
Temporal and Spatial Positioning of Service Crops in Cereals Affects Yield and Weed Control
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
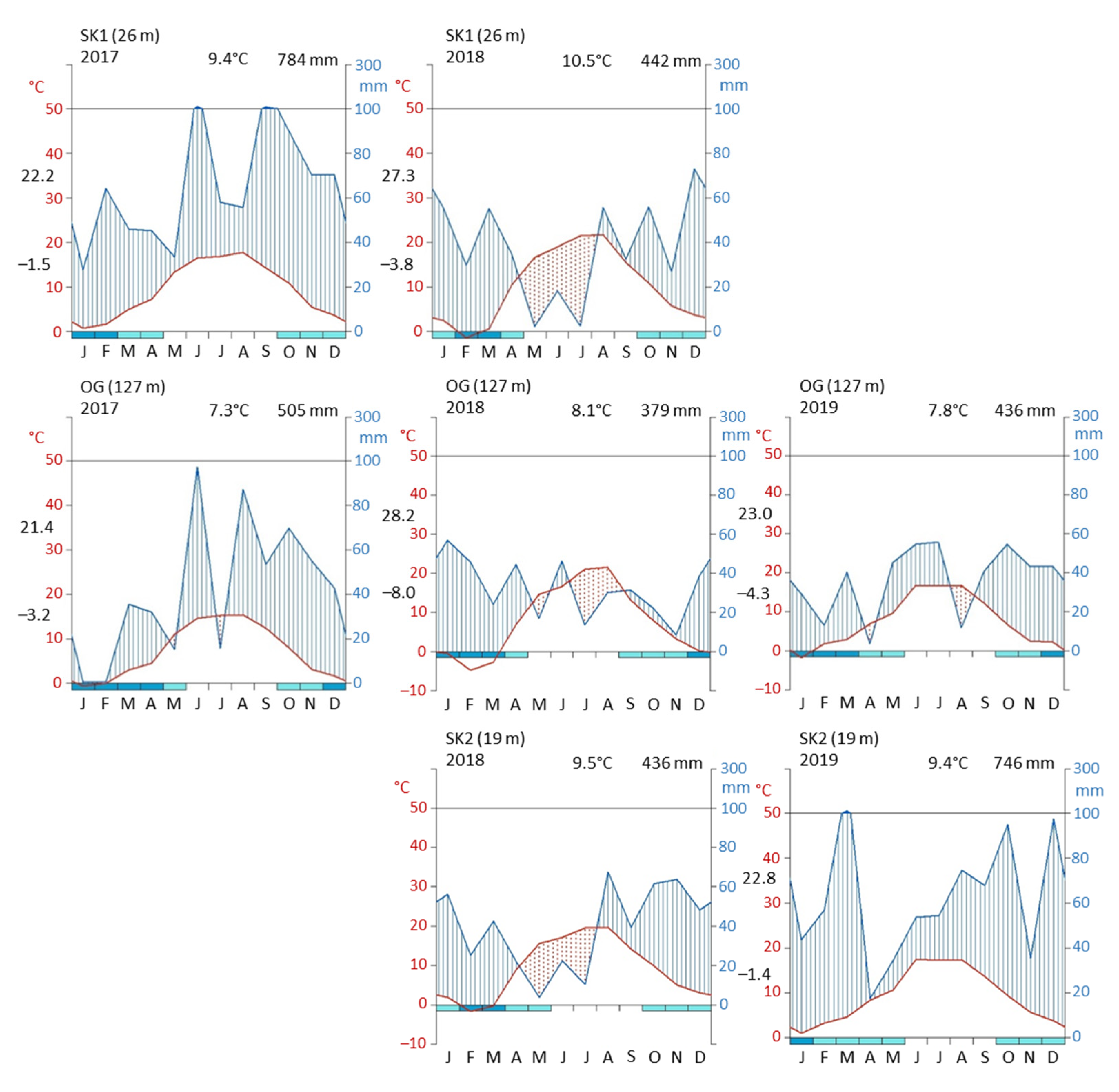
2.1. Experimental Sites
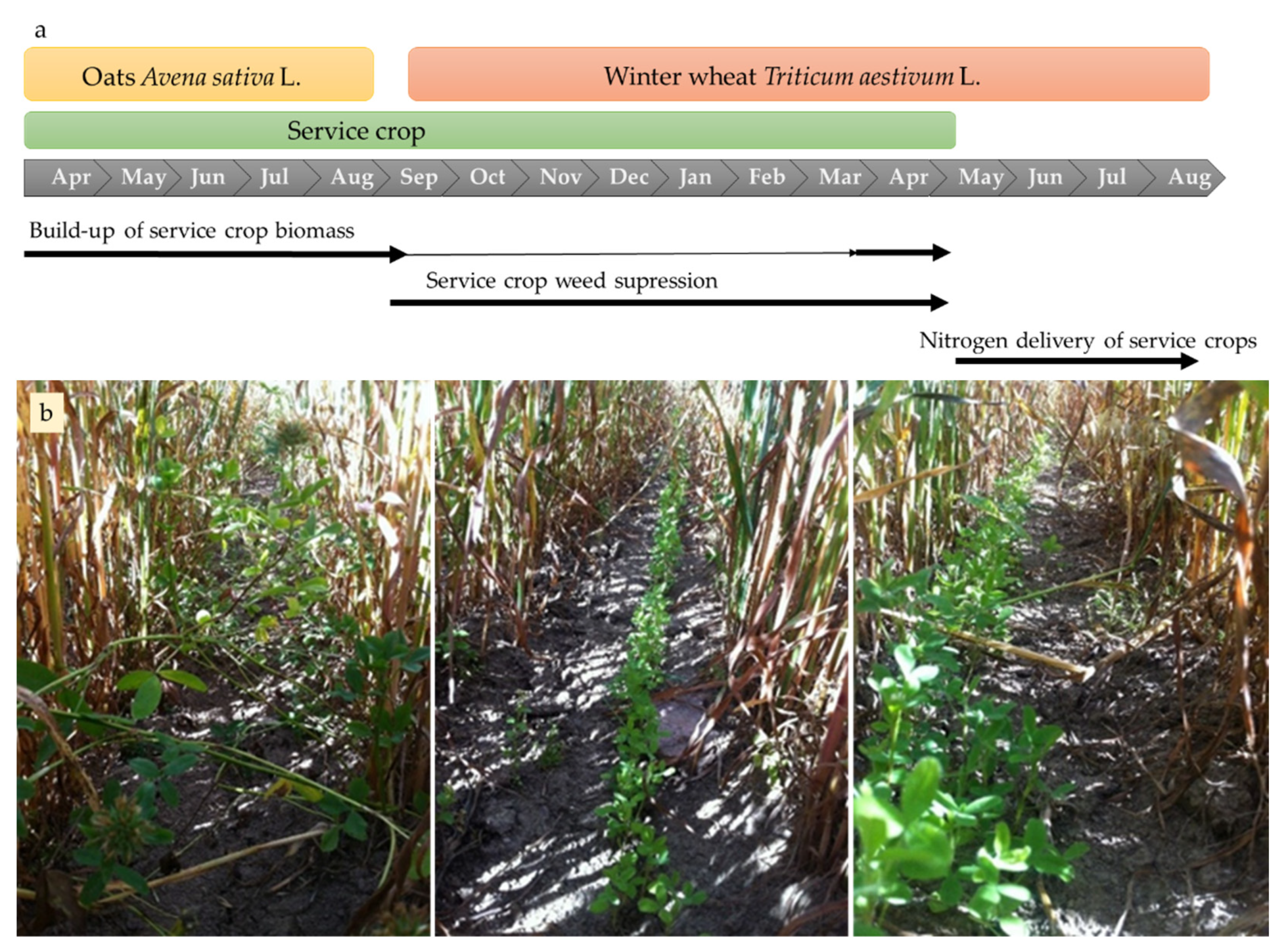
2.2. Crop Sequence and Management
2.3. Experimental Design
2.3.1. Service Crop Mixtures
2.3.2. System Strategy
2.4. Data Collection
2.4.1. Service Crop Performance
Nitrogen Isotope Analysis
2.4.2. Cash Crop Performance
2.4.3. Weed Biomass Assessment
2.4.4. Soil Mineral Nitrogen
2.5. Statistical Analysis
3. Results
3.1. Effect of Local Conditions on the System
3.2. Service Crop Performance
3.2.1. Establishment and Productivity of Service Crop Mixtures
3.2.2. Service Crop Soil Cover in Late Autumn
3.2.3. Service Crop Dinitrogen Fixation
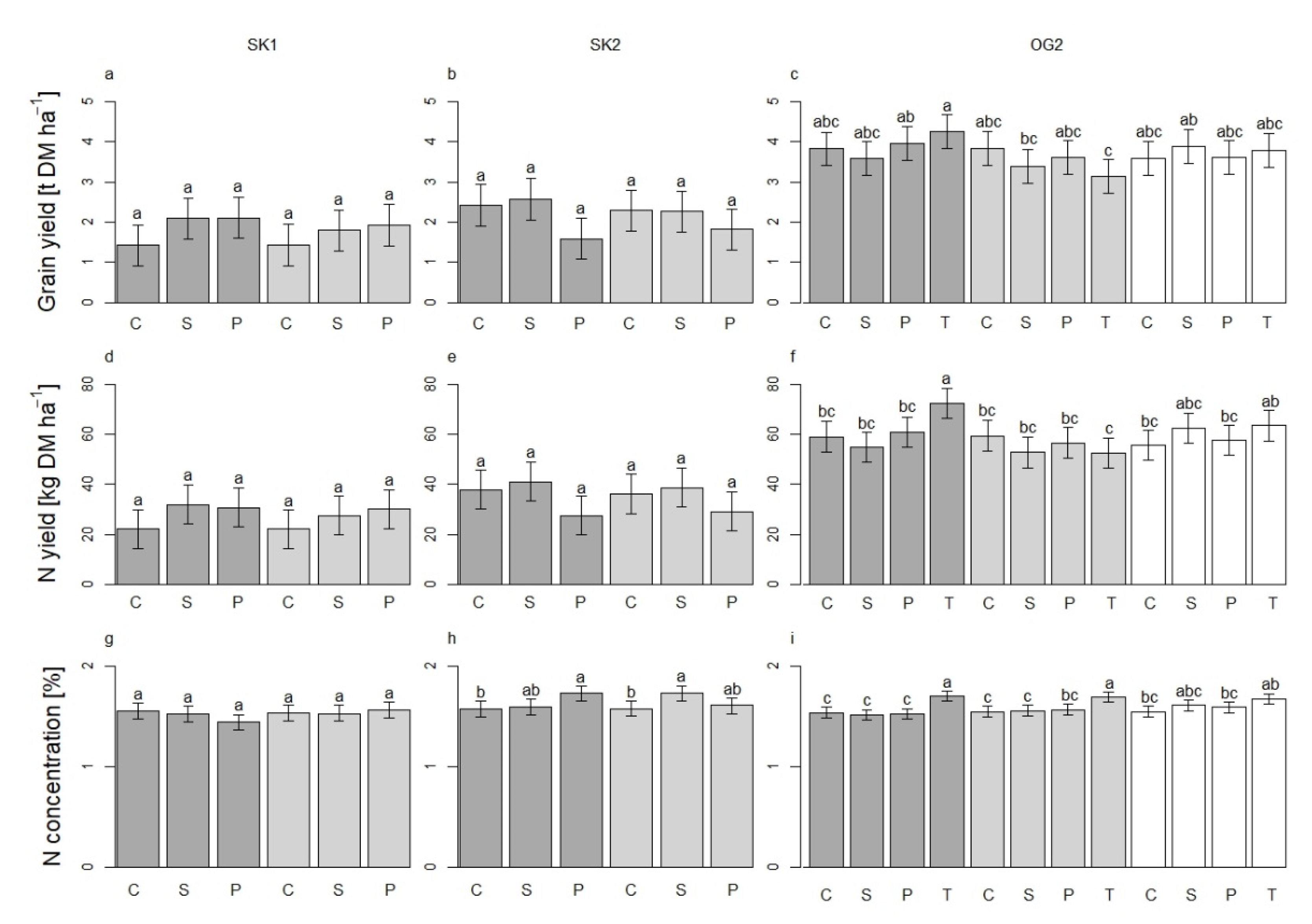
3.3. Delivery of Services and Disservices by Service Crops to the System
Effects of Service Crops on Winter Wheat Grain Yield and Nitrogen Content in Kernels
3.4. Oat Yields
3.5. Weed Biomass
3.6. Soil Mineral Nitrogen
4. Discussion
4.1. Nitrogen Provision for Yield and Grain Nitrogen Content in Winter Wheat
4.2. Weed Suppression
4.3. Oats–Service Crop Interaction
4.4. Service Crop Performance and Species Choice
4.5. Developing (Relay) Intercropping Systems with Forage Legumes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kremen, C. Managing Ecosystem Services: What Do We Need to Know about Their Ecology? Ecol. Lett. 2005, 8, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Nivelle, E.; Verzeaux, J.; Habbib, H.; Kuzyakov, Y.; Decocq, G.; Roger, D.; Lacoux, J.; Duclercq, J.; Spicher, F.; Nava-Saucedo, J.E.; et al. Functional Response of Soil Microbial Communities to Tillage, Cover Crops and Nitrogen Fertilization. Appl. Soil Ecol. 2016, 108, 147–155. [Google Scholar] [CrossRef]
- Verret, V.; Gardarin, A.; Pelzer, E.; Médiène, S.; Makowski, D.; Valantin-Morison, M. Can Legume Companion Plants Control Weeds without Decreasing Crop Yield? A Meta-Analysis. F. Crop. Res. 2017, 204, 158–168. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Holman, J.D.; Schlegel, A.J.; Tatarko, J.; Shaver, T.M. Replacing Fallow with Cover Crops in a Semiarid Soil: Effects on Soil Properties. Soil Sci. Soc. Am. J. 2013, 77, 1026–1034. [Google Scholar] [CrossRef]
- Askegaard, M.; Eriksen, J. Residual Effect and Leaching of N and K in Cropping Systems with Clover and Ryegrass Catch Crops on a Coarse Sand. Agric. Ecosyst. Environ. 2008, 123, 99–108. [Google Scholar] [CrossRef]
- Liu, J.; Bergkvist, G.; Ulén, B. Biomass Production and Phosphorus Retention by Catch Crops on Clayey Soils in Southern and Central Sweden. F. Crop. Res. 2015, 171, 130–137. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon Sequestration in Agricultural Soils via Cultivation of Cover Crops—A Meta-Analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Büchi, L.; Gebhard, C.A.; Liebisch, F.; Sinaj, S.; Ramseier, H.; Charles, R. Accumulation of Biologically Fixed Nitrogen by Legumes Cultivated as Cover Crops in Switzerland. Plant Soil 2015, 393, 163–175. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amossé, C.; Sinaj, S.; Walter, A.; Charles, R. Influence of Root and Leaf Traits on the Uptake of Nutrients in Cover Crops. Plant Soil 2016, 409, 419–434. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Abbate, C.; Lombardo, S.; Fontanazza, S.; Pandino, G.; Anastasi, U.; Mauromicale, G. Trifolium Subterraneum Cover Cropping Enhances Soil Fertility and Weed Seedbank Dynamics in a Mediterranean Apricot Orchard. Agron. Sustain. Dev. 2021, 41, 70. [Google Scholar] [CrossRef]
- Verret, V.; Pelzer, E.; Bedoussac, L.; Jeuffroy, M.H. Tracking on-Farm Innovative Practices to Support Crop Mixture Design: The Case of Annual Mixtures Including a Legume Crop. Eur. J. Agron. 2020, 115, 126018. [Google Scholar] [CrossRef]
- Finney, D.M.; White, C.M.; Kaye, J.P. Biomass Production and Carbon/Nitrogen Ratio Influence Ecosystem Services from Cover Crop Mixtures. Agron. J. 2016, 108, 39–52. [Google Scholar] [CrossRef]
- Bergkvist, G.; Stenberg, M.; Wetterlind, J.; Båth, B.; Elfstrand, S. Clover Cover Crops Under-Sown in Winter Wheat Increase Yield of Subsequent Spring Barley-Effect of N Dose and Companion Grass. F. Crop. Res. 2011, 120, 292–298. [Google Scholar] [CrossRef]
- Zhao, J.; De Notaris, C.; Olesen, J.E. Autumn-Based Vegetation Indices for Estimating Nitrate Leaching during Autumn and Winter in Arable Cropping Systems. Agric. Ecosyst. Environ. 2020, 290, 106786. [Google Scholar] [CrossRef]
- Aronsson, H.; Hansen, E.M.; Thomsen, I.K.; Liu, J.; Øgaard, A.F.; Känkänen, H.; Ulén, B. The Ability of Cover Crops to Reduce Nitrogen and Phosphorus Losses from Arable Land in Southern Scandinavia and Finland. J. Soil Water Conserv. 2016, 71, 41–55. [Google Scholar] [CrossRef]
- Reimer, M.; Ringselle, B.; Bergkvist, G.; Westaway, S.; Wittwer, R.; Baresel, J.P.; Van Der Heijden, M.G.A.; Mangerud, K.; Finckh, M.R.; Brandsæter, L.O. Interactive Effects of Subsidiary Crops and Weed Pressure in the Transition Period to Non-Inversion Tillage, a Case Study of Six Sites across Northern and Central Europe. Agronomy 2019, 9, 495. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Meersmans, J.; Serlet, L. Cover Crops and Their Erosion-Reducing Effects during Concentrated Flow Erosion. Catena 2011, 85, 237–244. [Google Scholar] [CrossRef]
- Justes, E.; Bedoussac, L.; Dordas, C.; Frak, E.; Louarn, G.; Boudsocq, S.; Journet, E.P.; Lithourgidis, A.; Pankou, C.; Zhang, C.; et al. The 4 C Approach as a Way to Understand Species Interactions Determining Productivity. Front. Agric. Sci. Eng. 2021, 8, 387–399. [Google Scholar] [CrossRef]
- Blackshaw, R.E.; Moyer, J.R.; Doram, R.C.; Boswall, A.L.; Smith, E.G. Suitability of Undersown Sweetclover as a Fallow Replacement in Semiarid Cropping Systems. Agron. J. 2001, 93, 863–868. [Google Scholar] [CrossRef]
- Walter, H.; Lieth, H.H.F. Klimadiagramm-Weltatlas; Jena, G., Ed.; Fischer Verlag: Jena, Germany, 1967. [Google Scholar]
- Werner, R.A.; Bruch, B.A.; Brand, W.A. ConFlo III—An Interface for High Precision d 13 C and d 15 N Analysis with an Extended Dynamic Range. Rapid Commun. Mass Spectrom. 1999, 13, 1237–1241. [Google Scholar] [CrossRef]
- Shearer, G.; Kohl, D.H. N2-Fixation in Field Settings: Estimations Based on Natural 15N Abundance. Aust. J. Plant Physiol. 1986, 13, 699–756. [Google Scholar] [CrossRef]
- Roscher, C.; Thein, S.; Weigelt, A.; Temperton, V.M.; Buchmann, N.; Schulze, E.; Weigelt, A. N2 Fixation and Performance of 12 Legume Species in a 6-Year Grassland Biodiversity Experiment. Plant Soil 2011, 341, 333–348. [Google Scholar] [CrossRef]
- Eriksen, J.; Høgh-Jensen, H. Variations in the Natural Abundance of 15 N in Ryegrass/White Clover Shoot Material as Influenced by Cattle Grazing. Plant Soil 1998, 205, 67–76. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 4 January 2021).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Package ‘Lme4’ Linear Mixed-Effects Models Using “Eigen” and S4. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 2 August 2022).
- Fox, J.; Weisberg, S.; Price, B.; Adler, D.; Bates, D.; Baud-Bovy, G.; Bolker, B.; Ellison, S.; Firth, D.; Friendly, M.; et al. Car: Companion to Applied Regression. Available online: https://cran.r-project.org/web/packages/car/index.html (accessed on 2 August 2022).
- Russell, A.; Lenth, V.; Buerkner, P.; Herve, M.; Love, J.; Singmann, H.; Lenth, M.R. V Package ‘ Emmeans ’ R Topics Documented. 2022. Available online: https://cran.r-project.org/web/packages/emmeans/ (accessed on 2 August 2022).
- Wickham, H. Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Askegaard, M.; Eriksen, J. Growth of Legume and Nonlegume Catch Crops and Residual-N Effects in Spring Barley on Coarse Sand. J. Plant Nutr. Soil Sci. 2007, 170, 773–780. [Google Scholar] [CrossRef]
- Noland, R.L.; Wells, M.S.; Sheaffer, C.C.; Baker, J.M.; Martinson, K.L.; Coulter, J.A. Establishment and Function of Cover Crops Interseeded into Corn. Crop Sci. 2018, 58, 863–873. [Google Scholar] [CrossRef]
- Arlauskienė, A.; Gecaitė, V.; Toleikienė, M.; Šarūnaitė, L.; Kadžiulienė, Ž. Soil Nitrate Nitrogen Content and Grain Yields of Organically Grown Cereals as Affected by a Strip Tillage and Forage Legume Intercropping. Plants 2021, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Thorup-Kristensen, K. The Effect of Nitrogen Catch Crops on The Nitrogen Nutrition of A Succeeding Crop I. Effects Through Mineralization and Pre-Emptive Competition. Acta Agric. Scand. Sect. B Soil Plant Sci. 1993, 43, 74–81. [Google Scholar] [CrossRef]
- Wivstad, M. Nitrogen Mineralization and Crop Uptake of N from Decomposing 15N Labelled Red Clover and Yellow Sweetclover Plant Fractions of Different Age. Plant Soil 1999, 208, 21–31. [Google Scholar] [CrossRef]
- Magid, J.; Henriksen, O.; Thorup-kristensen, K.; Mueller, T. Disproportionately High N-Mineralisation Rates from Green Manures at Low Temperatures—Implications for Modeling and Management in Cool Temperate Agro-Ecosystems. Plant Soil 2001, 228, 73–82. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Dresbøll, D.B. Incorporation Time of Nitrogen Catch Crops Influences the N Effect for the Succeeding Crop. Soil Use Manag. 2010, 26, 27–35. [Google Scholar] [CrossRef]
- Tosti, G.; Farneselli, M.; Benincasa, P.; Guiducci, M. Nitrogen Fertilization Strategies for Organic Wheat Production: Crop Yield and Nitrate Leaching. Agron. J. 2016, 108, 770–781. [Google Scholar] [CrossRef]
- Rochette, P.; Liang, C.; Pelster, D.; Bergeron, O.; Lemke, R.; Kroebel, R.; MacDonald, D.; Yan, W.; Flemming, C. Soil Nitrous Oxide Emissions from Agricultural Soils in Canada: Exploring Relationships with Soil, Crop and Climatic Variables. Agric. Ecosyst. Environ. 2018, 254, 69–81. [Google Scholar] [CrossRef]
- Hamza, M.A.; Anderson, W.K. Soil Compaction in Cropping Systems: A Review of the Nature, Causes and Possible Solutions. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Shen, Y.; McLaughlin, N.; Zhang, X.; Xu, M.; Liang, A. Effect of Tillage and Crop Residue on Soil Temperature Following Planting for a Black Soil in Northeast China. Sci. Rep. 2018, 8, 4500. [Google Scholar] [CrossRef]
- Dorn, B.; Jossi, W.; van der Heijden, M.G.A. Weed Suppression by Cover Crops: Comparative on-Farm Experiments under Integrated and Organic Conservation Tillage. Weed Res. 2015, 55, 586–597. [Google Scholar] [CrossRef]
- Cirujeda, A.; Melander, B.; Rasmussen, K.; Rasmussen, I.A. Relationship between Speed, Soil Movement into the Cereal Row and Intra-Row Weed Control Efficacy by Weed Harrowing. Weed Res. 2003, 43, 285–296. [Google Scholar] [CrossRef]
- Kurstjens, D.A.G.; Kropff, M.J. The Impact of Uprooting and Soil-Covering on the Effectiveness of Weed Harrowing. Weed Res. 2001, 41, 211–288. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Christiansen, J.L.; Rasmussen, J. Weed Harrowing and Inter-Row Hoeing in Organic Grown Quinoa (Chenopodium Quinoa Willd.). Outlook Agric. 2010, 39, 223–227. [Google Scholar] [CrossRef]
- Kolb, L.N.; Gallandt, E.R.; Molloy, T. Improving Weed Management in Organic Spring Barley: Physical Weed Control vs. Interspecific Competition. Weed Res. 2010, 50, 597–605. [Google Scholar] [CrossRef]
- Melander, B.; Jabran, K.; De Notaris, C.; Znova, L.; Green, O.; Olesen, J.E. Inter-Row Hoeing for Weed Control in Organic Spring Cereals—Influence of Inter-Row Spacing and Nitrogen Rate. Eur. J. Agron. 2018, 101, 49–56. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. A Functional Analysis of Large-Scale Temporal Shifts from 1970 to 2000 in Weed Assemblages of Sunflower Crops in France. J. Veg. Sci. 2009, 20, 49–58. [Google Scholar] [CrossRef]
- Storkey, J.; Moss, S.R.; Cussans, J.W. Using Assembly Theory to Explain Changes in a Weed Flora in Response to Agricultural Intensification. Weed Sci. 2010, 58, 39–46. [Google Scholar] [CrossRef]
- Amosse, C.; Mary, B.; David, C. Contribution of Relay Intercropping with Legume Cover Crops on Nitrogen Dynamics in Organic Grain Systems. Nutr. Cycl. Agroecosystems 2014, 98, 1–14. [Google Scholar] [CrossRef]
- Vrignon-Brenas, S.; Celette, F.; Amossé, C.; David, C. Effect of Spring Fertilization on Ecosystem Services of Organic Wheat and Clover Relay Intercrops. Eur. J. Agron. 2016, 73, 73–82. [Google Scholar] [CrossRef]
- Amossé, C.; Jeuffroy, M.H.; David, C. Relay Intercropping of Legume Cover Crops in Organic Winter Wheat: Effects on Performance and Resource Availability. F. Crop. Res. 2013, 145, 78–87. [Google Scholar] [CrossRef]
- Vrignon-Brenas, S.; Celette, F.; Piquet-Pissaloux, A.; David, C. Biotic and Abiotic Factors Impacting Establishment and Growth of Relay Intercropped Forage Legumes. Eur. J. Agron. 2016, 81, 169–177. [Google Scholar] [CrossRef]
- Roberts, R.; Jackson, R.W.; Mauchline, T.H.; Hirsch, P.R.; Shaw, L.J.; Döring, T.F.; Jones, H.E. Is There Sufficient Ensifer and Rhizobium Species Diversity in UK Farmland Soils to Support Red Clover (Trifolium pratense), White Clover (T. repens), Lucerne (Medicago sativa) and Black Medic (M. lupulina)? Appl. Soil Ecol. 2017, 120, 35–43. [Google Scholar] [CrossRef]
- Romanyà, J.; Casals, P. Biological Nitrogen Fixation Response to Soil Fertility Is Species-Dependent in Annual Legumes. J. Soil Sci. Plant Nutr. 2020, 20, 546–556. [Google Scholar] [CrossRef]
- Pandey, A.; Li, F.; Askegaard, M.; Olesen, J.E. Biological Nitrogen Fi Xation in Three Long-Term Organic and Conventional Arable Crop Rotation Experiments in Denmark. Eur. J. Agron. 2017, 90, 87–95. [Google Scholar] [CrossRef]
- Peoples, M.B.; Bowman, A.M.; Gault, R.R.; Herridge, D.F.; McCallum, M.H.; McCormick, K.M.; Norton, R.M.; Rochester, I.J.; Scammell, G.J.; Schwenke, G.D. Factors Regulating the Contributions of Fixed Nitrogen by Pasture and Crop Legumes to Different Farming Systems of Eastern Australia. Plant Soil 2001, 228, 29–41. [Google Scholar] [CrossRef]
- Sanz-Saez, A.; Morales, F.; Arrese-Igor, C.; Aranjuelo, I.P. Deficiency: A Major Limiting Factor for Rhizobial Symbiosis. In Legume Nitrogen Fixation in Soils with Low Phosphorus Availability; Sulieman, S., Tran, L.-S.P., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 21–40. ISBN 978-3-319-55728-1. [Google Scholar]
- Guiducci, M.; Tosti, G.; Falcinelli, B.; Benincasa, P. Sustainable Management of Nitrogen Nutrition in Winter Wheat through Temporary Intercropping with Legumes. Agron. Sustain. Dev. 2018, 38, 31. [Google Scholar] [CrossRef]
- Zhang, Y.; Carswell, A.; Jiang, R.; Cardenas, L.; Chen, D.; Misselbrook, T. The Amount, but Not the Proportion, of N2 Fixation and Transfers to Neighboring Plants Varies across Grassland Soils. Soil Sci. Plant Nutr. 2020, 66, 481–488. [Google Scholar] [CrossRef]
- Ripley, B.S.; Pammenter, N.W.; Smith, V.R. Function of Leaf Hairs Revisited: The Hair Layer on Leaves Arctotheca Populifolia Reduces Photoinhibition, but Leads to Higher Leaf Temperatures Caused by Lower Transpiration Rates. J. Plant Physiol. 1999, 155, 78–85. [Google Scholar] [CrossRef]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and Review of Trichomes in Plants. BMC Plant Biol. 2021, 21, 70. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; pp. 3–32. [Google Scholar]
- Wendling, M.; Charles, R.; Herrera, J.; Amossé, C.; Jeangros, B.; Walter, A.; Büchi, L. Effect of Species Identity and Diversity on Biomass Production and Its Stability in Cover Crop Mixtures. Agric. Ecosyst. Environ. 2019, 281, 81–91. [Google Scholar] [CrossRef]
- Ranaldo, M.; Carlesi, S.; Costanzo, A.; Bàrberi, P. Functional Diversity of Cover Crop Mixtures Enhances Biomass Yield and Weed Suppression in a Mediterranean Agroecosystem. Weed Res. 2020, 60, 96–108. [Google Scholar] [CrossRef]
- Wayman, S.; Kissing Kucek, L.; Mirsky, S.B.; Ackroyd, V.; Cordeau, S.; Ryan, M.R. Organic and Conventional Farmers Differ in Their Perspectives on Cover Crop Use and Breeding. Renew. Agric. Food Syst. 2017, 32, 376–385. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A Framework for Evaluating Ecosystem Services Provided by Cover Crops in Agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Daryanto, S.; Fu, B.; Wang, L.; Jacinthe, P.A.; Zhao, W. Quantitative Synthesis on the Ecosystem Services of Cover Crops. Earth-Sci. Rev. 2018, 185, 357–373. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Service Crop Mixture | Speed of Growth | Frost Sensitivity | SK | OG | |
|---|---|---|---|---|---|
| Frost-sensitive annual * | Fast | High | X | X | |
| Perennial ** | Slow | Low | X | X | |
| Frost-tolerant annual *** | Fast | Relatively low | X | ||
| Control, no service crop | X | X | |||
| System strategy | Sowing of service crops | Number of row hoeing events | Sowing of winter wheat | SK | OG |
| Early Intra | Same time as oats, in oat rows | 2 | Between oat rows | X | X |
| Late Inter | At first row hoeing, in inter-row centers | 1 | In oat stubble | X | X |
| Late Adjacent | At first row hoeing, adjacent to oat row | 2 | Between oat rows | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagerquist, E.; Menegat, A.; Dahlin, A.S.; Parsons, D.; Watson, C.; Ståhl, P.; Gunnarsson, A.; Bergkvist, G. Temporal and Spatial Positioning of Service Crops in Cereals Affects Yield and Weed Control. Agriculture 2022, 12, 1398. https://doi.org/10.3390/agriculture12091398
Lagerquist E, Menegat A, Dahlin AS, Parsons D, Watson C, Ståhl P, Gunnarsson A, Bergkvist G. Temporal and Spatial Positioning of Service Crops in Cereals Affects Yield and Weed Control. Agriculture. 2022; 12(9):1398. https://doi.org/10.3390/agriculture12091398
Chicago/Turabian StyleLagerquist, Elsa, Alexander Menegat, Anna Sigrun Dahlin, David Parsons, Christine Watson, Per Ståhl, Anita Gunnarsson, and Göran Bergkvist. 2022. "Temporal and Spatial Positioning of Service Crops in Cereals Affects Yield and Weed Control" Agriculture 12, no. 9: 1398. https://doi.org/10.3390/agriculture12091398