Abstract
The response of different plant growth regulators on callus induction and regeneration on three sugarcane genotypes (YT-53, CP-77-400, and NSG-59) was evaluated. Different concentrations of 2,4-D alone and in combination with other plant growth regulators (Kinetin and BAP) were used for callus induction. Kinetin along with IBA, BAP and NAA were analyzed with respect to shoot induction, while NAA and IBA were used for root induction. The best callus response in terms of number of days, callus fresh weight, and frequency in YT-53 was observed on MS media provided with 2,4-D (3 mg L−1) + Kinetin (0.5 mg L−1), while in NSG-59 the best response was on MS+2,4-D (4 mg L−1) + Kinetin (0.5 mg L−1), and in CP-77400, MS+2,4-D (5 mg L−1). For shoot induction, 2 mg L−1 Kinetin was found to be the best for YT-53 and NSG-59, while 1 mg L−1 BAP was found to be the best for CP-77-400 in terms of number of days, shoot numbers, and shoot length. The best media for root induction in terms of number of days, root numbers, and root length was 1 mg L−1 NAA + 1 mg L−1 IBA for YT-53, while this was 3 mg L−1 NAA for NSG-59. The highest root frequency and maximum root length in the minimum number of days was observed in CP-77-400 on MS media provided with 2 mg L−1 NAA.
1. Introduction
Sugarcane is a major industrial crop and a prime source of sugar, constituting about 70% of sugar consumed globally. Additionally, it is a source of sustainable bioenergy and many other byproducts [1]. The yield of commercial and productive genotypes of sugarcane are declining due to a lack of alternative technologies for the rejuvenation of genotypes [2]. To solve these multiple problems and to achieve high sugarcane yield, researchers are focusing on biotechnological approaches such as micropropagation [3]. This technology is employed for large-scale production, the rejuvenation of old genotypes, and the production of disease-free and disease-resistant plants. Because of it multiple advantages, this technique has received worldwide commercial application [4].
Each stage of micropropagation requires an efficient protocol for each genotype, because the response of every genotype varies according to the type and concentration of the plant growth regulator [5]. Therefore, for effective micropropagation, it is necessary to assess the most effective plant growth regulator or combination thereof for providing a high rate of growth in a shorter amount of time [6]. Cytokinin and auxins are the most extensively used PGRs for this purpose in plant tissue culture. The proportion of the auxin to the cytokinin determines the type of established culture and the formative pathway of the plant cells [7].
Most micropropagation studies focus on the effect of auxin on callus induction and cytokinin on regeneration [8]. However, the synergistic effect of auxins and cytokinin have rarely been studied. There is limited information available pertaining to the differential morphogenic response of auxins in combination with cytokinin. Moreover, callus and shoot proliferation are very important stages of plant propagation, because of commercial exploitation and genetic fidelity [8]. Different developmental stages of sugarcane under in vitro conditions are also highly dependent upon the interaction of plant growth regulators (PGR). Therefore, it has become a vast field of study, and the need for improved protocols with always be present [9].
Keeping in view the importance and uniqueness of micropropagation technology for each genotype in sugarcane, different combinations of PGR were studied for callus induction and regeneration in three sugarcane (Saccharum officinarum L.) genotypes (YT-53, CP-77-400 and NSG-59), with the aim of providing assistance with the development of a protocol and assessing the different roles of plant growth regulators.
2. Materials and Methods
2.1. Collection of Plant Material
Germplasm of three sugarcane genotypes (YT-53, CP-77-400 and NSG-59) was collected from the Shakerganj Sugarcane Research Institute Jhang. The experiment was conducted in the tissue culture lab Bagh-e-Jinn Park, Lahore, Pakistan.
2.2. Surface Sterilization
The innermost layers of 3–4-month-old sugarcane plant tops were used as explants. Plant material was first cleaned with 95% (v/v) ethyl alcohol and rinsed thoroughly with 15% v/v Robin bleach for about 20 min followed by three rinses with double-distilled sterilized water and kept in autoclaved jars.
2.3. Media Preparation
Cleaned glassware was sterilized at 121 °C under pressure of 15 lb/in2 for 15 min in an autoclave and kept in a culture room. Murashige and Skoog (MS) basal media [10] was used for all different phases. The pH was kept 5.65–5.75 with 1 N HCl and 1 N NaOH. Agar was used for solidification and media was poured in vessels (145 × 25 mm), which were pre-sterilized in an autoclave and covered with polypropylene sheets. Sterilized culture vessels were 1/4 filled with media.
Cultures were kept in the dark for callus induction and a 16 h photoperiod (35 µmol m−2 s−1) was used for regeneration [11]. All the cultures were maintained at 27 ± 2 °C at 80% humidity [12]. Twenty culture vessels were used for each medium, and the whole experiment was repeated twice.
2.4. Inoculation
Inoculation was carried out under aseptic conditions. The working area and tools were cleaned thoroughly with 70% v/v ethanol to achieve aseptic conditions. Ultraviolet light with laminar air flow (LCB-V900) was switched on for 20 min before working. The innermost layer near the nodal part was dissected in 0.5-mm-thick discs and inoculated on culture medium. These cultures were kept in the culture room under suitable conditions.
2.5. Callus Induction
Different media compositions for callus induction with 2,4-D alone and in combination with Kinetin and BAP were prepared. Frequency, weight of calli, and number of days required to induce callus were recorded after 30 days. Calli were subcultured on fresh medium when required. The different media compositions used for callus induction are given in Table 1.

Table 1.
Different Media compositions for callus induction.
2.6. Shoot Induction
For shoot induction, twelve different media with combinations of BAP, Kinetin, IBA, NAA were tested for each genotype. Shoot length, number of shoots per vessel, number of days, and regeneration frequency were recorded after 30 days for each medium. The best regeneration media was used for further experiments. Regenerated shoots were further subcultured on fresh media for further growth and development when required. The various media compositions used for shoot induction are given in Table 2.
Table 2.
Different media compositions for shoot induction.
2.7. Root Induction
For in vitro root induction, half-strength MS media with different concentrations and combinations of NAA and IBA were tested. The number of days required for root induction, as well as average root length, number of roots per plant, and rooting rate were noted after 30 days. The media composition for root induction is given in Table 3.
Table 3.
Media composition for root induction.
2.8. Acclimatization
For acclimatization, plants grown in vitro were taken out from agar and washed with water to remove traces of agar and dipped in antifungal solution for 5 min. Plants were then shifted to pots. For acclimatization, 3 types of substrate (sand, peat, composite) were used. Each pot was 3/4 filled with each substrate, and each pot carried one plant. Pots were kept in a PVC greenhouse at a temperature of 27 ± 2 °C. The survival rate was noted after 15 days. Plants were kept for 60 days for acclimatization.
2.9. Statistical Analysis
Data regarding callus weight, shoot numbers, shoot length, number of roots induced, and root length were subjected to analysis of variance, and the means were tested using Duncan’s Multiple Range Test (p < 0.05). The experiment was conducted using a complete randomized design (CRD).
3. Results
3.1. Callus Induction
All the treatments significantly affected the callus induction, while genotype and the effects of the interaction between treatment and genotype were also significant (Table 4).
Table 4.
Effect of treatments and genotypes and their interaction on callus induction.
For YT-53 only 50% of cultures showed callus induction at 2 mg 2,4-D(T1), but with increasing concentration of 2,4-D in MS media, the callus weight and frequency of callus formation increased. However, these parameters started to decrease when the concentration of 2,4-D exceeded 3.5 mg L−1 (T4).
Callus cultures kept on MS media provided with different concentrations of 2,4-D with BAP showed little increase in callus weight (T7-T14). The synergetic effect of 2,4-D with Kinetin was better than with BAP. The highest callus weight (0.54) and frequency were achieved on MS + 2,4-D (3 mg L−1) + Kinetin (0.5 mg L−1) (T16) after 9 days of explant inoculation (Figure 1). Therefore among 18 tested combinations of media, T16 was selected as being the most suitable for callus induction in sugarcane genotype YT-53. The different stages of callus formation after 10, 20, and 30 days can be seen in Figure 2. The callus of YT-53 was light yellow, granular, and friable.
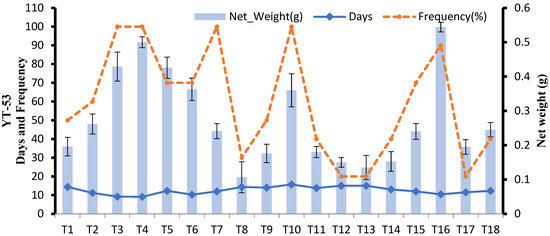
Figure 1.
Effect of different media compositions on callus induction in terms of callus, number of days, and frequency in YT-53.

Figure 2.
Callus of YT-53 on MS + 3 mg L −1 2,4-D + 0.5 mg L−1 Kinetin after 10 (a), 20 (b), and 30 (c) days of inoculation (2×).
In CP-77-400, no callus induction was observed at 2 mg L−1 2,4-D(T1); however, callus weight and frequency started to increase with increasing concentration of 2,4-D in MS media. The maximum callus weight of CP-77-400 was found with 5 mg L−1 2,4-D(T6), where callus induction started after 7 days with 100% frequency. CP-77-400 took fewer days to induce callus than YT-53, but the average weight of callus after 30 days was less than YT-53. Among three tested plant growth regulators, 2,4-D alone induced more callus than BAP and Kinetin in this genotype (Figure 3). Different stages of callus formation after 10, 20, and 30 days can be seen in Figure 4. The callus of CP-77400 was whitish, granular, and friable.
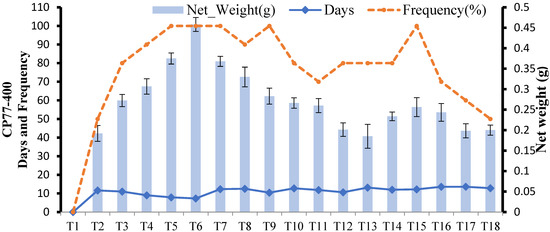
Figure 3.
Effect of different media compositions on callus induction in terms of callus, number of days, and frequency in CP-774-00.

Figure 4.
Callus of CP-77-400 on MS + 5 mg L−1 2,4-D after 10 (a), 20 (b), and 30 (c) days of inoculation (2×).
In NSG-59, callus weight and callus induction frequency started to increase when the concentration of 2,4-D in MS media was increased to 3.5 mg L−1 (T4), but decreased at higher concentrations (T5, T6).
The synergetic effect of 2,4-D and BAP resulted in weights lower than with 2,4-D alone. Kinetin and 2,4-D showed uneven behavior toward callus weight. At lower concentrations of 2,4-D with 0.5 mg L−1 Kinetin, the callus weight of NSG-59 was very low, but the highest mass of callus among all treatments was obtained on MS media supplemented with 4 mg L−1 2,4-d and 0.5 mg L−1 Kinetin (T17) with 80% frequency. At this point, it is important to note that although frequency was higher in T4, callus weight was higher in T17 than in T4. Therefore, T17 was selected as the best callus induction medium among the 18 tested media for NSG-59 (Figure 5). In NSG-59, callus induction was achieved on all tested media except MS + 2,4-D (5 mg L−1) + BAP (0.5 mg L−1 (T10)). Different stages of callus formation after 10, 20, 30 days can be seen in Figure 6. Callus of NSG-59 was cream in color, granular and friable.
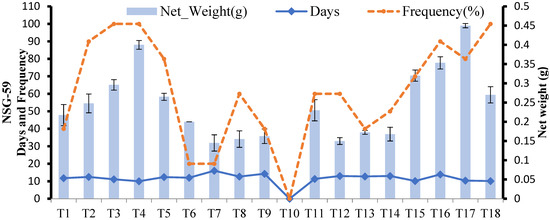
Figure 5.
Effect of different media compositions on callus induction in terms of callus, number of days, and frequency in NSG-59.

Figure 6.
Callus of NSG-59 on MS + 4 mg L−1 2,4-D + 0.5 mg/L Kinetin after 10 (a), 20 (b), and 30 (c) days of inoculation (2×).
The callus weight of YT-53 was highest among the three tested genotypes, while the callus weight of NSG-59 was the lowest after 30 days of inoculation. For further proliferation and growth, callus cultures were subcultured on their respective media. Pairwise comparison is given in Table S1.
Different types of calli were also observed in different media, but mostly light yellow or off white friable, granular callus was observed. Regeneration was achieved only in granular, light yellow and friable calli, while other types (white and non-granular) were non morphogenic and did not show regeneration response.
3.2. Plant Regeneration from Callus Cultures of Sugarcane (YT-53, CP-77-400, NSG-59)
Treatments, genotype, and their interactive effect were significantly (p-value < 0.001) causing variation in shoot length and shoot numbers (Table 5).
Table 5.
Effect of treatments and genotypes and their interaction on shoot length (a) and number of shoots (b) during regeneration.
In YT-53, regeneration potential varies with treatment. Among the 12 tested regeneration media, 1 mg L−1 IBA (T5) and 1 mg L−1 NAA mg L−1 (T6) did not show any shoot induction response whereas MS + 1 mg L−1 BAP + 1 mg L−1 Kinetin (T8), MS + 0.5 mg L−1 BAP + 0.5 mg L−1 IBA (T9), MS + 0.5 mg L−1 BAP + 0.5 mg L−1 IBA + 0.5 mg L−1 NAA (T10), MS + 1 mg L−1 BAP + 1 mg L−1 IBA + 1 mg L−1 NAA (T11) had same shoot induction frequency and average number of shoots per culture vial. Among all 12 tested treatments, MS + 2 mg L−1 Kinetin (T4) showed the highest average no. of shoots per culture vial, with 70 percent frequency after 8.57 days of inoculation and was selected as best shoot regeneration media for YT-53 (Figure 7). Different shoot induction stages are shown in Figure 8.
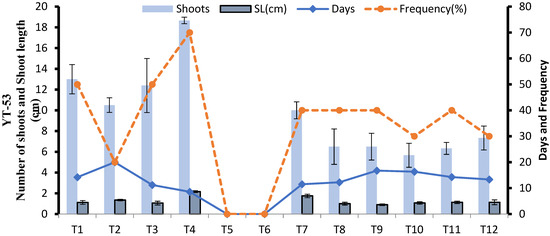
Figure 7.
Effect of different media compositions on shoot induction in terms of shoot length, number of shoots, number of days, frequency in YT-53.

Figure 8.
(a) Stereomicrograph showing shoot induction from callus culture in YT-53 on MS + 2 mg L−1 Kinetin (20×), (b) shoot induction in callus of YT-53 on MS + 2 mg L−1 Kinetin after 10 days (1×), and (c) shoot induction in callus YT-53 on MS + 2 mg L−1 Kinetin after 20 days of inoculation (1×).
In CP-77400 the greatest number of shoots in the minimum number of days (8) from 0.25 g callus was achieved on MS media provided with MS + 1 mg L−1 BAP(T1). No shoot induction was observed on IBA 1 mg L−1 IBA(T5), 1 mg L−1 NAA (T6), 1 mg L−1 BAP + 1 mg L−1 Kinetin (T8) and 0.5 mg L−1 BAP + 0.5 mg L−1 IBA(T9). Shoot length did not vary significantly among different treatments, but the average number of shoots and frequency varied. As T1 showed the highest shoot length and the maximum numbers of shoot with highest frequency in the minimum number of days, T1 was selected as regeneration media for CP-77-400 (Figure 9). Different shoot induction stages are shown in Figure 10.
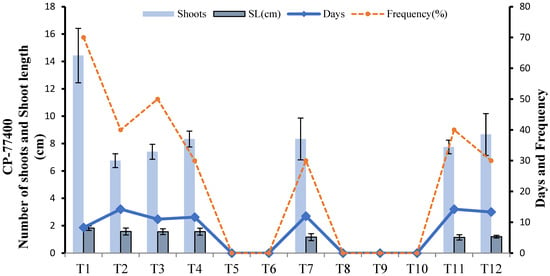
Figure 9.
Effect of different media compositions on shoot induction in terms of shoot length, number of shoots, number of days, frequency in Cp-77400.

Figure 10.
(a) Stereomicrograph showing shoot induction (arrows) from callus culture in CP-77-400 on MS + 5 mg L−1 2,4-D (20×), (b) shoot induction in callus of CP-77-400 on MS + 1 mg L−1 BAP after 10 days, and (c) shoot induction in callus of CP-77-400 on MS + 1 mg L−1 BAP after 20 days of inoculation (2×).
In NSG-59, the maximum number of shoots and longest shoot length per culture vial from 0.25 g callus after 15 days was achieved on T4 (MS + 2 mg L−1 Kinetin). Shoot induction was not observed on T5 and T6 (Figure 11). Different shoot induction and its development stages of NSG-59 can be seen in Figure 12.
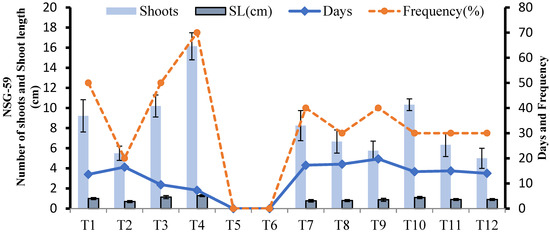
Figure 11.
Effect of different media compositions on shoot induction in terms of shoot length (cm), number of shoots, frequency, and number of days in NSG-59.

Figure 12.
(a) Stereomicrograph showing shoot induction (arrows) from callus culture in NSG-59 on MS + 2 mg/L Kinetin (20×), (b) shoot induction in callus of NSG-59 on MS + 2 mg/L Kinetin after 10 days, and (c) shoot induction in callus of CP-77-400 on MS + 2 mg/L Kinetin after 20 days of inoculation (2×).
YT-53 showed the highest number of shoots and shoot length, while NSG-59 had a higher number of shoots than CP-77-400. The shoots of CP-77-400 were longer than those of NSG-59. The maximum frequency of shoot induction was 70% in the three genotypes. Pairwise comparison of callus weight off all three genotypes is given in Table S1.
It is interesting to note that the genotypes (YT-53, NSG-59) that show the best callus induction with Kinetin also regenerated on media provided with Kinetin. Meanwhile, CP-77400, which showed the best callus induction on 2,4-D alone, regenerated on BAP. The regenerated shoots were shifted and maintained on respective media for 30 days for further growth development.
3.3. Root Induction
The interactive effect of treatment and genotype significantly (p-value < 0.001) influences root length and root numbers (Table 6).
Table 6.
Effect of treatments and genotypes on root length (a) and number of roots (b) during the rooting process.
The highest number of roots per plant in YT-53 was observed on half-strength MS medium supplemented with 1 mg L−1 NAA and 1 mg L−1 IBA (T4), while the highest frequency was recorded on 1/2 MS media provided with 3 mg L−1 NAA (T3). Root length was the same in both treatments (0.9 cm). As the number of days required to induce the root in T4, i.e., 12.33, was less than in T3 i.e., 15.5, T4 was selected as the root induction medium for YT-53 (Figure 13).
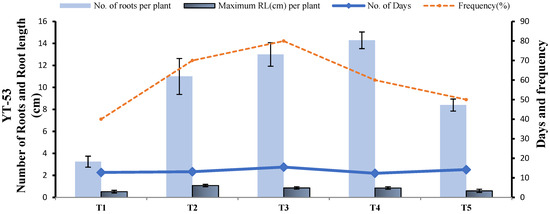
Figure 13.
Effect of five treatments on root induction in terms of root length (cm), number of roots, rooting frequency and number of days in YT-53.
In CP-77-400, 1/2 MS + 2 mg L−1 NAA (T2) induced the highest root frequency and maximum root length in the minimum number of days. Therefore, T2 was selected as the best root induction medium for CP-77-400. In contrast to YT-53, 1/2 MS + 3 mg L−1 NAA did not induce the root in CP-77400 (Figure 14).
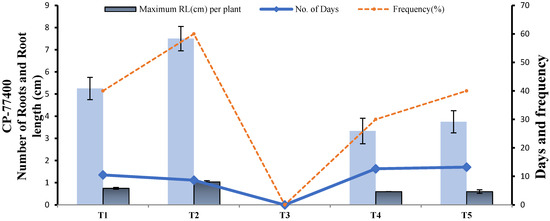
Figure 14.
Effect of different media compositions on root induction in terms of root length (cm), number of roots, rooting frequency and number of days in CP-77400.
NSG-59 showed equal numbers of shoots on 1/2 MS + 3 mg L−1 NAA (T3) and 1 mg L−1 NAA + 1 mg L−1 IBA (T4), but root length and root frequency were higher on 1/2 MS + 3 mg L−1 NAA (T3) than on 1 mg L−1 NAA + 1 mg L−1 IBA (T4). Therefore, T3 was selected as the root induction medium for NSG-59 (Figure 15).
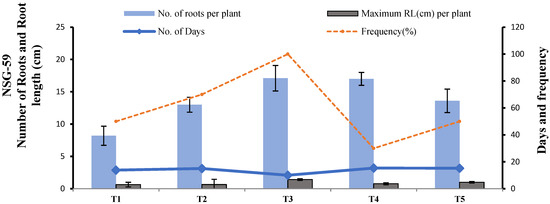
Figure 15.
Effect of different media compositions on root induction in terms of root length (cm), number of roots, rooting frequency and number of days in NSG-59.
Pairwise comparison of all three genotypes for root length and number of roots is given in Table S1.
3.4. Acclimatization
The highest survival rate was achieved on sand. The survival rate of CP-77400 was 60%, while the survival rates of YT-53 and NSG-59 were 50%. All the cultivars were successfully acclimatized to the glasshouse conditions.
4. Discussion
Callus induction in sugarcane is influenced by various factors and genotypic interactions with plant growth regulators, with the endogenous level of hormones reflecting the callus response [13,14].
The synergetic effect of 2,4-D and Kinetin for callus induction has rarely been explored, as callus induction is mostly carried out using different concentrations of 2,4-D alone. Auxin like 2,4-D can return the explant cells to dedifferentiation [15]. Meanwhile, auxin with cytokinin is known to accelerate callus growth and development [16,17]. Cytoplasmic parenchymatous cells have been observed in callus of B. racemosa [18] and Arabidopsis thaliana [19] using the combination of Kinetin and 2, 4-D. It was thought that Kinetin with 2,4-D enhanced the cytoplasm of the vacuolated cell of parenchyma. However, Kinetin alone was unable to induce the cytoplasm of parenchyma cells and remained highly vacuolated. According to Mastuti [20], 2,4-D is required for callus induction, whereas Kinetin promotes its proliferation. Thus, it is obvious that auxin induced the callus, while Kinetin helped with proliferation.
The present study indicates that callus induction and proliferation were also achieved at 4 and 5 mg L−1 of 2, 4-D, but at higher concentrations, its callus induction potency was negatively affected in the YT-53 and NSG-59 genotypes. In the present study, callus induction and proliferation decreased when the concentration of 2, 4-D increased beyond certain limits [21,22].
The highest callus mass being found in YT-53 in the minimum number of days was due to its greater sensitivity towards plant growth regulators compared to the other genotypes [23].
Cytokinin and auxins are the most prevalent hormones in the shoot regeneration process. The addition of synthetic cytokinin changes its endogenous level in explant, making the hormone level unbalanced. This inappropriate type or concentration of cytokinin might lead to epigenetic diversity, thus suppressing callus growth [16]. Therefore, optimization of the an appropriate concentration to obtain the highest callus growth in the minimum amount of time is also very important [24]. Cytokinin also promotes cell growth [25]. Molecular cytokinin signaling is required to stimulate cell proliferation and to differentiate shoot buds. Differential gene expression is required for the formation of organs under in vitro conditions [26]. Molecular studies have revealed the presence of important genes in the cytokinin signaling pathway in plants. Cytokinin receptors interconnect with cell cycles and develop shoot meristem [27].
Plant growth regulator BAP has been reported to be the most efficient cytokinin for shoot induction in sugarcane [28], but in our experiments, it seemed to work only for CP-77-400. The relative expression of GRF1 and GRF5 (growth regulating factors) indicated significant variations during shoot induction in MS media supplemented with BAP and GA3 in Citrus jambhiri Lush. [29].
The best regeneration response in YT-53 and NSG-59 was obtained on MS basal media supplemented with 2 mg L−1 Kinetin. Patel and coworkers [30] observed similar results when working on the regeneration response of callus in three sugarcane cultivars (Co 86002, CoN 07072, CoN 05071). In contrast, Kaur and Kapoor [31] reported no shoot induction in three sugarcane genotypes when using Kinetin in MS medium. This might be due to differences in genetic makeup and the level of endogenous growth hormones. Likewise, in the present investigation, three genotypes behaved differently towards different type of plant hormones. The difference in the response of the three genotypes toward different types and concentrations of plant growth hormones suggests that there are differences in the recognition or action mechanism of Kinetin and BAP, although both are purine-derived cytokinin.
It is obvious from the present findings that the initiation of shoot apical meristem is not only dependent upon genotype, but also on the type and concentration of plant growth regulators.
Former studies suggested that auxin has positive effects on root induction, where IBA and NAA are the most frequently used auxins for root induction in vitro, but the concentration depends upon the type of plant and its genotype [19]. NAA is the most efficient plant growth regulator for root induction in sugarcane, as appeared in CP-77400 and NSG-59 and confirmed by other scientists [32,33,34,35]. The synergetic effect of 1 mg L−1 NAA and 1 mg L−1 1 IBA was also found to be effective for root induction in sugarcane variety B41-227 [36].
5. Conclusions
From the above results, it was concluded that callus, root and shoot induction in any genotype of sugarcane was highly sensitive to the concentration and type of plant growth regulator. Moreover, different morphological types of calliunder different treatments determine the fate of the next developmental stages. This study also highlights the importance of Kinetin in callus induction, proliferation and shoot induction. Eventually, a complete optimized protocol will be able to be used in commercial laboratories to propagate sugarcane plant or conduct any experiments on the above genotypes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture12111812/s1, Table S1: Pair-wise comparison of treatment and variety means of studied traits as a result of least significant difference test (LSDα0.05).
Author Contributions
Conceptualization, funding acquisition, methodology, writing—original draft, Y.S.; Data curation, Formal analysis, Funding acquisition, M.Z.E.; Supervision, A.A.; Project administration, S.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gomathi, R.; Krishnapriya, V.; Arunkumar, R.; Govindaraj, P.; Ram, B. Physiological Traits Imparting Drought Stress Tolerance to Promising Sugarcane (Saccharum Spp.) Clones. Plant Physiol. Rep. 2020, 25, 509–515. [Google Scholar] [CrossRef]
- Hensel, F.; Covello, A.; Gargano, A. International Seminar on Agriculture, Biodiversity, Food Security and Health; IOP Conf. Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 883, p. 012075. [Google Scholar]
- Dinesh, P.; Thirunavukkarasu, D.P.; Saraniya, A.R.; Ramanathan, T. In vitro Studies of Sugarcane Variety Co-91017 Through Micropropagation of Shoot Tip Culture. Adv. Plants Agric. Res. 2015, 2, 234–238. [Google Scholar]
- Salokhe, S. Development of an Efficient Protocol for Tissue Culture of Sugarcane. Plant Cell Biotechnol. Mol. Bio. 2021, 4, 9–21. [Google Scholar]
- Lal, M.; Tiwari, A.K.; Gupta, G.N. Commercial Scale Micropropagation of Sugarcane: Constraints and Remedies. Sugar Tech 2015, 17, 339–347. [Google Scholar] [CrossRef]
- Hailu, M.; Chimdessa, M.; Muthswamy, M. In Vitro Propagation of Selected Sugarcane (Saccharum officinarum L.) Varieties (C 86-165 and C 86-12) Through Shoot Apical Meristem. Int. J. Hortic. Agric. 2018, 3, 1–7. [Google Scholar]
- Purnamaningsih, R.; Sukmadjaja, D.; Suhesti, S.; Rahayu, S. In vitro Propagation of Six Selected Sugarcane Mutant Clones Through Leaf Explants. IOP Conf. Ser. Earth Environ. Sci. 2021, 883, 012075. [Google Scholar] [CrossRef]
- Subedi, M.; Ojha, B.R.; Ghimire, S.K.; Joshi, B.K.; Niroula, R.K.; Sah, B.P.; Poudel, A.P. Effect of sugarcane genotypes and 2, 4-dichlorophenoxy acetic acid on callus induction. J. Inst. Agric. Anim. Sci. 2015, 33, 237–242. [Google Scholar] [CrossRef]
- Kumar, D.; Sengar, R.S.; Malik, N.; Yadav, M.K.; Gupta, S.; Chand, P.; Kumar, P. In Vitro Evaluation to Intensify the Differential Morphogenetic Response Through Plant Growth Regulators and Antibiotic Supplementation in Sugarcane. Plant Physiol. Rep. 2020, 25, 335–346. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Culture. Physiol. Plant. 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Aftab, F.; Zafar, Y.; Malik, K.A.; Iqbal, J. Plant regeneration from embryogenic cell suspensions and protoplasts in sugarcane (Saccharum spp. Hybrid cv. Col-54). PCTOC 1996, 44, 71–78. [Google Scholar] [CrossRef]
- Sengar, K.; Sengar, R.S.; Garg, S.K. The Effect of in vitro Environmental Conditions on Some Sugarcane Genotypes for Micropropagation. Afr. J. Biotechnol. 2011, 10, 17122–17126. [Google Scholar]
- Mostafa, H.H.; Wang, H.; Song, J.; Li, X. Effects of Genotypes and Explants on Garlic Callus Production and Endogenous Hormones. Sci. Rep. 2020, 10, 4867. [Google Scholar] [CrossRef] [PubMed]
- Baday, S.J. Sugarcane of Rapid Multiplication by Callogenesis. JPSC 2020, 1660, 012005. [Google Scholar] [CrossRef]
- Srivong, T.; Zhu, Y.J.; Pongdontri, P.; Pliansinchai, U.; Sakuanrungsirikul, S.; Borthakur, D.; Kosittrakun, M. Optimization of Callus Induction and Plant Regeneration in Sugarcane (Saccharum spp.) for a Study of Sucrose Accumulation in Relation to Soluble Acid Invertase Expression. Chiang Mai J. Sci. 2015, 42, 797–805. [Google Scholar]
- Mayerni, R.; Satria, B.; Wardhani, D.K.; Chan, S.R.O.S. Effect of Auxin (2,4-D) and Cytokinin (BAP) in Callus Induction of Local Patchouli Plants (Pogostemon cablin Benth.). IOP Conf. Ser. Earth Environ. Sci. 2020, 583, 012003. [Google Scholar] [CrossRef]
- Kaur, A.; Malhotra, P.K.; Manchanda, P.; Gosal, S.S. Micropropagation and Somatic Embryogenesis in Sugarcane. Biotechnol. Crop Improv. 2018, 1, 57–91. [Google Scholar]
- Dalila, Z.D.; Jaafar, H.; Manaf, A.A. Effects Of 2, 4-D and Kinetin on Callus Induction of Barringtonia racemosa leaf and Endosperm Explants in Different Types of Basal Media. Asian J. Plant Sci. 2013, 12, 21–27. [Google Scholar] [CrossRef][Green Version]
- Li, S.; Huang, P.; Ding, G.; Zhou, L.; Tang, P.; Sun, M.; Zheng, Y.; Lin, S. Optimization of Hormone Combinations for Root Growth and Bud Germination in Chinese Fir (Cunninghamia lanceolata) Clone Leaf Cuttings. Sci. Rep. 2017, 7, 5046. [Google Scholar] [CrossRef] [PubMed]
- Mastuti, R.; Munawarti, A.; Firdiana, E.R. The Combination Effect of Auxin and Cytokinin on in vitro Callus Formation of Physalis angulata L.—A Medicinal Plant. AIP Conf. Proc. 2017, 1908, 040007. [Google Scholar]
- Iqbal, M.; Aamir, A.; Naima, H.N.; Umair, A.K.; Muhammad, N.A.F.; Muhammad, I.; Mubashir, H. Effect of Explants and Growth Regulators on The Expression of Callogenesis Somatic Embryogenesis and Plantlets Formation in Sugarcane (Saccharum officinarum L.). Int. J. Biosci. 2016, 9, 147–156. [Google Scholar]
- Tripathy, S.K.; Ithape, D.M. High-throughput in vitro culture system targeting genetic transformation in sugarcane. J. Crop Sci. Biotechnol. 2020, 23, 325–335. [Google Scholar] [CrossRef]
- Perez-Jimenez, M.; Cantero-Navarro, E.; Perez-Alfocea, F.; Le-Disquet, I.; Guivarc’h, A.; Cos-Terrer, J. Relationship Between Endogenous Hormonal Content and Somatic Organogenesis in Callus of Peach (Prunus persica L. Batsch) Cultivars and Prunus persica× Prunus dulcis Rootstocks. J. Plant Phys. 2014, 171, 619–624. [Google Scholar] [CrossRef]
- Aslam, M.M.; Karanja, J.K.; Zhang, Q.; Lin, H.; Xia, T.; Akhtar, K.; Xu, W. In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes. Plants 2020, 9, 318. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Muhammad, A.; Hussain, I.; Shah, S.H.; Kumar, T.; Inam, S.; Ali, G.M. Rapid in vitro Multiplication of Sugarcane Elite Genotypes and Detection of Sugarcane Mosaic Virus Through Two Steps RT-PCR. Int. J. Agric. Biol. 2012, 14, 870–878. [Google Scholar]
- Muller, D.; Leyser, O. Auxin, Cytokinin and the Control of Shoot Branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Bidabadi, S.S.; Jain, S.M. Cellular, Molecular and Physiological Aspects of in vitro Plant Regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Abu-Romman, S.M.; Al-Hadid, K.A.; Arabiyyat, A.R. Kinetin is the most effective cytokinin on shoot multiplication from cucumber. J. Agric. Sci. 2015, 7, 159. [Google Scholar] [CrossRef]
- Devi, T.R.; Dasgupta, M.; Sahoo, M.R.; Kole, P.C.; Prakash, N. High Efficient de novo Root-to-shoot Organogenesis in Citrus jambhiri Lush.: Gene expression, Genetic Stability and Virus Indexing. PLoS ONE 2021, 16, e0246971. [Google Scholar] [CrossRef]
- Patel, A.A.; Patel, S.R.; Patel, C.L.; Prajapati, B.S. Effect of Media Composition on in vitro Multiplication of Sugarcane Genotypes. Indian J. Gen. Plant Breed. 2001, 61, 82–83. [Google Scholar]
- Kaur, R.; Kapoor, M. In Vitro Direct Plant Regeneration Using Shoot Tip Explants in Sugarcane (Saccharum officinarum L.) for rapid mass cloning. Agric. Sci. Digest. 2017, 37, 94–99. [Google Scholar] [CrossRef][Green Version]
- Biswas, P.; Harun-Or Rashid, M.D.; Chowdhury, A.K.; Hossain, M.D.A. Direct Plant Regeneration Through Micropropagation Using Selected Explants of Sugarcane. Int. J. Plant Geosci. 2020, 8, 244–248. [Google Scholar] [CrossRef]
- Gupta, C.; Nema, S.; Sapre, S.; Tantwai, K. Studies on Induction of Somaclonal Variation in Sugarcane (Saccharum officinarum) and Validation of Mutant Using Molecular Markers. Int. J. Agric. Environ. Biotechnol. 2020, 13, 105–110. [Google Scholar] [CrossRef]
- Rahman, M.M.; Ivy, N.A.; Mian, M.A.; Rasul, M.G.; Hossain, M.M.; Rahman, M.A. Effect of Auxin (NAA, IBA and IAA) in Root Regeneration Through in vitro Culture of Sugarcane. Int. J. Plant Biol. Res. 2018, 6, 1109. [Google Scholar]
- Silva, D.J.A.; Solis-Gracia, N.; Jifon, J.; Souza, S.C.; Mandadi, K.K. Use of Bioreactors for large-Scale Multiplication of Sugarcane (Saccharum spp.), Energy Cane (Saccharum spp.) and Related Species. In Vitro Cell. Dev. Biol. Plant 2020, 56, 366–376. [Google Scholar] [CrossRef]
- Tolera, B. Effects of Naphthalene Acetic Acid (NAA) and Indole-3-Butyric Acid (IBA) on in vitro Rooting of Sugarcane (Saccharum officinarum L.) Micro-shoots. J. Biotechnol. Biomater. 2016, 6, 215. [Google Scholar] [CrossRef]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).