
Double-Cropped Winter Camelina with and without Added Nitrogen: Effects on Productivity and Soil Available Nitrogen

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Experimental Design
2.3. Agronomic Management
- Study 1
- Study 2
2.4. Data Collection
2.5. Statistical Analysis
3. Results and Discussion
3.1. Weather Conditions
3.2. Growth and Development of Winter Camelina with and without Added N Fertilizer
3.3. Oil and Protein of Winter Camelina
3.4. Soil Available Nitrogen in Winter Camelina Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weyers, S.; Thom, M.; Forcella, F.; Eberle, C.; Matthees, H.; Gesch, R.; Ott, M.; Feyereisen, G.; Strock, J.; Wyse, D. Reduced Potential for Nitrogen Loss in Cover Crop-Soybean Relay Systems in a Cold Climate. J. Environ. Qual. 2019, 48, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in Agriculture: Balancing the Cost of an Essential Resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Basso, B.; Shuai, G.; Zhang, J.; Robertson, G.P. Yield stability analysis reveals sources of large-scale nitrogen loss from the US Midwest. Sci. Rep. 2019, 9, 5774. [Google Scholar] [CrossRef]
- Randall, G.W.; Huggins, D.R.; Russelle, M.P.; Fuchs, D.J.; Nelson, W.W.; Anderson, J.L. Nitrate losses through subsurface tile drainage in conservation reserve program, alfalfa, and row crop systems. J. Environ. Qual. 1997, 26, 1240–1247. [Google Scholar] [CrossRef]
- Peterson, A.T.; Berti, M.T.; Samarappuli, D. Intersowing cover crops into standing soybean in the US upper midwest. Agronomy 2019, 9, 264. [Google Scholar] [CrossRef]
- Doran, J.W. Soil health and global sustainability: Translating science into practice. Agric. Ecosyst. Environ. 2002, 88, 119–127. [Google Scholar] [CrossRef]
- Ott, M.A.; Eberle, C.A.; Thom, M.D.; Archer, D.W.; Forcella, F.; Gesch, R.W.; Wyse, D.L. Economics and agronomics of relay-cropping pennycress and Camelina with Soybean in Minnesota. Agron. J. 2019, 111, 1281–1292. [Google Scholar] [CrossRef]
- Sindelar, A.J.; Schmer, M.R.; Gesch, R.W.; Forcella, F.; Eberle, C.A.; Thom, M.D.; Archer, D.W. Winter oilseed production for biofuel in the US Corn Belt: Opportunities and limitations. GCB Bioenergy 2017, 9, 508–524. [Google Scholar] [CrossRef]
- Johnson, G.A.; Wells, M.S.; Anderson, K.; Gesch, R.W.; Forcella, F.; Wyse, D.L. Yield tradeoffs and nitrogen between pennycress, camelina, and soybean in relay- and double-crop systems. Agron. J. 2017, 109, 2128–2135. [Google Scholar] [CrossRef]
- Gesch, R.W.; Archer, D.W. Double-cropping with winter camelina in the northern Corn Belt to produce fuel and food. Ind. Crops Prod. 2013, 44, 718–725. [Google Scholar] [CrossRef]
- Moore, K.J.; Karlen, D.L. Double cropping opportunities for biomass crops in the North Central USA. Biofuels 2013, 4, 605–615. [Google Scholar] [CrossRef]
- CTIC; SARE; ASTA. National Cover Crop Survey Annual Report 2019–2020. 2020. Available online: https://www.sare.org/wp-content/uploads/2019-2020-National-Cover-Crop-Survey.pdf (accessed on 12 September 2022).
- Berti, M.; Gesch, R.; Johnson, B.; Ji, Y.; Seames, W.; Aponte, A. Double- and relay-cropping of energy crops in the northern Great Plains, USA. Ind. Crops Prod. 2015, 75, 26–34. [Google Scholar] [CrossRef]
- Johnson, G.A.; Kantar, M.B.; Betts, K.J.; Wyse, D.L. Field pennycress production and weed control in a double crop system with soybean in minnesota. Agron. J. 2015, 107, 532–540. [Google Scholar] [CrossRef]
- Gesch, R.W.; Archer, D.W.; Berti, M.T. Dual cropping winter camelina with soybean in the northern corn belt. Crop. Econ. Prod. Manag. 2014, 106, 1735–1745. [Google Scholar] [CrossRef]
- Zubr, J. Oil-seed crop: Camelina sativa. Ind. Crops Prod. 1997, 6, 113–119. [Google Scholar] [CrossRef]
- Ghamkhar, K.; Croser, J.; Aryanmanesh, N.; Campbell, M.; Kon’kova, N.; Francis, C. Camelina (Camelina sativa (L.) Crantz) as an alternative oilseed: Molecular and ecogeographical analyses. Genome 2010, 53, 558–567. [Google Scholar] [CrossRef]
- Prakhova, T.Y.; Prakhov, V.A.; Danilov, M.V. Changes in the fat-acidic composition of Camelina Sativa oilseeds depending on hydrothermal conditions. Russ. Agric. Sci. 2018, 44, 221–223. [Google Scholar] [CrossRef]
- Zanetti, F.; Eynck, C.; Christou, M.; Krzyżaniak, M.; Righini, D.; Alexopoulou, E.; Stolarski, M.J.; Van Loo, E.N.; Puttick, D.; Monti, A. Agronomic performance and seed quality attributes of Camelina (Camelina sativa L. crantz) in multi-environment trials across Europe and Canada. Ind. Crops Prod. 2017, 107, 602–608. [Google Scholar] [CrossRef]
- Heggenstaller, A.H.; Anex, R.P.; Liebman, M.; Sundberg, D.N.; Gibson, L.R. Productivity and nutrient dynamics in bioenergy double-cropping systems. Agron. J. 2008, 100, 1740–1748. [Google Scholar] [CrossRef]
- Wittenberg, A.; Anderson, J.V.; Berti, M.T. Crop growth and productivity of winter camelina in response to sowing date in the Northwestern Corn Belt of the USA. Ind. Crops Prod. 2020, 158, 113036. [Google Scholar] [CrossRef]
- Gesch, R.W.; Dose, H.L.; Forcella, F. Camelina growth and yield response to sowing depth and rate in the northern Corn Belt USA. Ind. Crops Prod. 2017, 95, 416–421. [Google Scholar] [CrossRef]
- Liu, R.; Wells, M.S.; Garcia y Garcia, A. Relay and sequential cropping corn with winter oilseed crops in northern climates. Nutr. Cycl. Agroecosyst. 2020, 116, 195–203. [Google Scholar] [CrossRef]
- Strock, J.S.; Porter, P.M.; Russelle, M.P. Cover Cropping to Reduce Nitrate Loss through Subsurface Drainage in the Northern U.S. Corn Belt. J. Environ. Qual. 2004, 33, 1010. [Google Scholar] [CrossRef] [PubMed]
- National Cooperative Soil Survey. National Cooperative Soil Characterization. Available online: https://ncsslabdatamart.sc.egov.usda.gov/ (accessed on 12 September 2022).
- U.S. Department of Agriculture, Agricultural Research Service. USDA Plant Hardiness Zone Map. 2012. Available online: https://planthardiness.ars.usda.gov/ (accessed on 12 September 2022).
- Gesch, R.W.; Cermak, S.C. Sowing date and tillage effects on fall-seeded Camelina in the Northern Corn Belt. Agron. J. 2011, 103, 980–987. [Google Scholar] [CrossRef]
- Martinelli, T.; Galasso, I. Phenological growth stages of Camelina sativa according to the extended BBCH scale. Ann. Appl. Biol. 2011, 158, 87–94. [Google Scholar] [CrossRef]
- Johnson, J.M.F.; Gesch, R.W. Calendula and camelina response to nitrogen fertility. Ind. Crops Prod. 2013, 43, 684–691. [Google Scholar] [CrossRef]
- Doane, T.A.; Horwáth, W.R. Spectrophotometric determination of nitrate with a single reagent. Anal. Lett. 2003, 36, 2713–2722. [Google Scholar] [CrossRef]
- Walia, M.K.; Wells, M.S.; Cubins, J.; Wyse, D.; Gardner, R.D.; Forcella, F.; Gesch, R. Winter camelina seed yield and quality responses to harvest time. Ind. Crops Prod. 2018, 124, 765–775. [Google Scholar] [CrossRef]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Patel, S.; Lenssen, A.W.; Moore, K.J.; Mohammed, Y.A.; Gesch, R.W.; Wells, M.S.; Johnson, B.L.; Berti, M.T.; Matthees, H.L. Interseeded pennycress and camelina yield and influence on row crops. Agron. J. 2021, 113, 2629–2647. [Google Scholar] [CrossRef]
- Zanetti, F.; Alberghini, B.; Marjanović Jeromela, A.; Grahovac, N.; Rajković, D.; Kiprovski, B.; Monti, A. Camelina, an ancient oilseed crop actively contributing to the rural renaissance in Europe. A review. Agron. Sustain. Dev. 2021, 41, 2. [Google Scholar] [CrossRef]
- Gesch, R.W.; Wells, M.S.; Hard, A. Desiccation of corn allows earlier direct seeding of winter camleina in the Norhtern Corn Belt. Crop Sci. 2021, 61, 2787–2797. [Google Scholar] [CrossRef]
- Gesch, R.W.; Johnson, J.M.F. Water use in camelina-soybean dual cropping systems. Agron. J. 2015, 107, 1098–1104. [Google Scholar] [CrossRef]
- Wittenberg, A.; Anderson, J.V.; Berti, M.T. Winter and summer annual biotypes of camelina have different morphology and seed characteristics. Ind. Crops Prod. 2019, 135, 230–237. [Google Scholar] [CrossRef]
- Gesch, R.W.; Mohammed, Y.A.; Walia, M.K.; Hulke, B.S.; Anderson, J.V. Double-cropping oilseed sunflower after winter camelina. Ind. Crops Prod. 2022, 181, 114811. [Google Scholar] [CrossRef]
- Gesch, R.W.; Matthees, H.L.; Alvarez, A.L.; Gardner, R.D. Winter camelina: Crop growth, seed yield, and quality response to cultivar and seeding rate. Crop Sci. 2018, 58, 2089–2098. [Google Scholar] [CrossRef]
- Assefa, Y.; Purcell, L.C.; Salmeron, M.; Naeve, S.; Casteel, S.N.; Kovács, P.; Archontoulis, S.; Licht, M.; Below, F.; Kandel, H.; et al. Assessing variation in US soybean seed composition (protein and oil). Front. Plant Sci. 2019, 10, 298. [Google Scholar] [CrossRef]
- Rod, K.S.; Shockley, J.; Knott, C.A. Seed yield, seed quality, profitability, and risk analysis among double crop soybean maturity groups and seeding rates. Agron. J. 2021, 113, 1792–1802. [Google Scholar] [CrossRef]
- Vyn, T.J.; Yin, X.; Bruulsema, T.W.; Jackson, C.J.C.; Rajcan, I.; Brouder, S.M. Potassium fertilization effects on isoflavone concentrations in soybean [Glycine max (L.) Merr.]. Agric. Food Chem. 2002, 50, 3501–3506. [Google Scholar] [CrossRef]
- Krueger, K.; Goggi, A.S.; Mallarino, A.P.; Mullen, R.E. Phosphorus and potassium fertilization effects on soybean seed quality and composition. Crop Sci. 2013, 53, 602–610. [Google Scholar] [CrossRef] [Green Version]
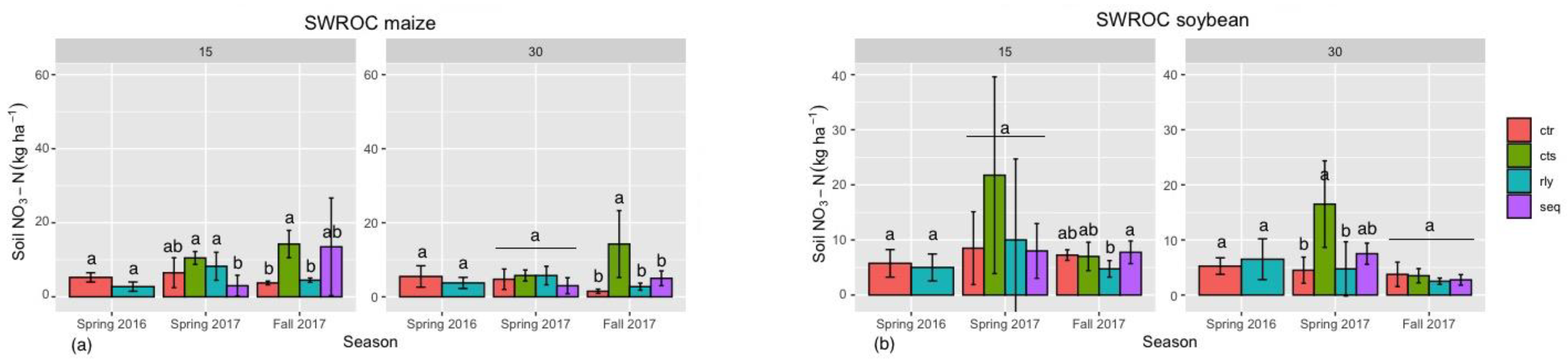
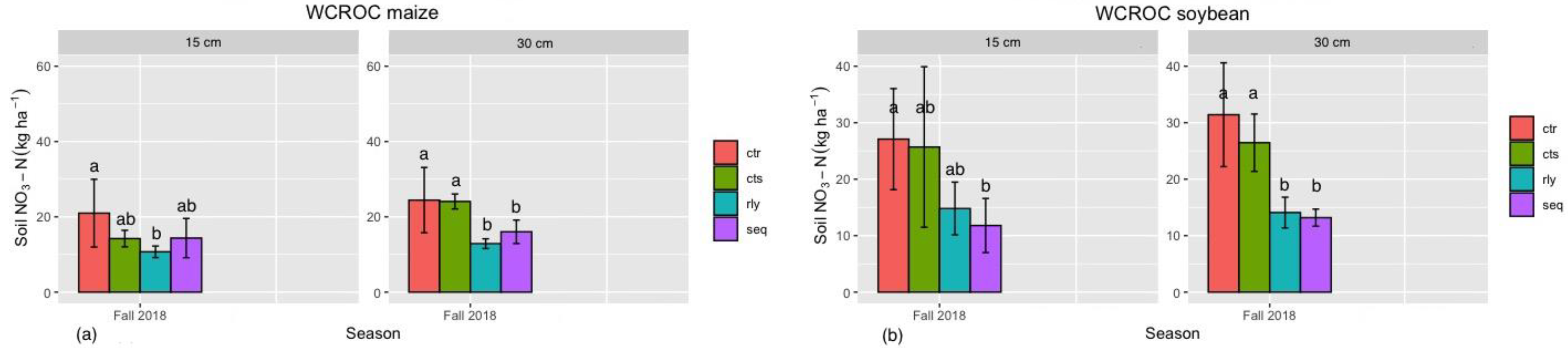


{kind=link}
{kind=link}
{kind=link}
| Study | Site–Year ‡ | Soil Textural Class | OM † (%) | pH | CEC (meq 100 g−1) | NO3–N | Bray P | K | Ca | Mg |
|---|---|---|---|---|---|---|---|---|---|---|
| ------------------- ppm ------------------- | ||||||||||
| Study 1 | SWROC 2015 | Loam | 3.9 | 5.2 | 27 | 10.7 | 20 | 168 | 2333 | 507 |
| Study 2 | SWROC 2018 | Loam | 3.6 | 5.5 | 20 | 1.5 | 10 | 95 | 1908 | 386 |
| WCROC 2018 | Sandy loam | 6.4 | 6.2 | 26 | 10.5 | 18 | 171 | 3115 | 630 | |
| Month | Southwest Research and Outreach Center | West Central Research and Outreach Center | |||||||
|---|---|---|---|---|---|---|---|---|---|
| --------------- Study 1 --------------- | -------------------------------- Study 2-------------------------------- | ||||||||
| LTA (1994–2019) | 2015 | 2016 | 2017 | 2019 | 2020 | LTA (1994–2019) | 2018 | 2019 | |
| Precipitation (mm) | Deviation (mm) from LTA | Precipitation (mm) | Deviation (mm) from LTA | ||||||
| January | 14 | −2 | −6 | −2 | −3 | −2 | 19 | −16 | −12 |
| February | 15 | −11 | 3 | −13 | 30 | 4 | 19 | 3 | 1 |
| March | 35 | −25 | 16 | −25 | 33 | 38 | 30 | −3 | 18 |
| April | 75 | −43 | 10 | 2 | 83 | −41 | 60 | −43 | −3 |
| May | 99 | 39 | 42 | 53 | 15 | −11 | 79 | −25 | 24 |
| June | 111 | 18 | −45 | −42 | 6 | −4 | 109 | 108 | 9 |
| July | 96 | 0 | 80 | 6 | 21 | 48 | 102 | 66 | 13 |
| August | 88 | 25 | 47 | 37 | −32 | 9 | 92 | −11 | 48 |
| September | 87 | 0 | 47 | −32 | 68 | 27 | 72 | −25 | 97 |
| October | 60 | −19 | 12 | 89 | 42 | −35 | 69 | 0 | 8 |
| November | 29 | 55 | 18 | −27 | −1 | 10 | 24 | −2 | −15 |
| December | 21 | 13 | 8 | −11 | 18 | −10 | 20 | 6 | 6 |
| Average Air Temperature (°C) | Deviation (°C) from LTA | Average Air Temperature (°C) | Deviation (°C) from LTA | ||||||
| January | −9 | 2 | 0 | 1 | −3 | 0 | −11 | 0 | −3 |
| February | −6 | −6 | 2 | 5 | −9 | −2 | −9 | −4 | −8 |
| March | 2 | 0 | 2 | −2 | −6 | −1 | 0 | −3 | −6 |
| April | 10 | −1 | −1 | −2 | −3 | −4 | 10 | −10 | −5 |
| May | 18 | −4 | −3 | −4 | −6 | −5 | 18 | −1 | −7 |
| June | 24 | −4 | −3 | −3 | −3 | −1 | 23 | −2 | −4 |
| July | 25 | −3 | −3 | −3 | −3 | −2 | 25 | −4 | −3 |
| August | 23 | −3 | −2 | −4 | −3 | −2 | 22 | −2 | −3 |
| September | 18 | 1 | 0 | 0 | 0 | −2 | 16 | 0 | 0 |
| October | 10 | 0 | 0 | −1 | −3 | −5 | 8 | −4 | −2 |
| November | 1 | 3 | 4 | −1 | −3 | 2 | −1 | −8 | −2 |
| December | −6 | 3 | −2 | −2 | −1 | 2 | −8 | −3 | −1 |
| Year | Cropping System ‡ | Followed by Maize | Followed by Soybean | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Biomass | Grain | Grain | |||||||
| Yield | Oil | Protein | Yield | Oil | Protein | ||||
| (kg ha−1) | (%) | (kg ha−1) | (%) | ||||||
| Study 1—Broadcast Seeded, non-N–fertilized | |||||||||
| Southwest Research and Outreach Center | |||||||||
| 2016 | Relay | 2553 a ± 51 | 494 b § ± 28 | − | − | 1531 b ± 367 | 247 b ± 147 | − | − |
| Sequential | 2808 a ± 65 | 394 b ± 21 | − | − | 2871 b ± 352 | 480 b ± 244 | − | − | |
| 2017 | Relay | 3106 a ± 83 | 609 a ± 18 | 27.2 a ± 0.3 | 21.5 a ± 3.6 | 3760 a ± 332 | 735 a ± 140 | 26.7 a ± 1.4 | 19.4 b ± 2.3 |
| Sequential | 3505 a ± 54 | 703 a ± 22 | 26.4 a ± 1.5 | 27.1 a ± 4.2 | 3840 a ± 1261 | 786 a ± 491 | 26.7 a ± 1.8 | 25.6 a ± 3.6 | |
| Study 2—Drill Seeded, N–fertilized | |||||||||
| West Central Research and Outreach Center | |||||||||
| 2019 | Relay | 2965 b ± 49 | 875 b ± 15 | 33.3 b ± 1.3 | 20.8 a ± 1.4 | 3028 b ± 182 | 851 b ± 64 | 35.1 a ± 1.2 | 17.9 a ± 1.8 |
| Sequential | 3943 a ± 61 | 1393 a ± 31 | 35.9 a ± 1.0 | 17.5 a ± 2.8 | 4134 a ± 735 | 1461 a ± 273 | 35.9 a ± 0.93 | 17.4 a ± 0.9 | |
| Southwest Research and Outreach Center | |||||||||
| 2020 | Relay | 4171 a ± 38 | 1170 a ± 91 | 31.7 a ± 2.1 | 16.3 a ± 2.0 | 3949 a ± 309 | 1061 b ± 168 | 32.7 a ± 1.1 | 18.1 a ± 0.9 |
| Sequential | 3840 a ± 36 | 1159 a ± 11 | 32.7 a ± 0.9 | 16.4 a ± 2.2 | 4488 a ± 286 | 1288 a ± 24 | 32.6 a ± 2.1 | 14.9 a ± 2.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregg, S.; Coulter, J.A.; Strock, J.S.; Liu, R.; Garcia y Garcia, A. Double-Cropped Winter Camelina with and without Added Nitrogen: Effects on Productivity and Soil Available Nitrogen. Agriculture 2022, 12, 1477. https://doi.org/10.3390/agriculture12091477
Gregg S, Coulter JA, Strock JS, Liu R, Garcia y Garcia A. Double-Cropped Winter Camelina with and without Added Nitrogen: Effects on Productivity and Soil Available Nitrogen. Agriculture. 2022; 12(9):1477. https://doi.org/10.3390/agriculture12091477
Chicago/Turabian StyleGregg, Stephen, Jeffrey A. Coulter, Jeffrey S. Strock, Ronghao Liu, and Axel Garcia y Garcia. 2022. "Double-Cropped Winter Camelina with and without Added Nitrogen: Effects on Productivity and Soil Available Nitrogen" Agriculture 12, no. 9: 1477. https://doi.org/10.3390/agriculture12091477
APA StyleGregg, S., Coulter, J. A., Strock, J. S., Liu, R., & Garcia y Garcia, A. (2022). Double-Cropped Winter Camelina with and without Added Nitrogen: Effects on Productivity and Soil Available Nitrogen. Agriculture, 12(9), 1477. https://doi.org/10.3390/agriculture12091477