; Xu, H.; Vyas, D.; Adesogan, A.T.; Xue, Y. Effects of adding pre-fermented fluid prepared from lucerne or red clover on fermentation quality and in vitro digestibility of the ensiled wilting-forages. Agriculture 2021, 11, 454. [Google Scholar] [CrossRef]
- Yin, G.; Bai, C.; Sun, J.; Sun, L.; Xue, Y.; Zhang, Y.; Zhao, H.; Yu, Z.; Liu, S.; Zhang, K. Fermentation quality and nutritive value of total mixed ration silages based on desert wormwood (Artemisia desertorum Spreng.) combining with early stage corn. Anim. Sci. J. 2017, 88, 1963–1969. [Google Scholar] [CrossRef]
- Xue, Y.L.; Yin, G.M.; Zhao, H.P.; Bai, C.S.; Sun, J.J.; Yu, Z.; Sun, Q.Z. Nutritive value of desert wormwood (Artemisia desertorum Spreng.) silage in mixture with high-moisture maize straw. Grass Forage Sci. 2017, 72, 174–178. [Google Scholar] [CrossRef]
- Bi, C.; Zhang, X.; Ingram, L.O.; Preston, J.F. Genetic engineering of Enterobacter asburiae strain JDR-1 for efficient production of ethanol from hemicellulose hydrolysates. Appl. Environ. Microb. 2009, 75, 5743–5749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Pech Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2017, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Cândido, E.S.; Pereira, J.L.; Quezado-Duval, A.M.; Noronha, E.F.; Krüger, R.H.; Quirino, B.F. Xanthomonas gardneri exoenzymatic activity towards plant tissue. World J. Microbiol. Biotechnol. 2008, 24, 163–170. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).