
Survival Dynamics of Trichoderma longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils with Different Moisture Levels
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Soil
2.2. Preparation of Conidia Powder of T. longibrachiatum Tr58
2.3. Preparation of Chlamydospore Powder of T. longibrachiatum Tr58
2.4. Effect of Soil with Different Moisture Contents on Spore Germination of T. longibrachiatum Tr58
2.5. Survival Dynamics of T. longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils
2.6. Experimental Design and Statistical Analysis
3. Results
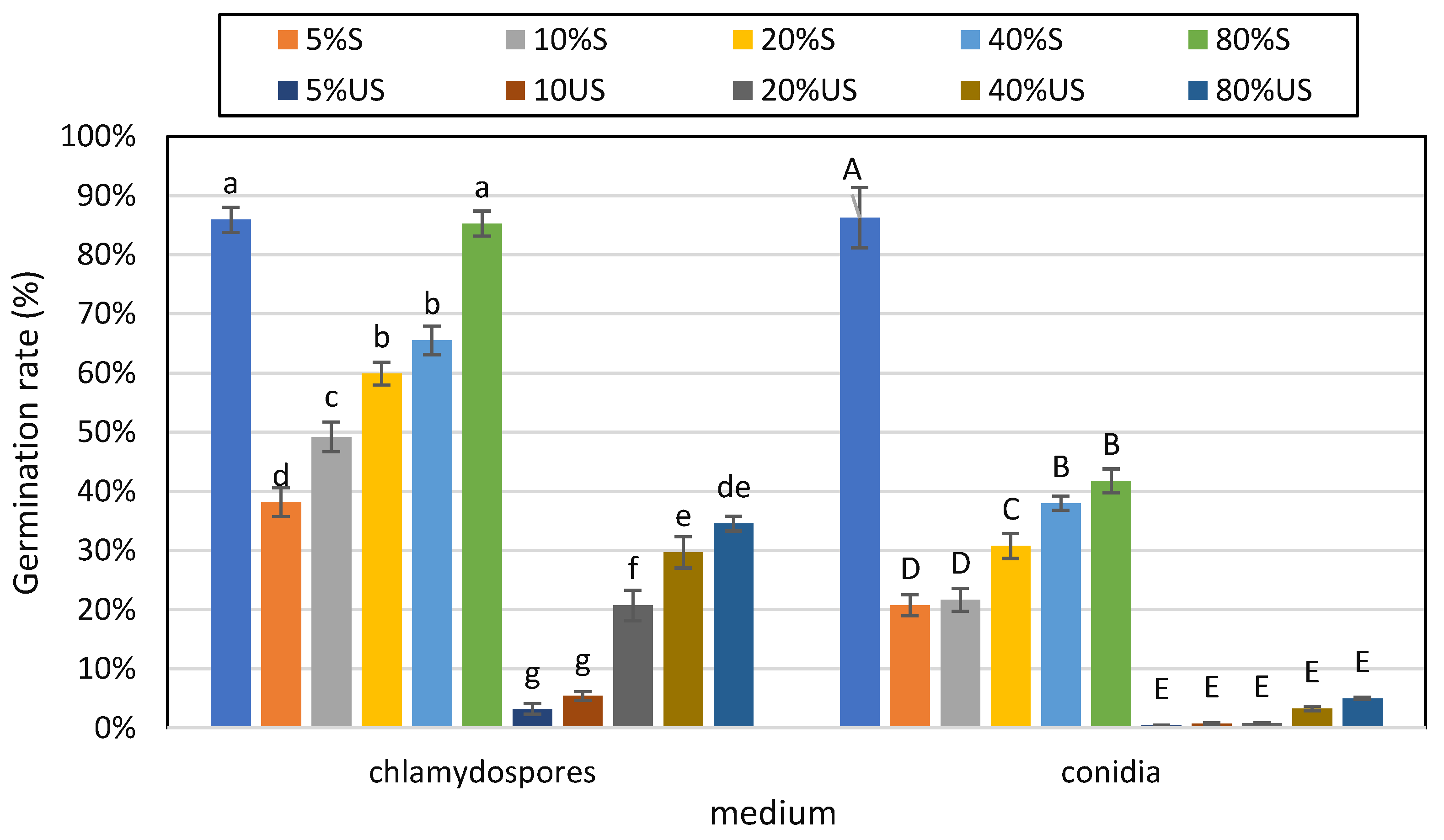
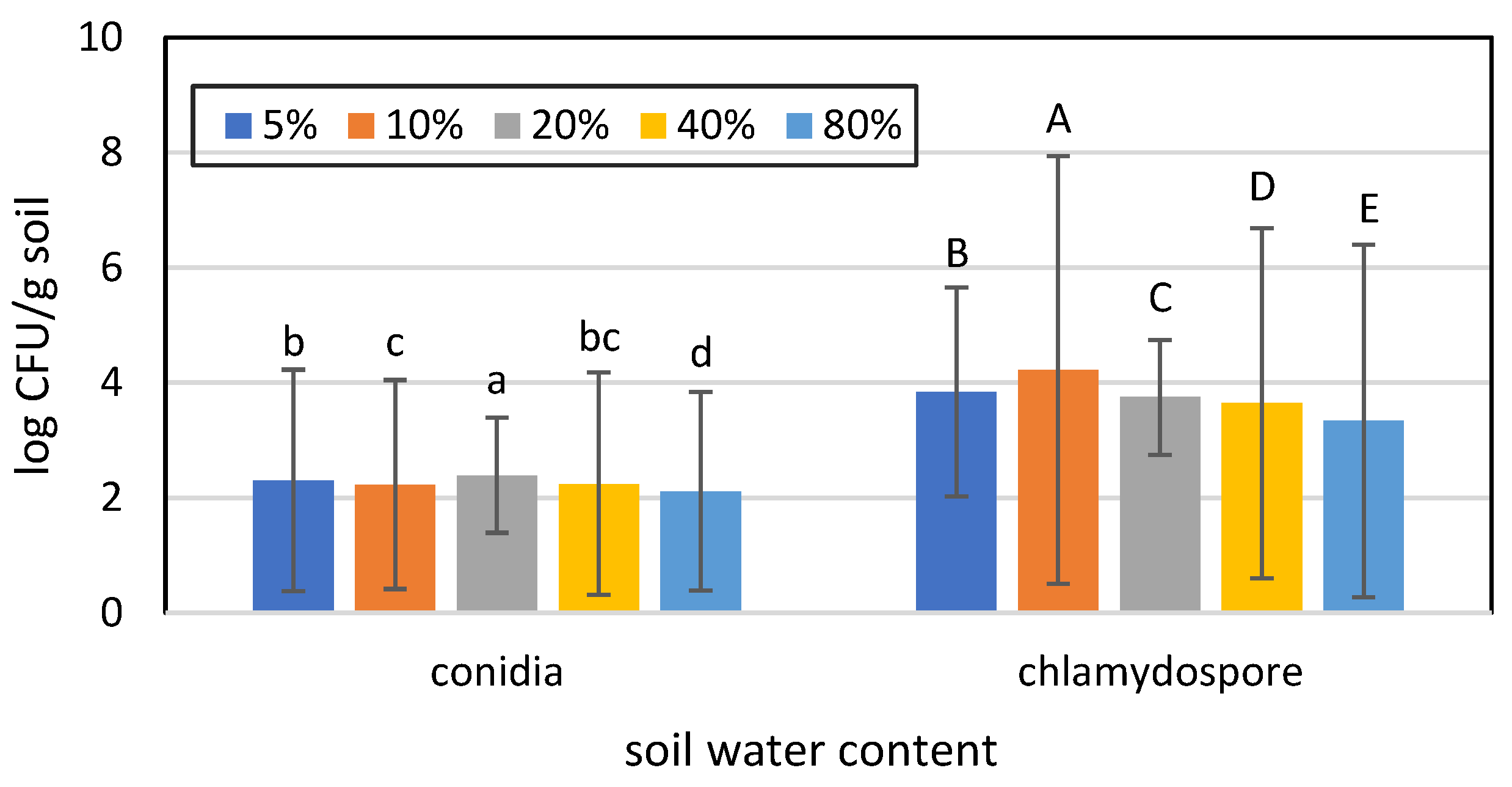
3.1. Effects of Soil Moisture Content and Sterilization on the Germination of Conidia and Chlamydospores of T. longibrachiatum Tr58
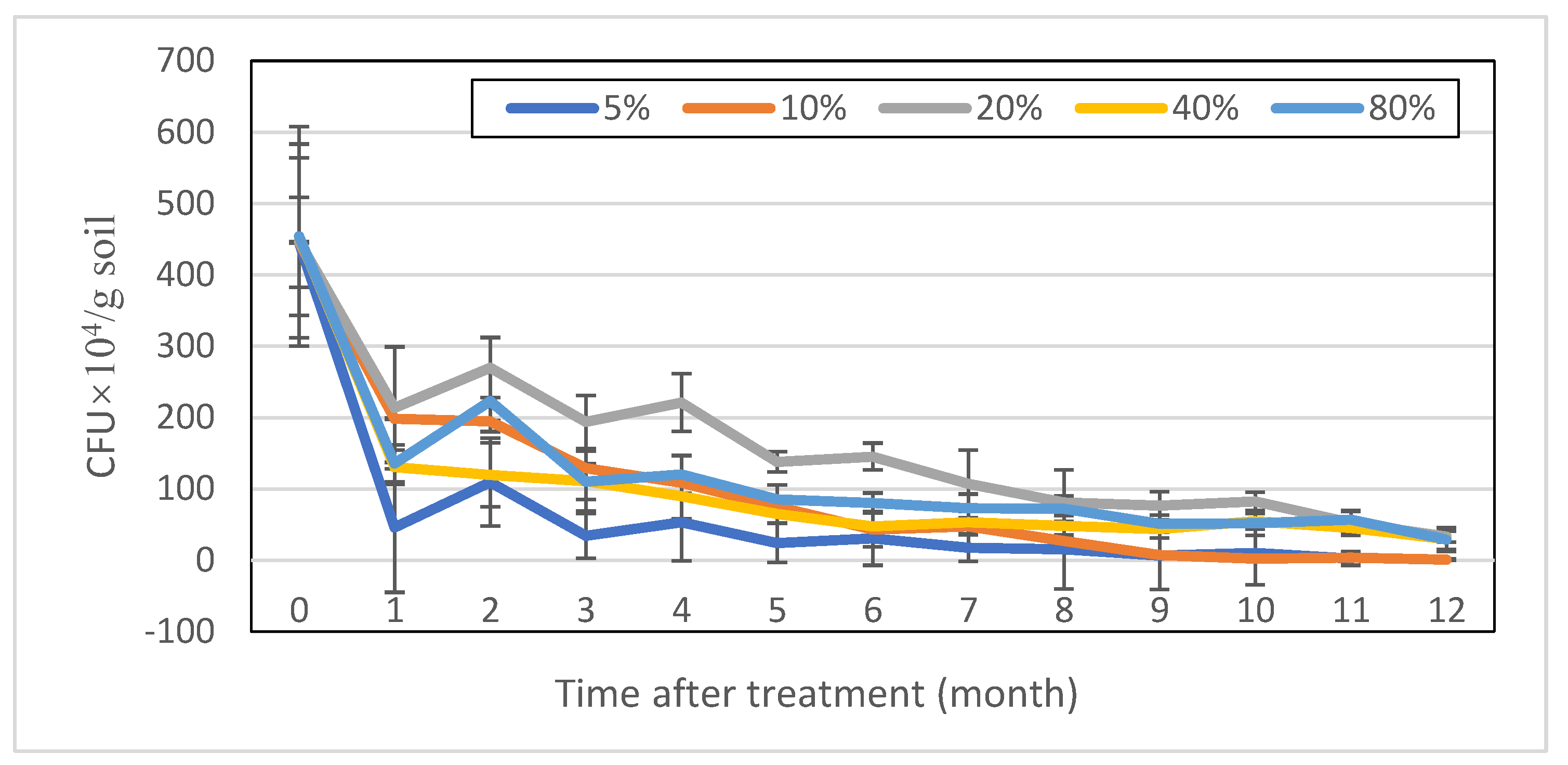
3.2. Survival Dynamics of T. longibrachiatum Tr58 in Conidia-Amended Soils with Different Moisture Contents
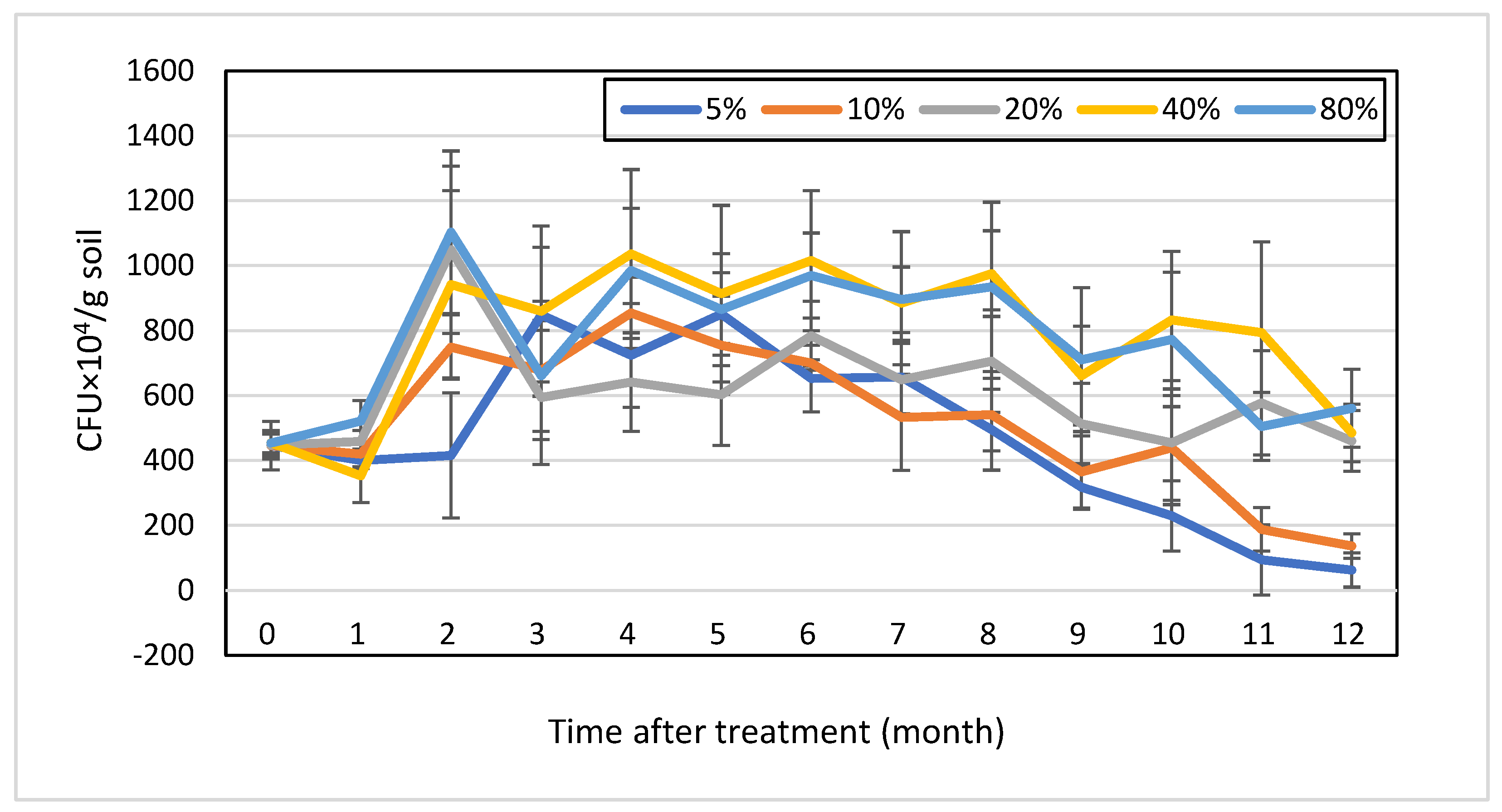
3.3. Survival Dynamics of T. longibrachiatum Tr58 in Chlamydospore-Amended Soils with Different Moisture Contents
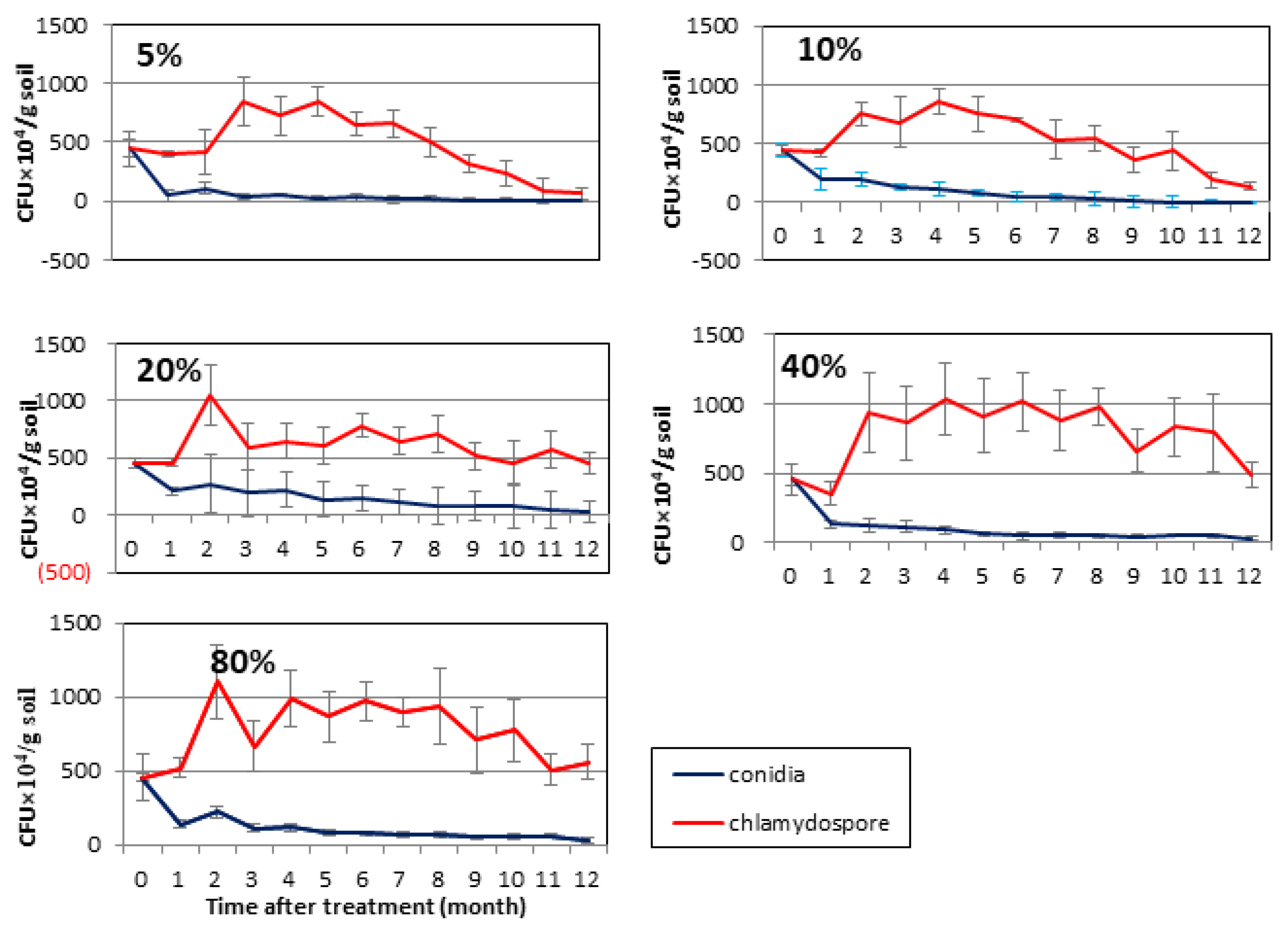
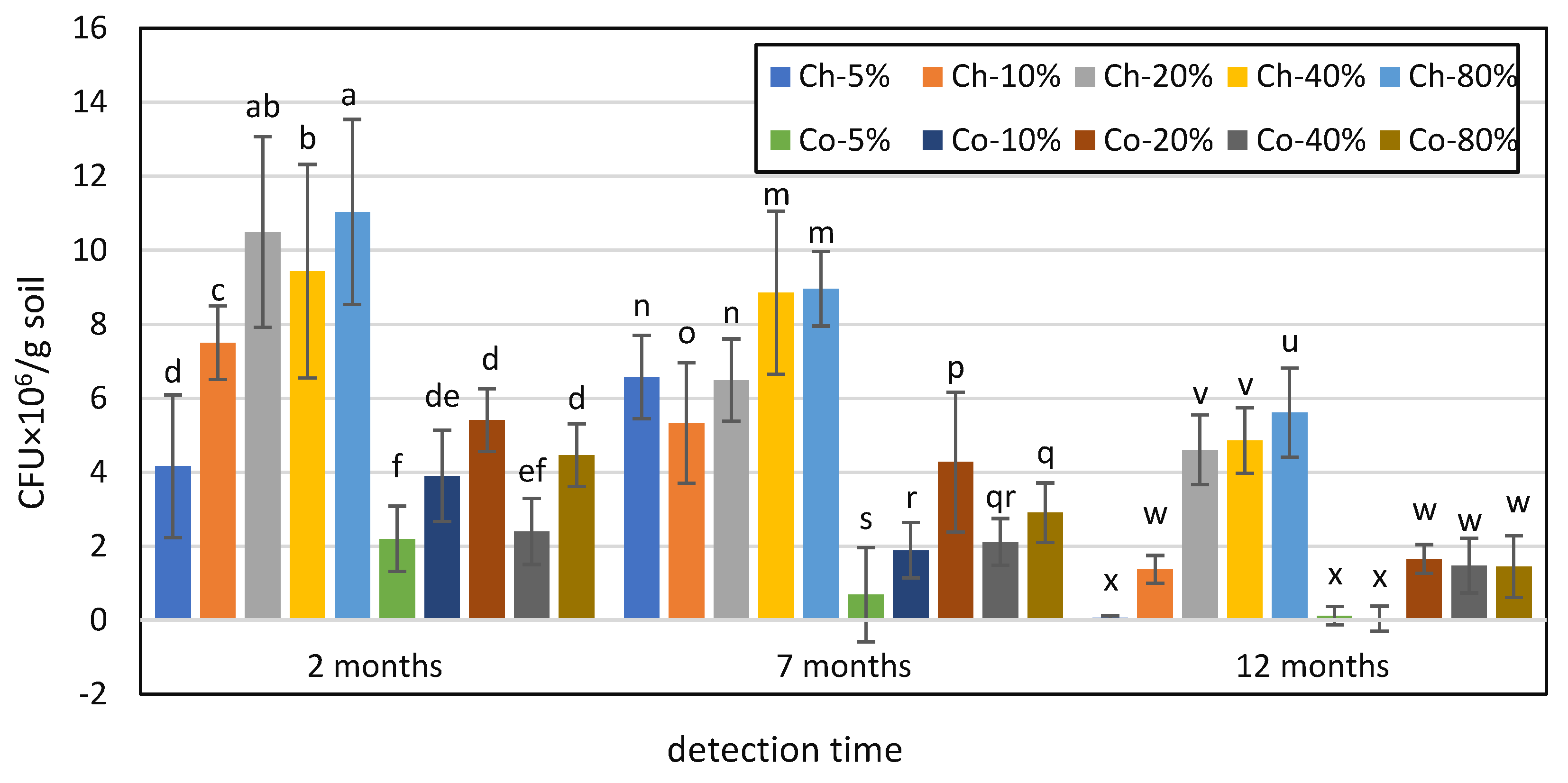
3.4. Comparison of Survival Dynamics of T. longibrachiatum Tr58 between Conidia- and Chlamydospore-Amended Soils with Different Moisture Contents
3.5. Comparison of the T. longibrachiatum Tr58 Content in Conidia- and Chlamydospore-amended Soils with Different Moisture Contents after 24 Months
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Cummings, N.J.; Ambrose, A.; Braithwaite, M.; Bissett, J.; Roslan, H.A.; Abdullah, J.; Stewart, A.; Agbayani, F.V.; Steyaert, J.; Hill, R.A. Diversity of root-endophytic Trichoderma from Malaysian Borneo. Mycol. Progress 2016, 15, 50. [Google Scholar] [CrossRef]
- Mukherjee, M.; Dasgupta, B. Survival potential of Trichoderma harzianum in different mass multiplication media. Environ. Ecol. 2006, 24, 737–741. [Google Scholar]
- Mukherjee, P.K.; Horwitz, B.A.; Herrera-Estrella, A.; Schmoll, M.; Kenerley, C.M. Trichoderma research in the genome Era. Annu. Rev. Phytopathol. 2013, 51, 105–129. [Google Scholar] [CrossRef]
- Prabhakaran, N.; Prameeladevi, T.; Sathiyabama, M.; Kamil, D. Screening of different Trichoderma species against agriculturally important foliar plant pathogens. J. Environ. Biol. 2015, 36, 191–198. [Google Scholar] [PubMed]
- Liu, H.; Duan, W.; Liu, C.; Meng, L.; Li, H.; Li, R.; Shen, Q. Spore production in the solid-state fermentation of stevia residue by Trichoderma guizhouense and its effects on corn growth. J. Integr. Agr. 2021, 20, 1147–1156. [Google Scholar] [CrossRef]
- Lewis, J.A.; Papavizas, G.C. Production of chlamydospores and conidia by Trichoderma spp. in liquid and solid growth media. Soil Boil. Biochem. 1983, 15, 351–357. [Google Scholar] [CrossRef]
- Tian, B.; Qu, Q.; Zhang, K.Q.; Li, L. Induction of chlamydospores in Trichoderma harzianum and Gliocladium roseum by antifungal compounds produced by Bacillus subtilis C2. J. Phytopathol. 2005, 153, 686–693. [Google Scholar]
- Staib, P.; Morschhäuser, J. Chlamydospore formation in Candida albicans and Candida dubliniensis–an enigmatic developmental programme. Mycoses 2007, 50, 1–12. [Google Scholar] [CrossRef]
- Darmono, T.W.; Parke, J.L. Chlamydospores of Phytophthora cactorum: Their production, structure, and infectivity. Can. J. Bot. 1990, 68, 640–645. [Google Scholar] [CrossRef]
- Lewis, J.A.; Papavizas, G.C. Chlamydospore formation by Trichoderma spp. in natural substrates. Can. J. Microbiol. 1984, 30, 1–7. [Google Scholar] [CrossRef]
- Papavizas, G.C. Trichoderma and Gliocladium: Biology, ecology, and potential for biocontrol. Ann. Rev. Phytopathol. 1985, 23, 23–54. [Google Scholar] [CrossRef]
- Pan, W.; Mu, C.; Jiang, X.; Tian, Y.; Zhu, C. Chlamydospore and conidia of Trichoderma and soil fungistasis. Chinese J. Biol. Control 2006, 22, 87–91. [Google Scholar]
- Bonanomi, G.; Gaglione, S.A.; Incerti, G.; Zoina, A. Biochemical quality of organic amendments affects soil fungistasis. Appl. Soil Ecol. 2013, 72, 135–142. [Google Scholar] [CrossRef]
- Garbeva, P.; Hol, W.H.G.; Termorshuizen, A.J.; Kowalchuk, G.A.; de Boer, W. Fungistasis and general soil biostasis—A new synthesis. Soil. Boil. Biochem. 2011, 43, 469–477. [Google Scholar] [CrossRef]
- Zhou, W.; Mo, M. Soil fungistasis on the spore germination of fungi. J. Yunnan Univ. Nat. Sci. Ed. 2002, 24, 312–315. [Google Scholar]
- Beagle-Ristaino, J.E.; Papavizas, G.C. Survival and proliferation of propagules of Trichoderma spp. and Gliocladium virens in soil and in plant rhizospheres. Phytopathology 1985, 75, 729–732. [Google Scholar] [CrossRef]
- Mishra, A.; Sharma, S.D.; Patel, S.L. Influence of fertilizers on the growth and spore germination of Trichoderma harzianum rifai. Agric. Sci. Digest. 2003, 23, 285–287. [Google Scholar]
- Yang, X.; Li, M.; Zhang, L.; Pang, L.; Sun, Q.; Jiang, X. Transcriptome analysis of Trichoderma harzianum Th-33 in chlamydospore formation. Chinese, J. Biol. Control 2015, 31, 85–95. [Google Scholar]
- Peng, X.; Wu, B.; Zhang, S.; Li, M.; Jiang, X. Transcriptome dynamics underlying chlamydospore formation in Trichoderma virens GV29-8. Front. Microbiol. 2021, 2, 654855. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, X.; Yang, X.; Li, M.; Chen, S. Inhibition of chlamydospore germination and mycelial growth of Trichoderma spp. by chemical fungicides. Agric. Sci. Technol. 2015, 16, 1494–1499. [Google Scholar]
- Liu, Z.; Sun, Y.; Zhang, X.; Chen, B.; Chen, J. Effect of Trichoderma chlamydospores preparation on amount of soil microorganism and cotton Verticillium wilt. Xinjiang Agric. Sci. 2015, 52, 97–101. [Google Scholar]
- Lian, H.; Chen, Y.; Li, M.; Liang, X.; Ma, G. The effect of Trichoderma conidia and chlamydospores on physiological characteristics of cucumber seedlings and control effect against Fusarium wilt. Agric. Res. Arid Areas 2021, 39, 227–235. [Google Scholar]
- Cavalcante, R.S.; Lima, H.L.S. Effect of moisture on Trichoderma conidia production on corn and wheat bran by solid state fermentation. Food Bioprocess Tech. 2008, 1, 100–104. [Google Scholar] [CrossRef]
- Griffin, D.M. Soil moisture and the ecology of soil fungi. Biol. Rev. 1963, 38, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cao, Q.; Lian, H.; Ma, G.; Jiang, X.; Li, M. Screening of Trichoderma and their efficiency against sunflower Sclerotinia rot. Chinese J. Biol. Control 2022, 38, 846–851. [Google Scholar]
- Sun, M.; Liu, X.; Tang, L. Fungistatic effect of soils on nematophagous fungi and their preparations. Mycosystema 1997, 2, 149–154. [Google Scholar]
- Redda, E.T.; Ma, J.; Mei, J.; Li, M.; Wu, B.; Jiang, X. Antagonistic potential of different isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. Eur. Exp. Biol. 2018, 8, 12. [Google Scholar]
- Dobbs, C.G.; Hinson, W.H.; Anderson, T.H.A. widespread fungistasis in soils. Nature 1953, 172, 197–199. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, H.; Li, X.; Zhang, Y.; Su, Z.; Zhang, C. Soil fungistasis and its relations to soil microbial composition and diversity: A case study of a series of soils with different fungistasis. J. Environ. Sci. 2008, 20, 871–877. [Google Scholar] [CrossRef]
- De Boer, W.; Verheggen, P.; Klein Gunnewiek, P.J.A.; Kowalchuk, G.A.; Van Veen, J.A. Microbial community composition affects soil fungistasis. Appl. Environ. Microbiol. 2003, 69, 835–844. [Google Scholar] [CrossRef]
- Lockwood, J.L. Fungistasis in soil. Biol. Rev. 1977, 52, 1–43. [Google Scholar] [CrossRef]
- Huang, Y.; Mo, M.; Zhang, K. Annulment and mechanism analysis of soil fungistasis. Southwest China J. Agric. Sci. 2006, 19, 206–210. [Google Scholar]
- Gyeltshen, J.; Dunstan, W.A.; Grigg, A.H.; Burgess, T.I.; St. J. Hardy, G.E.; Woodward, S.; Woodward, S. The influence of time, soil moisture and exogenous factors on the survival potential of oospores and chlamydospores of Phytophthora cinnamomic. Forest Pathol. 2021, 51, e12637. [Google Scholar] [CrossRef]
- Komy, M.; Saleh, A.A.; Anas, E.; Molan, Y.Y. Characterization of novel Trichoderma asperellum isolates to select effective biocontrol agents against tomato Fusarium wilt. Plant Pathol. J. 2015, 31, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.K.; Kumar, M.; Singh, N. Evaluation of fungitoxicants and Trichoderma against Macrophomina phaseolina causing root rot of cotton. Plant Dis. Res. 2021, 36, 39–44. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Z.; Zhang, Y.; Wang, Y.; Liu, Z. Biocontrol and growth-promoting effect of Trichoderma asperellum TaspHu1 isolate from Juglans mandshurica rhizosphere soil. Microbiol. Res. 2021, 242, 126596. [Google Scholar] [CrossRef]
- Li, M.; Ma, G.; Lian, H.; Su, X.; Tian, Y.; Huang, W.; Mei, J.; Jiang, X. The effects of Trichoderma on preventing cucumber fusarium wilt and regulating cucumber physiology. J. Integr. Agr. 2019, 18, 607–617. [Google Scholar] [CrossRef]
- Sanogo, S.; Pomella, A.; Hebbar, P.K.; Bailey, B.; Costa, J.C.B.; Samuels, G.J.; Lumsden, R.D. Production and germination of conidia of Trichoderma stromaticum, a mycoparasite of Crinipellis perniciosa on cacao. Phytopathology 2002, 10, 1032–1037. [Google Scholar] [CrossRef]
- Tamm, L. Influence of temperature and moisture on growth, spore production, and conidial germination of Monilinia laxa. Phytopathology 1993, 83, 1321–1326. [Google Scholar] [CrossRef]
- Dong, P.P.; Sun, M.H.; Li, S.; Peng, Y.; Luo, M. Biological characteristics of chlamydospores of Clonostachys rosea 67-1. Mycosystema 2014, 33, 1242–1252. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Q.; Liang, Y.; Tian, Y.; Lian, H.; Jiang, X.; Li, M. Survival Dynamics of Trichoderma longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils with Different Moisture Levels. Agriculture 2023, 13, 238. https://doi.org/10.3390/agriculture13020238
Cao Q, Liang Y, Tian Y, Lian H, Jiang X, Li M. Survival Dynamics of Trichoderma longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils with Different Moisture Levels. Agriculture. 2023; 13(2):238. https://doi.org/10.3390/agriculture13020238
Chicago/Turabian StyleCao, Qiulin, Yao Liang, Ying Tian, Hua Lian, Xiliang Jiang, and Mei Li. 2023. "Survival Dynamics of Trichoderma longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils with Different Moisture Levels" Agriculture 13, no. 2: 238. https://doi.org/10.3390/agriculture13020238
APA StyleCao, Q., Liang, Y., Tian, Y., Lian, H., Jiang, X., & Li, M. (2023). Survival Dynamics of Trichoderma longibrachiatum Tr58 in Conidia- and Chlamydospore-Amended Soils with Different Moisture Levels. Agriculture, 13(2), 238. https://doi.org/10.3390/agriculture13020238