

Physical–Chemical and Metataxonomic Characterization of the Microbial Communities Present during the Fermentation of Three Varieties of Coffee from Colombia and Their Sensory Qualities

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location, Sampling, Characterization of the Harvest and Fermentation
2.2. Physicochemical Analysis
2.2.1. Determination of Acidity
2.2.2. Determination of Sugars (Sucrose, Glucose and Fructose)
- Extract preparations
- Sugar analysis
2.2.3. Total Protein Quantification
2.3. Microbiological Analysis
2.3.1. Cultivation and Isolation of Microorganisms in Mucilage and Water
2.3.2. Microbial Identification
2.4. Molecular and Metataxonomic Analyses
2.5. Physical and Sensory Analyses of Coffee
3. Results
3.1. Characterization of the Harvest
3.2. Physicochemical Analysis
3.2.1. Temperature, pH, °Brix and Acidity in the Mucilage
3.2.2. Sugar and Protein Contents
3.3. Microbiological Analysis of Coffee Mucilage and Water
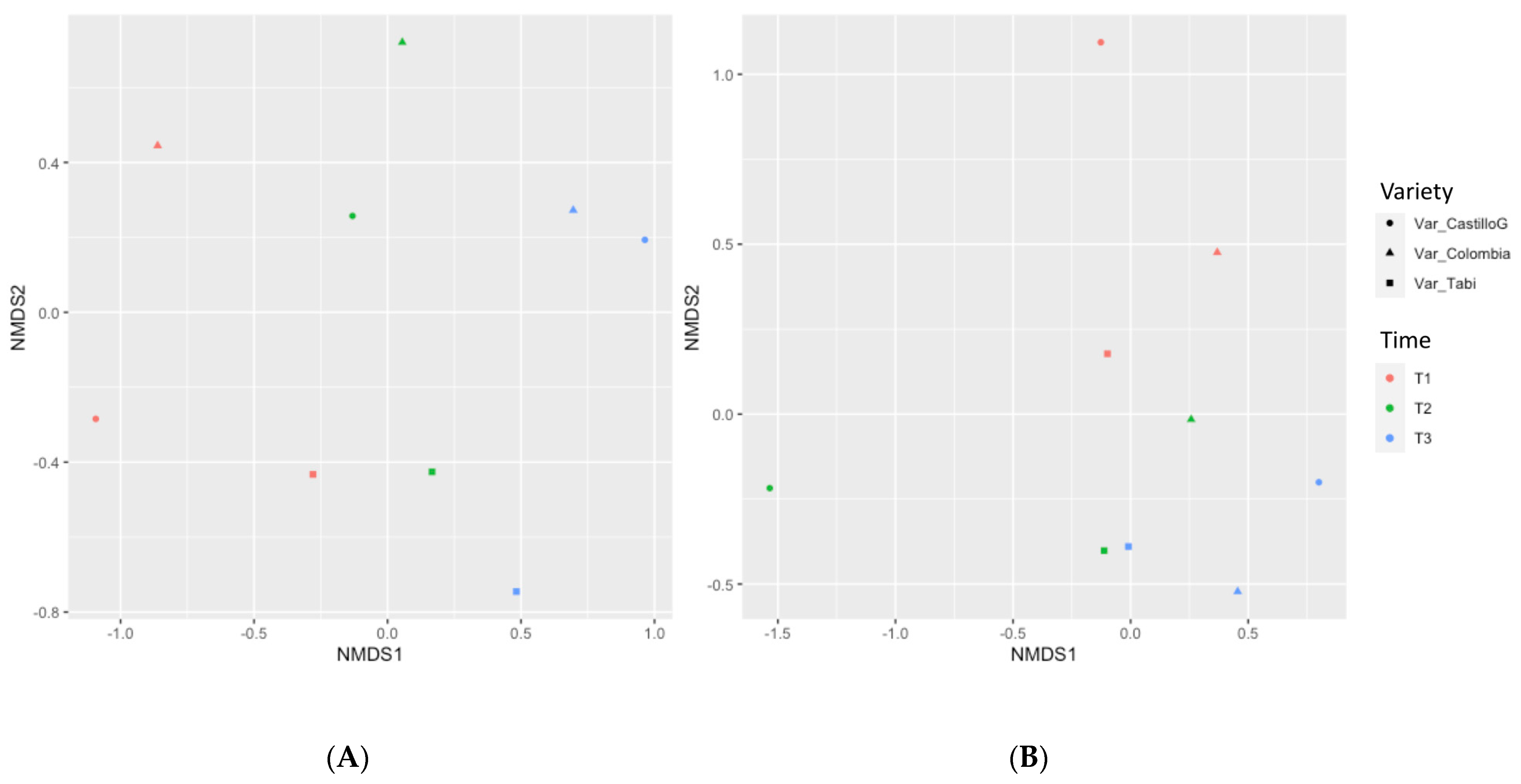
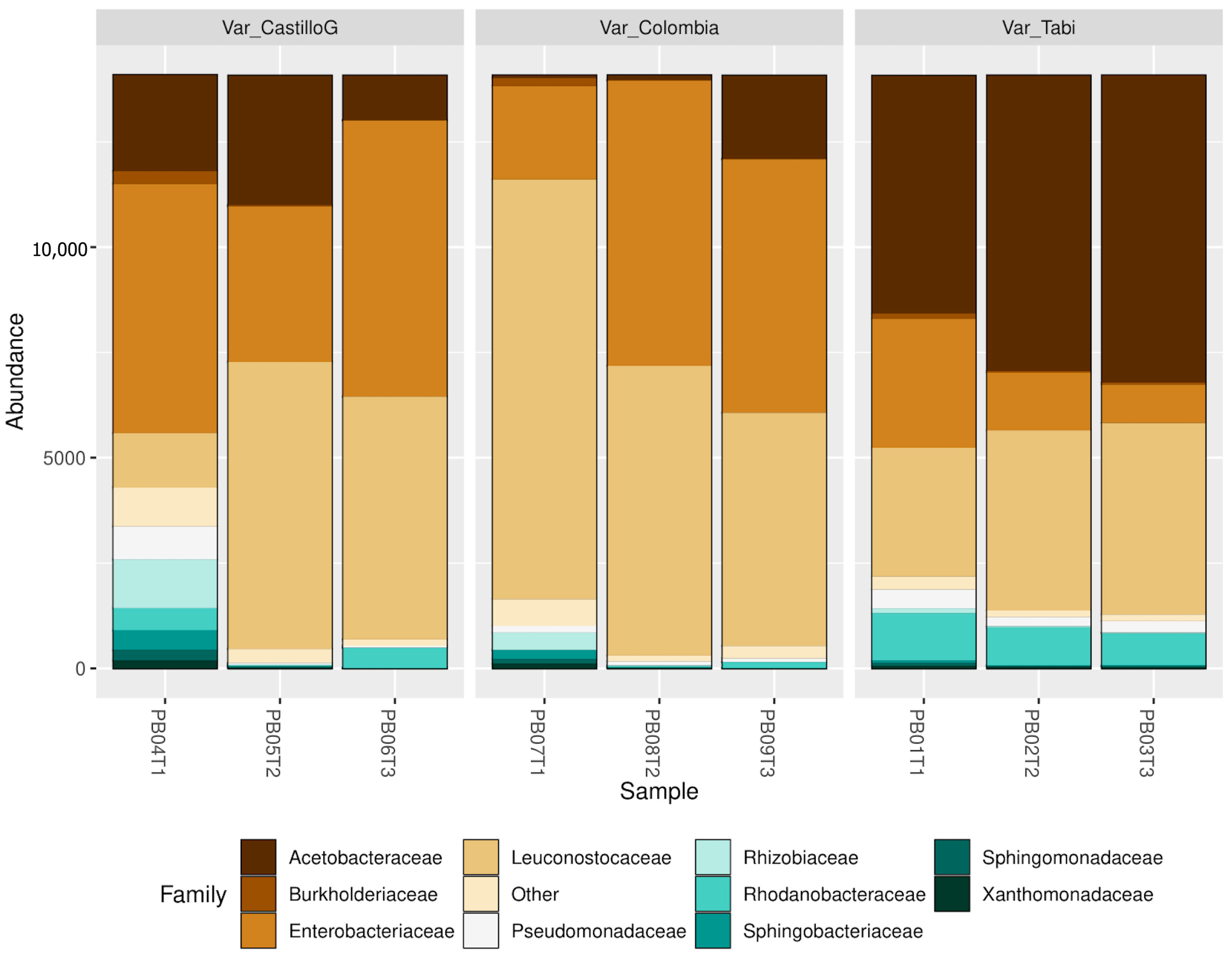
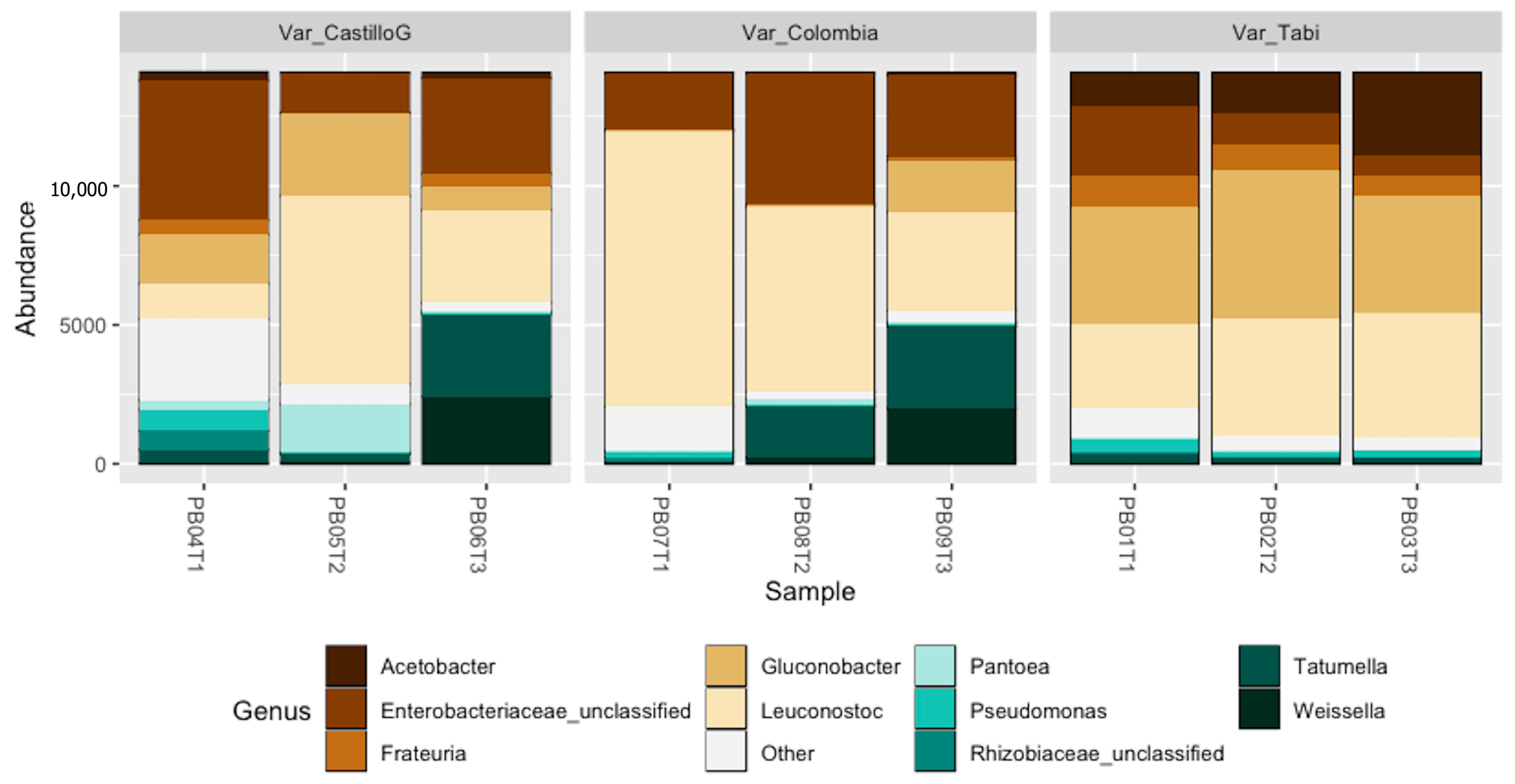
3.4. Molecular and Metataxonomic Analyses
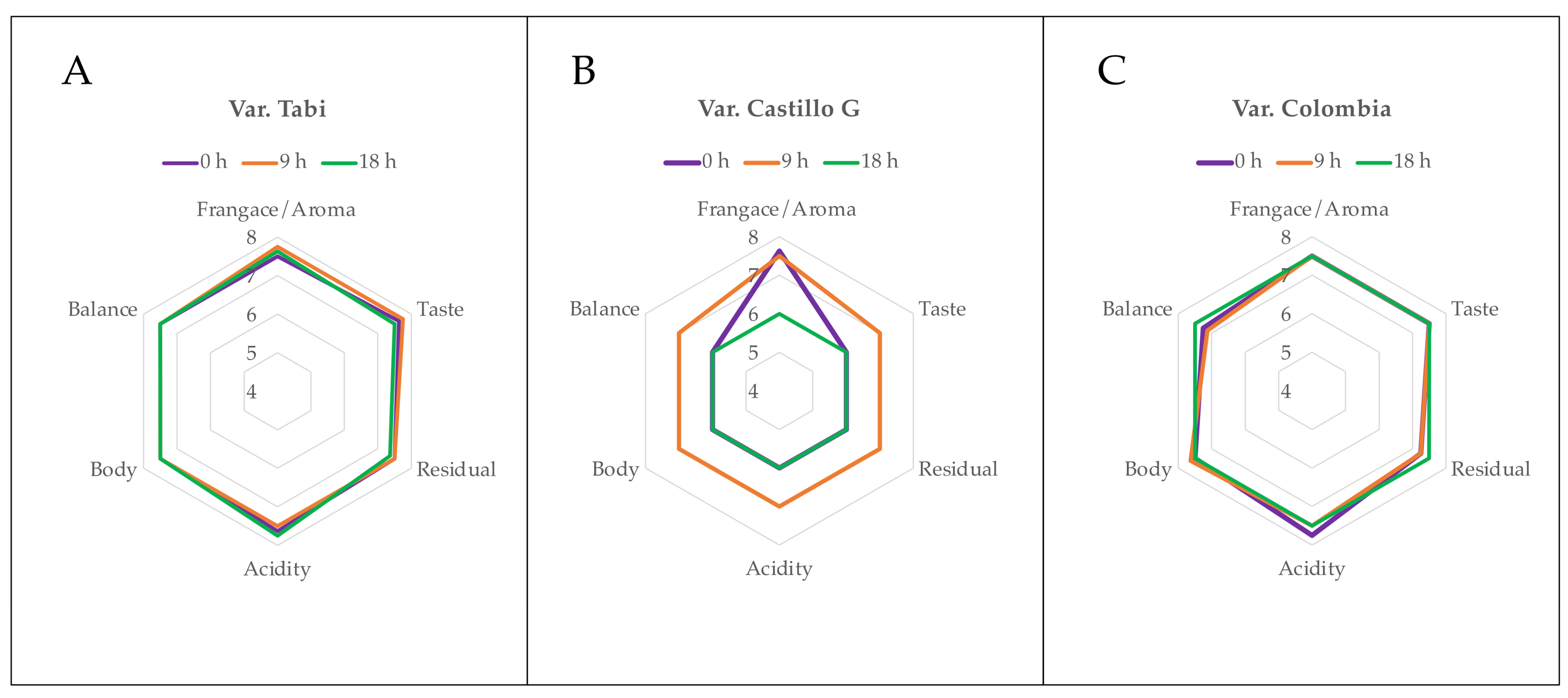
3.5. Physical and Sensory Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministerio de Agricultura y Desarrollo Rural CADENA CAFÉ 2021. Available online: https://sioc.minagricultura.gov.co/Cafe/Documentos/2021-03-31%20Cifras%20Sectoriales.pdf (accessed on 5 February 2022).
- Federación Nacional de Cafeteros Informe del Gerente 90 Congreso Nacional de Cafeteros 2022. Available online: https://federaciondecafeteros.org/app/uploads/2022/12/Informe-del-Gerente-D.pdf (accessed on 20 March 2023).
- Flórez, C.P.; Quiroga-Cardona, J.; Arias, J.C. Variedades Del Café. In Guía más Agronomia, más Productividad, más Calidad; Cenicafé: Manizales, Colombia, 2021; pp. 11–29. ISBN 978-958-8490-49-6. [Google Scholar]
- Vegro, C.L.R.; de Almeida, L.F. Global Coffee Market: Socio-Economic and Cultural Dynamics. In Coffee Consumption and Industry Strategies in Brazil; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–19. ISBN 978-0-12-814721-4. [Google Scholar]
- Puerta Quintero, G.I.; Ríos Arias, S. Composición química del mucílago de café, según el tiempo de fermentación y refrigeración. Cenicafé 2011, 62, 23–40. [Google Scholar]
- Avallone, S.; Guyot, B.; Brillouet, J.M.; Olguin, E.; Guiraud, J.P. Microbiological and Biochemical Study of Coffee Fermentation. Curr. Microbiol. 2001, 42, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Evangelista; Reis, S.; da Cruz Pedroso Miguel, M.G.; Silva, C.F.; Pinheiro, A.C.M.; Schwan, R.F. Microbiological Diversity Associated with the Spontaneous Wet Method of Coffee Fermentation. Int. J. Food Microbiol. 2015, 210, 102–112. [Google Scholar] [CrossRef]
- Nasanit, R.; Satayawut, K. Microbiological Study During Coffee Fermentation of Coffea arabica var. chiangmai 80 in Thailand. Kasetsart J. Nat. Sci. 2015, 49, 32–41. [Google Scholar]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Contreras, G.F.; Cai, Z.; Moccand, C.; Weckx, S.; De Vuyst, L. Influence of Various Processing Parameters on the Microbial Community Dynamics, Metabolomic Profiles, and Cup Quality during Wet Coffee Processing. Front. Microbiol. 2019, 10, 2621. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, F.; Zhang, S.J.; Pothakos, V.; Torres, J.; Lambot, C.; Moroni, A.V.; Callanan, M.; Sybesma, W.; Weckx, S.; De Vuyst, L. Exploring the Impacts of Postharvest Processing on the Microbiota and Metabolite Profiles during Green Coffee Bean Production. Appl. Environ. Microbiol. 2017, 83, e02398-16. [Google Scholar] [CrossRef]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Torres, J.; Falconi, C.; Moccand, C.; Weckx, S.; De Vuyst, L. Following Coffee Production from Cherries to Cup: Microbiological and Metabolomic Analysis of Wet Processing of Coffea Arabica. Appl. Environ. Microbiol. 2019, 85, e02635-18. [Google Scholar] [CrossRef]
- Pothakos, V.; De Vuyst, L.; Zhang, S.J.; De Bruyn, F.; Verce, M.; Torres, J.; Callanan, M.; Moccand, C.; Weckx, S. Temporal Shotgun Metagenomics of an Ecuadorian Coffee Fermentation Process Highlights the Predominance of Lactic Acid Bacteria. Curr. Res. Biotechnol. 2020, 2, 1–15. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Carvalho, J.C.; Soccol, V.T.; Soccol, C.R. High-Throughput RRNA Gene Sequencing Reveals High and Complex Bacterial Diversity Associated with Brazilian Coffee Beans Fermentation. Food Technol. Biotechnol. 2017, 56, 90–95. [Google Scholar] [CrossRef]
- Martinez, S.J.; Simão, J.B.P.; Pylro, V.S.; Schwan, R.F. The Altitude of Coffee Cultivation Causes Shifts in the Microbial Community Assembly and Biochemical Compounds in Natural Induced Anaerobic Fermentations. Front. Microbiol. 2021, 12, 671395. [Google Scholar] [CrossRef]
- Vera Pacheco, K.; Valdivieso Quintero, W.; Mantilla-Paredes, A.J.; Jaimes, W.; Torrado, J.; Zafra, G. Functional Metagenomic Analysis of the Coffee (Coffea arabica) Fermentation. Chem. Eng. Trans. 2018, 64, 355–360. [Google Scholar] [CrossRef]
- Elhalis, H.; Cox, J.; Zhao, J. Ecological Diversity, Evolution and Metabolism of Microbial Communities in the Wet Fermentation of Australian Coffee Beans. Int. J. Food Microbiol. 2020, 321, 108544. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.S.; Evangelista, S.R.; da Cruz Pedrozo Miguel, M.G.; van Mullem, J.; Silva, C.F.; Schwan, R.F. Microbiological and Chemical-Sensory Characteristics of Three Coffee Varieties Processed by Wet Fermentation. Ann. Microbiol. 2018, 68, 705–716. [Google Scholar] [CrossRef]
- Apolo, D.; Fernández, J.M.; Benítez, Á.; Figueroa, J.G.; Estrada, K.; Cruz, D. Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador. Diversity 2023, 15, 984. [Google Scholar] [CrossRef]
- Puerta Quintero, G.I.; Marín Mejía, J.; Osorio Betancur, G.A. Microbiología de La Fermentación Del Mucílago de Café Según Su Madurez y Selección. Rev. Cenicafé 2012, 63, 58–78. [Google Scholar]
- Hamdouche, Y.; Meile, J.C.; Nganou, D.N.; Durand, N.; Teyssier, C.; Montet, D. Discrimination of Post-Harvest Coffee Processing Methods by Microbial Ecology Analyses. Food Control. 2016, 65, 112–120. [Google Scholar] [CrossRef]
- De Oliveira Junqueira, A.C.; de Melo Pereira, G.V.; Coral Medina, J.D.; Alvear, M.C.R.; Rosero, R.; de Carvalho Neto, D.P.; Enríquez, H.G.; Soccol, C.R. First Description of Bacterial and Fungal Communities in Colombian Coffee Beans Fermentation Analysed Using Illumina-Based Amplicon Sequencing. Sci. Rep. 2019, 9, 8794. [Google Scholar] [CrossRef]
- Junior, D.B.; Guarçoni, R.C.; da Silva, M.D.C.S.; Veloso, T.G.R.; Kasuya, M.C.M.; da Silva Oliveira, E.C.; da Luz, J.M.R.; Moreira, T.R.; Debona, D.G.; Pereira, L.L. Microbial Fermentation Affects Sensorial, Chemical, and Microbial Profile of Coffee under Carbonic Maceration. Food Chem. 2021, 342, 128296. [Google Scholar] [CrossRef]
- Shen, X.; Wang, B.; Zi, C.; Huang, L.; Wang, Q.; Zhou, C.; Wen, W.; Liu, K.; Yuan, W.; Li, X. Interaction and Metabolic Function of Microbiota during the Washed Processing of Coffea Arabica. Molecules 2023, 28, 6092. [Google Scholar] [CrossRef]
- Cruz-O’Byrne, R.; Piraneque-Gambasica, N.; Aguirre-Forero, S. Microbial Diversity Associated with Spontaneous Coffee Bean Fermentation Process and Specialty Coffee Production in Northern Colombia. Int. J. Food Microbiol. 2021, 354, 109282. [Google Scholar] [CrossRef]
- Superintendencia de Industria y Comercio. Resolución 2484. 2017. Available online: https://www.sic.gov.co/sites/default/files/files/Denominacion%20de%20Origen/Resoluci%C3%B3n%202484%20Caf%C3%A9%20de%20la%20Sierra%20Nevada.pdf (accessed on 10 September 2021).
- Peñuela, A.E.; Sanz-Uribe, J.R. Obtenga Café de Calidad En El Proceso de Beneficio. In Guía más Agronomia, más Productividad, más Calidad; Cenicafé: Manizales, Colombia, 2021; pp. 189–218. ISBN 978-958-8490-49-6. [Google Scholar]
- Guerrero, Á.; Sanz-Uribe, J.R.; Peñuela-Martínez, A.E.; Ramírez, C.A. Mediverdes®: Un Método Para Medir La Calidad de La Recolección Del Café En El Campo. Av. Técnicos Cenicafé 2022, 536, 1–8. [Google Scholar] [CrossRef]
- Peñuela-Martínez, A.E.; Guerrero, Á.; Sanz-Uribe, J.R. Cromacafé® Herramienta Para Identificar Los Estados de Madurez de Las Variedades de Café de Fruto Rojo. Av. Técnicos Cenicafé 2022, 535, 1–8. [Google Scholar] [CrossRef]
- Peñuela-Martínez, A.E.; Pabón, J.; Sanz-Uribe, J.R. Método fermaestro: Para Determinar La Finalización de La Fermentación Del Mucílago de Café. Av. Técnicos Cenicafé 2013, 431, 1–8. [Google Scholar] [CrossRef]
- Oliveros Táscon, C.; Ramirez Gómez, C.; Tibaduiza Vianchá, C.; Sanz Uribe, J.R. Construcción de Secadores Solares Tipo Túnel Con Nuevos Materiales. Av. Técnicos Cenicafé 2017, 482, 1–8. [Google Scholar] [CrossRef]
- Tyl, C.; Sadler, G.D. pH and Titratable Acidity. In Food Analysis 2017; Nielsen, S.S., Ed.; Food Science Text Series; Springer: Cham, Switzerland, 2017; pp. 389–406. [Google Scholar] [CrossRef]
- Saint-Denis, T.; Goupy, J. Optimization of a Nitrogen Analyser Based on the Dumas Method. Anal. Chim. Acta 2004, 515, 191–198. [Google Scholar] [CrossRef]
- Gavidia-Valencia, J.G.; Venegas-Casanova, E.A.; Ríos, M.; Uribe-Villarreal, J.C.; Gutiérrez-Mendoza, D.D.; Rengifo-Penadillos, R.A.; Jara-Aguilar, D.R.; Martínez, J.L. Determinación del factor de conversión de nitrógeno a proteína en huevos de Coturnix coturnix L. (Codorniz japonesa). Arch. Venez. Farmacol. Ter. 2020, 39, 706–711. [Google Scholar] [CrossRef]
- Instituto Colombiano de Normas Técnicas y Certificación. NTC 3534. 2007. Available online: https://tienda.icontec.org/gp-cafe-tostado-en-grano-o-molido-ntc3534-2007.html (accessed on 20 July 2021).
- Instituto Colombiano de Normas Técnicas y Certificación. NTC 4092. 2016. Available online: https://tienda.icontec.org/gp-microbiologia-de-alimentos-y-productos-para-alimentacion-animal-requisitos-generales-y-directrices-para-analisis-microbiologicos-ntc4092-2016.html (accessed on 20 July 2021).
- Instituto Colombiano de Normas Técnicas y Certificación. NTC 4939. 2001. Available online: https://tienda.icontec.org/sectores/medio-ambiente-proteccion-de-la-salud-seguridad/proteccion-del-medio-ambiente/polucion-control-a-la-polucion-y-conservacion/gp-calidad-del-agua-enumeracion-de-coliformes-y-escherichia-coli-tecnica-con-tubos-de-fermentacion-y-tecnica-de-sustrato-enzimatico (accessed on 15 June 2021).
- Ministerio de la Protección Social Ministerio de Ambiente, Vivienda y Desarrollo Territorial. Resolución 2115. 2007. Available online: https://minvivienda.gov.co/sites/default/files/normativa/2115%20-%202007.pdf (accessed on 25 September 2021).
- Gerard, L. Caracterización de Bacterias del Ácido Acético Destinadas a la Producción de Vinagres de Frutas, Tesis Doctoral, Departamento de Tecnología de Alimentos, Universidad politecnica de Valencia. 2015. Available online: https://riunet.upv.es/bitstream/handle/10251/59401/GERARD%20- (accessed on 13 March 2022).
- Seearunruangchai, A.; Tanasupawat, S.; Keeratipibul, S.; Thawai, C.; Itoh, T.; Yamada, Y. Identification of Acetic Acid Bacteria Isolated from Fruits Collected in Thailand. J. Gen. Appl. Microbiol. 2004, 50, 47–53. [Google Scholar] [CrossRef]
- Walsh, T.J.; Hayden, R.T.; Larone, D.H. Larone’s Medically Important Fungi: A Guide to Identification, 6th ed.; American Society for Microbiology: Washington, DC, USA, 2018. [Google Scholar]
- Hunter, B. Illustrated Genera of Imperfect Fungi; The American Phytopathological Society: St. Paul, MN, USA, 1998. [Google Scholar]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Instituto Colombiano de Normas Técnicas y Certificación. NTC 2324. 2021. Available online: https://tienda.icontec.org/gp-cafe-verde-examen-olfativo-y-visual-y-determinacion-de-materia-extrana-y-defectos-ntc2324-2021.html (accessed on 26 July 2021).
- Specialty Coffee Association. Cupping Protocols. In Protocols & Best Practices; Specialty Coffee Association: Irvine, CA, USA, 2015. [Google Scholar]
- Louzada Pereira, L.; Rizzo Moreira, T. (Eds.) Quality Determinants In Coffee Production; Food Engineering Series; Springer International Publishing: Cham, Switzerland, 2021; Available online: http://link.springer.com/10.1007/978-3-030-54437-9 (accessed on 16 May 2022).
- Gänzle, M.G. Lactic Metabolism Revisited: Metabolism of Lactic Acid Bacteria in Food Fermentations and Food Spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Saichana, N.; Matsushita, K.; Adachi, O.; Frébort, I.; Frebortova, J. Acetic Acid Bacteria: A Group of Bacteria with Versatile Biotechnological Applications. Biotechnol. Adv. 2015, 33, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Masi, C.; Dinnella, C.; Monteleone, E.; Prescott, J. The Impact of Individual Variations in Taste Sensitivity on Coffee Perceptions and Preferences. Physiol. Behav. 2015, 138, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Batista, L.; Abreu, L.; Dias, E.; Schwan, R. Succession of Bacterial and Fungal Communities during Natural Coffee (Coffea arabica) Fermentation. Food Microbiol. 2008, 25, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Elhalis, H.; Cox, J.; Zhao, J. Coffee Fermentation: Expedition from Traditional to Controlled Process and Perspectives for Industrialization. Appl. Food Res. 2023, 3, 100253. [Google Scholar] [CrossRef]
- De Carvalho Neto, D.; de Melo Pereira, G.; Tanobe, V.; Thomaz Soccol, V.; da Silva, B.G.; Rodrigues, C.; Soccol, C. Yeast Diversity and Physicochemical Characteristics Associated with Coffee Bean Fermentation from the Brazilian Cerrado Mineiro Region. Fermentation 2017, 3, 11. [Google Scholar] [CrossRef]
- Mouret, J.R.; Camarasa, C.; Angenieux, M.; Aguera, E.; Perez, M.; Farines, V.; Sablayrolles, J.M. Kinetic Analysis and Gas–Liquid Balances of the Production of Fermentative Aromas during Winemaking Fermentations: Effect of Assimilable Nitrogen and Temperature. Food Res. Int. 2014, 62, 1–10. [Google Scholar] [CrossRef]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. The Crucial Role of Yeasts in the Wet Fermentation of Coffee Beans and Quality. Int. J. Food Microbiol. 2020, 333, 108796. [Google Scholar] [CrossRef]
- Feng, X.; Dong, H.; Yang, P.; Yang, R.; Lu, J.; Lv, J.; Sheng, J. Culture-Dependent and -Independent Methods to Investigate the Predominant Microorganisms Associated with Wet Processed Coffee. Curr. Microbiol. 2016, 73, 190–195. [Google Scholar] [CrossRef]
- Masoud, W.; Bjørg Cesar, L.; Jespersen, L.; Jakobsen, M. Yeast Involved in Fermentation Of Coffea arabica in East Africa Determined by Genotyping and by Direct Denaturating Gradient Gel Electrophoresis. Yeast 2004, 21, 549–556. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; Soccol, V.T.; Pandey, A.; Medeiros, A.B.P.; Andrade Lara, J.M.R.; Gollo, A.L.; Soccol, C.R. Isolation, Selection and Evaluation of Yeasts for Use in Fermentation of Coffee Beans by the Wet Process. Int. J. Food Microbiol. 2014, 188, 60–66. [Google Scholar] [CrossRef]
- Iamanaka, B.T.; Teixeira, A.A.; Teixeira, A.R.R.; Copetti, M.V.; Bragagnolo, N.; Taniwaki, M.H. The Mycobiota of Coffee Beans and Its Influence on the Coffee Beverage. Food Res. Int. 2014, 62, 353–358. [Google Scholar] [CrossRef]
- Bessaire, T.; Perrin, I.; Tarres, A.; Bebius, A.; Reding, F.; Theurillat, V. Mycotoxins in Green Coffee: Occurrence and Risk Assessment. Food Control 2019, 96, 59–67. [Google Scholar] [CrossRef]
- Pino, A.F.S.; Espinosa, Z.Y.D.; Cabrera, E.V.R. Characterization of the Rhizosphere Bacterial Microbiome and Coffee Bean Fermentation in the Castillo-Tambo and Bourbon Varieties in the Popayán-Colombia Plateau. BMC Plant Biol. 2023, 23, 217. [Google Scholar] [CrossRef]
- de Sousa, L.P.; Guerreiro-Filho, O.; Mondego, J.M.C. The Rhizosphere Microbiomes of Five Species of Coffee Trees. Microbiol. Spectr. 2022, 10, e00444-22. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.E.; da Luz, J.M.R.; da Silva Oliveira, E.C.; Guarçoni, R.C.; Moreira, T.R.; Moreli, A.P.; Siqueira, E.A.; de Cássia Soares da Silva, M.; Costa, M.R.G.F.; Pereira, L.L. Chemical Profile and Sensory Perception of Coffee Produced in Agroforestry Management. Eur. Food Res. Technol. 2023, 249, 1479–1489. [Google Scholar] [CrossRef]
- de Sousa, L.H.B.P.; da Luz, J.M.R.; da Silva, M.d.C.S.; Moreli, A.P.; Veloso, T.G.R.; Guarçoni, R.C.; Moreira, T.R.; Barros, M.V.P.; Kasuya, M.C.M.; Marcate, J.P.P.; et al. Relationship between Sensory and Microbial Profiles of Fermented Coffee by Dry and Washed Methods. Food Chem. Adv. 2023, 2, 100259. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Time (h) | Temperature (°C) | pH | °Brix (°Bx) | Total Acidity (mg/L CaCO3) |
|---|---|---|---|---|---|
| Var. Tabi | 0 | 25 ± 1.50 | 4.05 ± 0.16 | 4.00 ± 0.01 | 315.07 ± 15.27 |
| 9 | 28 ± 1.04 | 3.83 ± 0.15 | 2.00 ± 0.01 | 405.07 ± 11.54 | |
| 18 | 29 ± 0.50 | 3.83 ± 0.16 | 4.83 ± 0.28 | 1015.07 ± 28.86 | |
| Var. Castillo General | 0 | 24 ± 1.50 | 4.38 ± 0.54 | 5.00 ± 0.01 | 188.40 ± 17.32 |
| 9 | 27 ± 0.76 | 4.00 ± 0.22 | 5.00 ± 0.50 | 448.40 ± 17.32 | |
| 18 | 26 ± 1.00 | 3.56 ± 0.06 | 4.43 ± 0.40 | 748.40 ± 50.00 | |
| Var. Colombia | 0 | 24 ± 0.50 | 4.44 ± 0.65 | 5.17 ± 0.28 | 198.40 ± 86.60 |
| 9 | 25 ± 0.50 | 4.13 ± 0.18 | 6.83 ± 0.28 | 288.40 ± 17.32 | |
| 18 | 26 ± 0.50 | 3.49 ± 0.07 | 6.00 ± 0.01 | 655.07 ± 211.26 |
| Sample | Time (h) | Sucrose | Glucose | Fructose | Protein | ||||
|---|---|---|---|---|---|---|---|---|---|
| M | S | M | S | M | S | M | S | ||
| % | |||||||||
| Var. Tabi | 0 | 0.04 ± 0.01 | 8.13 ± 0.30 | 0.85 ± 0.32 | ND | 1.43 ± 0.38 | 0.23 ± 0.01 | 10.31 ± 1.84 | 13.94 ± 0.01 |
| 9 | ND | 7.52 ± 0.22 | 0.20 ± 0.11 | 1.32 ± 0.38 | 0.67 ± 0.24 | 0.22 ± 0.01 | 14.00 ± 1.15 | 13.94 ± 0.05 | |
| 18 | 0.02 ± 0.01 | 7.24 ± 0.13 | 0.07 ± 0.05 | 1.39 ± 0.03 | 0.51 ± 0.07 | 0.22 ± 0.01 | 14.56 ± 0.28 | 14.13 ± 0.09 | |
| Var. Castillo General | 0 | 0.17 ± 0.13 | 7.29 ± 0.39 | 0.40 ± 0.04 | 1.42 ± 0.04 | 1.58 ± 0.15 | 0.17 ± 0.09 | 9.19 ± 1.21 | 14.69 ± 0.09 |
| 9 | 0.44 ± 0.19 | 8.08 ± 0.22 | 1.37 ± 0.02 | 1.51 ± 0.03 | 1.88 ± 0.08 | 0.36 ± 0.05 | 11.63 ± 0.61 | 14.50 ± 0.04 | |
| 18 | 0.05 ± 0.01 | 7.30 ± 0.38 | 0.83 ± 0.26 | 1.39 ± 0.06 | 1.87 ± 0.01 | 0.13 ± 0.11 | 10.63 ± 0.50 | 14.63 ± 0.06 | |
| Var. Colombia | 0 | 0.07 ± 0.03 | 6.87 ± 0.08 | 2.43 ± 0.43 | 1.25 ± 0.01 | 2.93 ± 0.48 | 0.23 ± 0.08 | 11.19 ± 0.56 | 14.94 ± 0.25 |
| 9 | ND | 6.70 ± 0.50 | 1.55 ± 0.30 | 1.25 ± 0.03 | 1.97 ± 0.39 | 0.06 ± 0.04 | 10.06 ± 0.69 | 14.44 ± 0.13 | |
| 18 | ND | 8.54 ± 0.91 | 0.31 ± 0.08 | 1.38 ± 0.06 | 1.44 ± 0.26 | 0.15 ± 0.04 | 12.81 ± 1.37 | 14.56 ± 0.06 | |
| Sample | Time (h) | Microbial Counts | ||||
|---|---|---|---|---|---|---|
| Mesophiles | Coliforms | Acid Lactic Bacteria | Yeasts | Mycelial Fungi | ||
| Log10 cfu/mL | ||||||
| Var. Tabi | 0 | 5.89 | 3.60 | 6.78 | 5.77 | 3.30 |
| 9 | 5.61 | 3.48 | 6.45 | 5.58 | 3.00 | |
| 18 | 5.47 | 3.00 | 4.58 | 5.48 | 3.48 | |
| Var. Castillo General | 0 | 5.90 | 3.48 | 6.77 | 5.91 | 3.30 |
| 9 | 5.32 | 3.30 | 4.07 | 5.24 | 3.00 | |
| 18 | 5.48 | 3.00 | 6.30 | 5.38 | 3.30 | |
| Var. Colombia | 0 | 5.92 | 4.00 | 6.76 | 5.44 | 3.48 |
| 9 | 5.66 | 3.60 | 4.04 | 5.36 | 3.00 | |
| 18 | 5.57 | 3.30 | 6.38 | 5.24 | 3.30 | |
| Sample | Time (h) | Mesophilic Bacteria | * LAB | ** AAB | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enterobacter agglomerans | Enterobacter aerogenes | Enterobacter cloacae | Shigella spp. | Staphylococcus spp. | Bacillus spp. | Bacillus firmus | Klebsiella spp. | Klebsiella ozaenae | Escherichia coli | Micrococcus spp. | Citrobacter freudii | Cedecea lapagei | Averyella spp. | Leuconostoc mesenteroides | Leuconostoc mesenteroides spp. cremoris | Lactiplantibacillus pentosus | Lactiplantibacillus plantarum | Lactobacillus delbrueckii | Levilactobacillus brevis | Lactococcus spp. | Gluconobacter spp. | ||
| Var. Tabi | 0 | ||||||||||||||||||||||
| 9 | |||||||||||||||||||||||
| 18 | |||||||||||||||||||||||
| Var. Castillo General | 0 | ||||||||||||||||||||||
| 9 | |||||||||||||||||||||||
| 18 | |||||||||||||||||||||||
| Var. Colombia | 0 | ||||||||||||||||||||||
| 9 | |||||||||||||||||||||||
| 18 | |||||||||||||||||||||||
| Sample | Timee (h) | Yeast | Mycelial Fungi | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sacharomyces cerevisiae | Cryptococcus laurentii | Cryptococcus neoformans | Pichia sp. | Rhodotorula sp. | Candida rugosa | Candida krusei | Candida parapsilosis | Trichoderma sp. | Geotrichum sp. | Penicillium sp. | ||
| Var. Tabi | 0 | |||||||||||
| 9 | ||||||||||||
| 18 | ||||||||||||
| Var. Castillo General | 0 | |||||||||||
| 9 | ||||||||||||
| 18 | ||||||||||||
| Var. Colombia | 0 | |||||||||||
| 9 | ||||||||||||
| 18 | ||||||||||||
| Sample | Time (h) | Sobs * | Richness | Diversity | ||
|---|---|---|---|---|---|---|
| Ace | Chao1 | Simpson | Shannon | |||
| Var. Tabi | 0 | 266 | 420.170 | 3.958 | 2.677 | 0.879 |
| 9 | 224 | 366.067 | 3.711 | 2.196 | 0.820 | |
| 18 | 177 | 259.307 | 2.646 | 2.062 | 0.804 | |
| Var. Castillo G | 0 | 570 | 1516.287 | 13.276 | 3.292 | 0.894 |
| 9 | 194 | 318.577 | 3.130 | 2.113 | 0.811 | |
| 18 | 172 | 355.041 | 3.627 | 2.298 | 0.852 | |
| Var. Colombia | 0 | 314 | 505.159 | 4.958 | 1.862 | 0.589 |
| 9 | 238 | 698.933 | 5.579 | 1.931 | 0.754 | |
| 18 | 183 | 361.125 | 3.771 | 2.295 | 0.850 | |
| Sample | Time (h) | Sobs * | Richness | Diversity | ||
|---|---|---|---|---|---|---|
| Ace | Chao 1 | Simpson | Shannon | |||
| Var. Tabi | 0 | 574 | 446.570 | 4.653 | 2.167 | 0.782 |
| 9 | 436 | 286.931 | 2.644 | 1.700 | 0.685 | |
| 18 | 419 | 383.130 | 3.418 | 1.684 | 0.685 | |
| Var. Castillo G | 0 | 543 | 1756.051 | 29.155 | 2.519 | 0.849 |
| 9 | 227 | 577.207 | 4.810 | 2.406 | 0.842 | |
| 18 | 215 | 529.321 | 4.356 | 1.445 | 0.612 | |
| Var. Colombia | 0 | 577 | 389.022 | 3.703 | 1.858 | 0.679 |
| 9 | 524 | 359.125 | 3.017 | 2.028 | 0.768 | |
| 18 | 320 | 212.395 | 2.074 | 1.316 | 0.601 | |
| Sample | Time (h) | SCA | Sensory Description |
|---|---|---|---|
| Var. Tabi | 0 | 82.88 | Chocolat. Raw sugarcane |
| 9 | 83.25 | Raw sugarcane. Floral | |
| 18 | 82.74 | Apple. Caramel | |
| Var. Castillo General | 0 | 53.63 | Stored |
| 9 | 79.50 | Herbal. Cereal | |
| 18 | 52.00 | Stored | |
| Var. Colombia | 0 | 82.25 | Herbal. Raw sugarcane. Caramel |
| 9 | 82.13 | Raw sugarcane. Herbal | |
| 18 | 82.50 | Nuts |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holguín-Sterling, L.; Pedraza-Claros, B.; Pérez-Salinas, R.; Ortiz, A.; Navarro-Escalante, L.; Góngora, C.E. Physical–Chemical and Metataxonomic Characterization of the Microbial Communities Present during the Fermentation of Three Varieties of Coffee from Colombia and Their Sensory Qualities. Agriculture 2023, 13, 1980. https://doi.org/10.3390/agriculture13101980
Holguín-Sterling L, Pedraza-Claros B, Pérez-Salinas R, Ortiz A, Navarro-Escalante L, Góngora CE. Physical–Chemical and Metataxonomic Characterization of the Microbial Communities Present during the Fermentation of Three Varieties of Coffee from Colombia and Their Sensory Qualities. Agriculture. 2023; 13(10):1980. https://doi.org/10.3390/agriculture13101980
Chicago/Turabian StyleHolguín-Sterling, Laura, Bertilda Pedraza-Claros, Rosangela Pérez-Salinas, Aristófeles Ortiz, Lucio Navarro-Escalante, and Carmenza E. Góngora. 2023. "Physical–Chemical and Metataxonomic Characterization of the Microbial Communities Present during the Fermentation of Three Varieties of Coffee from Colombia and Their Sensory Qualities" Agriculture 13, no. 10: 1980. https://doi.org/10.3390/agriculture13101980
APA StyleHolguín-Sterling, L., Pedraza-Claros, B., Pérez-Salinas, R., Ortiz, A., Navarro-Escalante, L., & Góngora, C. E. (2023). Physical–Chemical and Metataxonomic Characterization of the Microbial Communities Present during the Fermentation of Three Varieties of Coffee from Colombia and Their Sensory Qualities. Agriculture, 13(10), 1980. https://doi.org/10.3390/agriculture13101980