Abstract
Passiflora edulis f. flavicarpa (yellow passion fruit) is a high-value tropical crop explored for both fruit and nutraceutical markets. As the fruit production in the US rises, the crop must be investigated for the effects of salinity under semi-arid climates. We assessed the effects of irrigation-water salinity, leaf age, and drying method on leaf antioxidant capacity (LAC) and plant genetic responses. Plants were grown in outdoor lysimeter tanks for three years, with waters of electrical conductivities of 3.0, 6.0, and 12.0 dS m−1. Both Na and Cl significantly increased with salinity; leaf biomass at 3.0 and 6.0 dS m−1 were similar but reduced significantly at 12.0 dS m−1. Salinity had no effect on LAC, but new leaves had the highest LAC compared to older leaves. Low-temperature oven-dried (LTO) and freeze-dried (FD) leaves had the same LAC. The analyses of twelve transporter genes, six involved in Na+ transport and six in Cl− transport, showed higher expressions in roots than in leaves, indicating a critical role of roots in ion transport and the control of leaf salt concentration. Passion fruit’s moderate tolerance to salinity and its high leaf antioxidant capacity make it a potential new fruit crop for California, as well as a rich source of flavonoids for the nutraceutical market. Low-temperature oven drying is a potential alternative to lyophilization in preparation for Oxygen Radical Absorbance Capacity (ORAC) analysis of passion fruit leaves.
1. Introduction
Passion fruit (Passiflora edulis spp., Passifloraceae) is one of the most consumed small fruits worldwide, with approximately 150 native species found in Brazil, although only a few species produce edible fruits [1]. These species have fruit peels of different colors, ranging from pale yellow to purple. Aside from the economic value given by the different colors, sizes, flavors, and aromas of the different fruits, the multicolored flowers give the species a high ornamental value. The yellow passion fruit (Passiflora edulis Sims. forma flavicarpa Deg.) is reported to be originally from Brazil, where its common names are based on the color of the fruit rind, used to differentiate the cultivars [2]. A recent hybrid named ‘BRS Gigante Amarelo 1′ is the ‘Brasil Yellow Giant 1′ developed by Embrapa Cerrados and collaborators and released in 2008. This cultivar produces fruits ranging in fresh weight from 120 to 350 g, and fruit yield ranging from 16–42 ton ha−1 when spaced at 3 m × 5 m, fertilized and irrigated according to the crop recommendation, and manually pollinated daily during flowering [3]. The global data for the trade of minor tropical fruits remain difficult to obtain, but the 2015–2017 worldwide estimated production of passion fruit was around 1.47 million metric tons (MT), the fifth highest after guava (6.7 MT), longan and lychee (both 3.4 MT), and durian (2.3 MT) [4]. Thus, there are no data for worldwide passion fruit production by country compiled by FAO. Brazil is the largest producer and consumer of passion fruit; however, the yield of Passiflora edulis f. flavicarpa decreased from 838,000 MT in 2013 to 593,400 MT in 2019 (https://www.statista.com/statistics/1078358/production-passion-fruit-brazil/), accessed on 27 October 2022. In contrast, the Colombian passion fruit cultivated area grew from 15,000 ha in 2016 to 21,000 ha in 2017 (https://farmfolio.net/articles/colombias-passion-fruit-production-continues-rise/), accessed 10 September 2022. Colombia reported passion fruit production of 150,000 MT in 2016, with most of its product staying home and only 6,000 MT exported to foreign markets. In 2018, Colombia’s production was mostly (72%) P. edulis f. flavicarpa (169,000 MT), followed by P. ligularis (sweet granadilla) with 47,460 MT, and P. edulis f. edulis (purple passion fruit) with 24,800 MT (https://storymaps.arcgis.com/stories/bf57e656b0334bac83f73f5f9e5fca50), accessed on 10 September 2022. Most of the produced P. edulis f. edulis was used for export purposes.
The passion fruit is primarily famous for its sweet (purple fruit) or sour (yellow fruit) pulp; the nutraceutical market has been long established for the use of the leaf and flower extracts due to their anxiolytic and sedative effects [5] as well as for being generally regarded as safe [6]. Although considered safe for most users, the extract of Passiflora incarnata, at therapeutic doses of 500–1000 mg three times a day, was reported to cause adverse effects in one 34-year-old female patient postpartum [7]. Because there was no toxicity associated with the analysis of the extract, the authors hypothesized that the patient had a defective cytochrome P450 enzyme, and was unable to metabolize Harman alkaloids present in P. incarnata extracts.
Passiflora edulis is the species with the highest commercial value due to its easy adaptation to different soils and climates [8], great use for industrial products (food, nutraceutical, and cosmetics), and for having mild tolerance to salinity [9]. Tolerance to salinity is vital for the commercial feasibility of the crop in arid and semi-arid regions. Regarding the effects of salinity on the crop, the yellow passion fruit was reported to have a minimal reduction in fruit quality when irrigated with waters of electrical conductivity (ECw) of 4 dS m−1 [10,11].
There is recent information on the effect of salinity on passion fruit growth and anatomical, physiological, and nutritional responses to salinity [12]. These authors evaluated the species P. edulis, P. mucronata, and their hybrid for 20 days after saline irrigation. They reported that P. mucronata was the most tolerant to salt due to its lowest leaf Na accumulation and highest stomatal conductance, photosynthesis, and leaf number when challenged with 150 mM NaCl. Another recent publication evaluated the antioxidant capacity and flavonoid characterization of passion fruit pulp, peel, and seeds [13]. However, there are no data on the effects of salinity on leaf antioxidant capacity and leaf biomass accumulation in adult passion fruit plants.
Our search of the literature revealed no data on the effects of different drying procedures on the antioxidant capacity of passion fruit leaves. In the United States, the food industry evaluates the antioxidant capacity of fruits, juices, and vegetables through the Oxygen Radical Absorbance Capacity (ORAC) method. For the ORAC analysis, the samples must be freeze-dried and lyophilized. Both the lyophilization process and ORAC method are expensive and time-consuming, with the ORAC analysis being unaffordable for small labs in the US and most labs in developing countries. In the past, it has been demonstrated that drying Artemisia annua leaves in a forced-air oven at 50 °C for 48 h preserved the highest leaf antioxidant capacity, without any significant reduction [14]. As the leaves of passion fruit vines are used to prepare medicinal remedies, it is important to evaluate practical, affordable, and rapid methods to determine the antioxidant capacity of the different species of interest. In this work, we compared low-temperature oven drying with freeze-drying and lyophilization and tested antioxidant capacity by both the ORAC and total phenolics (TP) methods.
Despite passion fruit’s ecological and economic importance, molecular markers have only recently been used in genetic studies [15]. Considering that the genus is a rich source of ornamental vines (large array of flower colors), folk medicine (extract of leaves and flowers), cosmetics (hydrating creams), and food for humans (such as fresh juice, ice creams, frozen concentrated juices, pulp, and seeds), it is surprising that there are no studies on the molecular genetic response of passion fruit to salinity. Despite taxonomical disagreements on the number of genera (between 18 and 23) and species (between 520 and 700), approximately 96% of the species are distributed in the Americas, with major centers of diversity in Brazil and Colombia (both having approximately 30% of the Passiflora species) [15]. However, a few species have been reported from India, China, Southeast Asia, Australia, the Pacific Islands, and neighboring regions [15]. Recently, an effort has been funded by FAPESP (Research Support Foundation of São Paulo, São Paulo, Brazil) for the genetic characterization of 150 Passiflora species from Brazil as a valuable gene bank to manage conservation and biodiversity [16]. Although the main producers of passion fruit in Brazil are in the semi-arid region, afflicted by natural and anthropogenic irrigation water salinity, the study does not mention tolerance to salinity.
The objectives of the study were to evaluate the effects of irrigation-water salinity on leaf Na and Cl accumulation, mineral status, leaf biomass, antioxidant capacity, and genetic responses of yellow passion fruit plants. The study also evaluated the effect of leaf age and drying methods on leaf antioxidant capacity.
2. Materials and Methods
The United State Salinity Laboratory (USSL) is located in Riverside, southern California, at the latitude 33.9°58′24′′ N, longitude 117°19′12′′ E, and altitude of 311 m above sea level, with average high temperatures from June to September ranging from 30.6 °C to 34.5 °C. However, extreme summer weather in July and August elevates these temperatures to over 40 °C and brings relative humidity to 25% or lower. Riverside has an average annual precipitation of 262.6 mm, concentrated mostly from November to March. The day length ranged from 9 h:53 min:36 s on the shortest day of the year (21 December 2018, the solstice) to 14 h:25 min on the longest day of the year (21 June 2020), according to https://www.timeanddate.com/sun/usa/riverside?month=9, accessed on 10 September 2022.
2.1. Plant Material
Passiflora edulis f. flavicarpa seeds were obtained from the state of Paraiba, a state located on the semi-arid coast of northeastern Brazil, transferred to the US through APHIS-USDA, and planted in six-inch pots in April 2015 in a greenhouse at the USSL. Seedlings were kept under natural illumination and temperature until they were three months old, with an approximate height of 45 cm. Seedlings were irrigated with Riverside municipal water (EC = 0.65 dS m−1) and received basic fertilization of 20-0-20 (NPK) in the pot and a leaf spray of 2% urea every two weeks until transplant. Seedlings were transplanted to outdoor sand tanks with two plants per tank and a spacing of 2.0 m between plants.
2.2. Irrigation Water Salinity and Sand Tank Cultivation
Plants were irrigated with Riverside municipal water (average EC = 0.65 dS m−1 and pH = 7.5) with ionic composition reported in Table 1, enriched with NO3− (9 mmolc L−1), K+ (6 mmolc L−1), and P (1 mmolc L−1) for the first month while plants were being established in the outdoor lysimeter tanks. After the plants were established, 30 days after transplanting (4 August 2015), they were irrigated with waters of salinity of 1.8, 3.8, and 6.8 dS m−1 for the first year. As plants did not show any significant difference in leaf biomass under these salinities, the irrigation water salinities were increased to 3.0, 6.0, and 12.0 dS m−1 for the second and third years of crop cultivation. Over time, and as nutrients were added, the final EC of the control water reached 3.0 dS m−1. The treatments of 3.8 and 6.8 dS m−1 were increased to 6.0 and 12.0 dS m−1 and maintained for the last two years of the experiment. The ionic composition of the control and saline treatment waters is shown in Table 1.

Table 1.
Averaged target electrical composition (ECw), pH, and mineral ion composition of passion fruit saline irrigation waters and Riverside municipal water (RMW). Ion composition in the table reflects the concentration of ions that were added to existing ion composition in RMW as NaCl, CaCl2, MgCl2, MgSO4, Na2PO4, and KNO3 in order to provide the ions specified in the table and to achieve each target ECw. No CO3H− was added to treatment waters in addition to what RMW already had. Micronutrients were added through a solution based on Hoagland and Arnon [17].
To this basic nutrient solution, we added salts of Na, Cl, SO4, Ca, Mg, and S in order to provide the target salinities (Table 1). Saline treatment solutions were pumped from 24 water reservoirs (Vol = 3605 L each) housed underneath the lysimeter facility to each of the 24 large sand tanks above, completely saturating and leaching the sand culture medium. Sand tanks were filled with coarse sand with dimensions of 1.5 m (W) × 3.0 m (L) × 2.0 m (D) (Figure 1). Plants were irrigated twice a week with treatment waters stored in underground reservoirs of 3605 L, each designated to irrigate one large sand tank above. After irrigating each tank, each saline nutrient solution returned to the reservoir through a subsurface drainage system at the bottom of the tanks (Figure 1), thus maintaining a uniform and constant salinity in the plant root zone.
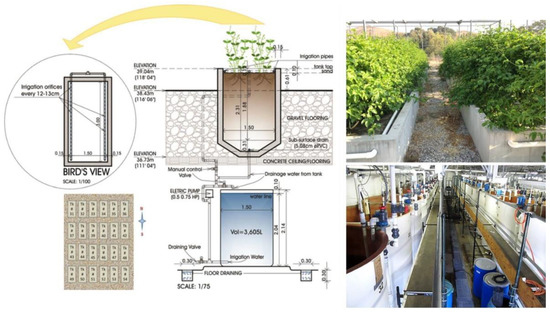
Figure 1.
Schematic cross-section representation of large outdoor sand tanks (top left) and water reservoirs (bottom left) used to grow yellow passion fruit vines, with a bird’s view of a single tank (top left) and the 25 sand tanks on a north/south orientation (bottom left). On the right, a representation of the front view of passion fruit in sand tanks (top right) and of the 24 water reservoirs (bottom right) housed underneath the sand tanks.
2.3. Plant Mineral Nutrition
The base nutrient solution was prepared following the nutrition recommendation for passion fruit: N as NO3− (9 mmol L−1), P as H2PO4− (1.0 mmol L−1), K (6.0 mmol L−1. For the micronutrients, we used the Hoagland and Arnon No. 1 solution [17] with the following composition: 46 µmol L−1 of B; 0.3 µmol L−1 of Cu; 12.6 µmol L−1 of Mn; 0.1 µmol L−1 of Mo; 90 µmol L−1 of Fe, and 1.3 µmol L−1 of Zn.
2.4. Lysimeter Tanks
Each lysimeter tank had two plants spaced 2.5 m apart. The coarse sand in each tank was mixed with 10% peat moss (v/v), resulting in an average bulk density of 1380 kg m−3 and an average volumetric water + air content of 0.30 m3 m−3, determined by packing dry sand, weighing, saturating with water, and reweighing. Plants were assigned to tanks in a complete randomized design with eight replicates per treatment (eight tanks per treatment) with two plants per tank, totaling sixteen plants per treatment.
2.5. Sample Collection
Leaves were collected in groups according to their position in the new tertiary stem generated in 2019, then compared to one large leaf from the secondary stem that originated in 2018 (Figure 2). Leaf samples for Na, Cl, and macronutrient analyses were fully expanded and taken from positions 7–9 in the new branch, counted from the newest leaf in the apex in April 2019, with approximately 15 leaves collected per tank. For antioxidant analyses, leaves were collected from positions 1–3 (new leaves, NL), 4–7 (young leaves, YL), 8–12 (physiologically mature, ML), and one previous-year (PY) leaf (Figure 2). After collection, a portion of the leaf samples was placed in cloth bags and immediately dipped into liquid nitrogen, freeze-dried (lyophilized), and ground in order to determine the antioxidant capacity through both ORAC and total phenolic tests. The other portion of the sampled leaves was oven-dried at 50 °C for 48 h for ORAC and total phenolic analyses to evaluate if this oven temperature would preserve most of the antioxidant capacity when compared to freeze-drying, as previously determined for Artemisia annua [14]. Leaves from positions 1–7 were also collected from different branches and from the east or west side of the plant in order to assess whether the direction of sun exposure would affect antioxidant capacity. All leaves were kept in the dark and stored in a −80 °C freezer before being directed to either freeze-drying or low-temperature oven drying. They were then ground in a Willey Mill to a 1 mm-particle size and maintained at −80 °C before ORAC and total phenolic analyses. For fresh biomass accumulation, plants were pruned up to the secondary stem, placed in a large plastic tarp, and immediately weighed on a large scale. The vine dry weight was later estimated by taking six samples from different tanks and drying them in an oven at 70 °C until they reached a stable weight.
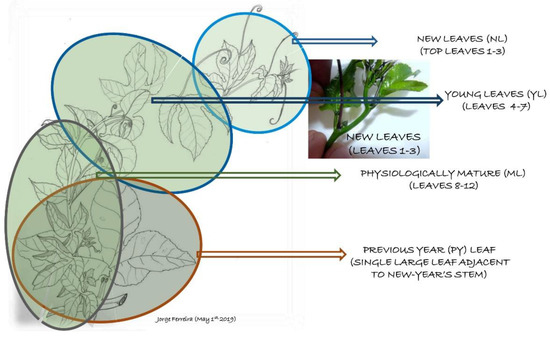
Figure 2.
Schematic drawing of a new passion fruit stem arising from the previous year’s stem. Elliptical lines show the group of leaves collected, starting from the new leaves at the tip of the stem to the oldest at the base. The large leaf at the base of that new stem is from the previous year’s (PY) growth and was also used to compare antioxidant capacity. Schematic illustration by Jorge F.S. Ferreira.
2.6. Antioxidant Capacity Tests
Leaves were analyzed by the ORAC test as well as by the TP (Folin–Ciocalteu) test following methodology previously validated [19,20] and currently used in our USDA laboratory in order to determine the shoot antioxidant capacity of several crops, including spinach, alfalfa, and artemisia [14,21,22].
2.7. Expression Analyses
Expression changes under salinity were studied using quantitative Reverse Transcription—Polymerase Chain Reaction (qRT-PCR). Some genes involved in Na+ transport and Cl− transport were selected for our expression analyses. The Arabidopsis sequence for each selected gene was used to identify the corresponding cassava (Manihot esculenta) gene, which was further used against the passion fruit whole genome shotgun sequence to identify the passion fruit ortholog. Primers were designed for the sequence that showed the highest homology (Table S1).
RNA samples were collected from leaves and roots, and RNA was isolated using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA). DNA contamination was removed using DNase I (Thermo Scientific, Waltham, MA, USA), and the samples were diluted to 5 ng/μL. The qRT-PCR analyses were performed in the BioRad CFX96 machine, utilizing the iTaqTM Universal SYBR® Green One-Step Kit (Bio-Rad Laboratories, Hercules, CA, USA). The PCR reaction was carried out in 10 μL volume containing 10 ng total RNA, 0.75 μM each primer, 5 μL 2X one-step SYBR® Green Reaction mix, and 0.125 μL Bio-Rad iScriptTM Reverse Transcriptase (Bio-Rad Laboratories, Hercules, CA, USA). Amplification was carried out using the following program: 50 °C for 10 min, 95 °C for 1 min, followed by 40 cycles of 95 °C for 10 s, 57 °C for 30 s, and 68 °C for 30 s. Three passion fruit housekeeping genes encoding histone (XP_002525279.1), 60S ribosomal protein (XP_002531173.1), and transcription initiation factor (XP_002299546.1) were used as reference genes for the qRT-PCR analyses [23].
3. Results
The results reflect the salt, mineral, antioxidant capacity, and gene expression in leaves of adult passion fruit plants after three years of exposure to saline waters and hot summer temperatures in Riverside, CA, USA.
3.1. Effects of Leaf Na and Cl Accumulation on Shoot Biomass
Leaf mineral analysis showed that an increase in irrigation water salinity resulted in significant increases in Na and Cl leaf concentrations. Passion fruit plants accumulated significantly more sodium and chloride with increasing salinity (Figure 3a). While leaves of plants irrigated with control water of ECw=3.0 dS m−1 had, on average, 9.8 g kg−1 of Na and 10.2 g kg−1 of Cl, leaves of plants irrigated with ECw = 6.0 dS m−1 had 18.4 g kg−1 of Na and 22.8 g kg−1 of Cl, and plants irrigated with ECw = 12.0 dS m−1 had, on average, 29.4 g kg−1 of Na and 43.3 g kg−1 of Cl (Figure 3a, Table 2). Plants irrigated with waters of the highest salinity (12 dS m−1) presented leaf edge scorching (Figure 4). Dry shoot (leaves and stems) biomass was, on average, 31.4% of fresh weight, regardless of salinity.
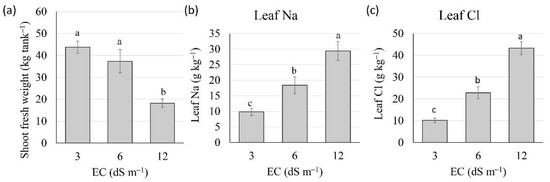
Figure 3.
Biomass and ion concentrations of passion fruit plants irrigated with saline waters of three electrical conductivities (ECw). (a) Shoot fresh weight. (b) Sodium (Na) concentrations. (c) Chloride (Cl) concentrations. Data are means ± 1SE, (n = 8). Different letters indicate significant differences (p ≤ 0.05) among the three salinity treatments.
Table 2.
Leaf macro- and micronutrient concentrations, including sodium and chloride, of passion fruit plants irrigated with waters of three electrical conductivities (ECw). Data are the means of eight outdoor lysimeter tanks (n = 8), except for the highest salinity (n = 6). Different letters indicate significant differences (p ≤ 0.05) among the three salinities. DM: dry matter. The water pH was 7.8.

Figure 4.
The general aspect of leaves of passion fruit plants irrigated with waters of different salinities. (a) 3.0 dS m−1. (b) 6.0 dS m−1. (c) 12.0 dS m−1. Plant biomass was significantly reduced by 58.45% at the highest salinity, with leaves showing burned edges typical of excess Cl accumulation.
Although there was no significant reduction in average leaf biomass between plants irrigated with 3.0 and 6.0 dS m−1 (43.8 vs. 37.3 kg tank−1), there was a significant reduction in leaf biomass (18.2 kg tank−1) for plants irrigated with ECw = 12.0 dS m−1 (Figure 3b). The leaves of plants irrigated with waters of ECw of 3.0 and 6.0 dS m−1 (Figure 4a,b) had no salt-toxicity symptoms, but the leaves of plants irrigated with ECw = 12.0 dS m−1 showed lateral burning of mature leaves (Figure 4c). Both plants from each of the two tanks under ECw = 12 dS m−1 died during the third year.
Although leaf concentrations of Na and Cl increased with each increase in irrigation-water salinity (Figure 3), the concentrations of macro- and micronutrients remained fairly constant across salinities (Table 2). In comparison to the expected macro- and micronutrient leaf composition of field-grown Passiflora alata Curtis [24], our plants were low in Mn. Although Mn was provided in a mixture of essential minerals, the water pH (7.8) was above the ideal pH (>6.2) for most plants, after which manganese has a decreased availability for plant uptake.
3.2. Effects of Na and Cl Accumulation, Drying Method, and Leaf Age on Antioxidant Capacity
Salinity did not affect the antioxidant capacity of passion fruit leaves. The leaves used for this analysis were the same leaves collected for mineral analysis, and were from the middle portion of the stem. These leaves, sampled from the middle of the tertiary branch (leaves 7–9), had ORAC concentrations ranging from 620–750 µmoles TE g−1 DM, and TP ranging from 15–19 mg GAE/g DM (Figure 5). There was also no difference in antioxidant capacity for either ORAC and TP between leaves from the east or west side of the vine (Figure 5).

Figure 5.
Comparison of drying methods and leaf developmental stages (1–12) for passion fruit leaves measured through the hydrophilic oxygen radical absorbance capacity (ORAC), and total phenolics (TP) concentration of freeze-dried and oven-dried leaves at different developmental stages. (a) Leaf ORAC at different leaf developmental stages. (b) Leaf TP at different leaf developmental stages. Vertical bars represent means plus/minus standard error (±1SE) of four replicates (n = 4) (4 tanks each with two vines). Different letters indicate significant differences (p ≤ 0.05) among different leaf growth stages. Lack of letters between drying methods indicate lack of significant differences. TE: Trolox equivalent, GAE: gallic acid equivalent. There was no significant difference (p > 0.05) between the two drying methods at any leaf developmental stage. PY, previous-year leaves.
When comparing freeze-drying at −50 °C for 3–4 days with oven drying at 50 °C for 48 h, leaves at different positions of the newest branch of passion fruit vines showed no difference in antioxidant capacity (ORAC) or TP (Figure 5). However, depending on their physiological age, leaf groups had significantly different (p < 0.05) ORAC values, with new leaves (NL, leaves 1–3) having average ORAC values of 1200 µmoles TE g−1 DW, while young leaves (YL, leaves 4–7) and physiologically mature leaves (ML, leaves 8–12) of the same branch had average ORAC value of 850 µmoles TE g−1, and leaves from the previous year (PY) had the lowest ORAC values of approximately 580 µmoles TE g−1 (Figure 6a). Leaf TP followed the same trend as ORAC, with NL having the highest values in TP, approximately 24 mg GAE g−1 DW, and ML and PY leaves having values of approximately 15 and 14 mg GAE g−1 DW, respectively (Figure 6b).
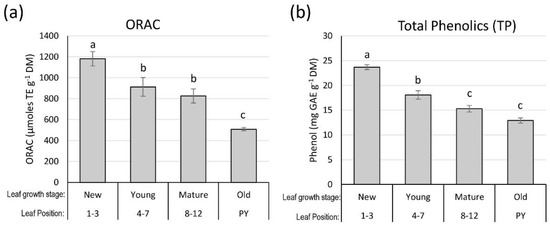
Figure 6.
Effect of passion fruit leaf developmental stage on the hydrophilic oxygen radical absorbance capacity (ORAC) and total phenolics (TP) concentration. (a) Leaf ORAC at different leaf developmental stages. (b) Leaf TP at different leaf developmental stages. Vertical bars represent means plus/minus one standard error (±1SE) of eight replicated samples (n = 8) (two vines per tank, freeze-dried and oven-dried sample data were combined). TE: Trolox equivalent, GAE: gallic acid equivalent. Different letters indicate significant differences (p ≤ 0.05) among different leaf growth stages. PY, previous-year leaves.
3.3. Effect of Salinity on Gene Expression
Twelve genes known to be involved in salt stress were used to study expression changes between control and salinity treatments. Of these, six genes (AKT1, NHX1, NHX2, SOS1, SOS2, and SOS3) are known to be involved in Na transport, and six genes (ALMT12, ALMT9, CCC, CLCc, CLCg, and SLAH3) are known for their roles in Cl transport. Expression analyses revealed higher expression of all 12 genes in roots than in leaves, irrespective of salinity treatment (Figure 7).
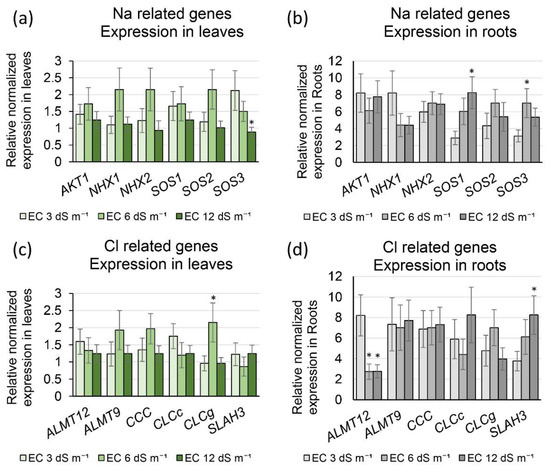
Figure 7.
Expression of genes involved in Na+ and Cl− transport under EC = 3 dS m−1, EC = 6 dS m−1, and EC = 12 dS m−1 in leaves and roots of passion fruit. (a) Expression of genes involved in Na+ transport in leaves. (b) Expression of genes involved in Na+ transport in roots. (c) Expression of genes involved in Cl− transport in leaves. (d) Expression of genes involved in Cl− transport in roots. * indicate a significant difference (p ≤ 0.05) between EC = 3 dS m−1 and EC = 6 dS m−1 or 3 dS m−1 and EC = 12 dS m−1. Bars represent standard errors.
Of the genes involved in Na transport, SOS1 and SOS3 were significantly upregulated in roots, at 12 dS m−1 compared to 3 dS m−1, whereas SOS3 was significantly upregulated at 6 dS m−1 compared to 3 dS m−1 (Figure 7b). On the other hand, SOS3 was downregulated in leaves at 12 dS m−1 compared to the control (Figure 7a). None of the other genes were differentially expressed under control and salinity conditions.
4. Discussion
4.1. Effects of Na and Cl Accumulation on Shoot Biomass
According to the salinity tolerance of several species under hydroponics, species that tolerated an ECw = 5.5 dS m−1 were considered tolerant [25]. In addition, young passion fruit plants that tolerated irrigation-water salinity of ECw = 4.43 dS m−1 were considered moderately tolerant to salinity [26]. As our yellow passion fruit plants had significant increases in the accumulation of both Na and Cl in leaves, but had no significant decrease in shoot biomass or visual salt toxicity symptoms up to ECw = 6.0 dS m−1, we believe that our plants can be classified as moderately tolerant to salinity. However, the accumulation of either Na or Cl (or both) was toxic for the vines and reduced their leaf biomass production at the ECw = 12.0 dS m−1. After two years of exposure to the highest irrigation water salinity of 12.0 dS m−1, although the plants had enough macronutrients to sustain their growth, four of the sixteen plants under this treatment died during the last months of the third year. Additionally, all surviving plants had their shoot biomass severely reduced and leaf marginal burning typical of chloride toxicity (Figure 4c), also consistent with chloride toxicity in strawberry leaves [27]. However, it is impossible to exclude the possibility that leaf damage could also be caused by sodium unless a separate study were conducted, with sodium and chloride salts used separately. As the yellow passion fruit plants evolved in the Brazilian Amazon, where soils are very low in sodium and chloride and excessive salts are leached by the abundant rains, we believe that plants have no effective mechanism to avoid either Na+ or Cl− absorption, as it was reported and discussed recently for habanero peppers irrigated with saline waters [28]. In fruit crops, such as avocado rootstocks, salt toxicity has been reported at much lower salinity levels. When Hass avocado plants field-grown in Riverside were grafted onto 13 rootstocks and irrigated with water salinity of 1.5 dS m−1 for 23 months, a significant variation in mortality rate was observed for different rootstocks with Hass plants that were grafted onto more salt-tolerant rootstocks, accumulating less leaf chloride [29]. Results prompted the authors to conclude that plant mortality was highly correlated to chloride (but not sodium) accumulation in avocado leaves [29]. As our passion fruit vines presented similar chloride toxicity symptoms to those seen in avocado leaves (necrosis of leaf tips extending to leaf edges, Figure 4c), maybe NaCl-tolerant passion fruit rootstocks could increase the yellow passion fruit salt tolerance, as it was found to do for avocados in Southern California.
4.2. Effects of Na and Cl Accumulation, Drying Method, and Leaf Age on Antioxidant Capacity
The antioxidant capacity of leaves, measured by both ORAC and TP methods, did not change in response to salinity, and ranged from 600–800 µmoles TE g−1 (ORAC) and 15–18 mg GAE g−1 (TP) of leaf dry matter (Figure S1). However, antioxidant capacity (ORAC and TP) decreased consistently and significantly from the youngest (1200 µmole TE g−1 DM) to the oldest (600 µmoles TE g−1 DM) leaves (Figure 6). The leaf with the lowest antioxidant capacity was the leaf from the previous year, at the base of the vine generated that year. These results indicate that young leaves are physiologically more active and generate more antioxidants, possibly as protection from sunlight stress, while the lower leaves may be more protected from sunlight and less physiologically active, and produce smaller concentrations of antioxidants. Regardless of the variation in antioxidant capacities based on the age of the leaf, passion fruit leaves have a higher hydrophilic antioxidant capacity than alfalfa, the ORAC of which ranged from 244–287 µmoles TE g−1 DM and TP from 5–5.6 mg GAE g−1 DM at 300 days after sowing and cultivation under moderately to highly saline irrigation [22]. As the yellow passion fruit leaves had 3- to 6-fold higher ORAC and 4- to 5-fold higher TP than alfalfa, its leaves should be evaluated as a rich source of minerals, NaCl, and antioxidants that would be beneficial for small ruminants and that could be provided after each annual pruning when stems and leaves are discarded. Although Passiflora sp. leaves contain cyanogenic glycosides, we could find no reliable toxicity studies on leaf extracts in animals, nor any report of toxicity by animals that would preclude its feeding to ruminants.
When the antioxidant capacity (both ORAC and TP) of leaves dried through freeze-drying and oven drying at 50 °C was assessed, there were no significant differences (Figure 5). This indicates that oven drying leaves at 50 °C for 48 h can effectively substitute for freeze-drying when liquid nitrogen and a freeze-drier are unavailable. These results with passion fruit confirmed our previous results with the antimalarial plant Artemisia annua, where leaves dried by oven drying at 50 °C for 48 h had the same antioxidant capacity (ORAC) as leaves that were frozen in liquid nitrogen and freeze-dried [14]. However, shade-drying and sun-drying, mainly the latter, destroyed most of the antioxidant capacity of A. annua leaves.
4.3. Effect of Na and Cl Accumulation on Gene Expression
Expression analyses showed that the genes involved in Na and Cl transport studied in this investigation had higher expression in roots compared to leaves (Figure 7), suggesting that roots play a more critical role than leaves under salinity stress. Similar observations were made in alfalfa, where most genes involved in Na and Cl transport were expressed at higher levels in roots compared to leaves [30].
Under salinity stress, a higher Na+ concentration is sensed by SOS3, which physically interacts with SOS2 [31]. SOS3-SOS2 complex then activates a Na+/H+ antiporter, SOS1, by phosphorylating it, causing extensive Na+ exclusion from the cytoplasm [32]. SOS1 and SOS3, known to be involved in Na+ exclusion, were upregulated in roots under salinity, indicating that the Na+ exclusion is vital during salinity stress in passion fruit (Figure 7b). At the same time, the lower expression of SOS3 in leaves under salinity than the control may suggest an additional role of SOS3 in the cellular metabolism of passion fruit (Figure 7a).
ALMT12 is found in root stelar cells, and is predicted to be involved in the loading of Cl− from root to xylem [33]. ALMT12 was significantly downregulated in roots under salinity compared to the control in our study (Figure 7d). Downregulation of this gene will restrict Cl− to roots and will protect leaves from higher Cl concentrations.
CLCg is localized to the tonoplast of mesophyll cells, and is involved in sequestering Cl− to vacuoles to protect the cytoplasm [34]. CLCg was upregulated at 6 dS m−1 compared to the control, suggesting its role in tissue tolerance under medium salinity. The high salinity of 12 dS m−1 does not show induction of CLCg compared to the control in leaves (Figure 7c).
Some of the genes, such as AKT1, NHX1, NHX2, SOS2, ALMT9, CCC, and CLCc, which do not show differential expression under control and saline treatments may be differentially expressed in different genotypes. Although not tested in this investigation, genotypic-specific differences may also be critical during salinity stress in passion fruit.
5. Conclusions
This work is first to report on the effect of saline irrigation on the biomass, mineral status, antioxidant capacity, and genetic response of Passiflora edulis f. cv flavicarpa, as well as the first to compare low-temperature oven drying with freeze-drying and lyophilization prior to leaf antioxidant analyses. Our experiment produced the main findings: Saline waters up to 6.8 dS m−1 (equivalent to a soil-paste salinity (ECe) of 3.1 dS m−1) were perfectly tolerated by young passion fruit vines in their first year. During the second and third years, salinities from 3.0 dS m−1 (equivalent to an ECe = 1.36 dS m−1) to 6.0 dS m−1 (equivalent to an ECe = 2.7 dS m−1) produced no visual toxicity symptoms, leaf biomass reduction, major changes in leaf mineral status, or decreased leaf antioxidant capacity under the hot summer temperatures of southern California. However, plants irrigated with water with a salinity of 12.0 dS m−1 (equivalent to an ECe = 5.45 dS m−1) resulted in significantly reduced shoot biomass and some plant death in the third year, confirming previous reports that the salinity threshold for the yellow passion fruit in the field had an ECe of approximately 4.0 dS m−1. However, our experience with salinity tolerance in other crops suggests that salinity trials must involve other cultivars, which may increase salt tolerance in a crop of Amazonian origin such as Passiflora edulis. The high antioxidant capacity found in young leaves makes them appealing to the nutraceutical market. We also showed that freezing-drying in liquid nitrogen and lyophilization at −50 °C can be substituted by drying leaves in a forced-air oven set at 50 °C for 48 h, which is a more effective, rapid, and economical way to preserve leaf antioxidant capacity. However, oven-dried leaves should be further analyzed for individual flavonoid glucosides by HPLC-UV in order to guarantee that key flavonoids are preserved by low-temperature oven drying. The similar trends in TP and ORAC analyses indicate that the passion fruit flavonoids have high antioxidant activity. Finally, the gene expression analyses of passion fruit leaves of plants submitted to irrigation waters of low to high salinity confirmed a more critical role of roots compared to leaves under salinity stress. The efflux of Na+ from roots to the soil, loading of Cl− from root to xylem, and sequestration of Cl− into vacuoles of mesophyll cells are vital components of salinity tolerance in passion fruit. In order to increase our understanding of Passiflora edulis f. flavicarpa responses to salinity, future studies should involve different cultivars, of both edible and wild species, and separate saline waters dominant in either sodium or chloride salts to determine which salt is most toxic to the crop.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture12111856/s1, Figure S1: Leaf hydrophilic oxygen radical absorbance capacity (ORAC) and total phenolics (TP) concentration of passion fruit leaves of plants grown under three salinities and of leaves facing east or west. (a) Leaf ORAC at different salinities. (b) Leaf ORAC for samples taken from leaves facing east or west. (c) Leaf TP at different salinities. (d) Leaf TP for samples taken from leaves facing east or west. Error bars represent standard errors of the means. Table S1: The list of primers used for the expression analysis.
Author Contributions
Conceptualization, J.F.S.F. and D.S.; methodology, J.F.S.F., D.S., X.L., and S.R.P.S.; formal analysis, X.L. and C.N.; investigation, J.F.S.F. and D.S.; data curation, J.F.S.F., D.S., X.L., and C.N.; writing—original draft preparation, J.F.S.F., X.L., and D.S.; writing—review and editing, J.F.S.F. and D.S.; visualization, J.F.S.F. and D.S.; supervision, J.F.S.F.; project administration, J.F.S.F. and D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This was funded by the USDA Project Number 2036-13210-011-00D (Enhancing Specialty Crop Tolerance to Saline Irrigation Waters).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
To Noah Gangoso, for his valuable contribution in taking the lead in monitoring irrigation-water salinity and pH and for overseeing the irrigation of plants since its inception and final data collection. Thanks are also due to Jason Verd and Alysia Sorya for general care and pruning, and to Dennise Jenkins for her valuable help installing the irrigation system for seedling greenhouse production and for overseeing the installation of the trellis system outdoors. Thanks are also due to Jaime Barros for his help during the shoot biomass data collection during the COVID-19 pandemic.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Santos, J.T.C.; Petry, F.C.; Tobaruela, E.C.; Mercadante, A.Z.; Gloria, M.B.A.; Costa, A.M.; Lajolo, F.M.; Hassimotto, N.M.A. Brazilian native passion fruit (Passiflora tenuifila Killip) is a rich source of proanthocyanidins, carote-noids, and dietary fiber. Food Res. Int. 2021, 147, 110521. [Google Scholar] [CrossRef] [PubMed]
- Bernacci, L.C.; Soares-Scott, M.D.; Junqueira, N.T.V.; Passos, I.R.S.; Meletti, L.M.M. Passiflora edulis Sims: The correct taxonomic way to cite the yellow passion fruit (and of others colors). Rev. Bras. Frutic. 2008, 30, 566–576. [Google Scholar] [CrossRef]
- Sá, C.P.; Neto, R.C.A.; Negreiros, J.R.S.; Nogueira, S.R. Coeficientes Técnicos, Custos de Produção e Indicadores Economicos Para o Cultivo do Maracujá BRS Gigante Amarelo, No Acre; Embrapa: Rio Branco, Acre, Brazil, 2015. [Google Scholar]
- Altendorf, S. Minor Tropical Fruits: Mainstreaming a Niche Market; Food and Agricultural Organization: Rome, Italy, 2018; pp. 67–75. Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Tropical_Fruits/Documents/Minor_Tropical_Fruits_FoodOutlook_1_2018.pdf (accessed on 10 September 2022).
- Li, H.; Zhou, P.; Yang, Q.; Shen, Y.; Deng, J.; Li, L.; Zhao, D. Comparative studies on anxiolytic activities and fla-vonoid compositions of Passiflora edulis ‘edulis’ and Passiflora edulis ‘flavicarpa’. J. Ethnopharmacol. 2011, 133, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Newall, C.A.; Anderson, L.A.; Phillipson, J.D. Herbal Medicines. A Guide for Health-Care Professionals; The Pharmaceutical Press: London, UK, 1996; p. 296. [Google Scholar]
- Fisher, A.A.; Purcell, P.; Couteur, D.G.L. Toxicity of Passiflora incarnata L. J. Toxicol. Clin. Toxicol. 2000, 38, 63–66. [Google Scholar] [CrossRef]
- Machado, C.F.; Jesus, F.N.; Ledo, C.A.S. Divergencia genetica de acessos de maracuja utilizando descritores quantitativos e qualitativos. Rev. Bras. Frutic. 2015, 37, 442–449. [Google Scholar] [CrossRef]
- Dias, T.J.; Cavalcante, L.F.; Pereira, W.E.; Freire, J.L.D.O.; Souto, A.G.D.L. Irrigação com água salina em solo com biofertilizante bovino no crescimento do maracujazeiro amarelo. Semin. Cienc. Agrar. 2013, 34, 1639–1652. [Google Scholar] [CrossRef]
- Freire, J.L.O.; Cavalcante, L.F.; Rebequi, A.M.; Dias, T.J.; Brehm, M.A.S.; Santos, J.B. Quality of yellow passion fruit juice with cultivation using different organic sources and saline water. IDESIA 2014, 32, 79–87. [Google Scholar] [CrossRef][Green Version]
- Cavalcante, L.F.; Cavalcante, Í.H.L.; Júnior, F.R.; Beckmann-Cavalcante, M.Z.; Santos, G.P. Leaf-macronutrient status and fruit yield of biofertilized yellow passion fruit plants. J. Plant Nutr. 2012, 35, 176–191. [Google Scholar] [CrossRef]
- Lima, L.K.S.; Jesus, O.N.; Soares, T.L.; Santos, I.S.; Oliveira, E.J.; Filho, M.A.C. Growth, physiological, ana-tomical and nutritional responses of two phenotypically distinct passion fruit species (Passiflora L.) and their hybrid under saline conditions. Sci. Hort. 2020, 263, 109037. [Google Scholar] [CrossRef]
- Reis, L.C.R.; Facco, E.M.P.; Salvador, M.; Flôres, S.H.; Rios, A.O. Antioxidant potential and physicochemical characterization of yellow, purple and orange passion fruit. J. Food Sci. Technol. 2018, 55, 2679–2691. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Luthria, D.L. Drying affects artemisinin, dihydroartemisinic acid, artemisinic acid, and the anti-oxidant capacity of Artemisia annua L. leaves. J. Agric. Food Chem. 2010, 58, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira-Silva, C.B.M.; Jesus, O.N.; Santos, E.S.L.; Corrêa, R.X.; Souza, A.P. Genetic breeding and diversity of the genus Passiflora: Progress and perspectives in molecular and genetic studies. Int. J. Mol. Sci. 2014, 15, 14122–14152. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira-Silva, C.B.M.; Faleiro, F.G.; Jesus, O.N.; Santos, E.S.L.; Souza, A.P. The Genetic Diversity, Conservation, and Use of Passion Fruit (Passiflora spp.). In Genetic Diversity and Erosion in Plants: Case Histories; Ahuja, M.R., Jain, S.M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 215–231. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Suarez, D.L.; Simunek, J. UNSATCHEM: Unsaturated water and solute transport model with equilibrium and kinetic chemistry. Soil Sci. Soc. Am. J. 1997, 61, 1633–1646. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for hydrophilic and lipophilic antioxidant capacity [oxygen radical absorbance capacity (ORAC)] of plasma and other biological and food samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Sandhu, D.; Liu, X.; Halvorson, J.J. Spinach (Spinacea oleracea L.) response to salinity: Nutritional value, physiological parameters, antioxidant capacity, and gene expression. Agriculture 2018, 8, 163. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Cornacchione, M.V.; Liu, X.; Suarez, D.L. Nutrient composition, forage parameters, and antioxi-dant capacity of alfalfa (Medicago sativa, L.) in response to saline irrigation water. Agriculture 2015, 5, 577–597. [Google Scholar] [CrossRef]
- Munhoz, C.F.; Santos, A.A.; Arenhart, R.A.; Santini, L.; Monteiro-Vitorello, C.B.; Vieira, M.L.C. Analysis of plant gene expression during passion fruit–Xanthomonas axonopodis interaction implicates lipoxygenase 2 in host de-fence. Ann. Appl. Biol. 2015, 167, 135–155. [Google Scholar] [CrossRef]
- Freitas, M.S.M.; Monnerat, P.H.; Vieira, I.J.C. Mineral deficiency in Passiflora alata Curtis: Vitexin bioproduction. J. Plant Nutr. 2008, 31, 1844–1854. [Google Scholar] [CrossRef]
- Hurtado-Salazar, A.; Silva, D.F.P.; Ceballos-Aguirre, N.; Ocampo-Pérez, J.; Bruckner, C.H. Promissory Passiflora L. species (Passifloraceae) for tolerance to water-salt stress. Rev. Colomb. Cienc. Hortic. 2020, 14, 44–49. [Google Scholar] [CrossRef]
- Soares, F.A.L.; Gheyi, H.R.; Viana, S.B.A.; Uyeda, C.A.; Fernandes, P.D. Water salinity and initial development of yellow passion fruit. Sci. Agric. 2002, 59, 491–497. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Fruit yield and survival of five commercial strawberry cultivars under field cultivation and salinity stress. Sci. Hort. 2019, 243, 401–410. [Google Scholar] [CrossRef]
- Suarez, D.L.; Celis, N.; Ferreira, J.F.S.; Reynolds, T.; Sandhu, D. Linking genetic determinants with salinity toler-ance and ion relationships in eggplant, tomato and pepper. Sci. Rep. 2021, 11, 16298. [Google Scholar] [CrossRef] [PubMed]
- Celis, N.; Suarez, D.L.; Wu, L.; Li, R.; Arpaia, M.L.; Mauk, P. Salt tolerance and growth of 13 avocado rootstocks related best to chloride uptake. HortScience 2018, 53, 1737. [Google Scholar] [CrossRef]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Qiu, Q.-S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.-K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the Move. Trends Plant Sci. 2017, 22, 236–248. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Agorio, A.; Jossier, M.; Depré, S.; Thomine, S.; Filleur, S. Characterization of the chloride channel-like, AtCLCg, involved in chloride tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 764–775. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).