
Effect of Nitrogen Fertilization on Tree Growth and Nutrient Content in Soil and Cherry Leaves (Prunus cerasus L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Location and Main Agrotechnical Treatments
2.2. Fertilization
2.2.1. Nitrogen Fertilization
2.2.2. Fertilization with Other Ingredients
2.3. Measurements, Observations, and Analyses
2.3.1. Vegetative Growth
2.3.2. Soil Sampling
Determination of Minerals
Determination of Nitrogen in Soil
2.3.3. Leaves Sampling
2.3.4. Determination of Pigments in Leaves
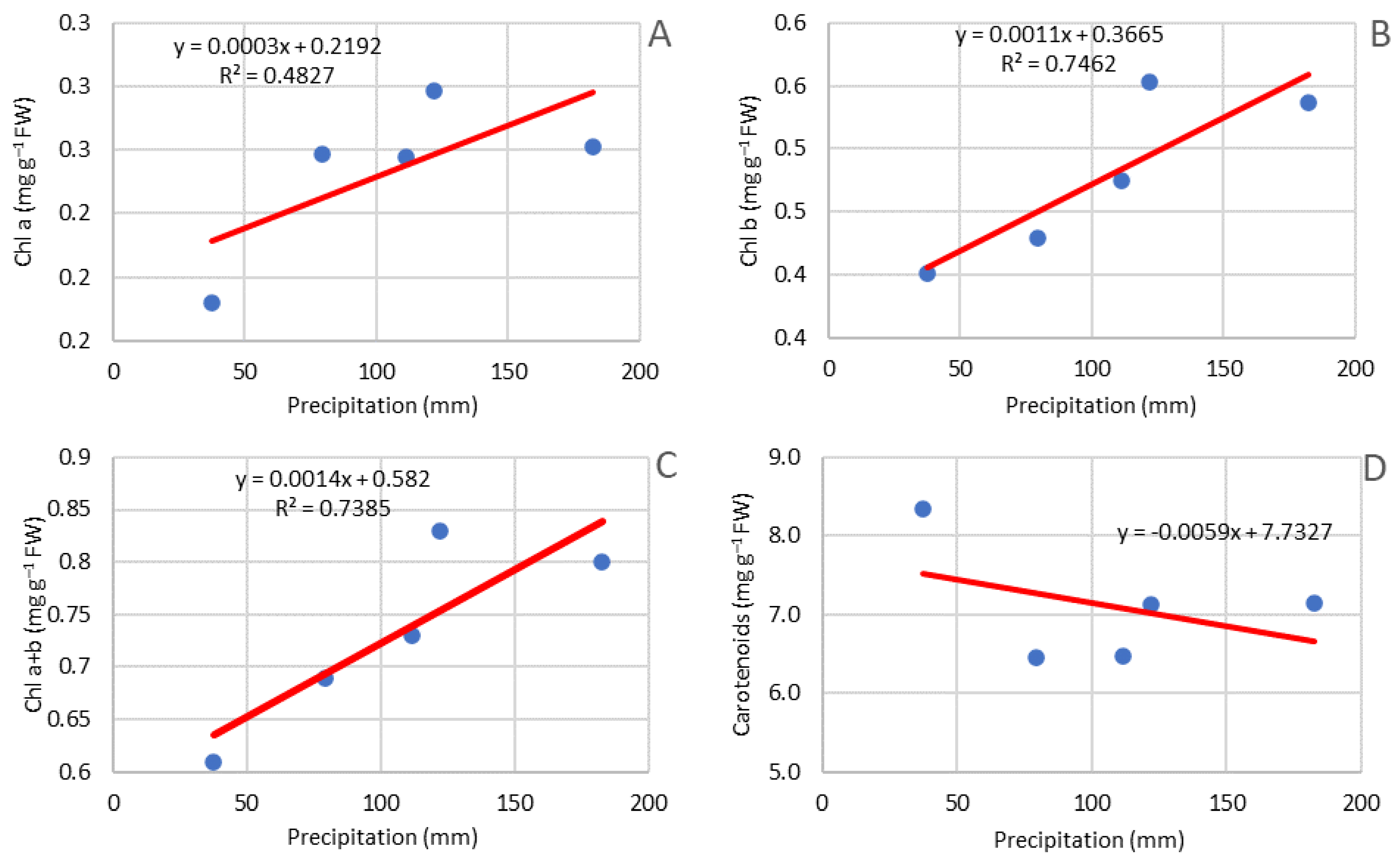
2.4. Analysis of Weather Conditions
2.5. Statistical Analysis of Results
3. Results and Discussion
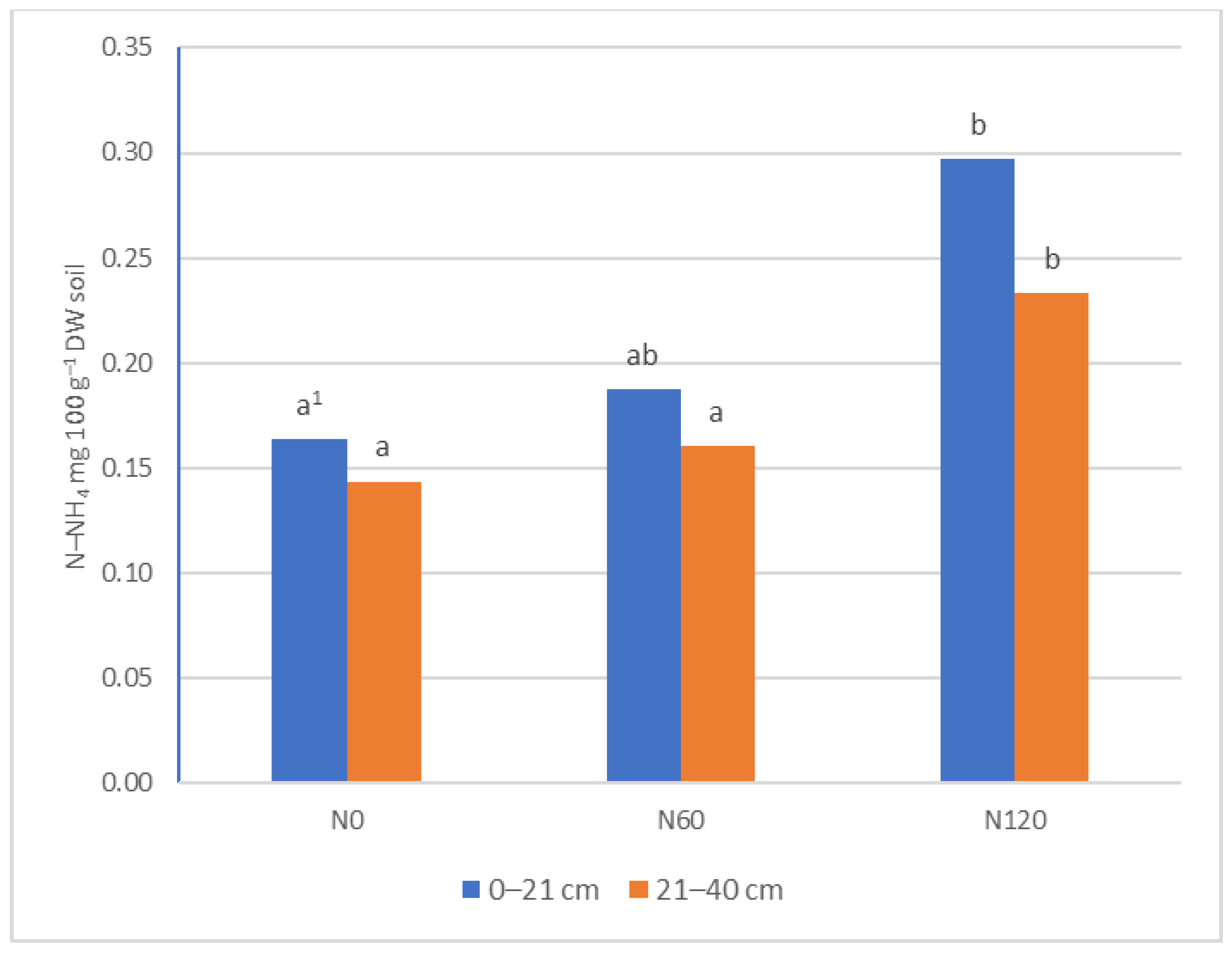
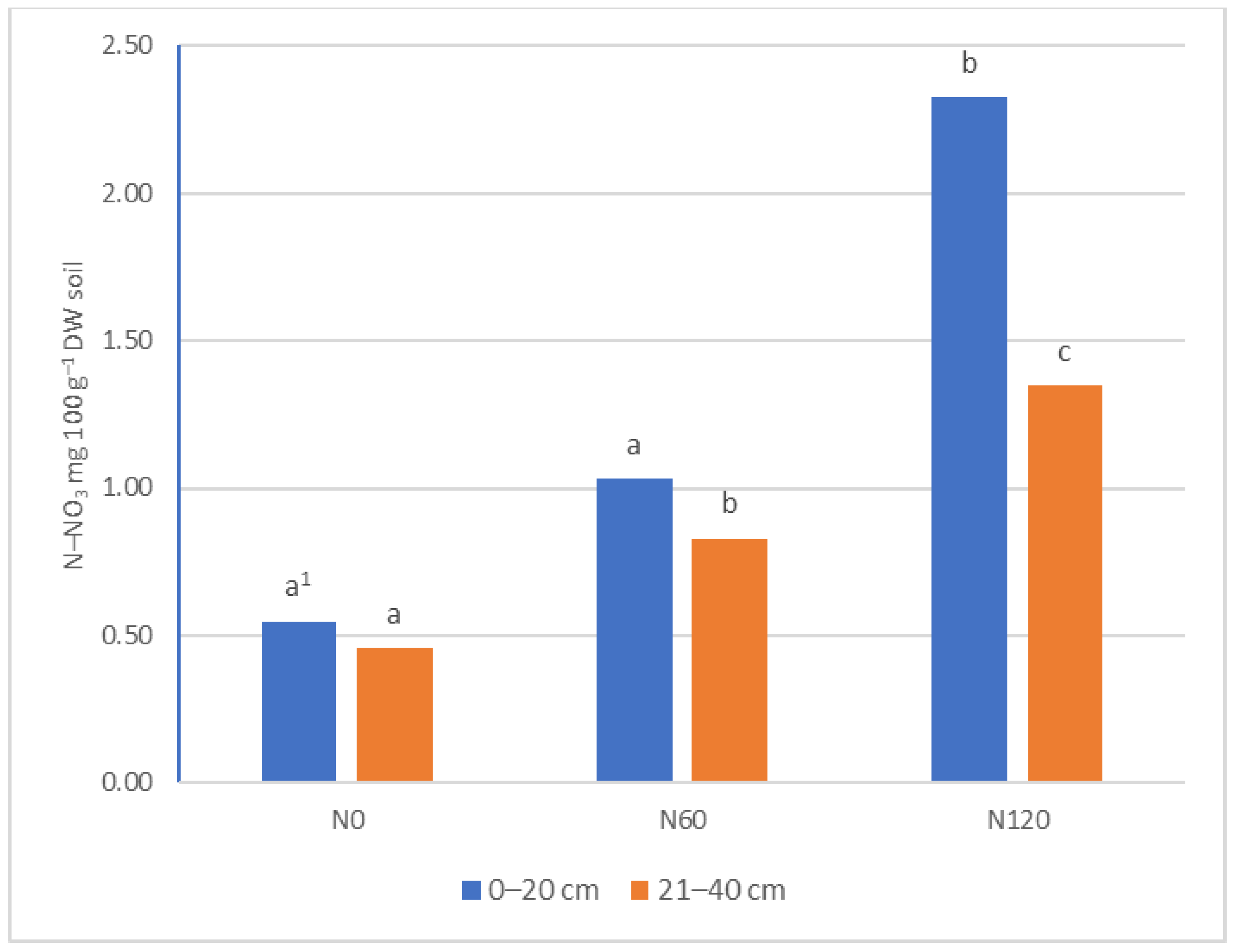
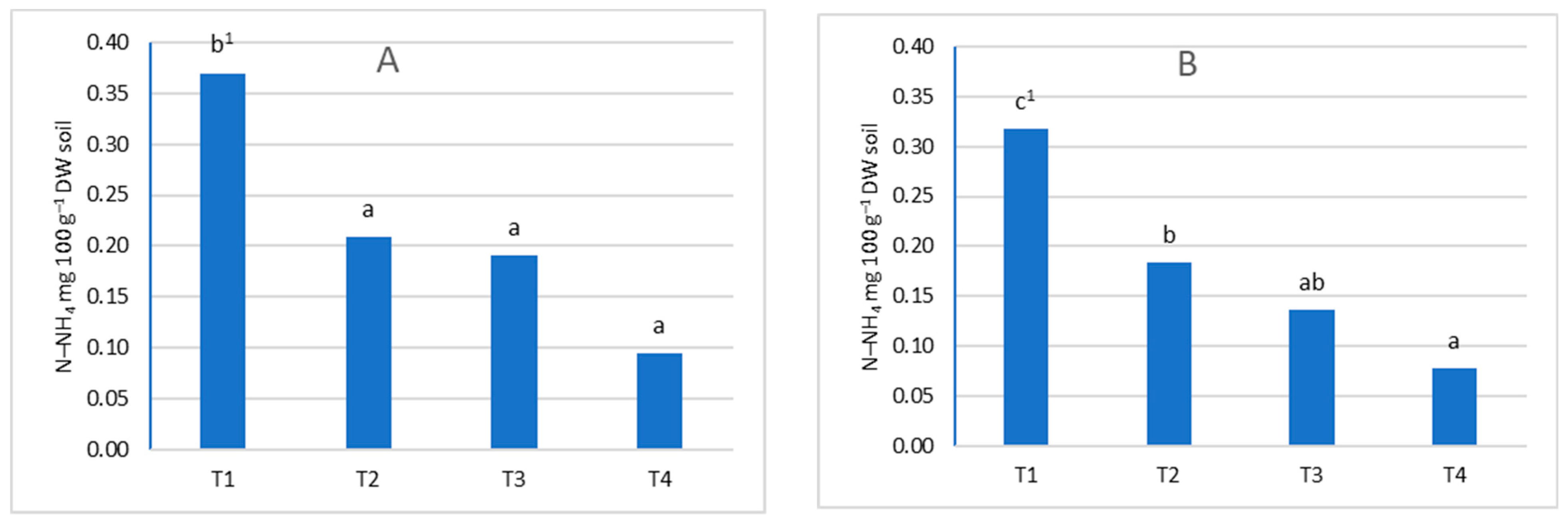
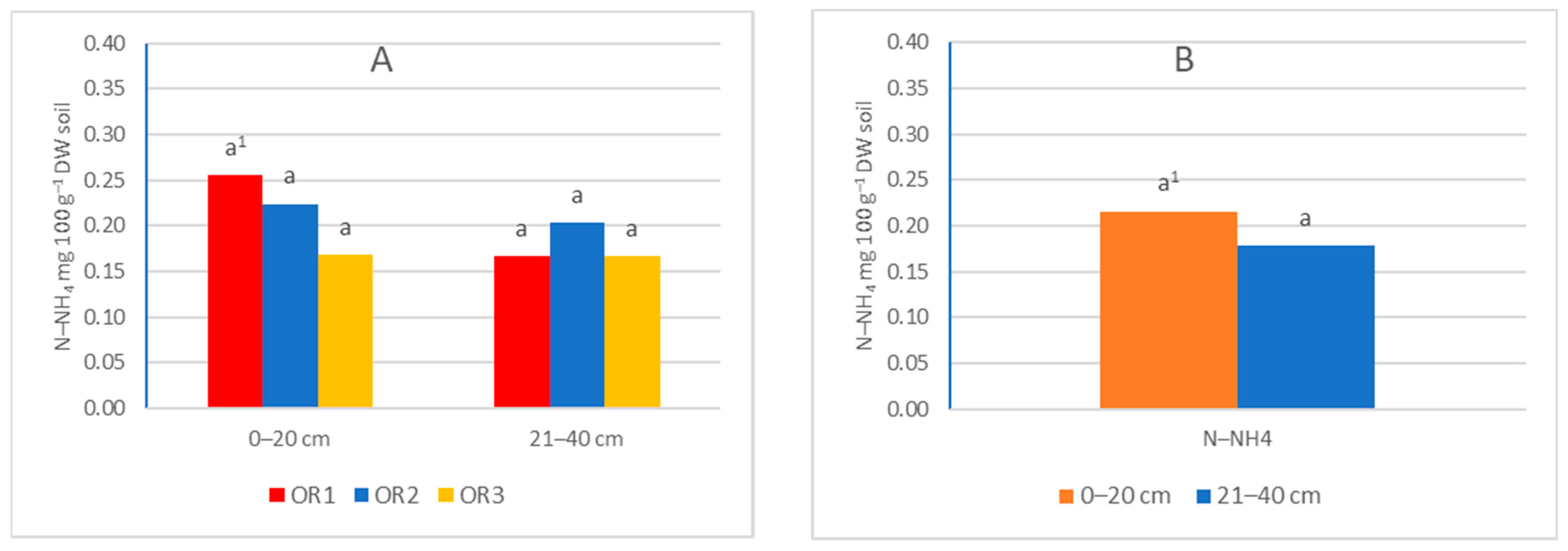
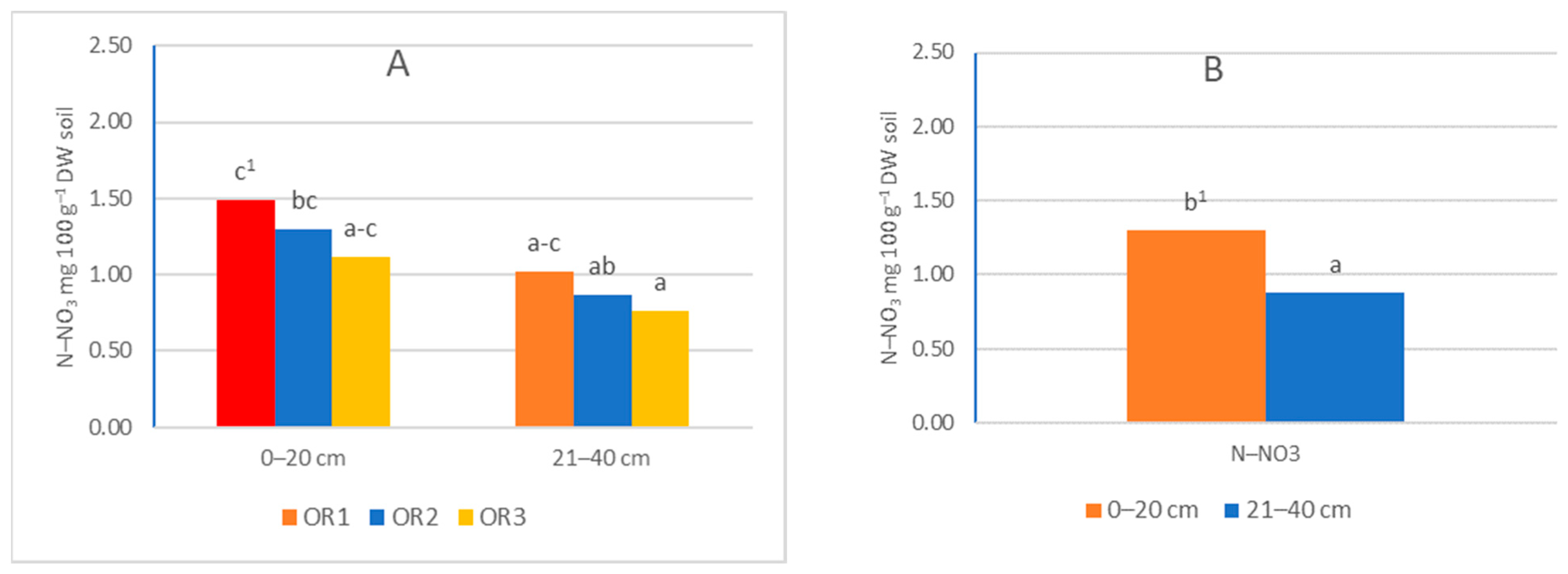
3.1. Ammonia and Nitrate Nitrogen Content in Soil
3.2. Content of Minerals Other Than Nitrogen in the Soil
3.3. Mineral Content in Leaves
3.4. Effect of Nitrogen Fertilization on the Growth of Cherry Trees
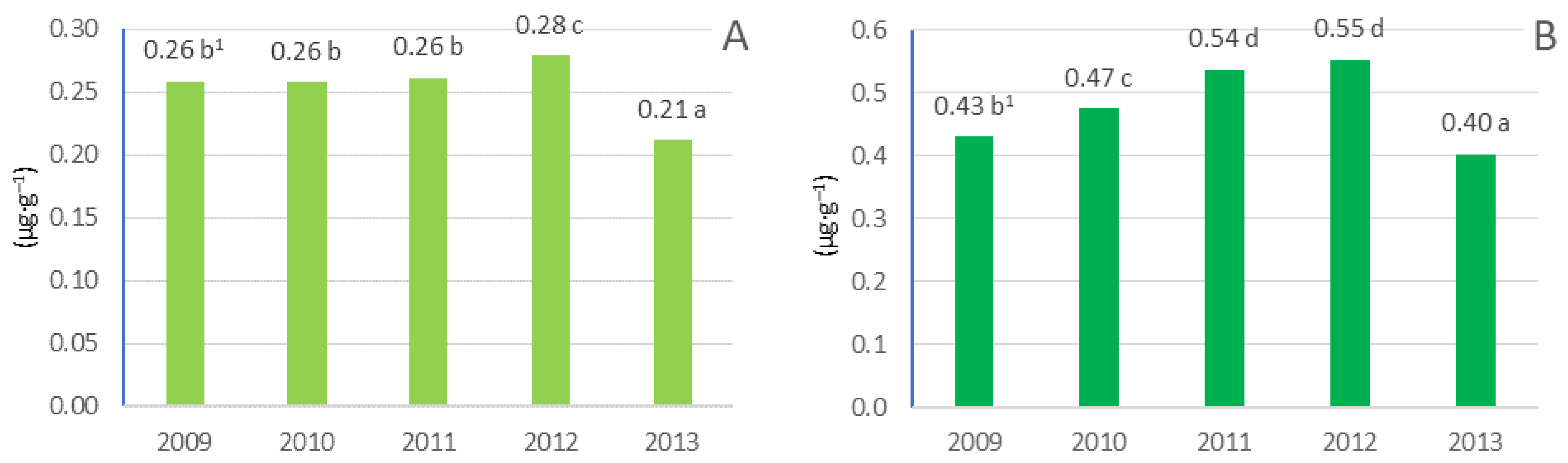
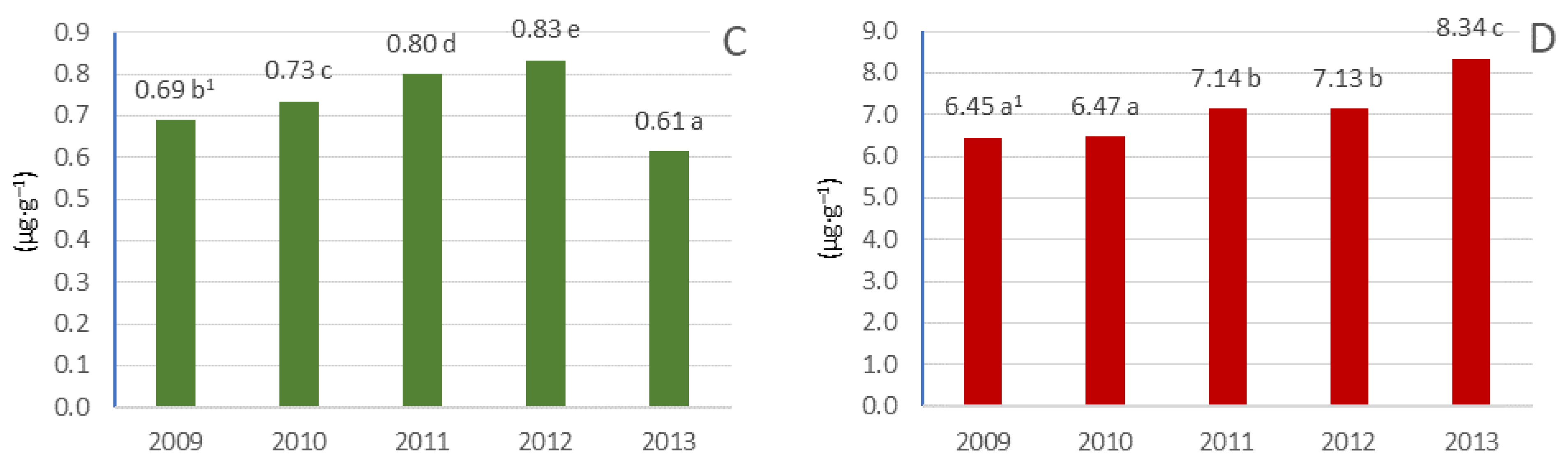
3.5. Pigment Content in Leaves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, J.; Dong, Y.; Xie, X.; Li, X.; Zhang, X.; Shen, X. Effect of Annual Variation in Soil PH on Available Soil Nutrients in Pear Orchards. Acta Ecol. Sin. 2011, 31, 212–216. [Google Scholar] [CrossRef]
- Ben Mimoun, M.; Marchand, M. Effects of Potassium Foliar Fertilization on Different Fruit Tree Crops over Five Years of Experiments. Acta Hortic. 2013, 211–217. [Google Scholar] [CrossRef]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, M.; Scandellari, F.; Bonora, E.; Tagliavini, M. Nutrient Release during Decomposition of Leaf Litter in a Peach (Prunus persica L.) Orchard. Nutr. Cycl. Agroecosyst. 2010, 87, 115–125. [Google Scholar] [CrossRef]
- Testolin, R.; Ferguson, A.R. Kiwifruit (Actinidia spp.) Production and Marketing in Italy. N. Z. J. Crop Hortic. Sci. 2009, 37, 1–32. [Google Scholar] [CrossRef]
- Pietrzak, M.; Chlebicka, A.; Kraciński, P.; Malak-Rawlikowska, A. Information Asymmetry as a Barrier in Upgrading the Position of Local Producers in the Global Value Chain—Evidence from the Apple Sector in Poland. Sustainability 2020, 12, 7857. [Google Scholar] [CrossRef]
- Gaworski, M.; Ciesielski, W.; Kaminska, N. Effect of Modern Apple Production Technologies on Development of Family Orchard Farming with Particular Focus on Polish Conditions. Eng. Rural Dev. 2017, 24, 408–414. [Google Scholar]
- Olszak, R. Methodology of Integrated Cherry Protection (in Polish), 4th ed.; Main Inspector of Plant Health and Seed Inspection: Warsaw, Poland, 2020. [Google Scholar]
- Huett, D.O. Prospects for Manipulating the Vegetative-Reproductive Balance in Horticultural Crops through Nitrogen Nutrition: A Review. Aust. J. Agric. Res. 1996, 47, 47–66. [Google Scholar] [CrossRef] [Green Version]
- Lysiak, P.G.; Pacholak, E. Effects of 13 Years Soil Fertilisation on Storage Quality of ‘Cortland’ Apples. Acta Hortic. 1999, 485, 265–272. [Google Scholar] [CrossRef]
- Kurlus, R.; Rutkowski, K.; Łysiak, G.P. Improving of Cherry Fruit Quality and Bearing Regularity by Chemical Thinning with Fertilizer. Agronomy 2020, 10, 1281. [Google Scholar] [CrossRef]
- Rutkowski, K.; Zydlik, Z.; Pacholak, E. Effect of Tree Pruning Intensity on the Yield and Fruit Quality of the Sour Cherry. Zemdirb.-Agric. 2015, 102, 417–422. [Google Scholar] [CrossRef]
- Dziedzic, E.; Bieniasz, M.; Kowalczyk, B. Morphological and Physiological Features of Sweet Cherry Floral Organ Affecting the Potential Fruit Crop in Relation to the Rootstock. Sci. Hortic. 2019, 251, 127–135. [Google Scholar] [CrossRef]
- Wójcik, P. Nawozy i Nawożenie Drzew Owocowych; Hortpress: Warsaw, Poland, 2009; ISBN 83-89211-88-2. [Google Scholar]
- Jankiewicz, L.; Lipecki, J. Fizjologia Roslin Sadowniczych; Państwowe Wydawn. Nauk.: Warsaw, Poland, 2011; Volume 1, ISBN 978-83-01-16685-4. [Google Scholar]
- Lin, Z.; YuanMao, J.; FuTian, P.; Xu, Z.; XiangJi, F.; HongBo, L. Studies on utilization and accumulation dynamics of spring soil 15N-urea application in apple orchard. Acta Hortic. Sin. 2009, 36, 1805–1809. [Google Scholar]
- Grassi, G.; Millard, P.; Gioacchini, P.; Tagliavini, M. Recycling of Nitrogen in the Xylem of Prunus avium Trees Starts When Spring Remobilization of Internal Reserves Declines. Tree Physiol. 2003, 23, 1061–1068. [Google Scholar] [CrossRef]
- Tromp, J. Nutrient Reserves in Roots of Fruit Trees, in Particular Carbohydrates and Nitrogen. Plant Soil 1983, 71, 401–413. [Google Scholar] [CrossRef]
- Neto, C.; Carranca, C.; Clemente, J. Senescent Leaf Decomposition in a Mediterranean Pear Orchard. Eur. J. Agron. 2009, 30, 34–40. [Google Scholar] [CrossRef]
- Tagliavini, M.; Tonon, G.; Scandellari, F.; Quiñones, A.; Palmieri, S.; Menarbin, G.; Gioacchini, P.; Masia, A. Nutrient Recycling during the Decomposition of Apple Leaves (Malus domestica) and Mowed Grasses in an Orchard. Agric. Ecosyst. Environ. 2007, 118, 191–200. [Google Scholar] [CrossRef]
- Kozieł, M.; Gałązka, A.; Martyniuk, S. Wolnożyjące Bakterie Wiążące Azot Atmosferyczny z Rodzaju Azotobacter–Występowanie, Liczebność i Znaczenie. Stud. Rap. IUNG-PIB 2018, 56, 57–70. [Google Scholar] [CrossRef]
- Rutkowski, K.; Łysiak, G.P.; Zydlik, Z. Effect of Nitrogen Fertilization in the Sour Cherry Orchard on Soil Enzymatic Activities, Microbial Population, and Fruit Quality. Agriculture 2022, 12, 2069. [Google Scholar] [CrossRef]
- Kozieł, M.; Martyniuk, S.; Siebielec, G. Occurrence of Azotobacter spp. in Cultivated Soils in Poland. Pol. J. Agron. 2021, 44, 3–8. [Google Scholar] [CrossRef]
- Gałązka, A.; Bigos, J.; Siebielec, S. Systematyka, Genetyka i Biologia Bakterii z Rodzaju Azospirillum (Systematics, Genetics and Biology of Bacteria from Genus Azospirillum). Pol. J. Agron. 2015, 23, 31–47. [Google Scholar]
- Saa, S.; Fernández, E.; Muhammad, S.; Río, A.O.-D.; DeJong, T.M.; Laca, E.; Brown, P. Increases in Leaf Nitrogen Concentration and Leaf Area Did Not Enhance Spur Survival and Return Bloom in Almonds (Prunus dulcis [Mill.] DA Webb). Acta Physiol. Plant. 2017, 39, 1–10. [Google Scholar] [CrossRef]
- Khaosumain, Y.; Sritontip, C.; Changjeraja, S. Effects of Different Nitrogen Fertilization Doses on Growth, Leaf Nutrient Concentration, Flowering and Fruit Quality in off-Season Longan. Acta Hortic. 2013, 984, 271–274. [Google Scholar] [CrossRef]
- Sete, P.B.; Comin, J.J.; Nara Ciotta, M.; Almeida Salume, J.; Thewes, F.; Brackmann, A.; Toselli, M.; Nava, G.; Rozane, D.E.; Loss, A.; et al. Nitrogen Fertilization Affects Yield and Fruit Quality in Pear. Sci. Hortic. 2019, 258, 108782. [Google Scholar] [CrossRef]
- Rosati, A.; Paoletti, A.; Al Hariri, R.; Famiani, F. Fruit Production and Branching Density Affect Shoot and Whole-Tree Wood to Leaf Biomass Ratio in Olive. Tree Physiol. 2018, 38, 1278–1285. [Google Scholar] [CrossRef] [Green Version]
- Pierce, L.L.; Running, S.W.; Walker, J. Regional-Scale Relationships of Leaf Area Index to Specific Leaf Area and Leaf Nitrogen Content. Ecol. Appl. 1994, 4, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Tromp, J.; Ovaa, J.C. Uptake and Distribution of Nitrogen in Young Apple Trees after Application of Nitrate or Ammonium, with Special Reference to Asparagine and Arginine. Physiol. Plant. 1979, 45, 23–28. [Google Scholar] [CrossRef]
- Rupp, D. Nitrogen fertilization in apple orchards-relationships between available nitrogen in soil samples, nitrates in soil water and leaching of nitrogen. Acta Hortic. 1995, 401–410. [Google Scholar] [CrossRef]
- Zipori, I.; Erel, R.; Yermiyahu, U.; Ben-Gal, A.; Dag, A. Sustainable Management of Olive Orchard Nutrition: A Review. Agriculture 2020, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Wojcik, P. Ecological Impact of Nitrogen Fertilization. J. Fruit Ornam. Plant Res. 2001, 9, 1–4. [Google Scholar]
- Cui, M.; Zeng, L.; Qin, W.; Feng, J. Measures for Reducing Nitrate Leaching in Orchards: A Review. Environ. Pollut. 2020, 263, 114553. [Google Scholar] [CrossRef] [PubMed]
- Viljevac, M.; Dugalić, K.; Mihaljević, I.; Šimić, D.; Sudar, R.; Jurković, Z.; Lepeduš, H. Chlorophyll Content, Photosynthetic Efficiency and Genetic Markers in Two Sour Cherry (Prunus cerasus L.) Genotypes under Drought Stress. Acta Bot. Croat. 2013, 72, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Wojdyło, A.; Nowicka, P.; Tkacz, K.; Turkiewicz, I.P. Fruit Tree Leaves as Unconventional and Valuable Source of Chlorophyll and Carotenoid Compounds Determined by Liquid Chromatography-Photodiode-Quadrupole/Time of Flight-Electrospray Ionization-Mass Spectrometry (LC-PDA-QTof-ESI-MS). Food Chem. 2021, 349, 129156. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, P. Physiological Basics of Sweet Cherry Productivity Depending on Rootstocks, Interstems and Plant Density. Open Agric. 2019, 4, 267–274. [Google Scholar] [CrossRef]
- Flore, J.; Layne, D.R. Photoassimilate Production and Distribution in Cherry. HortScience 1999, 34, 1015–1019. [Google Scholar] [CrossRef] [Green Version]
- Pacholak, E.; Zydlik, Z.; Rutkowski, K. Effect of Cherry Nitrogen Fertilization on the Content of Minerals in the Leaves and Soil. Acta Sci. Pol. Hortorum Cultus 2011, 10, 105–112. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2, Chemical and Microbiological Properties; Agronomy, 2nd ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; ISBN 978-0-89118-072-2. [Google Scholar]
- Sadowski, A.; Nurzyński, J.; Pacholak, E.; Smolarz, K. Określenie Potrzeb Nawożenia Roślin Sadowniczych II. Zasady, Liczby Graniczne i Dawki Nawożenia. Instr. Upowsz. 1990, 3, 1–25. [Google Scholar]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, W.; Wrona, D.; Przybyłko, S. Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits. Agriculture 2022, 12, 2169. [Google Scholar] [CrossRef]
- Komosa, A. Changes in Some Chemical Properties of the Soil under Grass Sward and Herbicide Strips in Apple Orchards. Acta Hortic. 1990, 274, 223–230. [Google Scholar] [CrossRef]
- Engel, A.; Blanke, M.; Lenz, F. Wie viele Nährstoffe werden mit dem Mulchen der Fahrgassen in den Baumstreifen einer Obstanlage verfrachtet? Erwerbs-Obstbau 2009, 51, 151–161. [Google Scholar] [CrossRef]
- Wrona, D. The Influence of Nitrogen Fertilization on Growth, Yield and Fruit Size of ‘Jonagored’ apple Trees. Acta Sci. Pol.-Hortorum Cultus 2011, 10, 3–10. [Google Scholar]
- Neilsen, D.; Neilsen, G.H. Efficient Use of Nitrogen and Water in High-Density Apple Orchards. HortTechnology 2002, 12, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Sobiczewski, P. Methodology of Integrated Protection of Apple Trees (Polish), 6th ed.; Main Inspector of Plant Health and Seed Inspection: Warsaw, Poland, 2020. [Google Scholar]
- Nicolardot, B.; Fauvet, G.; Cheneby, D. Carbon and Nitrogen Cycling through Soil Microbial Biomass at Various Temperatures. Soil Biol. Biochem. 1994, 26, 253–261. [Google Scholar] [CrossRef]
- Guttières, R.; Nunan, N.; Raynaud, X.; Lacroix, G.; Barot, S.; Barré, P.; Girardin, C.; Guenet, B.; Lata, J.-C.; Abbadie, L. Temperature and Soil Management Effects on Carbon Fluxes and Priming Effect Intensity. Soil Biol. Biochem. 2021, 153, 108103. [Google Scholar] [CrossRef]
- Cassman, K.G.; Munns, D.N. Nitrogen Mineralization as Affected by Soil Moisture, Temperature, and Depth. Soil Sci. Soc. Am. J. 1980, 44, 1233–1237. [Google Scholar] [CrossRef] [Green Version]
- Charyulu, P.B.B.N.; Rao, V.R. Influence of Various Soil Factors on Nitrogen Fixation by Azospirillum spp. Soil Biol. Biochem. 1980, 12, 343–346. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Domżał, H.; Świca, M. Influence of orchard use on the changes in selected soil properties. Acta Agrophys. 2001, 29–39. [Google Scholar]
- Wrona, D.; Sadowski, A. Effect of Nitrogen Fertilization and Soil Management on Soil Mineral Nitrogen in the Apple Orchard. J. Fruit Ornamen. Plant Res. 2004, 12, 191–199. [Google Scholar]
- Paltineanu, C.; Iancu, M. Orchard Groundcover Management Effects on Nitrate Leaching in an Irrigated Heavy-Clay Soil. Acta Hortic. 1997, 739–755. [Google Scholar] [CrossRef]
- Dong, S.; Scagel, C.F.; Cheng, L.; Fuchigami, L.H.; Rygiewicz, P.T. Soil Temperature and Plant Growth Stage Influence Nitrogen Uptake and Amino Acid Concentration of Apple during Early Spring Growth. Tree Physiol. 2001, 21, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Jabloun, M.; Schelde, K.; Tao, F.; Olesen, J.E. Effect of Temperature and Precipitation on Nitrate Leaching from Organic Cereal Cropping Systems in Denmark. Eur. J. Agron. 2015, 62, 55–64. [Google Scholar] [CrossRef]
- Thomsen, I.K.; Lægdsmand, M.; Olesen, J.E. Crop Growth and Nitrogen Turnover under Increased Temperatures and Low Autumn and Winter Light Intensity. Agric. Ecosyst. Environ. 2010, 139, 187–194. [Google Scholar] [CrossRef]
- Contosta, A.R.; Frey, S.D.; Cooper, A.B. Seasonal Dynamics of Soil Respiration and N Mineralization in Chronically Warmed and Fertilized Soils. Ecosphere 2011, 2, art36. [Google Scholar] [CrossRef]
- Patil, R.H.; Laegdsmand, M.; Olesen, J.E.; Porter, J.R. Sensitivity of Crop Yield and N Losses in Winter Wheat to Changes in Mean and Variability of Temperature and Precipitation in Denmark Using the FASSET Model. Acta Agric. Scand. Sect. B Soil Plant Sci. 2012, 62, 335–351. [Google Scholar] [CrossRef]
- Børgesen, C.D.; Olesen, J.E. A Probabilistic Assessment of Climate Change Impacts on Yield and Nitrogen Leaching from Winter Wheat in Denmark. Nat. Hazards Earth Syst. Sci. 2011, 11, 2541–2553. [Google Scholar] [CrossRef] [Green Version]
- Boman, B.J.; Battikhi, A.M. Growth, Evapotranspiration, and Nitrogen Leaching from Young Lysimeter-Grown Orange Trees. J. Irrig. Drain. Eng. 2007, 133, 350–358. [Google Scholar] [CrossRef]
- Sadowski, A.; Jadczuk, E. Results of 11-Year N-Fertiliser Trial in a Sour Cherry Orchard. Acta Hortic. 2001, 279–284. [Google Scholar] [CrossRef]
- Wrona, D.; Sadowski, A. Effects of Nitrogen Fertilisation in Young Apple Orchard. Acta Hortic. 1997, 481–486. [Google Scholar] [CrossRef]
- Rutkowski, K.; Łysiak, G.P. Weather Conditions, Orchard Age and Nitrogen Fertilization Influences Yield and Quality of ‘Łutówka’ Sour Cherry Fruit. Agriculture 2022, 12, 2008. [Google Scholar] [CrossRef]
- Haque, M.A.; Sakimin, S.Z. Planting Arrangement and Effects of Planting Density on Tropical Fruit Crops—A Review. Horticulturae 2022, 8, 485. [Google Scholar] [CrossRef]
- Cakmak, D.; Saljnikov, E.; Perovic, V.; Jaramaz, D.; Mrvic, V. Effect of Long-Term Nitrogen Fertilization on Main Soil Chemical Properties in Cambisol. In Proceedings of the 2010 19th World Congress of Soil Science, Soil Solutions for a Changing, Brisbane, Australia, 1–6 August 2010; pp. 291–293. [Google Scholar]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 978-94-010-1009-2. [Google Scholar]
- Smiley, R.W. Rhizosphere PH as Influenced by Plants, Soils, and Nitrogen Fertilizers. Soil Sci. Soc. Am. J. 1974, 38, 795–799. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Mengel, K. Ionic Balance in Different Tissues of the Tomato Plant in Relation to Nitrate, Urea, or Ammonium Nutrition. Plant Physiol. 1967, 42, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vang-Petersen, O. Fertilizing Sour Cherry {Prunus cerasus L. Cv. Stevnsbær) with N, P, and K. Tidsskr. Planteavl. 1977, 81, 346–350. [Google Scholar]
- Prsa, I.; Stampar, F.; Vodnik, D.; Veberic, R. Influence of Nitrogen on Leaf Chlorophyll Content and Photosynthesis of ‘Golden Delicious’ Apple. Acta Agric. Scand. Sect. B Plant Soil Sci. 2007, 57, 283–289. [Google Scholar] [CrossRef]
- Kurlus, R.; Łysiak, G. Rootstock Effect on Mineral Composition in Jonagored Apple Leaves; Estonian Agricultural University: Tartu, Estonia, 2000; pp. 68–71. [Google Scholar]
- Fallahi, E.; Mohan, S.K. Influence of Nitrogen and Rootstock on Tree Growth, Precocity, Fruit Quality, Leaf Mineral Nutrients, and Fire Blight in ‘Scarlet Gala’ Apple. HortTechnology 2000, 10, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Pacholak, E.; Zachwieja, M.; Zydlik, Z. Effect of Nitrogen Fertilization on the Content of Mineral Components in Soil, Leaves and Fruits of ‘Sampion’ apple Trees. Acta Sci. Pol. Hortorum Cultus 2004, 2, 207–217. [Google Scholar]
- Rutkowski, K.; Zydlik, Z.; Stachowiak, A. Effect of Tree Pruning Intensity on the Content of Mineral Components in the Sour Cherry Leaves of ‘Łutówka’. Folia Hortic. 2018, 30, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Fallahi, E.; Colt, W.M.; Baird, C.; Fallahi, B.; Chun, I. Influence of Nitrogen and Bagging on Fruit Quality and Mineral Concentrations of ‘BC2 Fuji’ Apple. HortTechnology 2001, 3, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Hansen, P. The Effects of Cropping on Uptake, Contents, and Distribution of Nutrients in Apple Trees. Tidsskr. Planteavl. 1971, 75, 615–625. [Google Scholar]
- Mészáros, M.; Hnátková, H.; Čonka, P.; Náměstek, J. Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple. Agronomy 2021, 11, 506. [Google Scholar] [CrossRef]
- Samuolienė, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between Apple Tree Rootstock, Crop-Load, Plant Nutritional Status and Yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Weinbaum, S.A.; Niederholzer, F.J.A.; Ponchner, S.; Rosecrance, R.C.; Carlson, R.M.; Whittlesey, A.C.; Muraoka, T.T. Nutrient Uptake by Cropping and Defruited Field-Grown ‘French’ Prune Trees. J. Am. Soc. Hortic. Sci. 1994, 119, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Sadowski, A.; Jadczuk, E. Effects of Nitrogen Fertilisation in a Sour Cherry Orchard. Acta Hortic. 1970, 475–480. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Matson, A.; Wigley, K.; Waller, L.; Dickie, I.A.; Whitehead, D.; Garrett, L. High Maintenance of Rhizosphere Soil C and N Equilibrium Regardless of Plant Species or Species Traits. Front. Soil Sci. 2021, 1, 762510. [Google Scholar] [CrossRef]
- Brust, G.E. Management Strategies for Organic Vegetable Fertility. In Safety and Practice for Organic Food; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–212. ISBN 978-0-12-812060-6. [Google Scholar]
- Watson, C.A.; Atkinson, D.; Gosling, P.; Jackson, L.R.; Rayns, F.W. Managing Soil Fertility in Organic Farming Systems. Soil Use Manag. 2002, 18, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Scalon, M.C.; Haridasan, M.; Franco, A.C. Influence of Long-Term Nutrient Manipulation on Specific Leaf Area and Leaf Nutrient Concentrations in Savanna Woody Species of Contrasting Leaf Phenologies. Plant Soil 2017, 421, 233–244. [Google Scholar] [CrossRef]
- Campbell, R.J.; Marini, R.P. Nitrogen Fertilization Influences the Physiology of Apple Leaves Subjected to European Red Mite Feeding. J. Am. Soc. Hortic. Sci. 1990, 115, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Mika, A.; Wawrzyńczak, P.; Buler, Z.; Krawiec, A.; Białkowski, P.; Michalska, B.; Plaskota, M.; Gotowicki, B. Results of Experiments with Densely-Planted Sour Cherry Trees for Harvesting with a Continuously Moving Combine Harvester. J. Fruit Ornam. Plant Res. 2011, 19, 31–40. [Google Scholar]
- Grafe, C.; Schuster, M. Physicochemical Characterization of Fruit Quality Traits in a German Sour Cherry Collection. Sci. Hortic. 2014, 180, 24–31. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Uysal, E. Effects of Nitrogen Fertilization on the Chlorophyll Content of Apple. Meyve Bilimi 2018, 5, 12–17. [Google Scholar]
- Kurlus, R.; Lysiak, G. Fertilizer Containing 5-Aminolevulinic Acid Affects Sour Cherry Fruit Quality and Biological Characteristics. Acta Hortic. 2014, 323–329. [Google Scholar] [CrossRef]
- Yakovenko, R.; Kopytko, P.; Pelekhatyi, V. The Content of Chlorophyll and Nutrients in Apple Leaves Depending on Long-Term Fertiliser. Sci. Horiz. 2021, 24, 93–98. [Google Scholar] [CrossRef]
- Wężyk, P.; Wertz, B.; Waloszek, A. Skaner hiperspektralny aisa (airborne imaging spectrometer for applications) jako narzędzie pozyskiwania informacji o ekosystemie leśnym (hiperspectral scanner aisa (airborne imaging spectrometer for applications)—Tool for forest ecosystem data gathering). Arch. Fotogram. Kartogr. Teledetekcji 2003, 13, 485–496. [Google Scholar]
- Bandurska, H. Akumulacja wolnej proliny jako przejaw metabolicznej reakcji roślin na działanie stresu wodnego. Wiad. Bot. 1991, 35, 35–46. [Google Scholar]
- Olszewska, M. Reakcja Wybranych Odmian Kostrzewy Lakowej i Tymotki Lakowej Na Stres Wodny. Acta Sci. Pol. Agric. 2003, 2, 141–148. [Google Scholar]
- Jaroszewska, A. Zawartość Barwników Asymilacyjnych w Liściach Drzew Pestkowych w Zależności Od Nawadniania i Nawożenia. Infrastrukt. Ekol. Teren. Wiej. 2011, 155–164. [Google Scholar]
- Szot, I.; Szot, P.; Lipa, T.; Sosnowska, B.; Dobrzański, B. Determination of physical and chemical properties of cornelian cherry (Cornus mas L.) fruits depending on degree of ripening and ecotypes. Acta Sci. Pol. Hortorum Cultus 2019, 18, 251–262. [Google Scholar] [CrossRef]
- Łysiak, G.; Kurlus, R.; Zydlik, Z.; Walkowiak-Tomczak, D. Apple Skin Colour Changes during Harvest as an Indicator of Maturity. Acta Sci. Pol. Hortorum Cultus 2014, 13, 71–83. [Google Scholar]
- Demirtas, N.M.; Bolat, I.; Ercisli, S.; Ikinci, A.; Olmez, H.; Sahin, M.; Altindag, M.; Celik, B. The Effects of Different Pruning Treatments on Seasonal Variation of Carbohydrates in Hacihaliloglu Apricot Cultivar. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 223–227. [Google Scholar] [CrossRef]
- Sanjay, K.S.; Sanjay, K.S.; Ram, R.S. Effects of Pruning Intensity on the Biochemical Status of Shoot Buds in Three Mango (Mangifera indica L.) Cultivars Planted at High Density. J. Hortic. Sci. Biotechnol. 2010, 85, 483–490. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | |
|---|---|---|---|---|---|---|
| Temperature before 7 days N fertilization | ||||||
| Air | 6.8 | 10.7 | 9.1 | 8.9 | 7.0 | 12.9 |
| Soil | 7.7 | 10.5 | 9.7 | 10.1 | 7.9 | 11.0 |
| Temperature after 7 days N fertilization | ||||||
| Air | 9.8 | 9.4 | 8.5 | 13.4 | 8.7 | 11.8 |
| Soil | 12.0 | 12.5 | 9.0 | 14.2 | 10.1 | 12.8 |
| Precipitation before 7 days N fertilization (mm) | ||||||
| Sum | 31.8 | 4.0 | 0.6 | 1.6 | 8.2 | 4.2 |
| Precipitation after 7 days N fertilization (mm) | ||||||
| Sum | 0.0 | 15.4 | 10.8 | 0.0 | 1.0 | 0.8 |
| April | ||||||
| Mean temperature (°C) | 7.9 | 11.2 | 8.8 | 11.5 | 8.9 | 7.9 |
| Sum of precipitation (mm) | 56.2 | 19.6 | 15.0 | 9.2 | 33.2 | 7.0 |
| Month | Precipitation (mm) | Temperature (°C) | ||||
|---|---|---|---|---|---|---|
| Mean 1982–2007 | Mean 2008–2013 | Change from Mean 1982–2007 in % | Mean 1982–2007 | Mean 2008–2013 | Change from Mean 1982–2007 | |
| January | 30.0 | 38.0 | +26 | −0.7 | −1.6 | −0.9 |
| February | 27.7 | 16.3 | −41 | −0.1 | −1.0 | −0.9 |
| March | 34.5 | 29.7 | −14 | 3.5 | 3.0 | −0.5 |
| April | 29.0 | 33.2 | +15 | 9.1 | 10.0 | 0.8 |
| May | 45.9 | 65.6 | +43 | 14.7 | 14.0 | −0.7 |
| June | 61.4 | 69.7 | +14 | 17.2 | 17.3 | 0.1 |
| July | 72.5 | 99.1 | +37 | 19.5 | 19.5 | 0.0 |
| August | 60.5 | 63.8 | +5.0 | 18.9 | 18.8 | −0.1 |
| September | 41.1 | 47.5 | +16 | 14.1 | 13.9 | −0.2 |
| October | 30.7 | 41.1 | +34 | 9.2 | 8.3 | −0.9 |
| November | 36.0 | 39.1 | +9.0 | 3.5 | 4.7 | 1.3 |
| December | 39.9 | 39.3 | −2.0 | 0.5 | −0.4 | −0.9 |
| Total | 509.2 | 609.20 | +14 | 9.1 | 8.9 | −0.2 |
| T1 | T2 | T3 | T4 | |||||
|---|---|---|---|---|---|---|---|---|
| 0–20 cm | 21–40 cm | 0–20 | 21–40 | 0–20 | 21–40 | 0–20 | 21–40 | |
| P30 | 0.16 | 0.41 * | −0.51 * | −0.17 | −0.10 | 0.17 | −0.41 * | −0.45 * |
| P14 | 0.09 | 0.33 * | −0.52 * | −0.27 * | −0.11 | 0.16 | 0.24 * | 0.12 |
| Ts | 0.27 * | 0.14 | 0.49 * | 0.11 | 0.26 * | 0.26 * | 0.17 | 0.03 |
| EWT | 0.50 * | 0.51 * | −0.08 | 0.54 * | 0.25 * | 0.26 * | 0.27 * | 0.18 |
| Pds | −0.38 * | −0.32 * | −0.52 * | −0.25 * | 0.18 | 0.27 * | 0.10 | −0.06 |
| WB | −0.53 * | −0.57 * | −0.52 * | −0.33 * | −0.27 * | −0.26 * | 0.07 | −0.24 * |
| Tair | 0.30 * | 0.20 | 0.49 * | 0.54 * | 0.01 | 0.22 | 0.06 | −0.10 |
| Orchard | Treatment | mg 100 g−1 DW Soil | pH in KCl | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | K | Mg | K/Mg | ||||||||
| OR 1 | N0 | 8.4 | b 1 | 8.5 | ab | 13.2 | b | 0.6 | bc | 6.7 | b |
| N60 | 9.5 | c | 7.8 | a | 11.7 | a | 0.7 | c | 6.7 | b | |
| N120 | 9.6 | c | 9.8 | cd | 11.9 | a | 0.8 | de | 6.5 | ab | |
| OR 2 | N0 | 7.1 | a | 7.8 | a | 11.8 | a | 0.7 | c | 6.4 | ab |
| N60 | 8.3 | b | 8.3 | ab | 11.0 | a | 0.8 | d | 6.2 | a | |
| N120 | 8.9 | bc | 10.5 | d | 10.8 | a | 1.0 | f | 6.1 | a | |
| OR 3 | N0 | 11.5 | e | 10.0 | cd | 15.5 | c | 0.9 | e | 6.6 | ab |
| N60 | 9.6 | c | 9.2 | bc | 15.8 | c | 0.5 | a | 6.4 | ab | |
| N120 | 11.8 | e | 13.6 | e | 15.7 | c | 0.6 | ab | 6.4 | ab | |
| Mean for orchard | OR 1 | 9.2 | b 2 | 8.7 | a | 12.3 | b | 0.7 | b | 6.6 | b |
| OR 2 | 8.1 | a | 8.9 | a | 11.2 | a | 0.8 | c | 6.2 | a | |
| OR 3 | 11.0 | c | 10.9 | b | 15.6 | c | 0.7 | a | 6.4 | ab | |
| Mean for treatment | N0 | 9.0 | a 3 | 8.8 | a | 13.5 | b | 0.7 | b | 6.5 | a |
| N60 | 9.1 | a | 8.4 | a | 12.8 | a | 0.7 | a | 6.4 | a | |
| N120 | 10.1 | b | 11.3 | b | 12.8 | a | 0.8 | c | 6.3 | a | |
| Orchard | Treatment | mg 100 g−1 DW Soil | pH in KCl | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | K | Mg | K/Mg | ||||||||
| OR 1 | N0 | 7.4 | cd 1 | 5.9 | a | 9.2 | bc | 0.6 | ab | 6.6 | bc |
| N60 | 7.3 | cd | 6.8 | b | 7.8 | a | 0.8 | c–e | 6.7 | c | |
| N120 | 7.6 | cd | 7.7 | c | 7.9 | a | 0.9 | ef | 6.7 | c | |
| OR 2 | N0 | 5.3 | a | 8.7 | d | 9.8 | b–d | 0.9 | ef | 6.0 | a |
| N60 | 6.4 | b | 7.7 | c | 9.0 | b | 0.9 | d–f | 6.2 | ab | |
| N120 | 7.2 | bc | 7.6 | c | 8.0 | a | 0.9 | f | 6.3 | a–c | |
| OR 3 | N0 | 8.2 | d | 8.3 | cd | 11.5 | e | 0.7 | bc | 6.2 | ab |
| N60 | 6.3 | b | 6.5 | ab | 10.5 | d | 0.6 | a | 6.5 | bc | |
| N120 | 9.3 | e | 7.6 | c | 10.2 | cd | 0.8 | cd | 6.4 | a–c | |
| Mean for orchard | OR 1 | 7.4 | b 2 | 6.8 | a | 8.3 | a | 0.8 | b | 6.7 | b |
| OR 2 | 6.3 | a | 8.0 | c | 9.0 | b | 0.9 | c | 6.2 | a | |
| OR 3 | 7.9 | b | 7.5 | b | 10.7 | c | 0.7 | a | 6.4 | a | |
| Mean for treatment | N0 | 7.0 | a 3 | 7.6 | b | 10.2 | b | 0.8 | a | 6.3 | a |
| N60 | 6.7 | a | 7.0 | a | 9.1 | a | 0.7 | a | 6.5 | a | |
| N120 | 8.0 | b | 7.7 | b | 8.7 | a | 0.9 | b | 6.5 | a | |
| Orchard | Treatment | Nutrient Content (% DW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | |||||||
| OR 1 | N0 | 2.14 | b 1 | 0.31 | e | 1.63 | d | 2.73 | f | 0.45 | ab |
| N60 | 2.36 | d | 0.18 | ab | 1.24 | bc | 2.71 | f | 0.48 | bc | |
| N120 | 2.65 | f | 0.17 | a | 1.43 | c | 2.39 | de | 0.43 | a | |
| OR 2 | N0 | 1.96 | a | 0.28 | d | 1.08 | ab | 2.01 | a | 0.52 | de |
| N60 | 2.18 | bc | 0.22 | c | 1.06 | ab | 2.22 | b–d | 0.55 | ef | |
| N120 | 2.30 | cd | 0.19 | ab | 1.02 | ab | 2.13 | ab | 0.50 | cd | |
| OR 3 | N0 | 1.99 | a | 0.28 | d | 1.10 | ab | 2.17 | a–c | 0.55 | ef |
| N60 | 2.18 | bc | 0.20 | b | 0.89 | a | 2.44 | e | 0.58 | f | |
| N120 | 2.51 | e | 0.17 | ab | 0.95 | a | 2.34 | c–e | 0.52 | de | |
| Mean for orchard | OR 1 | 2.39 | b 2 | 0.22 | a | 1.43 | b | 2.61 | c | 0.45 | a |
| OR 2 | 2.14 | a | 0.23 | a | 1.06 | a | 2.12 | a | 0.52 | b | |
| OR 3 | 2.23 | a | 0.22 | a | 0.98 | a | 2.32 | b | 0.55 | c | |
| Meant for treatment | N0 | 2.03 | a 3 | 0.29 | c | 1.27 | b | 2.30 | a | 0.51 | a |
| N60 | 2.24 | b | 0.20 | b | 1.06 | a | 2.46 | b | 0.53 | b | |
| N120 | 2.49 | c | 0.17 | a | 1.13 | a | 2.29 | a | 0.48 | a | |
| N | P | K | Ca | Mg | |
|---|---|---|---|---|---|
| T | 0.01 | 0.09 | 0.52 * | −0.66 * | −0.61 * |
| T max. | −0.06 | 0.12 | 0.55 * | −0.68 * | −0.62 * |
| T min. | 0.18 | −0.17 | −0.45 * | 0.55 * | 0.50 * |
| Precipitation | 0.01 | 0.08 | 0.52 * | −0.65 * | −0.61 * |
| ET | 0.09 | −0.14 | −0.54 * | 0.67 * | 0.62 * |
| BW | 0.21 * | −0.18 | −0.39 * | 0.46 * | 0.42 * |
| N-NO3 (0–20 cm) | 0.53 * | −0.08 | −0.05 | −0.07 | −0.16 |
| Total N (0–20 cm) | 0.53 * | −0.02 | −0.01 | −0.09 | −0.18 |
| N-NO3 (20–40 cm) | 0.58 * | −0.19 * | 0.00 | −0.04 | −0.20 * |
| Total N (20–40 cm) | 0.60 * | −0.20 * | −0.03 | −0.04 | −0.19 |
| P (0–20 cm) | 0.31 * | −0.02 | 0.23 * | −0.19 | −0.06 |
| K (0–20 cm) | 0.40 * | −0.17 | −0.09 | 0.02 | 0.21 * |
| Mg (0–20 cm) | −0.05 | 0.08 | 0.10 | 0.04 | 0.15 |
| K/Mg (0–20 cm) | 0.28 * | −0.09 | −0.13 | −0.09 | 0.09 |
| P (20–40 cm) | 0.43 * | −0.01 | 0.31 * | −0.17 | −0.37 * |
| K (20–40 cm) | 0.24 * | −0.10 | −0.18 | −0.02 | 0.15 |
| Mg (20–40 cm) | −0.22 * | 0.21 * | −0.07 | −0.01 | 0.39 * |
| K/Mg (20–40 cm) | 0.38 * | −0.22 * | −0.05 | −0.02 | −0.19 |
| Orchard | Treatment | Leaf Area (cm2) | TCSA | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2007 | 2013 | Increase 2007–2013 | |||||||
| OR 1 | N0 | 29.0 | a 1 | 56.5 | b | 84.1 | bc | 27.7 | ab |
| N60 | 34.1 | bc | 68.0 | c | 102.4 | d | 34.4 | a–d | |
| N120 | 34.7 | bc | 54.9 | b | 92.1 | c | 37.2 | b–d | |
| OR 2 | N0 | 28.0 | a | 43.5 | a | 69.3 | a | 25.9 | a |
| N60 | 32.8 | b | 46.8 | a | 78.0 | ab | 31.2 | a–c | |
| N120 | 34.9 | bc | 46.7 | a | 88.6 | c | 41.9 | d | |
| OR 3 | N0 | 28.7 | a | 41.5 | a | 72.2 | a | 30.8 | a–c |
| N60 | 32.6 | b | 43.1 | a | 78.7 | ab | 35.6 | a–d | |
| N120 | 36.3 | c | 39.7 | a | 77.9 | ab | 38.2 | cd | |
| Mean for orchard | OR 1 | 32.6 | a 2 | 59.8 | c | 92.9 | b | 33.1 | a |
| OR 2 | 31.9 | a | 45.7 | b | 78.7 | a | 33.0 | a | |
| OR 3 | 32.5 | a | 41.4 | a | 76.3 | a | 34.9 | a | |
| Mean for treatment | N0 | 28.6 | a 3 | 47.1 | a | 75.2 | a | 28.1 | a |
| N60 | 33.2 | b | 52.6 | b | 86.4 | b | 33.8 | b | |
| N120 | 35.3 | c | 47.1 | a | 86.2 | b | 39.1 | c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rutkowski, K.; Łysiak, G.P. Effect of Nitrogen Fertilization on Tree Growth and Nutrient Content in Soil and Cherry Leaves (Prunus cerasus L.). Agriculture 2023, 13, 578. https://doi.org/10.3390/agriculture13030578
Rutkowski K, Łysiak GP. Effect of Nitrogen Fertilization on Tree Growth and Nutrient Content in Soil and Cherry Leaves (Prunus cerasus L.). Agriculture. 2023; 13(3):578. https://doi.org/10.3390/agriculture13030578
Chicago/Turabian StyleRutkowski, Krzysztof, and Grzegorz P. Łysiak. 2023. "Effect of Nitrogen Fertilization on Tree Growth and Nutrient Content in Soil and Cherry Leaves (Prunus cerasus L.)" Agriculture 13, no. 3: 578. https://doi.org/10.3390/agriculture13030578
APA StyleRutkowski, K., & Łysiak, G. P. (2023). Effect of Nitrogen Fertilization on Tree Growth and Nutrient Content in Soil and Cherry Leaves (Prunus cerasus L.). Agriculture, 13(3), 578. https://doi.org/10.3390/agriculture13030578