
Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata)

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Genetic Material
2.2. Field Study Area
2.3. Field Establishment
2.4. Postharvest Study Area and Sampling
2.4.1. Dry Matter and Moisture Content
2.4.2. Extraction and Quantification of Non-Structural Sugars
2.4.3. Total Sugar and Starch Estimation
2.4.4. Reducing and Non-Reducing Sugars Estimation
2.4.5. Amylose and Amylopectin
3. Results
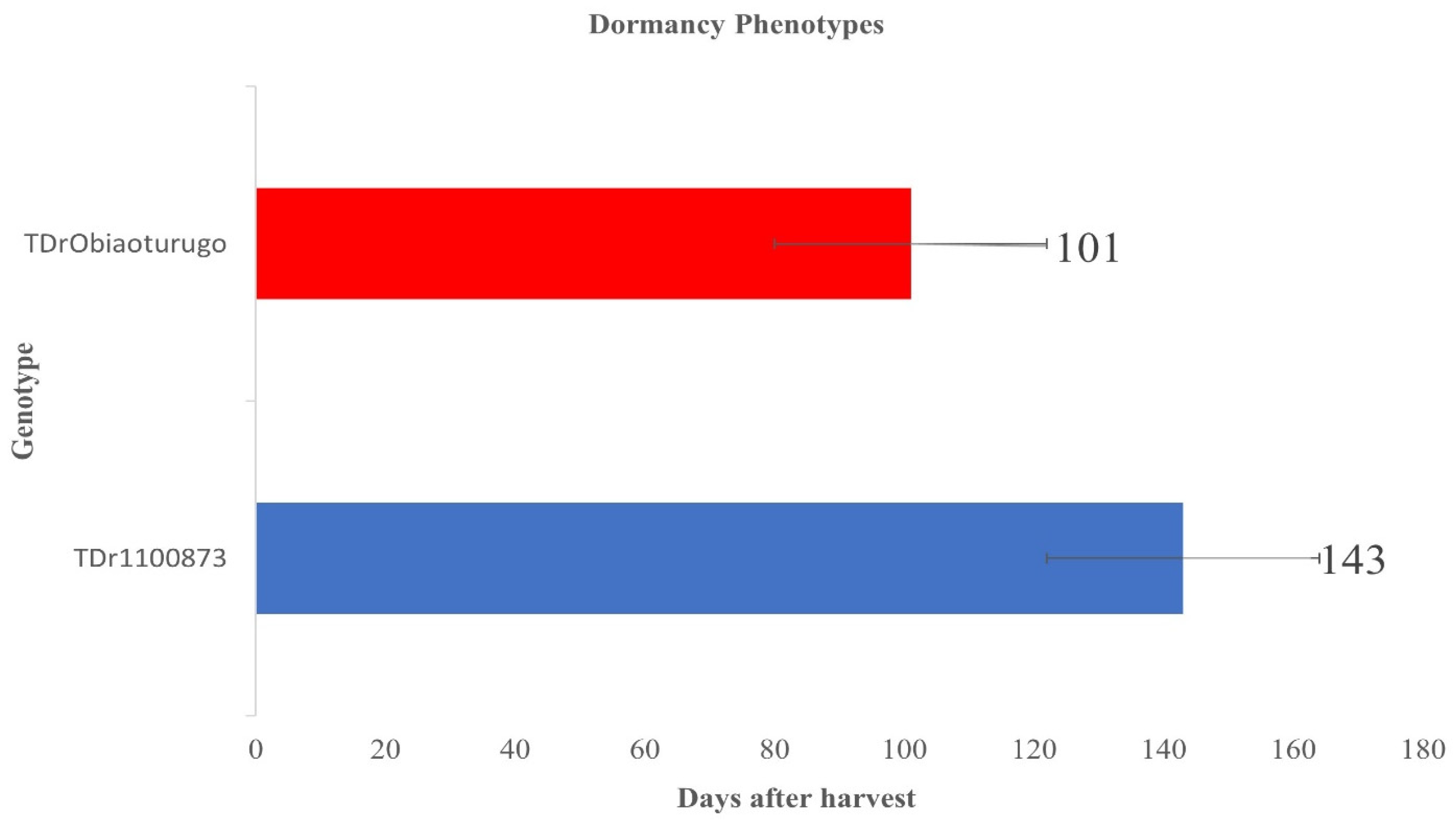
3.1. Tuber Dormancy Duration
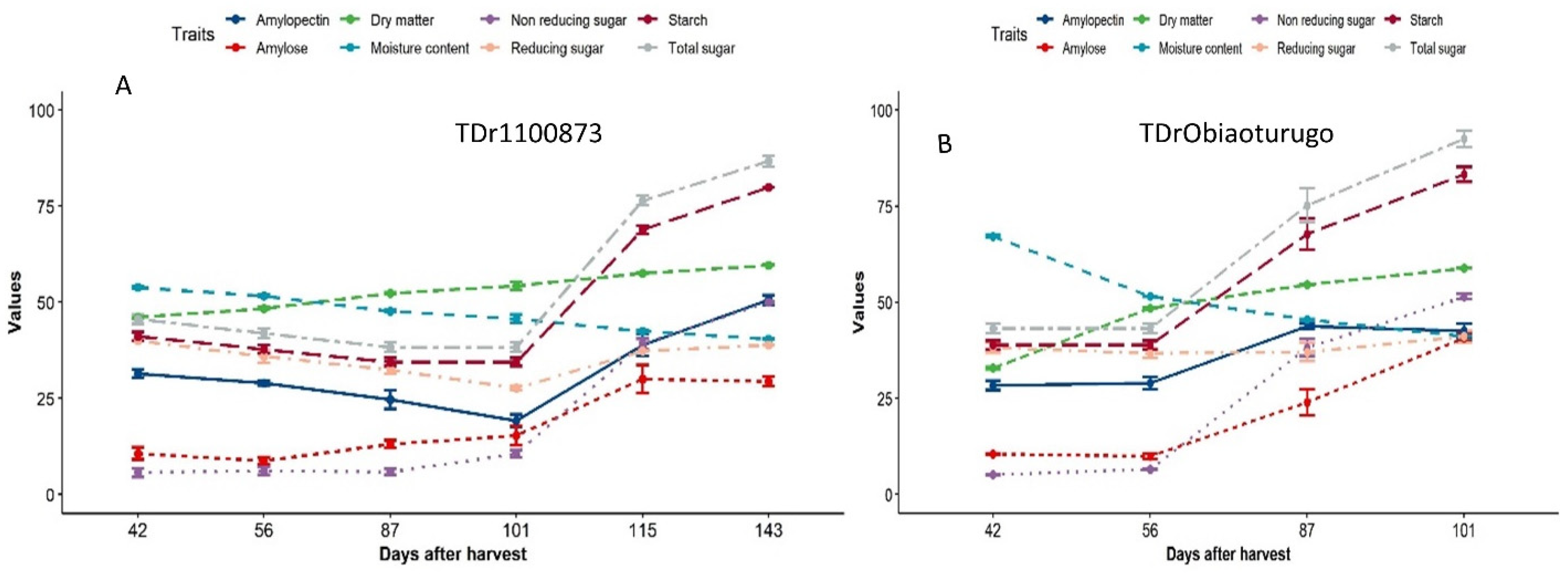
3.2. An Overview Changes in the Metabolic Status of Non-Structural Sugar, Dry Matter and Moisture Content of White Yam Tubers during Dormancy
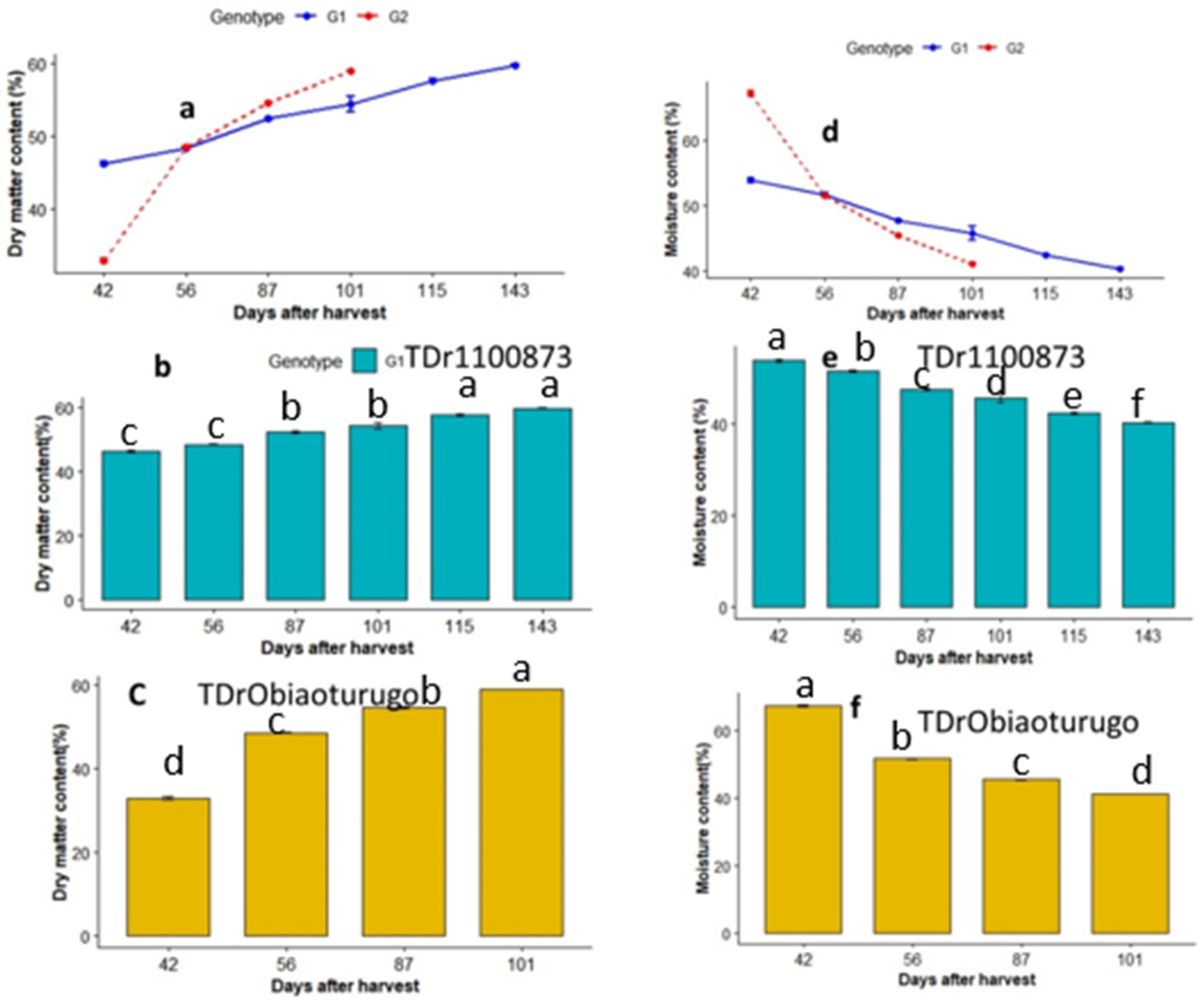
3.3. Dry Matter and Moisture Content
3.4. Metabolic Status of Non-Structural Sugar in White Yam Tubers from Dormancy to Sprouting
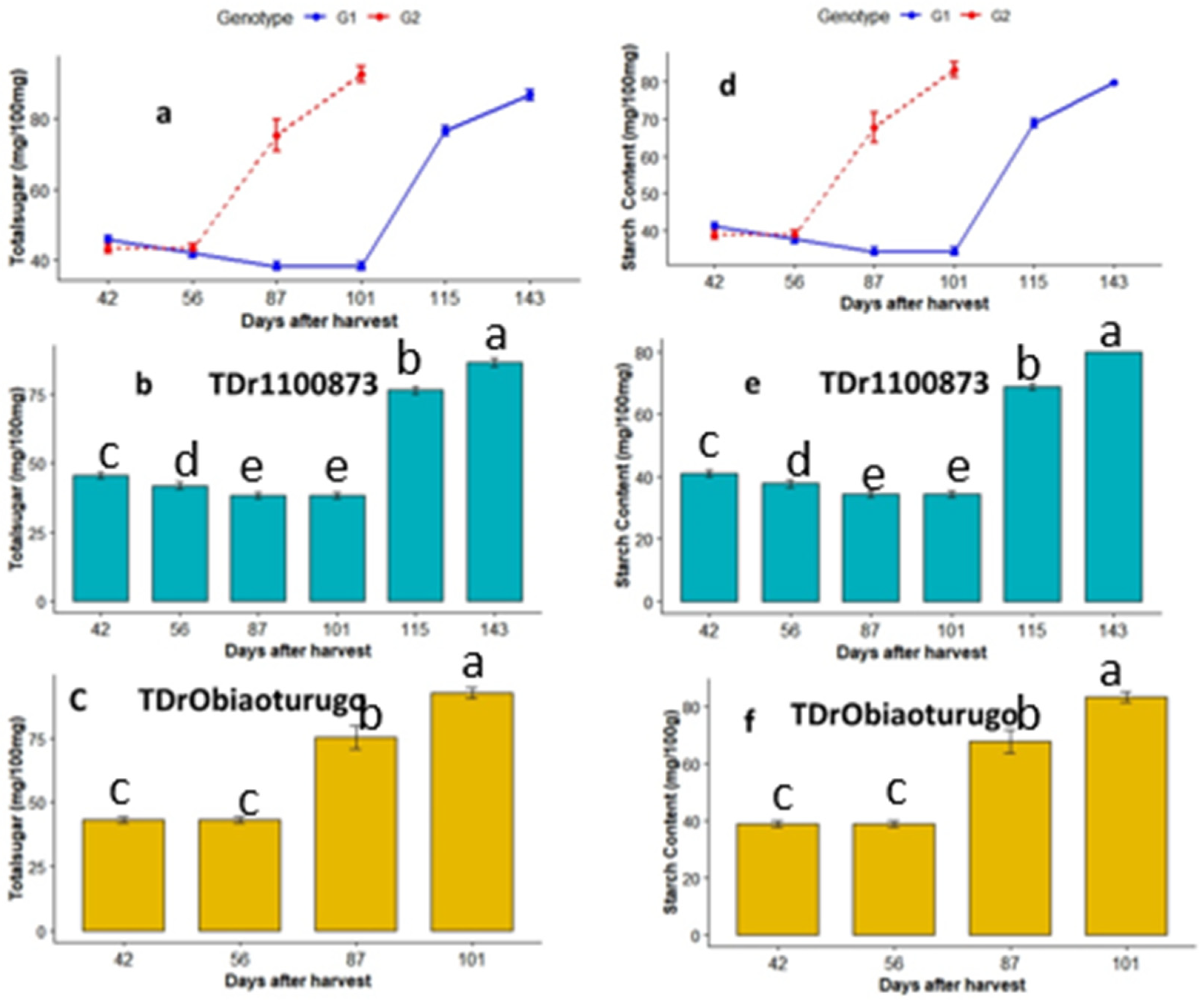
3.4.1. Total Sugar and Starch
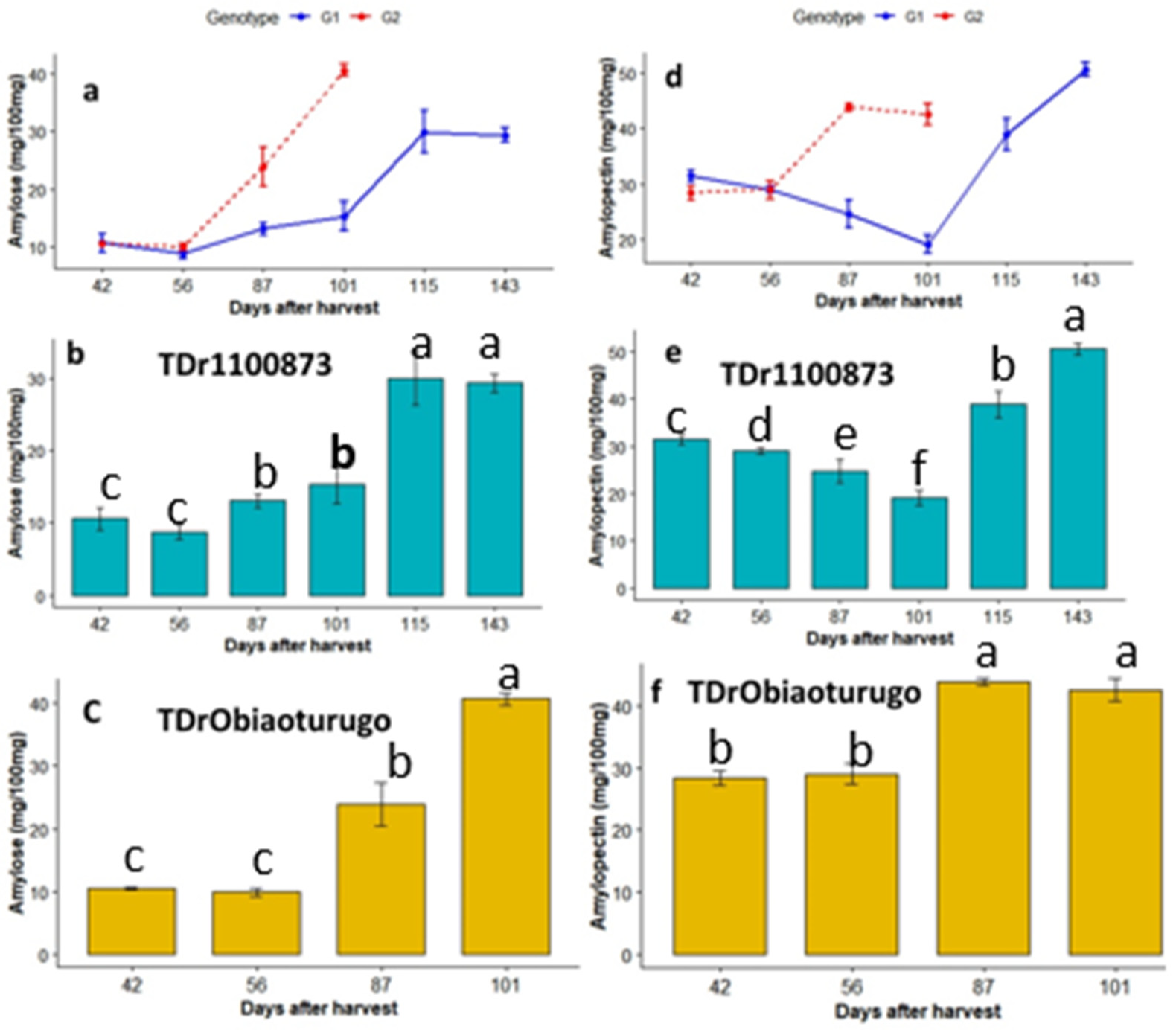
3.4.2. Amylose and Amylopectin
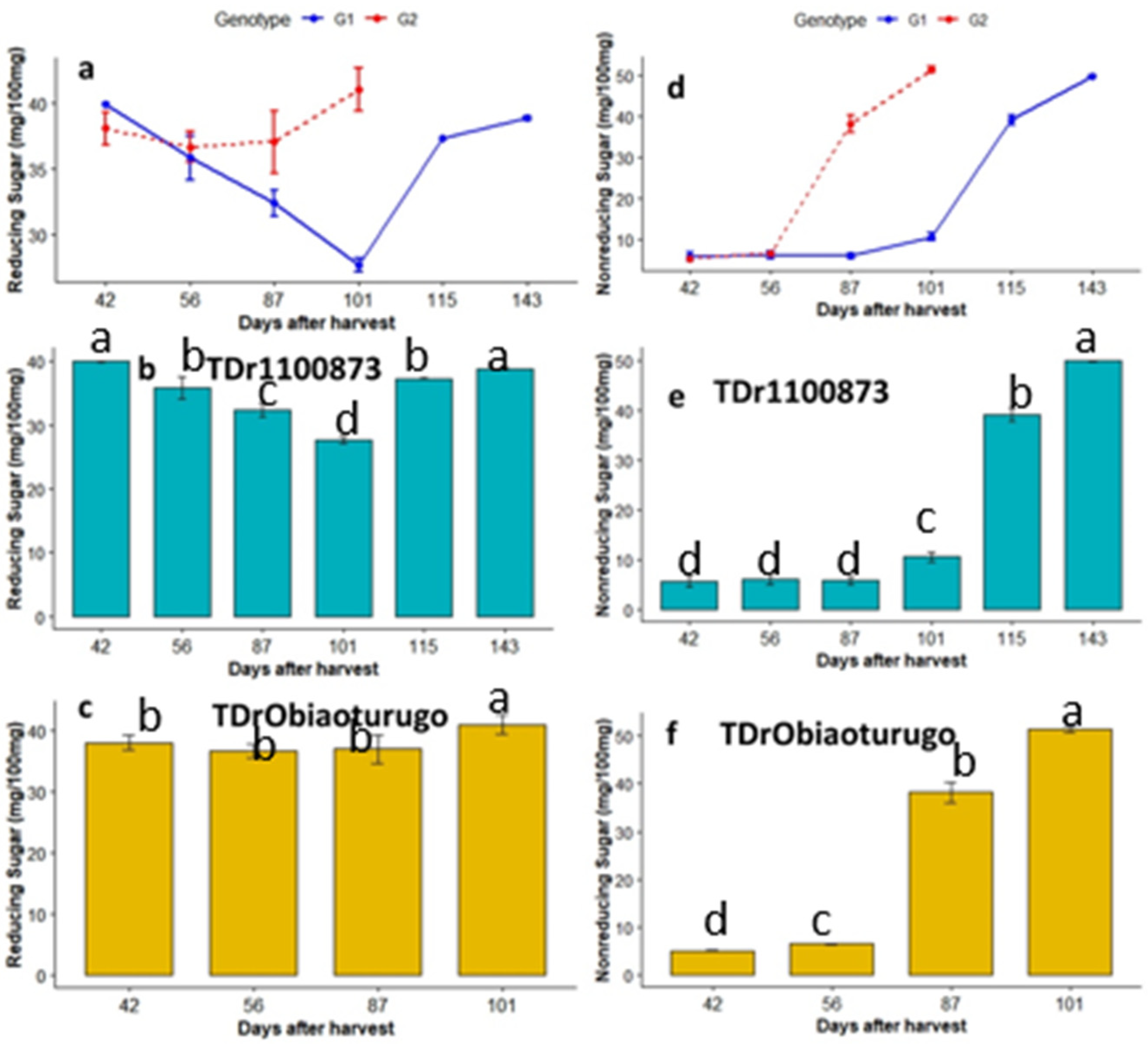
3.4.3. Reducing and Non-reducing Sugars
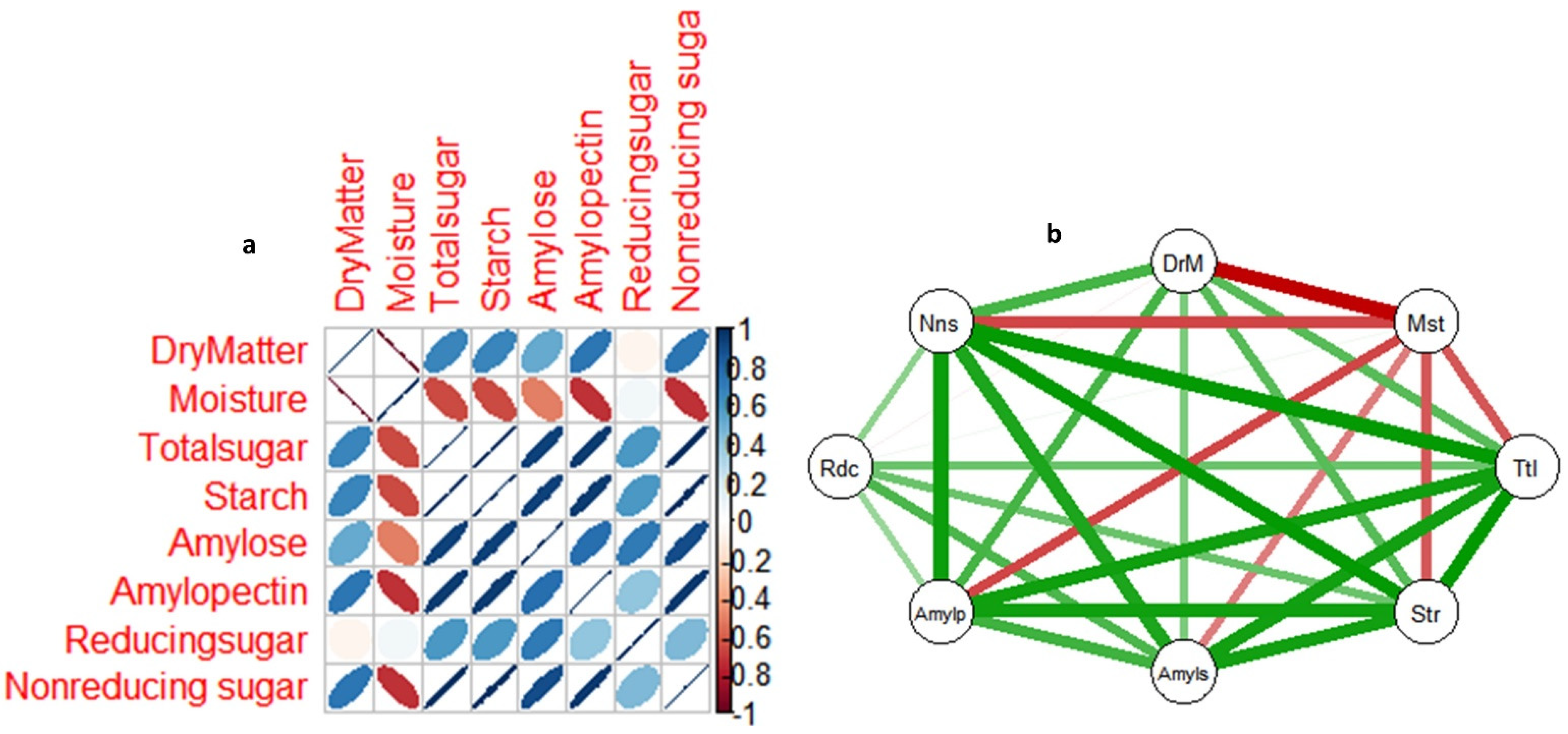
3.5. Correlations among Non-Structural Sugars, Dry Matter and Moisture Content in Yam Tuber from Dormancy to Sprouting
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam)—An appraisal of nutritional and therapeutic potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Obidiegwu, J.E.; Akpabio, E.M. The geography of yam cultivation in southern Nigeria: Exploring its social meanings and cultural functions. J. Ethn. Foods 2017, 4, 28–35. [Google Scholar] [CrossRef]
- Nwogha, J.S.; Obidiegwu, J.E.; Okereke, R.N.; Bhattacharjee, R.; Oselebe, H.O. Preliminary verification of the adoption status of some yam (Dioscorea rotundata and Dioscorea alata) varieties in Nigeria using microsatellites markers. Afr. J. Biotechnol. 2022, 21, 198–207. [Google Scholar] [CrossRef]
- Price, E.J.; Wilkin, P.; Sarasan, V.; Fraser, P.D. Metabolite profiling of Dioscorea (yam) species reveals underutilised biodiversity and renewable sources for high-value compounds. Sci. Rep. 2016, 6, 29136. [Google Scholar] [CrossRef] [PubMed]
- Mignouna, H.D.; Abang, M.M.; Asiedu, R. Harnessing modern biotechnology for tropical tuber crop improvement: Yam (Dioscorea spp.) molecular breeding. Afr. J. Biotechnol. 2003, 2, 478–485. [Google Scholar]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest Trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef]
- Ile, E.; Craufurd, P.; Battey, N.; Asiedu, R. Phases of dormancy in yam tubers (Dioscorea rotundata). Ann. Bot. 2006, 97, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Passam, H. Dormancy of yams in relation to storage. Yams. Ignames. 1982, 285–293. [Google Scholar]
- Yolou, M.; Zoundjihekpon, J.; Tiama, D.; Anizehou, S.I.; Assaba, E.I.; Adechokan, H.A.A.M.; Zongo, J.D.; Akoegninou, A. Evaluation of yam (Dioscorea cayenensis–Dioscorea rotundata) seed germination grown in Centre Benin. Int. J. Adv. Res. 2015, 3, 277–284. [Google Scholar]
- Hamadina, E.I. The Control of Yam Tuber Dormancy: A Framework for Manipulation; IITA: Ibadan, Nigeria, 2011. [Google Scholar]
- Zhang, Y.; He, J. Sugar-induced plant growth is dependent on brassinosteroids. Plant Signal. Behav. 2015, 10, e1082700. [Google Scholar] [CrossRef]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [PubMed]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87, 3583. [Google Scholar] [CrossRef]
- Wingler, A. Transitioning to the next phase: The role of sugar signaling throughout the plant life cycle. Plant Physiol. 2018, 176, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Damaris, R.N.; Lin, Z.; Yang, P.; He, D. The rice alpha-amylase, conserved regulator of seed maturation and germination. Int. J. Mol. Sci. 2019, 20, 450. [Google Scholar] [CrossRef]
- Zhaowei, L.; Qian, Z.; Fangmin, C. Sugar starvation enhances leaf senescence and genes involved in sugar signaling pathways regulate early leaf senescence in mutant rice. Rice Sci. 2020, 27, 201–214. [Google Scholar] [CrossRef]
- Yaliang, W.; Yikai, Z.; Qinghua, S.; Huizhe, C.; Jing, X.; Guohui, H.; Yanhua, C.; Xiaodan, W.; Junke, W.; Zihao, Y. Decrement of sugar consumption in rice young panicle under high temperature aggravates spikelet number reduction. Rice Sci. 2020, 27, 44–55. [Google Scholar] [CrossRef]
- Rodriguez, M.; Parola, R.; Andreola, S.; Pereyra, C.; Martínez-Noël, G. TOR and SnRK1 signaling pathways in plant response to abiotic stresses: Do they always act according to the “yin-yang” model? Plant Sci. 2019, 288, 110220. [Google Scholar] [CrossRef] [PubMed]
- Pokhilko, A.; Flis, A.; Sulpice, R.; Stitt, M.; Ebenhöh, O. Adjustment of carbon fluxes to light conditions regulates the daily turnover of starch in plants: A computational model. Mol. BioSyst. 2014, 10, 613–627. [Google Scholar] [CrossRef]
- Gibson, S.I. Control of plant development and gene expression by sugar signaling. Curr. Opin. Plant Biol. 2005, 8, 93–102. [Google Scholar] [CrossRef]
- Dennis, R. The Role of Primary Carbohydrate Metabolism in Wheat Grain Dormancy and Germination. Ph.D. Thesis, The Australian National University, Canberra, Australia, 2019. [Google Scholar]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- de Paiva Neto, V.B.; Otoni, W.C. Carbon sources and their osmotic potential in plant tissue culture: Does it matter? Sci. Hortic. 2003, 97, 193–202. [Google Scholar] [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. I. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef]
- Wingler, A.; Henriques, R. Sugars and the speed of life—Metabolic signals that determine plant growth, development and death. Physiol. Plant. 2022, 174, e13656. [Google Scholar] [CrossRef] [PubMed]
- Martín-Fontecha, E.S.; Tarancón, C.; Cubas, P. To grow or not to grow, a power-saving program induced in dormant buds. Curr. Opin. Plant Biol. 2018, 41, 102–109. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Manivannan, P.; Kishorekumar, A.; Gomathinayagam, M.; Panneersel Vam, R. Changes in biochemical constituents and induction of early sprouting by triadimefon treatment in white yam (Dioscorea rotundata Poir.) tubers during storage. J. Zhejiang Univ. Sci. B 2007, 8, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Tschannen, A.B.; Girardin, O.; Nindjin, C.; Daouda, D.; Farah, Z.; Stamp, P.; Escher, F. Improving the application of gibberellic acid to prolong dormancy of yam tubers (Dioscorea spp). J. Sci. Food Agric. 2003, 83, 787–796. [Google Scholar] [CrossRef]
- Hariprakash, C.; Nambisan, B. Carbohydrate metabolism during dormancy and sprouting in yam (Dioscorea) tubers: Changes in carbohydrate constituents in yam (Dioscorea) tubers during dormancy and sprouting. J. Agric. Food Chem. 1996, 44, 3066–3069. [Google Scholar] [CrossRef]
- Landhäusser, M.S.; Pak, S.C.; Dickman, L.T.; Furze, E.M.; Kuhlman, I.; Schmid, S.; Wiesenbauer, J.; Wild, B.; Gleixner, G.; Hartmann, H.; et al. Standardized protocols and procedures can precisely and accurately quantify non-structural carbohydrates. Tree Physiol. 2018, 38, 1764–1778. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Widdowson, E.M. A method for the determination of small quantities of mixed reducing sugars and its application to the estimation of the products of hydrolysis of starch by taka-diastase. Biochem. J. 1931, 25, 863–879. [Google Scholar] [CrossRef]
- Sowbhagya, C.M.; Bhattacharya, K.R. A Simplified Colorimetric Method for Determination of Amylose Content in Rice. Starch 1971, 23, 53–56. [Google Scholar] [CrossRef]
- Huang, C.-C.; Chiang, P.-Y.; Chen, Y.-Y.; Wang, C.-C. Chemical compositions and enzyme activity changes occurring in yam (Dioscorea alata L.) tubers during growth. LWT Food Sci. Technol. 2007, 40, 1498–1506. [Google Scholar] [CrossRef]
- Muzac-Tucker, I.; Helen, N.; Ahmad, M. Biochemical composition and storage of Japanican yams (Dioscorea spp.). J. Sci. Food Agric. 1993, 62, 219–224. [Google Scholar] [CrossRef]
- Paul, M.J.; Lawlor, D.W. Genetic modification of primary metabolism|Photosynthesis. In Encyclopedia of Applied Plant Sciences; Academic Press: Cambridge, MA, USA, 2003; pp. 484–493. [Google Scholar]
- Jabrin, S.; Ravanel, S.; Gambonnet, B.; Douce, R.; Rébeillé, F. One-carbon metabolism in plants. Regulation of tetrahydrofolate synthesis during germination and seedling development. Plant Physiol. 2003, 131, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Landhäusser, S. Aspen shoots are carbon autonomous during bud break. Trees 2011, 25, 531–536. [Google Scholar] [CrossRef]
- Ohanenye, I.C.; Alamar, M.C.; Thompson, A.J.; Terry, L.A. Fructans redistribution prior to sprouting in stored onion bulbs is a potential marker for dormancy break. Postharvest Biol. Technol. 2019, 149, 221–234. [Google Scholar] [CrossRef]
- Smeekens, S.; Hellmann, A.H. Sugar sensing and signaling in plants. Front. Plant Sci. 2014, 5, 185–205. [Google Scholar] [CrossRef]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef]
- Doblin, M.S.; Kurek, I.; Jacob-Wilk, D.; Delmer, D.P. Cellulose biosynthesis in plants: From genes to rosettes. Plant Cell Physiol. 2002, 43, 1407–1420. [Google Scholar] [CrossRef]
- Tsai, A.Y.-L.; Gazzarrini, S. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: The emerging picture. Front. Plant Sci. 2014, 5, 119. [Google Scholar] [CrossRef] [PubMed]
- Durán-Soria, S.; Pott, D.M.; Osorio, S.; Vallarino, J.G. Sugar signaling during fruit ripening. Front. Plant Sci. 2020, 11, 564917. [Google Scholar] [CrossRef]
- Hartmann, H.; Adams, H.D.; Hammond, W.M.; Hoch, G.; Landhäusser, S.M.; Wiley, E.; Zaehle, S. Identifying differences in carbohydrate dynamics of seedlings and mature trees to improve carbon allocation in models for trees and forests. Environ. Exp. Bot. 2018, 152, 7–18. [Google Scholar] [CrossRef]
- Yadav, U.P.; Ivakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.-M.; Stitt, M. The sucrose-trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot. 2014, 65, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Baena-González, E.; Lunn, J.E. SnRK1 and trehalose 6-phosphate–two ancient pathways converge to regulate plant metabolism and growth. Curr. Opin. Plant Biol. 2020, 55, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Goddijn, O.J.; van Dun, K. Trehalose metabolism in plants. Trends Plant Sci. 1999, 4, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, C.M.; Feil, R.; Ishihara, H.; Watanabe, M.; Kölling, K.; Krause, U.; Höhne, M.; Encke, B.; Plaxton, W.C.; Zeeman, S.C. Trehalose 6–phosphate coordinates organic and amino acid metabolism with carbon availability. Plant J. 2016, 85, 410–423. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR signaling promotes accumulation of BZR1 to balance growth with carbon availability in Arabidopsis. Curr. Biol. 2016, 26, 1854–1860. [Google Scholar] [CrossRef]
- Bledsoe, S.W.; Henry, C.; Griffiths, C.A.; Paul, M.J.; Feil, R.; Lunn, J.E.; Stitt, M.; Lagrimini, L.M. The role of Tre6P and SnRK1 in maize early kernel development and events leading to stress-induced kernel abortion. BMC Plant Biol. 2017, 17, 74. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | DAPM | DM% | Moisture% | Total Sugar(mg/100 mg) | Starch (mg/100 mg) | Amylopectin (mg/100 mg) | Amylose (mg/100 mg) | Reducing Sugar (mg/100 mg) | Non-Reducing Sugar (mg/100 mg) |
|---|---|---|---|---|---|---|---|---|---|
| TDr1100873 | 42 days | 46.12 c | 53.88 a | 45.63 c | 41.07 c | 31.45 c | 10.62 c | 39.97 a | 5.66 d |
| TDr1100873 | 56 days | 48.35 c | 51.65 b | 41.93 d | 37.74 d | 28.98 d | 8.76 c | 35.87 b | 6.07 d |
| TDr1100873 | 87 days | 52.3 b | 47.69 c | 38.23 e | 34.41 e | 24.67 e | 13.07 b | 32.42 c | 5.81 d |
| TDr1100873 | 101 days | 54.29 b | 45.72 d | 38.23 e | 34.41 e | 19.12 f | 15.29 b | 27.68 d | 10.56 c |
| TDr1100873 | 115 days | 57.55 a | 42.45 e | 76.47 b | 68.82 b | 38.85 b | 27.5 a | 37.32 b | 39.15 b |
| TDr1100873 | 143 days | 59.91 a | 40.09 f | 86.7 a | 79.92 a | 50.57 a | 29.35 a | 38.87 a | 49.94 a |
| TDrObia | 42 days | 43.24 d | 67.17 a | 44.4 c | 38.85 c | 28.37 b | 10.48 c | 38.1 ab | 5.07 d |
| TDrObia | 56 days | 48.5 c | 51.5 b | 43.17 c | 38.85 c | 28.98 b | 9.86 c | 36.71 b | 6.46 c |
| TDrObia | 87 days | 54.47 b | 45.46 c | 75.23 b | 67.71 b | 43.78 a | 23.93 b | 37.14 ab | 38.18 b |
| TDrObia | 101 days | 59.88 a | 40.12 d | 92.5 a | 83.25 a | 42.55 a | 40.7 a | 41.1 a | 51.44 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwogha, J.S.; Abtew, W.G.; Raveendran, M.; Oselebe, H.O.; Obidiegwu, J.E.; Chilaka, C.A.; Amirtham, D.D. Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata). Agriculture 2023, 13, 343. https://doi.org/10.3390/agriculture13020343
Nwogha JS, Abtew WG, Raveendran M, Oselebe HO, Obidiegwu JE, Chilaka CA, Amirtham DD. Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata). Agriculture. 2023; 13(2):343. https://doi.org/10.3390/agriculture13020343
Chicago/Turabian StyleNwogha, Jeremiah S., Wosene G. Abtew, Muthurajan Raveendran, Happiness O. Oselebe, Jude E. Obidiegwu, Cynthia A. Chilaka, and Damodarasamy D. Amirtham. 2023. "Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata)" Agriculture 13, no. 2: 343. https://doi.org/10.3390/agriculture13020343
APA StyleNwogha, J. S., Abtew, W. G., Raveendran, M., Oselebe, H. O., Obidiegwu, J. E., Chilaka, C. A., & Amirtham, D. D. (2023). Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata). Agriculture, 13(2), 343. https://doi.org/10.3390/agriculture13020343