1. Introduction
Growing plants in a greenhouse provides a more favourable environment compared to an open field due to the control and management of different environmental conditions, such as radiation, temperature, humidity, carbon dioxide, and more precise availability of water and nutrients. Consequently, crops have an increased vegetative development, are of higher quality (colour, firmness, taste, etc.), and have earlier harvests.
Lettuce can be grown both in the open field and in a greenhouse. However, in recent decades, greenhouse lettuce cultivation has become more popular as it extends seasonal availability and improves quality by producing cleaner lettuce and an earlier yield.
Nevertheless, the different materials covering a greenhouse affect the amount and spectral quality of light that passes inside the greenhouse and reaches the plant growth area. Greenhouse plastic cover materials are sensitive to UV radiation, and their exposure to UV radiation for a long period reduces their durability and transparency in the light. Protective agents are added to plastic covers to protect them from senescence. These agents expand the use life of the plastic covers; however, they reduce their transparency in UV radiation [
1]. Consequently, in the commonly used greenhouse plastic covers, transparency in UV radiation is much lower than transparency in photosynthetically active radiation (PAR).
Preventing the entry of ultraviolet radiation into the greenhouse is achieved by choosing covers that are not transparent to this radiation. These covers are defined as UV-block, while the covers that allow UV radiation to enter the greenhouse are characterized as UV-transparent or UV-open. Most plastic greenhouse covers, as well as glass, do not transmit UV-B radiation (280–315 nm). The glass transmits only a part of UV-A radiation (315–400 nm). In the Mediterranean region, plastic materials, and mostly the polyethene variant types, have prevailed as greenhouse-covering materials. Photoselective sheets, which block UV radiation, have also been used as greenhouse covers to control diseases and pests in greenhouses. The absence of ultraviolet radiation prevents the sporulation of several phytopathogenic fungi, such as Botrytis and Alternaria, and reduces the populations of pests that need ultraviolet radiation for their orientation [
2,
3]. Moreover, in recent decades, types of polyethene covers with low to zero UV transmittance (UV-block polyethene) have been developed to control pest populations [
4] and greenhouse crop diseases [
5].
On the other hand, UV-block polyethene films can have a detrimental effect on the accumulation of antioxidant compounds in lettuce [
6]. Ultraviolet radiation and the high density of photosynthetically active radiation increase the concentration of flavonoids in lettuce [
7]. In addition, lettuce grown in an open field contains higher concentrations of flavonols than lettuce grown in a polycarbonate greenhouse [
8] due to the different radiation densities and different wavelengths entering the greenhouse. Polycarbonate sheets absorb a small percentage of photosynthetically active radiation but most of the ultraviolet radiation. Moreover, exposure of red lettuce cultivars to high levels of ultraviolet radiation during cultivation causes reddening of the leaves and increases the concentration of total phenols and main flavonoids (quercetin and cyanidin), as well as phenolic acids [
9]. Consequently, the antioxidant content of the plants is responsible for the intensity of the colouring of various fruits, leaves, and flowers, so ultraviolet radiation is required to initiate the violet colour in some ornamental and vegetable crops, such as eggplant and red-pigmented lettuce [
10].
On the other hand, blocking the entry of ultraviolet solar radiation into the greenhouse favours the faster growth of red lettuce plants [
11]. A greater explanation for the growth inhibition of red lettuce under ambient levels of UV radiation may be the high metabolic cost of photoprotection, such that the plants divert energy produced by photosynthesis to synthesize phenolic compounds [
12]. In a similar experiment, eggplant plants grown in greenhouses with UV-block polyethene achieved higher height, larger leaf size, and higher yields compared to plants grown in a greenhouse with UV-transparent polyethene [
10].
In order to achieve high yield and antioxidant content in red lettuce, Tsormpatsidis et al. [
12] proposed initial cultivation in a UV-block greenhouse and transfer to a UV-transparent greenhouse a few days before harvest. However, transferring soil-grown red lettuce plants shortly before harvest is impossible, while transferring soilless-grown plants is difficult and requires a lot of labour or an expensive automated installation.
Therefore, there is a need to develop a technique that will allow the use of UV-block polyethene sheets with simultaneous ultraviolet radiation availability to plants at their critical growth stages (in which ultraviolet radiation is necessary). Ultraviolet radiation is not currently used in horticulture, but its effects on plant development and secondary metabolism could be implemented (especially UV-A in small amounts) for the production of high-quality compact plants [
13]. However, little research has been completed on the effect of pre-harvest supplemental lighting containing the UV-A spectrum on red lettuce in a greenhouse production system [
14]. The present study aimed to investigate the use of UV-block greenhouse covers with the possibility of artificially adding UV radiation before the harvest of red lettuce to achieve high nutritional quality.
2. Materials and Methods
Two experiments were performed in two greenhouses located at the experimental field of the Agriculture Department of the University of Peloponnese, Kalamata, 5 m above sea level (latitude 37°03′40″ N, longitude 22°03′41″ E). Each greenhouse had an area of 108 m
2 (6 × 9 m), with a height of 1.7 and 3 m in the gutter and ridge, respectively. During hot days, both greenhouses were ventilated and cooled by fan-pad system [
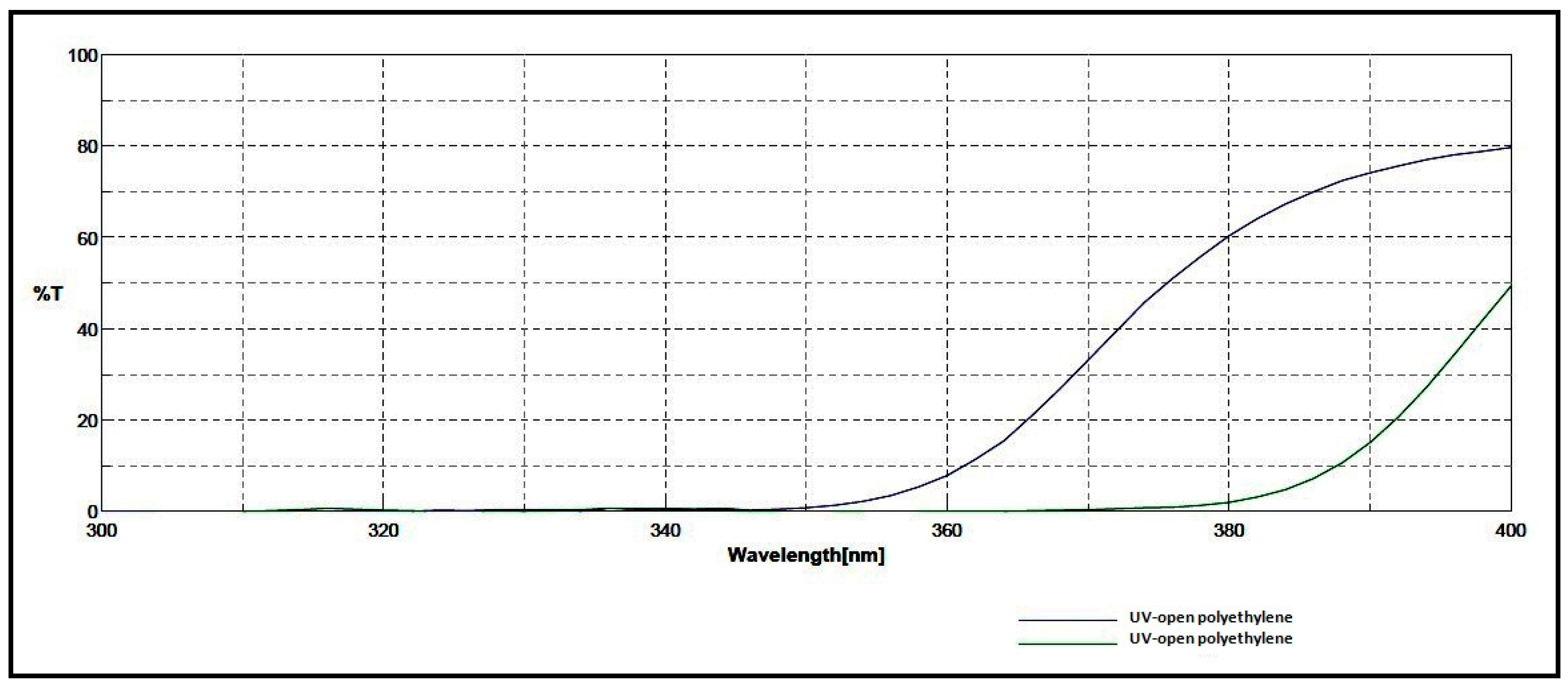
15]. The two greenhouses were covered with polyethene films of different UV transmittance. One greenhouse was covered by a common polyethene film (TUV3965), which allowed a large part (26%) of UV-A radiation to pass through, while the second greenhouse was covered with a UV-block polyethene film (TUV3957) that allowed only 4.5% of outside UV-A radiation to transmit through it.
Figure 1 shows the spectral transmission in UV radiation of each polyethene film used in the current study accordingly to film supplier (plastikakritis S.A.). This transmittance difference to UV radiation of the used polyethene films was confirmed by measurements with portable instruments (HD2102.1 and LP 471 UV-A probe, Delta-OHM, ITALY). According to the polyethene film supplier’s laboratory, the transparency of UV-open and UV-block film at PAR was 86.5% and 85%, respectively.
2.1. Plant Material and Growing Conditions of 1st Experiment
In the first experiment, the effect of different levels of ultraviolet radiation on the growth and quality characteristics (leaf colour, antioxidant accumulation) of red lettuce was studied. Thus, the experiment consisted of two treatments; the first treatment included the red lettuce plants grown in the UV-open greenhouse and the second treatment included the plants grown in the UV-block greenhouse.
Red lettuce plants (Redino Lollo rosso from Geoponiki S.A.) were grown in a deep floating hydroponic system. Tanks (105 cm × 52 cm × 40 cm) were used, which were sealed by a black–white polyethene sheet. Each tank was filled with 60 L of nutrient solution suitable for lettuce. EC and pH values of the nutrient solution in the solution tanks were adjusted to 2.1 dS m−1 and 5.6, respectively. The composition of the lettuce nutrient solution was as follows: 8.47 mM K+, 3.81 mM Ca2+, 1.27 mM Mg2+, 0.85 mM NH4+, 16.10 mM NO3−, 1.00 mM H2PO4−, 1.19 mM SO42−, 35 μM Fe, 20 μM Μn, 4 μM Zn, 5 μM B, 0.50 μM Cu, and 0.50 μM Mo.
Moreover, an air pump was placed in each tank to oxygenate the nutrient solution. A polystyrene plate (100 cm × 50 cm × 2 cm) with ten planting sites was placed on the surface of the nutrient solution in each tank, in which ten lettuce plants were transplanted, respectively.
The transplantation of lettuce seedlings took place on 17 March 2020, in the stage of 3–4 leaves, and 7 tanks with growing plants were placed in each greenhouse. Two plants from each greenhouse and tank (14 plants from each greenhouse) were randomly sampled 42 days after transplanting for growth measurements (plant weight, number of leaves per plant), leaf colour determination, antioxidants analysis, and nutritional analyses.
On the same date, three lettuce planting tanks were moved from the UV-block greenhouse to the UV-open greenhouse to determine whether a change in UV intensity could trigger the production of antioxidants and plant colouration. Seven days later, ten plants were randomly sampled from the three removed tanks in the UV-open greenhouse and from the tanks that were in the UV-open greenhouse from the beginning of the experiment.
2.1.1. Leaf Colour Determination
To determine the leaf colour of red lettuce, two intermediate leaves of each sample plant (14 plants from each greenhouse) were used, and the measurement was made 1–2 cm from the margins of the leaf circumference. Colour was measured using a compact colorimeter Minolta CR-400 (Minolta, Osaka, Japan). The hue angle [tan
−1(b/a)] and chroma index [(a
2 + b
2)
1/2] were determined from parameters L, a, and b. The hue angle was represented on a 360° polar chart, where 0° and 360° represent red colour, while 90°, 180°, and 270° represent yellow, green, and blue, respectively [
16].
2.1.2. Extraction of Phytochemicals for Measurements of Total Phenolics (TP), Total Flavonoids (TF), and Total Antioxidant Capacity (TAC)
Leaf tissues, which were used for antioxidants analysis, were frozen immediately and stored in a freezer (−25 °C) until analysis. The extraction of phenolic compounds was carried out according to [
17] with some modifications. A sample of approximately 1 g of frozen tissue was homogenized in an Ultra-Turrax (T25, IKA Labortechnik, Germany) with 10 mL of cold 80% (
v/
v) methanol. The crude extract was placed in a supersonic bath (Elma, Transsonic 420) at 4 °C for 20 min, centrifuged at 4000 rpm for 6 min, and the supernatant was collected. The extraction was repeated twice, and the extracts were collected. After centrifugation, the supernatant was assessed for total phenols (TP), total flavonoids (TF), and total antioxidant capacity (TAC) determination, as described below.
Total phenols content was determined by the Folin–Ciocalteu assay method according to [
18]. To 3.95 mL distilled water, 50 μL of extract was added and agitated thoroughly. After that, 250 μL of Folin–Ciocalteu reagent and 750 μL of 20%
w/
v Na
2CO
3 were added and thoroughly mixed. The intensity of blue colour developed was recorded on a spectrophotometer (Heλios γ, Unicam, UK) after 2 h at 760 nm. The results were expressed as milligrams of gallic acid equivalents per gram of fresh weight.
Total flavonoid content was determined using aluminium chloride (AlCl
3) according to [
17]. To 2 mL distilled water 500 μL of extract (1:1 diluted) was added and 150 μL 5%
w/
v NaNO
2. After 5 min 150 μL of 10%
w/
v AlCl
3 was added. After a further 6 min in the reaction mixture, 1 mL of 1 N NaOH was added. Finally, the reaction mixture was diluted to 1.2 mL with water, and the absorbance was measured at 510 nm on a spectrophotometer (Heλios γ, Unicam, UK). The results were expressed as mg of catechin equivalents per gram of fresh weight.
The antioxidant capacity of lettuce leaves was evaluated in the supernatant produced by the extraction of phenolic compounds using the DPPH assay based on the method described by [
19]. For the DPPH assay, 2 mL of 0.1 mM DPPH freshly prepared solution was added to 0.1 mL of the methanolic extract, and the absorbance was measured at 517 nm after a 60 min period in the dark. The scavenging capacity of the extracts was expressed in μmol Trolox equivalents per g fresh weight (μmol TE g
−1).
2.1.3. Nutrient Status
Leaf samples were oven-dried at 70 °C to constant weight and were grounded for mineral analysis after ashing at 500 °C and extraction with 1N HCl solution. The concentrations of K
+, Ca
2+, and Mg
2+ were determined by atomic absorption spectrophotometry (Varian, SpectrAA-200, Australia). Total nitrogen was determined by means of Kjeldahl digestion using a Gerhard Vapodest 30 apparatus [
20].
2.2. Plant Material and Growing Conditions of 2nd Experiment
Red lettuce seedlings (Redino Lollo rosso from Geoponiki S.A.), in the stage of 2–3 true leaves, were transplanted on 25 August 2020 in a deep floating hydroponic system, similar to the 1st experiment. In the UV-block greenhouse, 14 growth plant tanks were installed, and each tank had 10 lettuce plants. Four similar plant tanks were installed in the UV-open greenhouse. On the red lettuce plants grown in the UV-block greenhouse, supplemental UV lighting treatments started 26 days after the transplanting and lasted 10 days. Red lettuce plants were sampled randomly before and after supplemental UV lighting to determine the head weight of the plants.
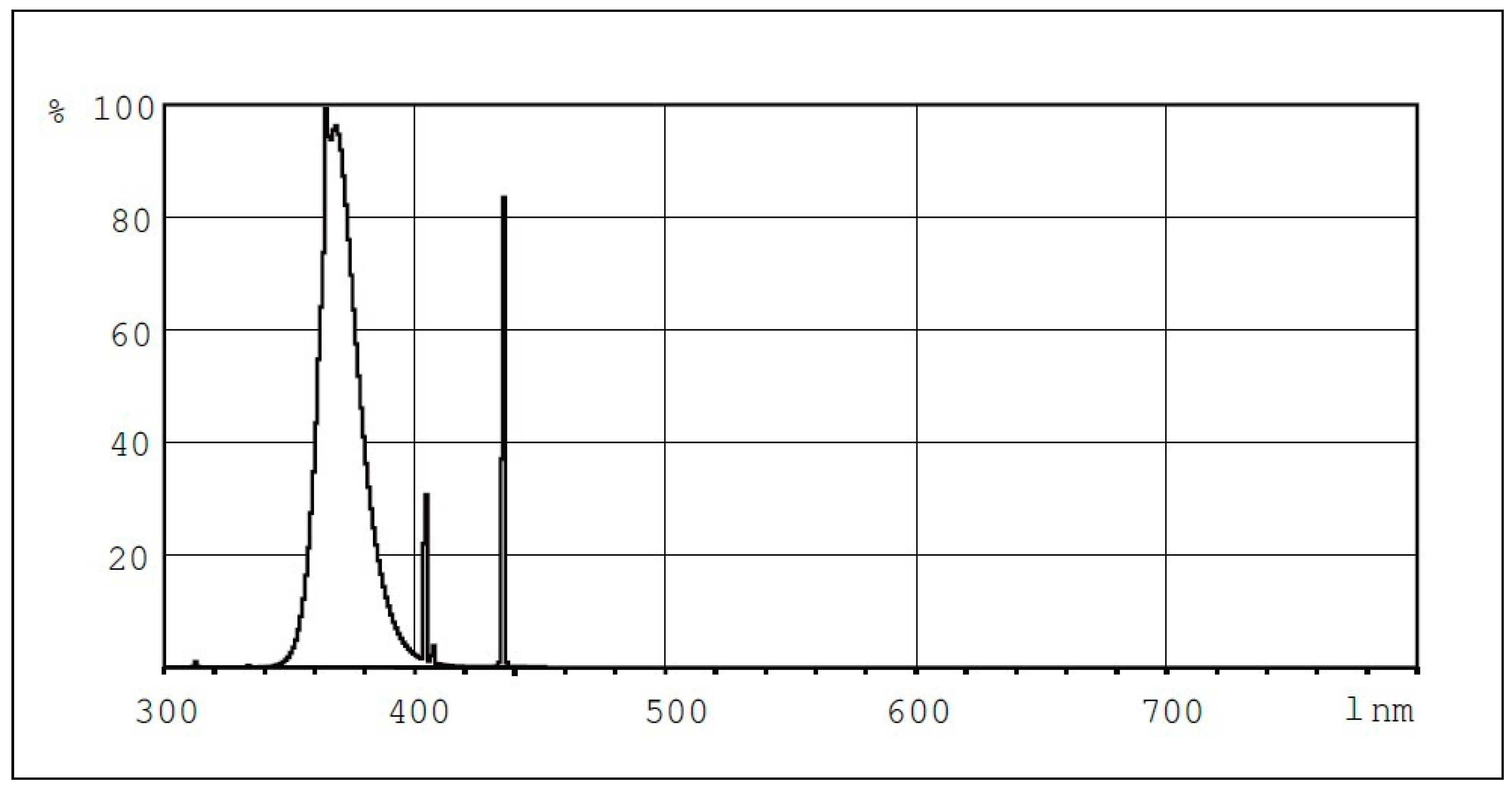
Ultraviolet lamps (Philips, UVA, TLK40W/10R) were used to apply supplemental lighting. These lamps emit in the 350 to 400 nm waveband, with peak at 370 nm and 18.5% efficiency in UV-A radiation. Thus, each lamp emitted 7.4 W of UV-A radiation according to the manufacturer. The UV emission intensity of the lamps was confirmed by measurements with portable instruments (HD2102.1 and LP 471 UV-A probe, Delta-OHM, ITALY).
Figure 2 shows the spectral power distribution of the lamps used.
During supplemental lighting with UV radiation, care must be taken so that the applied dose does not exceed 30 W m
−2 because there is a risk of causing burns to the lettuce leaves. The plants in the UV-block greenhouse were divided into seven different groups (two plant growth tanks per group). Each group received a different treatment with supplemental ultraviolet radiation. With different combinations of operating times and bulb density, each group of plants received 0 kJ m
−2 d
−1, 210 kJ m
−2 d
−1, 265 kJ m
−2 d
−1, 320 kJ m
−2 d
−1, 425 kJ m
−2 d
−1, 530 kJ m
−2 d
−1, or 640 kJ m
−2 d
−1 of UV-A radiation daily. In
Table 1, details are given about calculation of the doses of UV supplemental lighting in kJ m
−2 d
−1 and mmol m
−2 d
−1, number of used lamps per m
2, and the duration of the lighting in each treatment.
After 10 days of supplemental UV lighting, the irradiated plants were compared to the non-irradiated (0 kJ m−2 d−1) control plants. At the end of the supplemental ultraviolet lighting treatments, five plants per treatment were randomly sampled for growth, colour, and antioxidant content measurements. These parameters were determined as in the first experiment.
2.3. Statistical Analyses
The data were subjected to single-factor analyses of variance using the STATISTICA software version 7.0 (StatSoft Inc., Tulsa, OK, USA), and, when a significant F-test was obtained, means were separated using LSD test (p < 0.05).
4. Discussion
In the present study, red lettuces were grown in two greenhouses with different transparency in UV-A radiation, and both greenhouses were not transparent to UV-B radiation. Recent research suggests that different UV transparencies of greenhouse cover do not affect the yield of red lettuce [
21]. However, our results (
Table 2 and
Table 6) agree with earlier research that found a significant increase in the yield of red lettuce in the absence of UV radiation or low UV intensity [
9,
11]. The effects of UV-A on biomass production depend on further environmental factors, as well as on the species or even the genotype, as different responses to UV-A are observed within particular studies [
22]. In the UV-block greenhouse, the red lettuce plants developed more leaves (
Table 6) and gained more weight (
Table 2 and
Table 6) compared to the plants in the UV-open greenhouse. However, their root size was not affected (
Table 2). UV intensity probably affects the distribution of assimilates only in the upper part of the red lettuce plant, and root size can supply water and nutrients to plants with a larger head. Differential partitioning of biomass between shoots and roots in response to UV-A has been reported in various species [
22]. For instance, in all four cultivars of Cucumis sativus studied, UV-A decreased the amount of shoot biomass, although there was no effect on root biomass [
23].
Previous research demonstrated the detrimental effect of the greenhouse covered with UV-block polyethene film in the antioxidant production of red lettuce [
6] and the potential of using UV-transparent covers to increase the beneficial flavonoid content of red leaf lettuce when the crop is grown in greenhouses [
9]. Similar conclusions emerge from the results of our first experiment. The cultivation of red lettuce plants in the UV-block greenhouse significantly reduced the content of total flavonoids and total phenolics, causing a corresponding significant reduction in antioxidant capacity (
Table 4). On the other hand, growing red lettuce plants in a greenhouse with a transparent cover in UV-A radiation allowed them to synthesize flavonoids and phenolics and thus acquire their natural violet colour. The plants that presented high concentrations of the above secondary compounds (
Table 4) also developed more intense violet colouration (
Table 3). The violet colour of red lettuce origins mainly from the composition of phenolic substances, flavonoids, and anthocyanins. It has been suggested that when lettuce is grown under a high level of ultraviolet radiation, it will contain more phenolic compounds, which are produced by the plant as protective agents [
11].
In the current study, from leaf analysis of red lettuces grown in UV-open or UV-block greenhouse, no differences in concentrations of four macronutrients were found (
Table 5). On the other hand, Lee et al. [
24], in an indoor experiment, found higher levels of calcium and magnesium in the leaves of red lettuces that received supplemental UV lighting. This finding indicates that the effect of UV radiation intensity on red lettuce nutrition may be cultivar-dependent, or different effects may be caused by different conditions in the greenhouse and growth chamber. However, little research has been conducted on the effect of UV intensity on the nutritional status of red lettuce, and further research is needed. The authors of [
25] reported that the intensity of UV radiation causes a different response in stomatal conductance among the various plant species. In order to draw safe conclusions about the effect of UV intensity, simultaneous transpiration measurements, plant tissue analyses, and nutrient solution analyses are required.
Red lettuce yield is high in UV-block greenhouses, which is interesting for farmers, but its antioxidant capacity is low. Plant secondary metabolites that are affected by UV light are essential due to their health-promoting properties, and this is important for consumers. Tsormpatsidis et al. [
12] supported that growing plants continuously under a UV-blocking film and then transferring them to a UV transparent film six days before the final harvest showed that high yields and high phytochemical content could be achieved complementarily. In the current study, the transfer of red lettuce plants from UV-block to UV-open greenhouse seven days pre-harvest caused a significant increase in their antioxidant content (total phenols, total flavonoids). However, this increase was not enough to reach the quality of the red lettuces grown in the UV-open greenhouse for the whole period (
Table 4). Probably, more time in a UV-open greenhouse was required than a period of seven days. Ordidge et al. [
26] showed that in the red lettuce Lollo Rosso, total phenolics, anthocyanin, luteolin, and quercetin levels were all raised by changing from a UV-blocking film to a film of low UV transparency and to a film of high UV transparency. Nevertheless, this finding confirms that a pre-harvest supplemental UV lighting treatment of red lettuces grown in a UV-block greenhouse can produce red lettuces with high yield and high quality. Light manipulation is a key environmental control method to increase the functional phytochemical concentrations of plants under controlled environments such as greenhouses and plant factories [
27].
The second experiment of the present study sought to answer the question: what is the appropriate pre-harvest dose of additional supplemental UV light to produce red lettuces in a UV-block greenhouse with equal antioxidant content to those produced in a UV-open greenhouse?
According to our results in
Table 7, the high intensity of supplemental UV radiation significantly decreased the red lettuces’ head weight. However, in an experiment with supplemental UV treatments in a UV-open greenhouse, Lee et al. [
24] reported that UV-A treatments did not affect the fresh or dry shoot biomass of lettuce varieties. However, plant physiological processes are variably affected by light, and the responses are species- and cultivar-dependent [
28]. This indicates that plant response to supplemental UV-A may be cultivar-dependent or affected by pre-treatment acclimatization to UV radiation. In the present study, red lettuces before the starting of supplemental UV lighting were grown in a UV-block greenhouse, while in the study of [
24], they were grown in a UV-transparent greenhouse.
The results of the second experiment indicated that supplemental UV-A lighting stimulated the accumulation of total flavonoids and total phenols. Red lettuce plants which received 210 and 265 kJ m
−2 d
−1 of supplemental UV lighting had no differences in yield, colour, total phenols content, total flavonoids content, and antioxidation capacity in comparison to plants grown without supplemental UV light in the UV-block greenhouse. Our findings are in agreement with that of [
29], who used 11 μmol m
−2 s
−1 of supplemental UV light with a photoperiod duration of 20 h per day, that is, approximately equal to 172 kJ m
−2 d
−1, to improve the quality of red lettuce in a plant growth chamber. They reported that no treatment differences were found for total phenolics, anthocyanins content, and antioxidant capacity. Obviously, a higher dose of UV radiation is required to express the photomorphogenetic effect.
The higher dose of supplemental UV lighting (640 kJ m
−2 d
−1) induced more intense leaf colouration and a higher accumulation of antioxidants in the leaves of red lettuces in comparison to lettuces grown in the UV-open greenhouse. This indicates that this dose can produce the desired effect in a shorter time than 10 days. Lee et al. [
24] improved the quality (antioxidant content) of red lettuces grown in a UV-open greenhouse using 700 kJ m
−2 d
−1 of supplemental UV-A radiation for 5–6 days pre-harvest.
The doses of 425 and 530 kJ m
−2 d
−1 of supplemental UV lighting for 10 days prior to the harvest produced red lettuces of similar quality to that in the UV-open greenhouse. Moreover, the yield in these treatments was 30% higher in comparison to that of red lettuces grown in the UV-open greenhouse. Gómez and Jiménez [
29] supported that end-of-production radiation is a cost-effective, pre-harvest practice that can allow growers to manipulate product quality and thus increase the market value of lettuce without negatively affecting plant growth. From this research, we concluded that adding UV light 10 days prior to harvest is effective for the production of functional phytochemical-rich lettuce. Consequently, we can suggest the cultivation of the red lettuces in a UV-block greenhouse and adding 425 kJ m
−2 d
−1 of supplemental UV-A lighting 10 days prior to harvest. LED lights have great potential to provide supplemental light more efficiently than traditional lights, and their spectrum can be adjusted based on plant growth requirements [
30]. Additionally, LEDs allow potential control of both irradiance and spectra and, when used in fully enclosed environments, photoperiod [
13], so the results of the current study can be applied to both greenhouse and plant factory cultivations. Real-time electricity prices and crop value should be considered in the economic evaluation of the effectiveness of supplemental UV lighting.
In the present study, the addition of supplementary UV lighting was carried out during the light period, that is, during the working hours of the staff in the greenhouse. Although the above-recommended UV dose is not higher than the usual outside UV intensity, care must be taken for the safety of greenhouse staff with supplemental UV lighting. Night-time supplemental lighting in red lettuce was more effective than daytime supplemental lighting, as it resulted in better crop quality [
31]. In addition, the application of short-duration supplemental blue LED light may become more effective in the pigmentation of red lettuce if applied during night breaks [
32]. So, in future research, it is worth studying whether adding the same doses of supplemental UV lighting during the night produces the same results.








{kind=link}
{kind=link}