Abstract
The production of cucumber (Cucumis sativus L.) is often harmed by biotic and abiotic stresses. Although the dehydration-responsive element-binding (DREB) transcription factors, playing vital roles in stress responses, have been characterized in several plant species, little is known about the pan-genome characteristics of DREB genes and their expression patterns under different stresses in cucumber. In this study, we identified 55 CsDREBs from the cucumber pan-genomes of 13 accessions, but only four accessions had all the genes. Most of the CsDREB proteins had sequence length and/or amino acid variations, and only four of them had no variation among different accessions. Using the 55 CsDREBs from ‘9930’, we analyzed their gene structures, conserved domains, phylogenetic relationships, gene promoter’s cis-elements and syntenic relationships, and classified them into six groups. Expression pattern analysis revealed that eight CsDREBs showed constitutive expression (FPKM > 1 in all samples), and different CsDREBs showed specifically high expression in root, stem, leaf, tendril, male-flower, female flower, and ovary, respectively, suggesting that these genes might be important for morphogenesis and development in cucumber. Additionally, a total of 31, 22, 30 and nine CsDREBs were differentially expressed in responding to the treatments of heat, NaCl and/or silicon, power mildew and downy mildew, respectively. Interestingly, CsDREB33 could respond to all the tested stresses. Our results provide a reference and basis for further investigation of the function and mechanism of the DREB genes for resistance breeding in cucumber.
1. Introduction
Plants encounter many adverse environments in nature, which can affect their normal growth and development, and can even lead to their death [1,2]. Environmental stresses are generally divided into biotic stresses, such as bacterial or fungal stress, and abiotic stresses, such as temperature, drought, or salt stress [3,4]. Meanwhile, plants have developed many defense mechanisms including a broad regulation of numerous genes to mediate plant physiological and biochemical processes to better survive such stresses. These defense mechanisms are orchestrated within a complex regulatory network, such as reactive oxygen species (ROS), the stress-related hormones (e.g., ethylene) and secondary metabolites [5,6,7]. Transcription factors (TFs) are one of the key components of defense mechanisms in plants, which can regulate signal transduction by binding to specific cis-elements in the promoter regions of target genes to adapt to diverse stresses [8]. Many TFs, such as the AP2/ERF (APETALA2/ethylene response factor), NAC, MYB (v-myb avian myeloblastosis viral oncogene homolog), bHLH (basic helix–loop–helix), bZIP (basic leucine zipper), and WRKY families, have been reported to play very important roles in the defense mechanisms of plants against stresses [9,10,11]. For instance, in wheat, bHLH39 regulates the expression level of stress-responsive genes, making them show higher expression levels, thus increasing the tolerance to salt stress [12]. The WRKY TFs from Arabidopsis [13] and tomato [14] were shown to function in the response to various biotic and abiotic stresses.
The TF family AP2/ERF is one of the most important TF groups in plants [9]. Each AP2 family member contains one or two conserved domains of 60–70 amino acids called AP2/ERF domains. The AP2/ERF family genes can be divided into four subfamilies [15]: AP2, RAV, ERF, and Soloist. The ERF subfamily can be further divided into ERF and C-repeat-binding factor (CBF)/dehydration-responsive element-binding factor (DREB) subfamilies [16]. The DREB proteins contain only one AP2/ERF domain, but are different from the ERF subfamily based on the conserved amino acids of valine (Val) at position 14 (a key site for interaction) [17]. The DREB subfamily can be divided into six groups (A1–A6) [16,18] or four clades (I–IV) [19,20]. The first DREB was found to regulate the transcription of drought-responsive genes by binding to the DRE/CRT (dehydration-responsive element/C-repeat element; core sequence A/GCCGAC) sequence in Arabidopsis [21]. Since then, the functions of DREBs in abiotic stresses have gradually been discovered. For instance, GmDREB responds to drought [22], AtCBF1 and OsDREB respond to low temperatures [23,24], CaDREBLP1 responds to high salinity [25], ZmDREB2A responds to heat [26], CkDREB responds to osmotic stress [27], and AtDREB1A responds to multiple stresses, such as drought and heat [28]. Additionally, DREBs have now been identified as playing important roles in stress resistance in many plant species, such as Arabidopsis [29], rice [30], bell pepper [25], soybean [31], pearl millet [32], wheat [33], maize [34], chrysanthemum [35], tomato [36], lettuce [37], Chinese jujube [38], mulberry [39], sugarcane [40,41], pineapple [42], and potato [43].
Cucumber (Cucumis sativus L.) is a popular economic crop because of its unique taste [44,45]. However, many biotic and abiotic stresses are threats to cucumber production [46,47]. Therefore, it is important to identify stress-resistance genes—such as DREBs—in cucumber. In a previous study, 42 DREBs were identified from the version 1 (v1) genome of cucumber [48,49]. However, with the emergence of better genome versions, v1 has been deleted from the cucumber genome database because of its low quality. Moreover, more genomes of different accessions have been re-sequenced in this species, because a single genome cannot represent the genetic diversity of the species [50]. Therefore, the concept of a pan-genome was put forward; this concept was first developed in bacteria [51], and refers to the total genes of a species. Recently, a graph-based pan-genome has been successfully constructed for cucumber, providing a very important reference for the identification of cucumber’s functional genes [52].
In this study, we performed a pan-genome-wide identification of DREBs to analyze their variation and potential functions in the different cucumber accessions [52]. We identified a total of 55 CsDREB genes in the pan-genome of cucumber and found that most of them vary in protein length between the 13 accessions. Then, we further analyzed the chromosomal location, gene structure, and phylogeny of these DREB genes. The 55 CsDREBs could be classified into six groups based on their phylogenetic relationships. In addition, we analyzed the expression patterns of the CsDREB genes using transcriptomic data under different stresses and identified some CsDREB genes that may play roles in different stresses. Therefore, our study provides a basis for discovering potential functions of CsDREBs for breeding stress resistance in cucumber.
2. Materials and Methods
2.1. Gene Identification and Chromosomal Distribution
The cucumber genome data were downloaded from the Cucurbit Genomics Database (http://cucurbitgenomics.org/, accessed on 21 November 2021) and NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 21 November 2021). The hidden Markov model (HMM) profile files of the AP2 domain (PF00847) were downloaded from the Pfam database (http://pfam.xfam.org/, accessed on 22 November 2021). The DREB genes of cucumber were identified from the genome database using HMMER 3.0 with the default parameters and a cutoff value of 0.01. Candidate genes were screened with only one AP2 domain and its 14th amino acid Val; all CsDREB genes were further examined to confirm the AP2 domain through the CDD (https://www.ncbi.nlm.nih.gov/cdd/, accessed on 22 November 2021), Pfam (http://pfam.xfam.org/search#tabview=tab1, accessed on 22 November 2021), and SMART (http://smart.embl-heidelberg.de/, accessed on 22 November 2021) online tools.
Multiple DREB protein sequences were aligned using ClustalW (http://www.genome.jp/tools/clustalw/, accessed on 26 November 2021) with the default parameters, and the results of the alignment were visualized using the GeneDoc program. The protein sequences of DREB in different cucumber accessions were uploaded to the MEGA software (v5.0) to be aligned using ClustalW, and the phylogenetic relationships among all DREB proteins were examined via the neighbor-joining method with 1000 bootstrap replicates. The cluster analysis of the phylogenetic branches was used to confirm the correspondence of the DREB genes in different cucumber accessions.
Chromosome location information was extracted from the genomic database, and CsDREB genes were mapped using the MapChart program [53].
2.2. Protein Length, Sequence, and Motif Composition Analysis
The protein sequences of CsDREBs in different cucumber accessions were extracted and the proteins’ lengths were counted using ExPASy (https://web.expasy.org/compute pi/, accessed on 26 November 2021). The variation of amino acids was analyzed using DNAMAN program. The conserved motifs were identified using the MEME program V5.0.4 (http://alternate.meme-suite.org/tools/meme, accessed on 26 November 2021). The protein sequences of the CsDREB genes were submitted to the MEME online system with the following parameters—number of repetitions: any; maximum number of motifs: 10; optimal motif width: 6–100 amino acid residues.
2.3. Gene Structure Analysis and Cis-Element Prediction
The location information of CDSs and UTRs was extracted from the genomic annotation database and graphed using TBtools [54]. The elements in the 2.0 kb promoter fragments (i.e., upstream sequences of the CsDREBs-coding sequences) were extracted and analyzed using the online program PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 29 November 2021) [55]. The results were visualized using TBtools [56].
2.4. Multiple Sequence Alignment and Phylogenetic Analysis
The protein sequences of DREB in cucumber and Arabidopsis [33] were uploaded to the MEGA software (v5.0) to be aligned using ClustalW, and the phylogenetic relationships among all DREB genes were examined via the neighbor-joining method with 1000 bootstrap replicates. Then, the phylogenetic trees were landscaped in Evolview (https://evolgenius.info//evolview-v2/#login, accessed on 29 November 2021).
2.5. Gene Duplication and Synteny Analysis
The genomic databases of cucumber, Arabidopsis, rice, tomato, and maize were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 29 November 2021), and then, the gene duplication events and the syntenic relationships were obtained using the Multiple Collinearity Scan toolkit (MCScanX) [57] with the default parameters. The results were constructed using TBtools [54].
2.6. Transcriptome Analysis of CsDREB Genes in Cucumber
The expression patterns of the CsDREB genes were analyzed using the transcriptomic data of the roots, stems, leaves, flowers, ovaries, and tendrils of cucumber. The published RNA-Seq data (SRA046916) [58] were downloaded from the Cucurbit Genomics Database (http://cucurbitgenomics.org/, accessed on 7 December 2021). The remapped clean tags and the recalculated FPKM values were cited to analyze the expression patterns of the CsDREBs. The genome-wide expression of the CsDREB genes was shown on a heatmap using TBtools [54]. The heatmap values were calculated according to the following steps: the fold change values of the FPKM value of the treatment group and the control group were calculated first, and then the logarithm based on two of the fold change values were taken.
2.7. Transcriptome Analysis of CsDREBs in Response to Abiotic and Biotic Stresses
The publicly available transcriptomic data of cucumber treated with salt (GSE116265) [59], heat (GSE151055) [60], DM (SRP009350) [46], and PM (GSE81234) [61] were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 21 November 2021) to analyze the expression patterns of CsDREBs under different stresses. After aligning the gene IDs to the cucumber genome, the genome-wide expression of the CsDREB genes was shown on a heatmap using TBtools [54]. For the transcriptome analysis of the CsDREBs, a threshold of FDR (or p-value) ≤ 0.05 and an absolute value of log2 (fold-change) ≥ 1 or log2 (fold-change) ≤ −1 were used to define DEGs.
2.8. Plant Materials, Stress Treatments, and qRT-PCR Analysis
Cucumber ‘Chinese long 9930’, ‘JinYou 1’ [59], along with the DM-susceptible cultivar ‘Vlaspik’ [46], the PM-susceptible cultivar D8, and the PM-resistant cultivar SSL508-28 [61], were used. Cucumber seedlings were grown in a growth chamber under day/night temperatures of 26/18 °C, with a 16 h/8 h light/dark period. The relative humidity was maintained at 60–70%. The light intensity was 200 µmol·m−2·s−1 in the day. Water and pest control were provided according to standard protocols. The leaves of the three-week-old seedlings (‘Chinese Long 9930’) were taken at 0, 3 and 6 h after treatment with 42 °C for transcriptome sequencing in Novogene (Beijing, China) and quantitative real-time PCR (qRT-PCR) analysis. Seedlings of cucumber ‘JinYou 1’ were subjected to salt stress (75 mM NaCl) for three days with or without the addition of 0.3 mM Si. First, seedlings at the two-leaf stage were transferred to 15-L plastic containers with aerated 1/4-strength modified Hoagland nutrient solution for three days [59]. Second, the strength of Hoagland solution was increased to 1/2 for four days [59]. Finally, Na2SiO3·9H2O and NaCl were added to the nutrient solution, and the Si (0.3 mM) and salt (75 mM) treatments were started [59]. There were four treatments: control (CT), Si treatment (Si), salt stress (NaCl), and salt stress plus Si (NaSi) [59]. For the DM treatment, cucumber ‘Vlaspik’ leaves were inoculated with Pseudoperonospora cubensis over an 8-day period. Samples were collected at 1, 2, 3, 4, 6, and 8 days post-inoculation (dpi) [46]. For the PM treatment, normally watered control leaves and leaves inoculated with PM for 48 h from the SSL508-28 and D8 lines were collected [61]. Samples were immediately frozen in liquid nitrogen and stored at −80 °C until further use. All experiments were conducted at least three times with independently collected and extracted tissues unless noted otherwise. The total RNA was extracted with the TRIzol reagent, and then cDNA was reverse-transcribed using polyT primers. The qRT-PCR experiment was conducted with SYBR Premix Ex Taq (Mei5bio, Beijing, China) with an ABI 7500 Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). The 2−∆∆Ct method was used to analyze the qRT-PCR results, and the primer sequences designed by Primer 5 are provided in Table S1.
3. Results
3.1. Identification of Genes and the Comparison of Gene Characterizations in Different Cucumber Accessions
With the construction of a graph-based pan-genome, the gene information in cucumber has been updated [52]. Therefore, we could identify cucumber DREB genes in different accession genomes. We downloaded the HMM profile files and extracted CsDREB proteins from the cucumber genome database. Afterwards, the ones with only one AP2 domain with a V (valine acid) at the 14th amino acid position were selected as CsDREB proteins. All the candidate genes were further detected through NCBI CDD, SMART, and Pfam. A total of 55 CsDREB genes were identified in the genomes of different cucumber accessions (Table 1). There were 55 CsDREB genes identified from ‘9930’, ‘Cuc37’, ‘Gy14’, ‘PI183967’, and ‘Cuc64’; 54 from ‘Cu2’, ‘W4’, ‘W8’, and ‘Hx14’; 52 from ‘Hx117’; 51 from ‘XTMC’; 41 from ‘9110gt’, and 40 from ‘Cuc80’. After sequence alignment, we renamed them CsDREB01–CsDREB55 based on their order on the chromosomes to avoid confusion in this study (Table S2).

Table 1.
The origins of the different cucumber accessions.
The 55 CsDREB genes were unevenly distributed on seven chromosomes of cucumber (Figure S1). Most of them were distributed on chromosomes 2, 3, and 4; only three and four CsDREB genes were distributed on chromosomes 1 and 7, respectively. It is worth noting that CsDREB08 was localized in different positions in different genomes (Figure S1)—it was on chromosome 1 in the ‘Chinese Long 9930’ genome, but on chromosome 2 in the genomes of another 10 accessions. In addition, there were two copies of CsDREB09 in the ‘PI183967’ genome and they were located in different positions on chromosome 2 (Figure S1).
To further understand whether there are differences in CsDREB genes between different cucumber accessions, we measured the length of the identified CsDREB proteins (Table 2). There were 13 CsDREBs with the same protein length among 13 different cucumber accessions, namely, CsDREB01, CsDREB04, CsDREB09, CsDREB12, CsDREB16, CsDREB20, CsDREB28, CsDREB30, CsDREB31, CsDREB32, CsDREB35, CsDREB53, and CsDREB54. The length of CsDREB06, CsDREB18, CsDREB23, CsDREB25, CsDREB27, CsDREB33, and CsDREB37 differed in only one of the accessions. CsDREB02, CsDREB03, CsDREB13, CsDREB15, CsDREB24, CsDREB36, CsDREB40, CsDREB41, and CsDREB55 showed differences in protein length in two accessions, while others showed differences in protein length among multiple accessions. Length difference data are marked in red in Table 2. Among the proteins with different lengths, CsDREB05 in ‘Hx14’; CsDREB06 in ‘Hx117’; CsDREB08 in ‘PI183967’, ‘Cuc64’, and ‘W8’; CsDREB11 in ‘9930’; CsDREB13 in ‘9110gt’; CsDREB18 in ‘Gy14’; CsDREB37 in ‘Cu2’; CsDREB41 in ‘PI183967’; and CsDREB45 in ‘Cu2’ had shorter lengths compared to those in other accessions, while CsDREB07 in ‘XTMC’, ‘Cuc37’, ‘9110gt’, and ‘W8’; CsDREB22 in ‘Cuc37’, ‘W4’, ‘W8’, ‘Hx14’, and ‘Hx117’; CsDREB24 in ‘W4’; and CsDREB25 in ‘9110gt’ were longer than in other accessions. In addition, the lengths of some proteins showed accession-specificity. For example, the protein length of CsDREB17 was the same in ‘Cu2’, ‘Cuc80’, ‘Cuc37’, ‘Hx14’, and ‘Hx117’, but different in other accessions.
Table 2.
The protein lengths of CsDREBs in different cucumber accessions.
Besides protein length, amino acid substitution also can change a protein’s function. The amino acid variations of CsDREBs in different cucumber accessions was analyzed, and the results were shown in Table S3. Among CsDREB proteins, only four of them had no variation among different accessions, they were CsDREB09, CsDREB12, CsDREB20 and CsDREB31. In addition, CsDREB04, CsDREB08, CsDREB28, CsDREB30 and CsDREB32 had only one amino acid difference among different accessions. Meanwhile, though there was only one amino acid difference in CsDREB10, CsDREB19 and CsDREB45, respectively, it led to a reading frame shift, resulting in big differences of many amino acids. Reversely, several proteins, such as CsDREB23, CsDREB27, CsDREB44, CsDREB46 and CsDREB48, had great differences with InDel and amino acid variations among different accessions.
Among the 13 examined accessions, only four (‘9930’, ‘Cuc37’, ‘Gy14’, and ‘PI183967’) had 55 DREBs (Table 2). Therefore, the subsequent analyses were mainly based on the genes identified in the genome of ’9930’, because it had all 55 CsDREBs, and its genome was the first to be sequenced and has been updated into v3.
3.2. Multiple Sequence Alignment and Phylogenetic Relationships of CsDREBs
To explore the phylogenetic relationships of the CsDREB proteins, we first performed multiple sequence alignment. The results showed that 55 CsDREBs have a conserved AP2 domain and other typical features of DREBs. In the AP2 conserved domain, in addition to the conserved V14, the YRG, WLG, and RAYD motifs are quietly conserved (Figure 1).

Figure 1.
The multiple protein sequence alignment of the AP2 domains of DREBs from cucumber: Conserved V14, YRG, RAYD, and WLG motifs are highlighted by the asterisks and lines. Conserved sequences are highlighted by black and grey shading; the black shading represents completely conserved sequences, while the grey shading represents incompletely conserved sequences.
To clarify the phylogenetic relationships among the CsDREBs, a phylogenetic tree was constructed by the multiple sequence alignment of the DREB proteins from cucumber and Arabidopsis (Figure 1). The proteins of CsDREBs and AtDREBs were multiply aligned using ClustalW, and a phylogenetic tree was constructed via the neighbor-joining (NJ) method using MEGA 7.0.4 software with 1000 bootstrap tests. As shown in Figure 1, CsDREBs can be divided into six groups (A1–A6). This classification is based on a previous study by Nakano et al. [1]. The classification standard is provided by the different conserved amino acids in different groups. Based on the previous study, group A6 contains CMI motifs, group A5 contains CMII motifs, groups A4 and A1 contain CMIII motifs, and group A2 contains CMIV motifs (Figure S2). Among the 55 CsDREB proteins, 20 were placed in group A5 to form the largest branch, and only one CsDREB protein was placed in group A3 to form the smallest branch. For the other groups, four CsDREB proteins were placed in group A1, six in Group A2, 16 in Group A4, and eight in Group A6 (Figure 2).
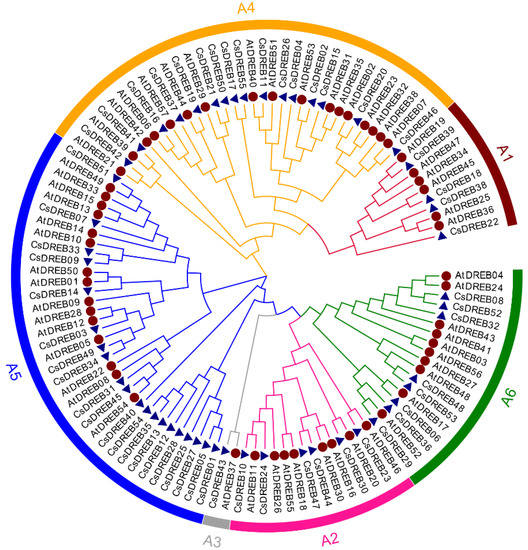
Figure 2.
The phylogenetic tree of the DREB proteins from cucumber and Arabidopsis: Different colors represent different subgroups of the DREB family. The blue triangles and red circles represent DREB proteins of cucumber and Arabidopsis, respectively.
3.3. Gene Structure, Motif Composition, and Cis-Element Analysis of CsDREBs
To reveal the evolutionary relationships, we analyzed the distribution of introns and exons among the 55 CsDREB genes. As shown in Figure 3, most (43 of 55) CsDREB genes harbor no introns, only one CsDREB gene (CsDREB27) harbors two introns, and others harbor one intron. Moreover, the genes containing introns mainly belong to groups A2 and A5. The UTR (untranslated region) is the untranslated 5′- or 3′- end of the mature mRNA and generally plays an important role in mRNA transport, stability, and translational regulation. However, few of the CsDREBs have UTRs.
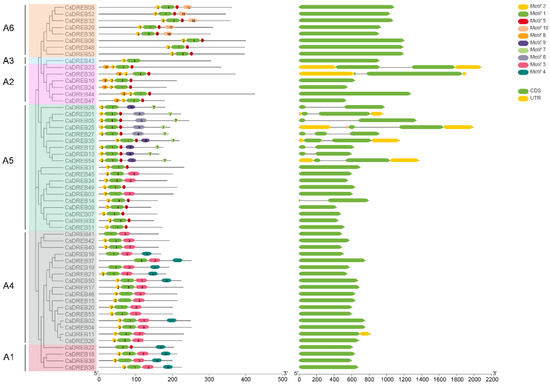
Figure 3.
Analysis of phylogenetic relationships, gene structures, and motifs in cucumber: Left panel—the phylogenetic tree was constructed from the sequences of CsDREB proteins. The different groups are shown in different colors. Middle panel—different colored boxes represent the motifs of CsDREB; The details of each motif are shown in Table S4. Right panel—gene structure of CsDREB TFs; 5′- and 3′-regions, exons, and introns are indicated by green boxes, yellow boxes, and grey lines, respectively. The gray numbered line at the bottom represents gene length.
To better understand the conservation and diversification of CsDREBs, the putative conversed motifs of CsDREB proteins were predicted via MEME motif analysis, and the CsDREBs that were categorized into the same groups shared highly similar motif compositions (Figure 3). For example, only the members of group A2 contained motif 8.
In addition, we submitted the 2.0 kb sequence upstream of the translation start site of CsDREB genes to PlantCARE to identify cis-elements. After removing redundant findings from the resulting file, two types of putative cis-elements were related to hormone signaling and stress response: one type included MeJA-RE (methyl-jasmonic-acid-responsive element), ARE (auxin-responsive element), ABRE (abscisic-acid-responsive element), SARE (salicylic-acid-responsive element), and GARE (gibberellin-responsive element); the other type included LTR (low-temperature-responsive element), DRE (dehydration-responsive element), HSE (heat-stress-responsive element), the WUN-motif (wound-responsive element), and TC-rich repeats (defense- and stress-responsive elements) (Figure 4). These results suggest that CsDREBs may be involved in many resistance processes.
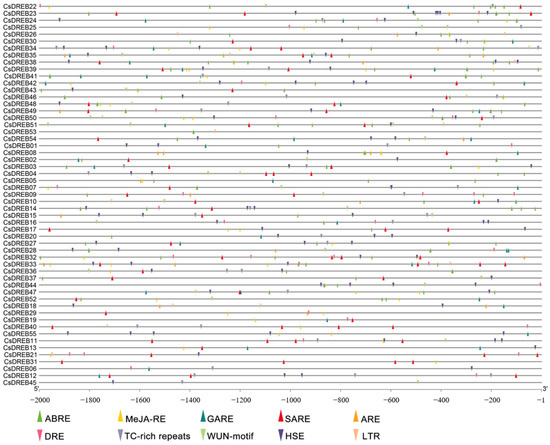
Figure 4.
The cis-acting elements of the DREB genes: TC-rich repeats, defense- and stress-responsive elements; WUN-motif, wound-responsive element; LTR, low-temperature-responsive element; DRE, dehydration-responsive element; HSE, heat-stress-responsive element; GARE, gibberellin-responsive element; SARE, salicylic-acid-responsive element; ABRE, abscisic-acid-responsive element; ARE, auxin-responsive element; MeJA-RE, methyl-jasmonic-acid-responsive element. The gray numbered line at the bottom represents promoter length.
3.4. Gene Duplication and Synteny Analysis
To further explore the evolutionary relationships of DREBs in cucumber and other different species, we first analyzed the ratio of the number of nonsynonymous substitutions per nonsynonymous site (Ka) to the number of synonymous substitutions per synonymous site (Ks). The Ka/Ks values were used to determine species selection pressure. If Ka/Ks > 1, the gene is under positive selection; if Ka/Ks = 1, the gene is under neutral evolution; if Ka/Ks < 1, the gene is under purifying selection. We identified a total of 10 pairs of tandem repeat genes; their Ka/Ks values were <1 or even <0.1. This suggests that they had experienced strong purifying selection (Table S5).
Other than that, we also analyzed the synteny of the DREB genes in cucumber and other species. A total of 16 pairs of genes had co-linearity in cucumber (Figure 5).
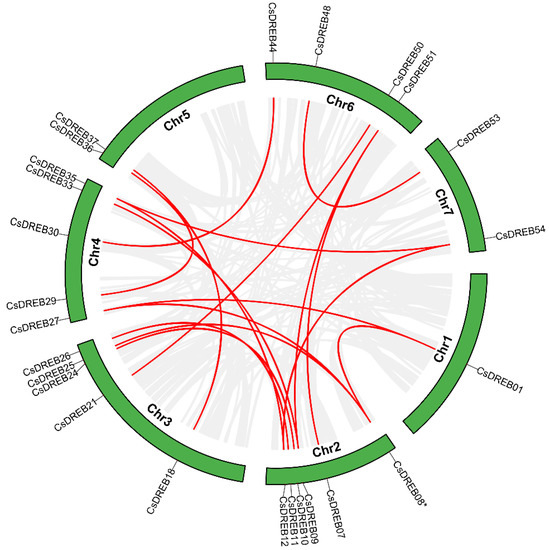
Figure 5.
Schematic representations for the inter-chromosomal relationships of CsDREBs: Gray lines show synteny blocks in the cucumber genome. Duplicated CsDREB gene pairs are connected with red lines.
Between cucumber and other species, there are also different numbers of collinear genes. There were 36, 43, 16, and 8 pairs of DREB genes that showed syntenic relationships with Arabidopsis, tomato, rice, and maize, respectively. It is clear that the number of homologous genes between dicotyledonous plants (i.e., 36 between cucumber and Arabidopsis, 43 between cucumber and tomato) is significantly higher than that between dicotyledonous and monocotyledonous plants (i.e., 16 between cucumber and rice, 8 between cucumber and maize); this is also consistent with the characteristics of biological evolution (Figure 6). Interestingly, all the collinear gene pairs between cucumber and maize were observed between cucumber and all the other three species, suggesting that these genes are highly conserved during evolution.
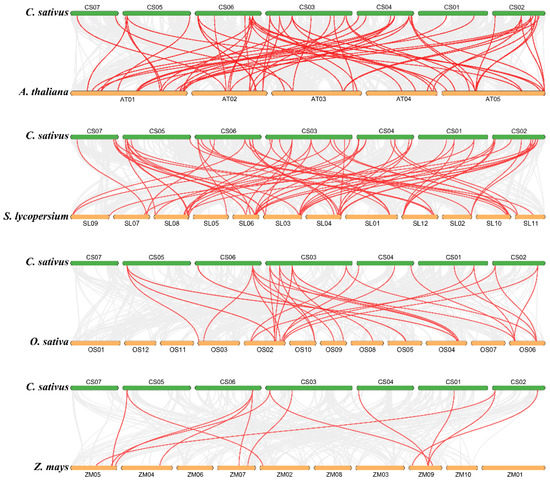
Figure 6.
Synteny analysis of DREBs between cucumber and other plant species: The collinear blocks are marked by gray lines, while the collinear gene pairs with DREB genes are highlighted by red lines. ‘C. sativus’, ‘A. thaliana’, ‘S. lycopersicum’, ‘O. sativa’, and ‘Z. mays’ indicate Cucumis sativus, Arabidopsis thaliana, Solanum lycopersicum, Oryza sativa, and Zea mays, respectively.
3.5. Expression Patterns of CsDREB in Different Tissues
To explore the expression patterns of DREB genes in cucumber, we analyzed the expression of these genes in various tissues of cucumber using public transcriptomic data. The gene expression heatmap reveals that different DREB genes are expressed in different tissues (Figure 7). Twelve genes are mainly expressed in the stem, fourteen genes are mainly expressed in the tendrils, eleven genes are mainly expressed in the male flowers, sixteen genes are mainly expressed in the roots, and several sporadic genes are mainly expressed in other tissues. However, the expression of CsDREB21 and CsDREB36 could hardly be detected in all tissues. This indicates that DREB family genes may play various roles in cucumber’s growth and development.
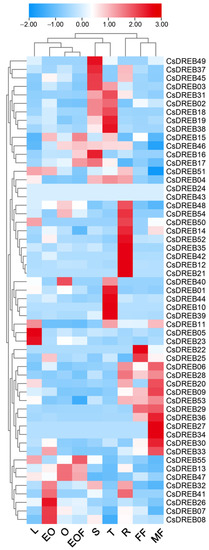
Figure 7.
Tissue-specific expression of DREB genes in cucumber: The genome-wide expression of CsDREB genes on a heatmap using FPKM values. A range of −2.00 to 3.00 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. S, stem; T, tendril; FF, female flower; EO, expanded unfertilized ovary; L, leaves; O, ovary; EOF, expanded fertilized ovary; MF, male flower; R, root.
3.6. Expression Profiles of CsDREB Genes under Abiotic and Biotic Stresses
As DREB genes in many species have been identified as being involved in various stress responses, we analyzed the comprehensive expression patterns of CsDREB genes under different stresses—including salt, heat, downy mildew (DM, Pseudoperonospora cubensis), and powdery mildew (PM, Podosphaera fusca)—based on public transcriptome information, to further explore the roles of CsDREB genes under different stresses.
First, we analyzed the roles of CsDREB genes under salt stress. The transcriptomic data were presented as a heatmap (Figure 8B). Under treatment with NaCl, most of the DREB genes in cucumber were downregulated, and only a few were upregulated, including CsDREB07, CsDREB33, CsDREB41, and CsDREB05. Silicon (Si) is considered to be the fourth most essential element for plants, enhancing resistance to stresses and promoting plant growth and development. Under the conditions treated with Si only, there were also many changes in gene expression. The expression of CsDREB38, CsDREB34, CsDREB30, CsDREB03, CsDREB07, and CsDREB33 was upregulated, whereas the expression of CsDREB06, CsDREB52, CsDREB10, CsDREB28, and CsDREB36 was downregulated. Nevertheless, CsDREB33 and CsDREB07 were more upregulated after the exogenous salt and Si treatment. It is noteworthy that the uniformity of CT-1 is poor, so CT-2 and CT-3 were considered to be the main reference samples.
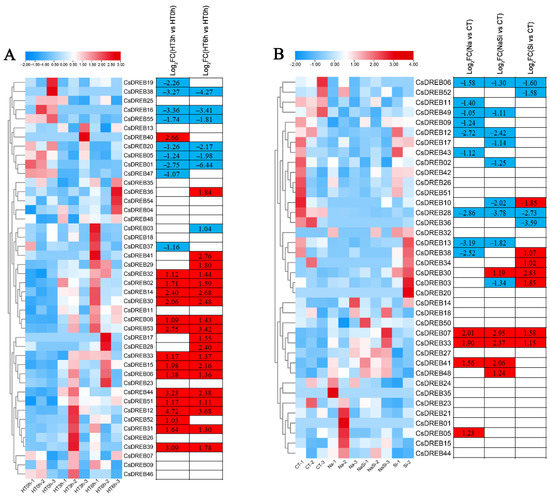
Figure 8.
Expression profiles of CsDREB genes in response to various abiotic stress treatments: The transcriptional levels of CsDREB genes in response to heat (A) and salt (B) stresses are shown on the heatmap. A range of −3.00 to 3.00 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. CT, control treatment; HT, heat treatment; HT0h, heat treatment for 0 h (hours); HT3h, heat treatment for 3 h; HT6h, heat treatment for 6 h; FC, fold-change.
We also analyzed the responses of CsDREB genes to heat stress (Figure 8A). At three hours after high-temperature treatment, CsDREB19, CsDREB38, CsDREB16, CsDREB55, CsDREB20, CsDREB05, CsDREB01, and CsDREB47 were downregulated, while CsDREB14, CsDREB30, CsDREB53, CsDREB15, CsDREB44, CsDREB12, and CsDREB39 were upregulated. Similarly, at six hours after heat stress, similar changes had taken place in the same gene set. Particularly, the genes upregulated at three and six hours after heat stress were virtually the same, suggesting that these genes may play important roles in thermotolerance. The transcriptional levels of six selected CsDREB genes under heat stress were further determined by qRT-PCR. Those genes effectively had the same expression patterns as those shown in the RNA-seq results (Figure 9).
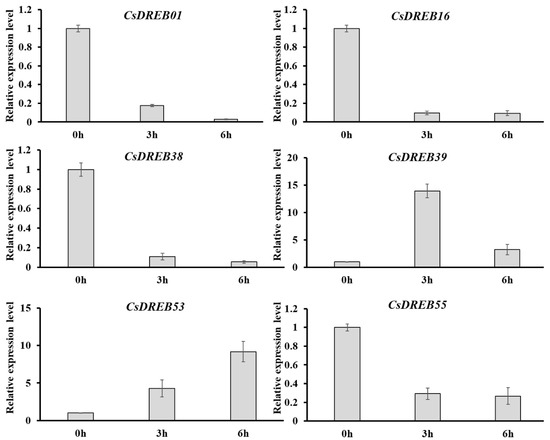
Figure 9.
The qRT-PCR analysis of CsDREB genes under heat stress: The seedlings were kept at 42 °C for heat stress, and leaf samples were collected at 0, 3, and 6 h. The value of each CsDREB gene at 0 h was normalized as ‘1.0’. The cucumber β-actin gene was used as an internal control, and three biological replicates were used for the gene expression analyses. Error bars represent the standard error (SE).
To explore the potential functions of CsDREBs in the resistance to biotic stresses, we performed expression analyses of CsDREBs using the published RNA-Seq data of cucumber seedlings inoculated with PM for 48 h and with DM for 8 days [46,61]. After inoculation with PM, a total of 22 genes were differentially expressed in the susceptible cucumber line compared with the control, and a total of 24 genes were differentially expressed in the resistant cucumber line compared with the control (Figure 10A). Among the differentially expressed genes affected by PM, 14 genes (CsDREB18, CsDREB26, CsDREB34, CsDREB03, CsDREB52, CsDREB45, CsDREB39, CsDREB10, CsDREB33, CsDREB14, CsDREB35, CsDREB51, CsDREB02, and CsDREB07) had similar expression trends between the susceptible and resistant lines, while three genes (CsDREB11, CsDREB49, and CsDREB15) had opposite expression trends. In the transcriptomic data from cucumber seedlings inoculated with DM, only 20 DREB genes were detected. Among these 20 genes, 15 (CsDREB36, CsDREB46, CsDREB39, CsDREB03, CsDREB18, CsDREB37, CsDREB38, CsDREB44, CsDREB53, CsDREB33, CsDREB52, CsDREB07, CsDREB51, CsDREB30, CsDREB12, and CsDREB35) were upregulated at a minimum of one treatment timepoint, while four genes (CsDREB47, CsDREB06, CsDREB09, and CsDREB10) were downregulated at a minimum of one treatment timepoint.
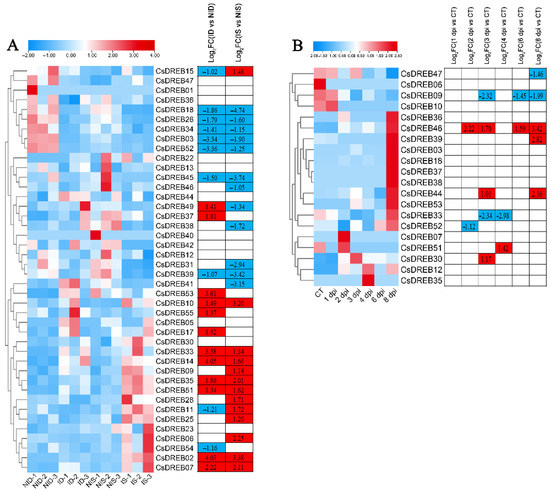
Figure 10.
Expression analysis of CsDREBs under biotic stresses: The transcriptional levels of CsDREB genes after infection with powdery mildew (PM) for 48 h (A) and with downy mildew (DM) for 1–8 days post-inoculation (B) are shown on the heatmaps. A range of −3.00 to 3.00 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. ID, PM-inoculated susceptible cucumber line D8 leaves; NID, non-inoculated D8 leaves; IS, PM-inoculated resistant cucumber line SSL508–28 leaves; NIS, non-inoculated SSL508–28 leaves; CT, without inoculation; DPI, days post inoculation; FC, fold-change.
4. Discussion
Plants respond to environmental stresses by activating specific genes during their growth and development; TFs play an indispensable role in this process. The dehydration-responsive element-binding factor (DREB) family is an important TF family for plants’ stress responses and has been identified in multiple species. Although the DREB family genes of cucumber have been identified previously [48], v1.0 of the ‘Chinese Long 9930‘ genome has been deleted because of its low quality, and the genome has been updated to v3 [62], resulting in the previously identified DREB gene names being unable to be found in the current genome database (http://www.cucurbitgenomics.org, accessed on 26 September 2022). The comprehensive identification and characterization of DREB genes in cucumber were hindered by low quality genomes and the lack of a high-quality pan-genome, and little is known about CsDREBs’ expression pattern under different stresses. At present, there is a pan-genome of cucumber available to use for genetic research. Therefore, we performed a pan-genome-wide identification of DREBs in the different cucumber accessions for the first time to further explore their variation and potential functions.
We identified a total of 55 CsDREB genes in the pan-genome of cucumber. We identified varying numbers of CsDREB genes in different cucumber accessions. Among the 13 cucumber accessions, the fewest genes were identified in ‘9110gt’ and ‘Cuc80’. Compared with ‘9930’, other accessions had similar numbers of CsDREB genes, lacking only one or two genes. Interestingly, we identified genes with the same sequence but different IDs in ‘9930’, ‘XTMC’, ‘Cuc64’, and ‘PI183967’—for example, CsaV3_4G026790.1/CsaV3_4G026910.1 and CSPI02G11740.1/CSPI02G18900.1. Only ‘9930’ and ‘XTMC’ had two copies of CsDREB30, and only ‘PI183967’ and ‘Cuc64’ had two copies of CsDREB09 (Table S2). For these cases, there might be two possible explanations: errors in genome assembly, or recent duplications of these two genes. Compared with gene numbers, the protein lengths showed greater variation, and there were only a few genes with consistent protein lengths in different accessions. However, we observed that the protein lengths in Indian accessions were more consistent—for example, CsDREB05, CsDREB10, CsDREB14, and CsDREB22 (Table 1 and Table 2). Moreover, the protein length of CsDREB41 in ‘PI183967’ was very short (Table 2), due to an early termination of translation caused by a base mutation. Strangely, the protein length of CsDREB50 in ‘W8’ was far longer than in other accessions (Table 2), and it was found that the protein sequence contained partial sequences of CsDREB50 from ‘9930’ and ‘XTMC’. Of course, the differences in protein length might also be due to the deviations in gene prediction and gene sequencing, and specific gene analysis will be needed to be certain.
To analyze the gene structure and potential function of DREBs more clearly, we mainly carried out follow-up analysis of the genes identified in v3 of the ‘Chinese Long 9930’ genome. We mapped 55 CsDREBs to cucumber chromosomes, and we numbered them CsDREB01–CsDREB55 according to their sequence on the seven chromosomes in this study (Figure S1). Based on the amino acid sequences, conserved structural domains, and phylogenetic relationships with A. thaliana, the 55 CsDREB proteins were classified into six groups: A1–A6 (Table 1 and Figure 2). The largest and smallest groups were A5 and A3, respectively; this was not similar to Arabidopsis, where the largest group was A4 [16]. Additionally, we observed that nine proteins (CsDREB01, 05, 12, 13, 25, 27, 28, 35, and 54) in cucumber group A5 were not closely related to any AtDREB and formed an independent branch by themselves (Figure 1). There were unique motifs in their protein structure: motif 6 in CsDREB01, 05, 25, and 27; and motif 9 in CsDREB12, 13, 28, 35, and 54 (Figure 3). This indicated that the members of group A5 had undergone expansion during their evolution in cucumber. According to Nakano et al. [15], groups A1–A6 could be cataloged into four clades (I–IV) according to the type of conserved domains. Previous studies have reported that the C-terminal regions of repressor-type ERF proteins have the EAR (ERF-associated amphiphilic repression) motif (DLNxxP), which can negatively regulate development, hormone, and stress signaling pathways, and plays a key role in various biological functions [63,64]. We found that the EAR motif existed only in clade II, and the regions of acidic amino-acid-rich, Gln-rich, Pro-rich, and/or Ser/Thr-rich amino acid sequences are believed usually to be designated as transcriptional activation domains [65]. We found Gln-rich motifs and Pro-rich motifs in clade III. Because proteins that share the same domains or motifs are likely to have similar functions, we speculated that most of the genes in clade II might be involved in negative regulation of the associated pathways, while most of the genes in clade III might be involved in transcriptional activation. In addition, most of the CsDREB genes did not contain introns; only CsDREB27 contained two introns, being consistent with the findings in Vitis vinifera [66], Ziziphus jujube [67], pineapple [42], and maize [38]. Additionally, all members of the independent branch of group A5 contained introns (Figure 3). Previous studies suggested that a small number of introns would help to respond quickly to various stresses and activate downregulated genes [56], and the presence of introns can lead to selective splicing, which can possibly delay the response to stresses.
It is well known that gene expression is correlated with gene function. To date, the DREB genes have been found to regulate multiple stresses, including drought [22,23], cold [24,68], heat [26,28], and salt [25] stresses, along with plant development [69,70,71,72,73]. For instance, in rice, the gene OsDREB1C (Dehydration-Responsive Element-Binding Protein 1C), a member of the DREB family, increased grain yield of 41.3 to 68.3% and, in addition, shortened growth duration, improved nitrogen use efficiency, and promoted efficient resource allocation, which provided an excellent strategy toward achieving much-needed increases in agricultural productivity through manipulating a single key gene [73]. In maize, ZmDREB2A played a dual functional role in mediating the expression of genes responsive to both drought stress and heat stress [26]. In this study, we analyzed the expression patterns of CsDREBs in cucumber in different tissues and under various stress conditions. Our results indicated that eight CsDREBs showed constitutive expression, and 12, 14, 11 and 16 CsDREBs showed specifically high expression in stem, tendril, male-flower, and root, respectively (Figure 7), suggesting that they might play important roles in the specific given tissues. Many genes showed varying degrees of expression under different stress conditions (Figure 8 and Figure 10). Although a total of 31, 20, 30, and 9 CsDREBs were differentially expressed in responding to the treatments of heat, NaCl and silicon, power mildew and downy mildew, respectively, only one gene, CsDREB33, responded to all the stresses tested, being up-regulated in the stresses of power mildew, heat and NaCl, but down-regulated in the downy mildew stress (Figure 8 and Figure 10). These results might give us some clues that it might be worthy to explore the function of CsDREB33 in multiple stress resistance of power mildew, heat and NaCl. In summary, the results of the present study showed that CsDREBs may play important roles in the growth and development of cucumber, and some of these genes participate in the stress response under one or more conditions, but there are also some CsDREB genes that do not participate in stress responses or even are not expressed, and it is possible that they work in other ways.
5. Conclusions
In this study, we performed pan-genome-wide identification of the DREB gene family in cucumber. In total, 55 members were identified and positioned on seven chromosomes. Four of the 13 examined accessions had all the 55 CsDREBs, and most of the CsDREB proteins had length and/or amino acid variations in different accessions. All the CsDREB genes were divided into six groups by analyzing their gene structures, conserved motifs, and relationships. Analysis of the expression patterns of CsDREBs in different tissues revealed that they are most likely involved in the growth and development of cucumber. Furthermore, transcriptomic data under different stress conditions revealed the differences and similarities in the stress-induced expression of CsDREBs in response to abiotic and biotic stresses, and CsDREB33 was found to response to all the four tested stresses. In conclusion, this study provides a reference for investigating the potential role of DREBs for breeding stress resistance in cucumber.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agriculture12111879/s1. Figure S1: Chromosome distribution of CsDREB genes; Figure S2: The different conserved motifs (CMs) of CsDREB in different groups; Table S1: The gene-specific primers used for qRT-PCR. Table S2: Identification of CsDREB genes in different cucumber varieties; Table S3: The variation of amino acids in different cucumber accessions. Table S4: Analysis and distribution of conserved motifs in cucumber DREB proteins; Table S5: Ka/Ks calculation and divergent time of the duplicated cucumber DREB gene pairs.
Author Contributions
Conceptualization, Z.R., L.W. and C.C.; methodology, C.W. and J.H.; software, C.W., J.H., T.W., J.L., Z.X. and Q.Z.; data analysis, C.W. and J.H.; investigation, C.W. and J.H.; data curation, C.W. and J.H.; writing—original draft preparation, C.W. and J.H.; writing—review and editing, Z.R.; supervision, Z.R.; project administration, Z.R., L.W. and C.C.; funding acquisition, Z.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31872950), the Shandong “Double Tops” Program (SYL2017YSTD06) and the ‘Taishan Scholar’ Foundation of the People’s Government of Shandong Province (ts20130932).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and Supplementary Materials.
Acknowledgments
We extend our appreciation to the anonymous reviewers for their valuable suggestions to help improve this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| AP2/ERF | APETALA2/ethylene response factor |
| CDS | Coding sequence |
| Chr | Chromosome |
| CM | Conserved motif |
| CT | Control treatment |
| DM | Downy mildew |
| DPI | Days post-inoculation |
| DREB | Dehydration-responsive element-binding factor |
| FC | Fold-change |
| FPKM | Fragments per kilobase of exon model per million mapped fragments |
| Glu | Glutamine |
| HMM | Hidden Markov model |
| Kb | Kilobase |
| mRNA | Messenger RNA |
| NCBI | National Center for Biotechnology Information |
| PM | Powdery mildew |
| UTR | Untranslated region |
| Val | Valine |
References
- Ohama, N.; Kusakabe, K.; Mizoi, J.; Zhao, H.; Kidokoro, S.; Koizumi, S.; Takahashi, F.; Ishida, T.; Yanagisawa, S.; Shinozaki, K.; et al. The Transcriptional Cascade in the Heat Stress Response of Arabidopsis Is Strictly Regulated at the Level of Transcription Factor Expression. Plant Cell 2016, 28, 181–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; Weisskopf, L. Airborne medicine: Bacterial volatiles and their influence on plant health. New Phytol. 2020, 226, 32–43. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Plant Immune System: Crosstalk between Responses to Biotic and Abiotic Stresses the Missing Link in Understanding Plant Defence. Curr. Issues Mol. Biol. 2017, 23, 1–15. [Google Scholar] [CrossRef]
- Paes de Melo, B.; Carpinetti, P.A.; Fraga, O.T.; Rodrigues-Silva, P.L.; Fioresi, V.S.; de Camargos, L.F.; Ferreira, M. Abiotic Stresses in Plants and Their Markers: A Practice View of Plant Stress Responses and Programmed Cell Death Mechanisms. Plants 2022, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Lai, W.; Zhu, C.; Hu, Z.; Liu, S.; Wu, H.; Zhou, Y. Identification and Transcriptional Analysis of Zinc Finger-Homeodomain (ZF-HD) Family Genes in Cucumber. Biochem. Genet. 2021, 59, 884–901. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Ratcliffe, O.J. A genomic perspective on plant transcription factors. Curr. Opin. Plant Biol. 2000, 3, 423–434. [Google Scholar] [CrossRef]
- Wang, G.D.; Xu, X.P.; Wang, H.; Liu, Q.; Yang, X.T.; Liao, L.X.; Cai, G.H. A tomato transcription factor, SlDREB3 enhances the tolerance to chilling in transgenic tomato. Plant Physiol. Biochem. 2019, 142, 254–262. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, C.L.; Wang, G.L.; Wang, Y.X.; Qi, C.H.; You, C.X.; Li, Y.Y.; Hao, Y.J. Apple AP2/EREBP transcription factor MdSHINE2 confers drought resistance by regulating wax biosynthesis. Planta 2019, 249, 1627–1643. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional regulation of bHLH during plant response to stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Deyholos, M.K. Comprehensive transcriptional profiling of NaCl-Stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol. 2006, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-Wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genomics. 2012, 287, 495–513. [Google Scholar] [CrossRef]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-Wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef]
- Cao, Z.F.; Li, J.; Chen, F.; Li, Y.Q.; Zhou, H.M.; Liu, Q. Effect of two conserved amino acid residues on DREB1A function. Biochemistry 2001, 66, 623–627. [Google Scholar] [CrossRef]
- Liu, Z.; Yuan, G.; Liu, S.; Jia, J.; Cheng, L.; Qi, D.; Shen, S.; Peng, X.; Liu, G. Identified of a novel cis-element regulating the alternative splicing of LcDREB2. Sci. Rep. 2017, 7, 46106. [Google Scholar] [CrossRef]
- Lakhwani, D.; Pandey, A.; Dhar, Y.V.; Bag, S.K.; Trivedi, P.K.; Asif, M.H. Genome-wide analysis of the AP2/ERF family in Musa species reveals divergence and neofunctionalisation during evolution. Sci. Rep. 2016, 6, 18878. [Google Scholar] [CrossRef]
- Wang, L.; Ma, H.; Lin, J. Angiosperm-Wide and Family-Level Analyses of AP2/ERF Genes Reveal Differential Retention and Sequence Divergence after Whole-Genome Duplication. Front. Plant Sci. 2019, 10, 196. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.L.; Li, Y.N.; Zhang, X.P.; Song, Y.; Wang, W.; Fang, J.; Cui, W.M.; Jia, X.D. Immunotoxicologic assessment of genetically modified drought-resistant wheat T349 with GmDREB1. Zhonghua Yu Fang Yi Xue Za Zhi 2012, 46, 556–560. [Google Scholar] [PubMed]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef]
- Hong, J.P.; Kim, W.T. Isolation and functional characterization of the Ca-DREBLP1 gene encoding a dehydration-responsive element binding-factor-like protein 1 in hot pepper (Capsicum annuum L. cv. Pukang). Planta 2005, 220, 875–888. [Google Scholar] [CrossRef]
- Qin, F.; Kakimoto, M.; Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J. 2007, 50, 54–69. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Liu, Y.; Gao, H.; Wang, Z.; Sun, G. CkDREB gene in Caragana korshinskii is involved in the regulation of stress response to multiple abiotic stresses as an AP2/EREBP transcription factor. Mol. Biol. Rep. 2011, 38, 2801–2811. [Google Scholar] [CrossRef]
- Hong, B.; Ma, C.; Yang, Y.; Wang, T.; Yamaguchi-Shinozaki, K.; Gao, J. Over-expression of AtDREB1A in chrysanthemum enhances tolerance to heat stress. Plant Mol. Biol. 2009, 70, 231–240. [Google Scholar] [CrossRef]
- Hwang, J.E.; Lim, C.J.; Chen, H.; Je, J.; Song, C.; Lim, C.O. Overexpression of Arabidopsis dehydration- responsive element-binding protein 2C confers tolerance to oxidative stress. Mol. Cells 2012, 33, 135–140. [Google Scholar] [CrossRef]
- Sharoni, A.M.; Nuruzzaman, M.; Satoh, K.; Shimizu, T.; Kondoh, H.; Sasaya, T.; Choi, I.R.; Omura, T.; Kikuchi, S. Gene structures, classification and expression models of the AP2/EREBP transcription factor family in rice. Plant Cell Physiol. 2011, 52, 344–360. [Google Scholar] [CrossRef]
- Zhou, Y.X.; Zhou, W.; Liu, H.; Liu, P.; Li, Z.G. Genome-Wide analysis of the soybean DREB gene family: Identification, genomic organization and expression profiles in response to drought stress. Plant Breed. 2020, 139, 1158–1167. [Google Scholar] [CrossRef]
- Agarwal, P.; Agarwal, P.K.; Nair, S.; Sopory, S.K.; Reddy, M.K. Stress-Inducible DREB2A transcription factor from Pennisetum glaucum is a phosphoprotein and its phosphorylation negatively regulates its DNA-binding activity. Mol. Genet. Genomics 2007, 277, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Durmaz, E.; Akpınar, B.A.; Budak, H. The drought response displayed by a DRE-binding protein from Triticum dicoccoides. Plant Physiol. Biochem. 2011, 49, 346–351. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Wang, H.; Xin, H.; Yang, X.; Yan, J.; Li, J.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; et al. Genome-Wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. PLoS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Zhu, K.; Liu, L.; Chen, F.; Yu, D. Identification and characterization of two chrysanthemum (Dendronthema x moriforlium) DREB genes, belonging to the AP2/EREBP family. Mol. Biol. Rep. 2009, 36, 71–81. [Google Scholar] [CrossRef]
- Yang, H.; Sun, Y.; Wang, H.; Zhao, T.; Xu, X.; Jiang, J.; Li, J. Genome-Wide identification and functional analysis of the ERF2 gene family in response to disease resistance against Stemphylium lycopersici in tomato. BMC Plant Biol. 2021, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Shi, A.; Mou, B. Genome-wide identification and expression analysis of the CBF/DREB1 gene family in lettuce. Sci. Rep. 2020, 10, 5733. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Jia, J.P.; Kong, D.C.; Zhang, Z.D.; Song, S.; Li, Y.Y.; Pang, X.M. Genome-wide identification and analysis of the DREB genes and their expression profiles under abiotic stresses in Chinese jujube (Ziziphus jujuba Mill.). J. For. Res. 2019, 30, 1277–1287. [Google Scholar] [CrossRef]
- Liu, X.Q.; Zhu, J.J.; Wei, C.J.; Guo, Q.; Bian, C.K.; Xiang, Z.H.; Zhao, A.C. Genome-wide identification and characterization of the DREB transcription factor gene family in mulberry. Biol. Plant. 2015, 59, 253–265. [Google Scholar] [CrossRef]
- Huang, X.; Song, X.P.; Chen, R.F.; Zhang, B.Q.; Li, C.N.; Liang, Y.S.; Qiu, L.H.; Fan, Y.G.; Zhou, Z.F.; Zhou, H.W.; et al. Genome-Wide Analysis of the DREB Subfamily in Saccharum spontaneum Reveals Their Functional Divergence During Cold and Drought Stresses. Front. Genet. 2020, 10, 1326. [Google Scholar] [CrossRef]
- Li, Z.; Wang, G.; Liu, X.H.; Wang, Z.C.; Zhang, M.Q.; Zhang, J.S. Genome-Wide identification and expression profiling of DREB genes in Saccharum spontaneum. BMC Genom. 2021, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chai, M.N.; Cheng, H.; Yan, M.K.; Priyadarshani, S.; Zhang, M.; He, Q.; Huang, Y.M.; Chen, F.Q.; Liu, L.P.; Huang, X.Y.; et al. Identification and expression analysis of the DREB transcription factor family in pineapple (Ananas comosus (L.) Merr.). PeerJ 2020, 8, e9006. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, N.; Munir, F.; Gul, A.; Amir, R.; Paracha, R.Z. Genome-Wide analysis, identification, evolution and genomic organization of dehydration responsive element-binding (DREB) gene family in Solanum tuberosum. PeerJ 2021, 9, e11647. [Google Scholar] [CrossRef]
- Liu, M.Y.; Zhang, C.J.; Duan, L.X.; Luan, Q.Q.; Li, J.L.; Yang, A.G.; Qi, X.Q.; Ren, Z.H. CsMYB60 is a key regulator of flavonols and proanthocyanidans that determine the colour of fruit spines in cucumber. J. Exp. Bot. 2019, 70, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Luan, Q.Q.; Chen, C.H.; Liu, M.Y.; Li, Q.; Wang, L.N.; Ren, Z. CsWRKY50 mediates defense responses to Pseudoperonospora cubensis infection in Cucumis sativus. Plant Sci. 2019, 279, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.N.; Savory, E.A.; Vaillancourt, B.; Childs, K.L.; Hamilton, J.P.; Day, B.; Buell, C.R. Expression Profiling of Cucumis sativus in Response to Infection by Pseudoperonospora cubensis. PLoS ONE 2012, 7, e34954. [Google Scholar] [CrossRef]
- Chen, C.H.; Chen, X.Q.; Han, J.; Lu, W.L.; Ren, Z.H. Genome-wide analysis of theWRKYgene family in the cucumber genome and transcriptome-wide identification of WRKY transcription factors that respond to biotic and abiotic stresses. BMC Plant Biol. 2020, 20, 1–19. [Google Scholar] [CrossRef]
- Hu, L.; Liu, S. Genome-Wide identification and phylogenetic analysis of the ERF gene family in cucumbers. Genet. Mol. Biol. 2011, 34, 624–633. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef]
- Bayer, P.E.; Golicz, A.A.; Scheben, A.; Batley, J.; Edwards, D. Plant pan-genomes are the new reference. Nat. Plants 2020, 6, 914–920. [Google Scholar] [CrossRef]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, S.; Chai, S.; Yang, Z.; Zhang, Q.; Xin, H.; Xu, Y.; Lin, S.; Chen, X.; Yao, Z.; et al. Graph-Based pan-genome reveals structural and sequence variations related to agronomic traits and domestication in cucumber. Nat. Commun. 2022, 13, 682. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; Yan, P.; Huang, S.; Fei, Z.; Lin, K. RNA-Seq improves annotation of protein-coding genes in the cucumber genome. BMC Genom. 2011, 12, 540. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, J.; Liang, Y.; Liu, J.; Jia, J.; Huo, H.; Wu, Z.; Yang, R.; Gong, H. Transcriptomic dynamics provide an insight into the mechanism for silicon-mediated alleviation of salt stress in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 174, 245–254. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Tang, R.; Wang, L.; Chen, C.; Ren, Z. Genome-Wide identification and expression analysis of Hsf and Hsp gene families in cucumber (Cucumis sativus L.). Plant Growth Regul. 2021, 95, 223–239. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Shi, Y.; Qi, X.; Chen, X. Elucidation of the molecular responses of a cucumber segment substitution line carrying Pm5.1 and its recurrent parent triggered by powdery mildew by comparative transcriptome profiling. BMC Genom. 2017, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, H.; Huang, W.; Xu, Y.; Zhou, Q.; Wang, S.; Ruan, J.; Huang, S.; Zhang, Z. A chromosome-scale genome assembly of cucumber (Cucumis sativus L.). GigaScience 2019, 8, giz072. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 2000, 12, 393–404. [Google Scholar] [CrossRef]
- Ohta, M.; Matsui, K.; Hiratsu, K.; Shinshi, H.; Ohme-Takagi, M. Repression domains of class II ERF transcriptional repressors share an essential motif for active repression. Plant Cell 2001, 13, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; White, M.J.; MacRae, T.H. Transcription factors and their genes in higher plants functional domains, evolution and regulation. Eur. J. Biochem. 1999, 262, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Xia, H.; Liu, J.; Ma, F. The gene family of dehydration responsive element-binding transcription factors in grape (Vitis vinifera): Genome-Wide identification and analysis, expression profiles, and involvement in abiotic stress resistance. Mol. Biol. Rep. 2014, 41, 1577–1590. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X. Genome-wide identification of AP2/ERF superfamily genes and their expression during fruit ripening of Chinese jujube. Sci. Rep. 2018, 8, 15612. [Google Scholar] [CrossRef]
- Cao, P.B.; Azar, S.; SanClemente, H.; Mounet, F.; Dunand, C.; Marque, G.; Marque, C.; Teulières, C. Genome-wide analysis of the AP2/ERF family in Eucalyptus grandis: An intriguing over-representation of stress-responsive DREB1/CBF genes. PLoS ONE 2015, 10, e0121041. [Google Scholar] [CrossRef]
- Gilmour, S.J.; Zarka, D.G.; Stockinger, E.J.; Salazar, M.P.; Houghton, J.M.; Thomashow, M.F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J 1998, 16, 433–442. [Google Scholar] [CrossRef]
- Knight, H.; Zarka, D.G.; Okamoto, H.; Thomashow, M.F.; Knight, M.R. Abscisic acid induces CBF gene transcription and subsequent induction of cold-regulated genes via the CRT promoter element. Plant Physiol. 2004, 135, 1710–1717. [Google Scholar] [CrossRef]
- Xiao, H.; Tattersall, E.A.R.; Siddiqua, M.K.; Cramer, G.R.; Nassuth, A. CBF4 is a unique member of the CBF transcription factor family of Vitis vinifera and Vitis riparia. Plant Cell Environ. 2008, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Han, J.; Deng, X.; Tan, S.; Li, L.; Li, L.; Zhou, J.; Peng, H.; Yang, G.; He, G.; et al. Expansion and stress responses of AP2/EREBP superfamily in Brachypodium distachyon. Sci. Rep. 2016, 6, 21623. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Li, X.; Lu, Z.; Zhang, H.; Ye, X.; Zhou, Y.; Li, J.; Yan, Y.; Pei, H.; Duan, F.; et al. A transcriptional regulator that boosts grain yields and shortens the growth duration of rice. Science 2022, 377, eabi8455. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).