

An Overview of Comet Assay Application for Detecting DNA Damage in Aquatic Animals


 ,
,  , and
, and
Abstract
1. Introduction
2. Criteria for Selection of Papers
3. Evaluation of DNA Damage and Repair Processes
4. Comet Assay
4.1. History of Comet Assay
4.2. Advantages of Comet assay
4.3. Disadvantages of Comet Assay
4.4. Applications of the Comet Assay
4.5. Genotoxicity Testing and Ecological Monitoring
4.6. Protocol of Comet Assay
- (i)
- Isolation of cells and preparation of single-cell suspensions;
- (ii)
- Embedding of the cells in agarose;
- (iii)
- Cell lysis;
- (iv)
- Incubation of the nucleoids with lesion-specific enzyme (for the enzyme-modified comet assay) or with cell or tissue extract (for the in vitro DNA repair assay);
- (v)
- Alkaline treatment;
- (vi)
- Electrophoresis;
- (vii)
- Neutralization;
- (viii)
- Staining and visualization;
- (ix)
- Scoring and data analysis [8].
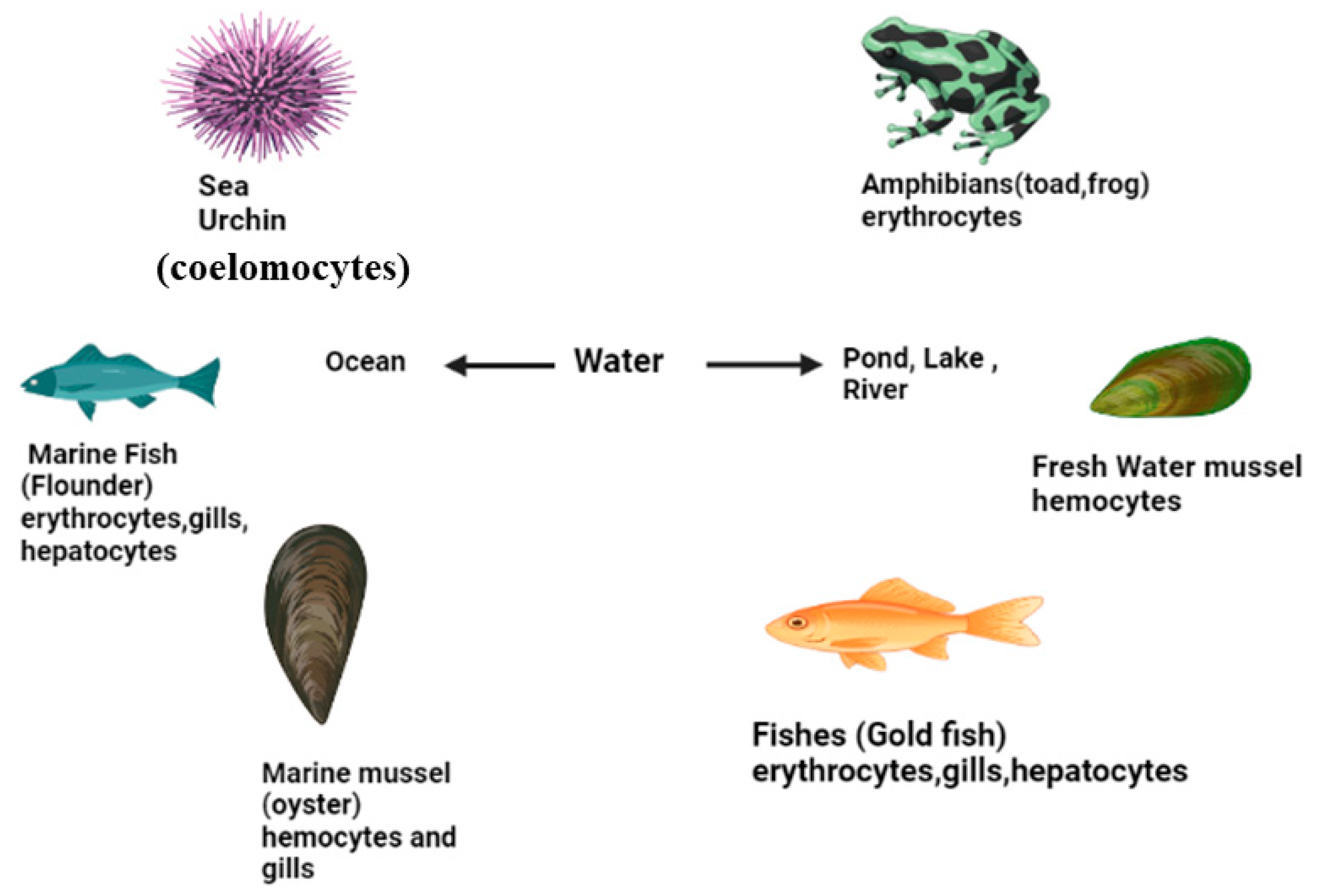
4.7. Invertebrates
4.8. Comet Assay in Mollusca
4.9. Comet Assay in Arthropoda

{kind=link}
{kind=link}
| Location | Phylum | Organism | Cell Type | Concentration Range | Exposure Time | Descriptor | Agent | Response | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Siolim, Goa, India | Mollusca | Paphiamalabarica Meretrix casta | Hemolymph | Gama (2, 4, 6, 8, and 10 Gray Ethyl Methanesulfonate (18, 32, and 56 mg/L) | 24, 48, 72 h | % Tail DNA | Ethyl Methanesulfonate (EMS) Gamma radiation | Both EMS and gamma radiation indicated a dose-dependent manner. DNA damage for both species was seen in the following order: 24 > 48 and 72 h. | [64] |
| Wenchang, Hainan Province, China | Arthropoda | Litopenaeusvannamei | Gill, hepatopancreas and hemolymph | 3.0 and 1.5 ppm | 24 h | % Tail DNA | Hypoxia | Dose-dependent relation was observed, i.e., DNA damage increased at all concentrations. | [65] |
| Santa Barbara, California, United States of America | Mollusca | Mytilus galloprovincialis Mytilus californianus | Haemocytes | Heat stress: From 13 °C to 24 °C, 28 °C and 32 °C for Mytilus californianus For Mytilus galloprovincialis, 13 °C to 28 °C and 32 °C Cold stress: 2 °C, 6 °C | 0.5, 2 and 8 h | % Tail DNA | Temperature | At 32 °C single-strand breakage and double-strand breakage. DNA in tail length trend: Mytilus californianus > Mytilus galloprovincialis. Cold stress: temperature and time-dependent double-strand breakage in both species. | [87] |
| Zhoushan, Zhejiang Province, China | Mollusca | Mytilus coruscus | Haemocytes | 2 and 8 μg/L | 18 days | OTM | Copper | A time- and concentration-dependent pattern of DNA damage For 2 µ/L order of OTM value:18 > 12 > 6 days For 8 µ/L OTM value is greater after six days | [67] |
| Bizerta Lagoon, Tunisia | Mollusca | Mytilus galloprovincialis | Digestive gland cells | 75 Nm;19 µg/L per animal | 24, 48, and 72 h | % Tail DNA | Benzo[a]pyrene | DNA damage was maximum at 72 h after exposure | [68] |
| Unpolluted freshwater pond, Pakistan | Mollusca | Anodonta anatina | Gill cells | 120 µg/L 240 µg/L 360 µg/L | 15 days | OTM | Lead Chromium Copper | Concentration-wise DNA Damage: Low dose > medium dose > highest dose. Metal-wise DNA damage: Copper and Lead > Chromium and metal combination (Lead + Chromium + Copper) | [71] |
| Taishan, Guangzhou, China | Arthropoda | Scylla paramamosain | Haemocytes | 1.25, 2.5, 5 and 10 mg/L | 72 h | OTM | Cadmium | OTM value was highest at 10 mg/L | [88] |
| Local market in Xichang City, China | Arthropoda | Procambarusclarkii | Haemocytes | 7, 14, and 28 ng/L | 96 h | OTM | Deltamethrin | Olive tail moment (OTM) was highest at 28 ng/L | [89] |
| Ennore Estuary, Chennai city, India | Mollusca | Perna viridis | Gill and hepatopancreatic cells | Copper 65.1 µg/g Lead 0.123 µg/g Zinc 823 µg/g Cadmium 0.047 µg/g Manganese 10.3 µg/g Iron 191 µg/g | 48 h | TM, TL, % Tail DNA | Copper, Lead, Zinc, Cadmium, Manganese and Iron | Cd and Pb were negatively associated and accumulated at lower conc., while Fe showed a higher positive correlation compared to other metals, indicating enhanced accumulation. The trend of Comet Tail: Gills > hepatocreatic cells The trend of %Tail DNA was: Gills > hepatocreatic cells The trend of Tail Moment: Gills < hepatocreatic cells | [72] |
| Elanfoshy bay in Alexandria, Egypt | Mollusca | Lithophagalithophaga | Gill cells | 5 and 20 μg/L | 28 days | OTM, % Tail DNA | Silver oxide nanoparticles | Concentration and time-dependent DNA damage. The highest was at 20 μg/L | [83] |
| Ecotoxicology Laboratory of the Federal Technological University of Paraná, Brazil | Arthropoda | Daphnia magna | Whole organism (homogenized) cells | 0.23 and 0.47 μg/L | 48 h | % Tail DNA | Malathion | Results showed DNA damage at each concentration | [90] |
| Betanzos estuary (North coast of Galicia), Spain, Europe | Mollusca | Mytilus galloprovincialis | 2 gills per individual | 1:500 and 2:500 | 12 days | TL, %TDNA, and OTM | Spilled prestige crude oil | DNA Damage: 1:500 > 2:500 because of more accumulation of TPAH | [91] |
| Devon Great Consols (DGC), Britain | Annelid | Lumbricus rubellus, Dendrodrilus rubidus and Lumbricus terrestris | Soil sample, intestine | 204 mg/kg to 9025 mg/kg | 1, 3, 4, and 5 weeks | % Tail DNA | Arsenic | DNA damage in Earthworms living in As-contaminated soils at DGC < Non-Native Earthworms | [92] |
| Arabian Sea, Goa, India | Mollusca | Perna viridis | Gonad tissue | 2, 5, 10 and 15 µg per kg | 16 days | Percent comet nuclei and deformed nuclei | Cigar tobacco | Dose-response relationship was highest on day 12 | [93] |
| Rio de Janeiro, Brazil | Arthropoda | Amphipod Quadrivisio aff. lutzi (Gammaridea) | Blood cells | 6.25, 12.5, 25, 50 and 100% | 72 h | TL | Water-soluble fraction of heavy oil | DNA damage trend: 6 and 48 h > 24 and 72 h | [94] |
| The Bermuda Institute of Ocean Sciences | Echinodermata | Lytechinus variegatus, Lucunter lucunter, Tripneustes ventricosus and Isostichopus badionotus | Coelomocytes | (0–100 mM) (0–9999 J/m2) | 24 h | %DNA Damage | Hydrogen peroxide UV-C(Ultraviolet-C) | There was a definite concentration- and dose-dependent rise in the DNA damage for all of the examined echinoderm species. The trend of the % DNA damage levels in different coelomocytes of the tested species is as follows: I. badionotus < L. variegatus L. lucunter, T. ventricosus | [95] |
| Lake Trasimeno, Italy | Mollusca | Dreissena polymorpha | Haemocytes | (0.5 and 5 g/mL) (10 g/mL) 4, 18, 28 and 37 °C | 15 h | TL and OTM | Melphalan Hypochlorite Sodium hypochlorite Temperature | As the reaction is temperature dependent, at 4 °C, DNA damage from Hypochlorite > Melphalan and Sodium hypochlorite. At 18°C, DNA damage from Sodium Hypochlorite > Hypochlorite and Melphalan. At 28 °C, 5 g/mL, and 5 min electrophoresis DNA damage from Melphalan > Hypochlorite and Sodium hypochlorite. At 37 °C, DNA damage from Melphalan (5 g/mL) > 0.05 g/mL of Melpahlan and Hypochlorite. | [96] |
| Valencia, Spain | Arthropoda | Chironomus riparius | Fourth instars larvae | Bisphenol A (0.5 and 3 mg/L) Nonylphenol (1, 10 and 100 g/L) Pentachlorophenol (25 and 250 g/L) Tributyltin (0.1, 1 and 10 ng/L) Triclosan (10, 100 and 1000 g/L) | 24 and 96 h | TL, %TDNA, and OTM | Bisphenol A Nonylphenol Pentachlorophenol Tributyltin Triclosan | Time and concentration-dependent variations were significant, i.e., DNA damage was increased with an increase in concentration and time of exposure. | [86] |
| Northern Croatia | Mollusca | Dreissena polymorpha | Haemocytes | 10, 80, 100 and 150 mg/L | 7 days | TL, %TDNA, and OTM | Pentachlorophenol | DNA damage at 80, 100 and 150 mg/L > 10 mg/L | [97] |
| Pune, India | Annelid | Dichogaster curgensis | Coelomocytes | In vivo (1, 3, 10, 30, 70, 100 ppm) In vitro (1, 3, 10, 30, 70 and 100 ppm) | In vivo (14 days) In vitro (1 h) | TL | Cr (Chromium) (VI) | In vivo exposure revealed that DNA damage at 30 ppm > 70 and 100 ppm, while in vitro exposure showed dose-dependent DNA damage. It is hypothesized that a decrease in DNA mobility brought on by the formation of numerous cross-links may be the cause of loss in arbitrary units at greater concentrations. | [98] |
| Korea Institute of Toxicology, Daejeon, Korea | Arthropoda | Daphnia magna, and Chironomus riparius | Erythrocytes | Daphnia magna (0.3, 3, and 30 µg/L for Nonlyphenol and BPA) Chironomus riparius (1, 10 and 100 µg/L for Nonlyphenol and 5, 50 and 500 µg/L for bisphenol A) | 24 h | OTM | Nonlyphenol Bisphenol A | Only for Daphnia magna, OTM at 3 and 30 µg /L > 0.3 µg/L of Bisphenol A exposure. For Chronomus riparius, they statistically increased at all doses of both chemicals examined. | [99] |
| Rio Gola, Northeast-Italy | Arthropod | Chironomus riparius | Fourth-instar larvae | 0.05, 1, and 25 mg/L | 3 h | TL, %TDNA, and OTM | Copper | Significant increases in all the comet parameters at concentrations more than 1 mg/L serve as evidence that copper can cause DNA damage even at sublethal doses. | [100] |
| Anzali wetland, North of Iran | Mollusca | Anodonta cygnea | Haemolymph and gill cells | 0.25, 0.5, and 1.0 ppm | 10 days | TL | Crude oil | Statistically significant increases in DNA damage and micronuclei were reported with 0.25, 0.5, and 1.0 ppm of crude oil. | [101] |
| Randaberg, Norway | Echinodermata and mollusca | Strongylocentrotus droebachiensis and Mytilus edulis Lamellibranchiata | Sea urchin coelomocytes and mussel haemocytes | Sea urchins (0.06 and 0.25 mg/L) Mussels (0.015, 0.06, and 0.25 mg/L) | Sea urchin (4 weeks) Mussel (5 weeks) | Percentage of DNA in comet tail | Crude oil | DNA concentration in the comet tails increased significantly with oil concentrations of 0.06 and 0.25 mg/L. Mussels subjected to oil concentrations up to 0.06 mg/L revealed a considerable concentration-related rise in the comet tail’s DNA %. | [102] |
| Kat O, Hong Kong | Mollusca | Perna viridis | Haemocytes | 0, 0.3, 3, and 30 µg/L | 0, 1, 3, 6 and 12 days | TL, %TDNA, and OTM | Water-borne benzo[a]pyrene | For 0.3 µg/L TL, %TDNA and OTM at day 0–3 > 6 and 12 days. For 3 and 30µg/L TL, %TDNA and OTM at day 0–6 > 6–12 days. | [103] |
| Lake Ponikve Krk Island (Northern Adriatic Sea) | Annelid | Hirudo verbana | Leech haemocytes | 0.5 L | 30, 60, 180 days | TL, %TDNA, and OTM | Aluminium contaminated sediment and water | DNA damage at 180 > 60 > 30 days | [104] |
| BiAzerta Lagoon, Tunisia | Mollusca | Mytilus galloprovincialis | Digestive gland cells | 75 nM; 19 mg/L per animal | 24, 48, and 72 h | %TDNA | Benzo[a]pyrene | DNA damage at 72 h > 48 > 24 h | [105] |
| Marennes-Oléron Bay, France | Mollusca | Crassostrea gigas | Hemocytes, gills and spermatozoa | 0.4 and 0.6 μg/L 0.005% for solvent control | Two 7-day exposures | % tDNA | Diuron Acetonitrile | DNA damage in Gills, Hemocytes and Spermatozoa: Diuron > Control groups | [106] |
4.10. Vertebrates
4.11. Comet Assay in Fish
| Location | Phylum | Organism | Descriptor | Cell Type | Concentration Range | Exposure Time | Agent | Response | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Gorukle, Bursa, Turkey | Chordata | Carassius auratus | TL | Erythrocytes | 5, 10, and 15 μg/L | 2, 4, and 6 days | Gesaprim Atrazine | Gesaprim increased proportion of damaged cells in the peripheral erythrocytes of Carassius auratus. However, no significant effect on erythrocytes was found. | [114] |
| Mandapam, southeast coast of India | Chordata | Therapon jarbua | % Tail DNA | Gill, kidney, and erythrocytes | 0.125, 0.25 ppm | 96 h | Mercuric chloride | At concentrations 0.125 and 025 ppm: Gill cells > kidneys and blood | [115] |
| CIFA’s (Central Institute of Freshwater Aquaculture) culture ponds in Bhubaneswar, India | Chordata | Labeo rohita | DNA damage (migration comet tail length and relative DNA content of the comet tail) | Blood, liver, and gill cells | 0.001, 0.002, 0.01 ppm | 96 h | Phorate | After 72 h, ↓DNA damage DNA breaks order: Liver > Gills and blood | [116] |
| Hatchery of the Freshwater Fisheries Institute of Lishui Zhejiang Province, China | Chordata | Cyprinus carpio | DNA damage percentage, TL, and TM | Liver cells | 0.41, 0.52, 0.69, 1.03 and 2.06 mg/L | 7 days | Cadmium | At concentrations (0.41 mg/L), ↓SOD activity, ↑ MDA and GSH levels These findings showed that cadmium (Cd) caused DNA damage in Cyprinus carpio liver. At concentration > 0.41mg/L all parameters were increased. | [117] |
| Commercial fish seed hatchery, India | Chordata | Catla catla | % Tail DNA TL TM | Erythrocytes | 30, 60, 90, 150 mg/L | 7, 14, and 21 days | Chromium | A significant (p 0.001) increase in % Tail DNA was observed after 21 days of exposure | [118] |
| Taian, Shandong, China | Chordata | Danio rerio | OTM | Liver cells | 0.3, 1.25, and 5 mg/L | 28 days | Imidacloprid | At the 21st day of exposure, at concentrations (of 1.25 and 5 mg/L), ROS generation and MDA contents were increased. Both time and dosage were factors in DNA damage. | [66] |
| Fish Seed Hatchery, Faisalabad | Chordata | Labeo rohita, Cirrhinus mrigala, Catla catla Ctenopharyngodon idella | Percent damaged cells, genetic damage index (GDI) and cumulative tail length | Erythrocytes | 17, 25, 33 and 50% LC50 | 30 days | Arsenic, Copper, and Zinc | The order of percentage damage in the erythrocytes of all four fish species is as follows: C. mrigala > L. rohita > C. idella > C. catla. L. rohita tail length revealed the only non-significant variation (C. mrigala > L. rohita > C. catla > C. idella) as exposure dramatically increased GDI, followed by exposures to Zn and Copper. | [119] |
| Rosario, Argentina laboratory | Chordata | Danio rerio | DNA damage (DNA damage score (Damage Index, DI)) | Gill cells | 0.3 or 0.6 µg/L | 12 days | Cypermethrin | The DNA damage index in the gill cells >dose- and time-dependent pathways. | [120] |
| China | Chordata | Gobio cyprisrarus | Tail DNA%, OTM | Liver cells | 0.1 mg/L 0.5 mg/L 2.0 mg/L | 60 days | Imidacloprid, Nitenpyram, and Dinotefuran | The DNA damage assay results showed that treatments with 2.0 mg/L imidacloprid significantly increased tail moments whereas treatments with 0.5, 2.0, 2.0 mg/L imidacloprid, 2.0 mg/L nitenpyram, and all treatments with dinotefuran significantly increased tail DNA%. Furthermore, treatments with 0.5 and 2.0 mg/L imidacloprid and 2.0 mg/L dinotefuran markedly increased the olive tail moments. | [121] |
| Water Treatment Station SANEAGO, in Goiânia city, GO, Brazil | Chordata | Poecilia reticulata | TL, TM, and OTM | Brain, Liver, and blood cells | 0.5, 1.0, and 1.5 μg/L | 24 and 96 h | Cylindrospermopsin | After 96 h, an increase in DNA damage was seen in peripheral blood cells using all three doses. | [122] |
| Snidjers Scientific, Tilburg, Nederland | Chordate | Oryzias latipes | TD% | Medaka cells | Acute toxicity (CdCl2 (0.1, 1, 10 and 100 μM) or Flu (2, 10 and 50 μM)) In vivo (CdCl2 (0.03, 0.1, 0.3, and 1 μM) or Flu (2, 10, and 50 μM)) In vitro (H2O2 (1, 10, and 100 μM) and (CdCl2 (0.03, 0.1, 0.3 and 1 μM) in 400 μL of Minimum Essential Medium (MEM) | 24 h | CdCl2 or Flu | Findings indicated that all tested chemicals significantly increased the proportion of damage in DNA of fragmented cells than the control one. | [123] |
| Eastern Croatia, Europe | Chordate | Cyprinus carpio | %TDNA | Erythrocytes | Nil | 3 weeks | Wastewaters, paper and pulp mill industry, chemicals (detergent and soap, textile) and metal industries | The results of comet assay revealed a statistically significant increase in the DNA damage in carp erythrocytes. | [124] |
| McElroy laboratory, The Ohio State University, United States | Chordate | Oryzias latipes | OTM | Embryos | 10 μg/L(final) | 7-day exposure | Polycyclic Aromatic Hydrocarbons (PAHs) Acenaphthenequinone and 7,12-Benz[a]anthracenquinone | After 48 h of exposure at the lowest tested dose (5 g/L), DNA damage was increased. | [125] |
| River Rhine, Germany | Chordate | Danio rerio | The tail moments, which are calculated as the ratio of the tail’s mean migration distance to its relative DNA content | Embryonic cells | 600 mg/mL = 100% (1:1), 300 mg/mL = 50% (1:2), and 150 mg/mL = 25% (1:4) 40, 20, 10 and 5 mg | 72 h | Exposure to whole (freeze-dried) sediment, exposure to organic sediment extracts | Only four native soils had genotoxic potential, compared to 17 out of 18 sediment extracts that significantly damaged the embryo cells’ DNA. The organic extracts, on the other hand, appear to have enhanced quantities of even barely soluble compounds | [126] |
| Coast of Gwangyang Bay, Korea | Chordate | Paralichthys olivaceus | TL | Whole blood | 5, 10, 50 and 100 ppb | 2 h for in vivo (0, 2, and 4 days | PAHs (Polycyclic Aromatic Hydrocarbons) BaP’s(benzo[a]pyrene, fluoranthene, anthracene, pyrene and phenanthrene) and sediments and also in vivo benzo[a]pyrene is used | Five groups treated with PAHs at concentrations of 50 and 100 ppb showed considerably shorter tail lengths than the control group. BaP’s genotoxic effects were associated with exposure time and concentration. Significant variations in DNA breakage between cells exposed to sediment extracts or PAHs and non-exposed control were seen throughout the investigation. | [127] |
| Zebrafish International Resource Center (ZIRC), United States | Chordate | Danio rerio | TL, %TDNA, and OTM | Embryos | 0 (control or vehicle), 250, 350, 625, 850, and 1250 μg/mL | 2, 4, 6, 12, 24, and 48 h | Monceren 250 SC | Caused DNA damage, as evidenced by the three genotoxicity parameters significantly increasing in zebrafish embryos. The survival percentage of larvae embryos was down to 40–45% after 48 h following fungicide treatment. | [128] |
| Fish Hatchery Station of the State University of Londrina, Brazil | Chordate | Prochilodus lineatus | TL | Erythrocytes and liver cells | 1.25, 12.5, 125, and 1250 mg /L | 120 h | Midacloprid (IMI) | Integrated Biomarker Response (IBR) index revealed: Liver and kidney > gills. | [129] |
| Santa Fe Province, Argentina | Chordate | Caiman latirostris | TL | Erythrocytes | 50, 100, 200, 300, 400, 500, 750, 1000, 1250 and 1750 µg/egg | 68 and 70 days in Experiment 1 and 66 and 77 days in Experiment 2 | Roundup | The MN test and the comet assay findings proved concentration-dependent and negative impacts. | [130] |
| Southern Brazil | Chordate | Gray mullet (Mugil sp.) and sea catfish (Netuma sp.) | Image length (IL), damage index (DI)and damage frequency (DF) | Erythrocytes | 2, 4, and 8 × 10−5 M | 1, 2, 6 and 24 h | Methyl methanesulfonate (MMS) | Baseline DNA damage enhanced at higher temperatures, as did MMS-induced DNA damage, and there was a definite dose/time response to MMS therapy. | [131] |
| Santa Fe city, Argentina | Chordate | Prochilodus lineatus | Comet tail and % of TDNA | Erythrocytes | 0.300, 0.150, 0.075 and 0.000 μg/L | 96 h | Cypermethrin | DNA damage was considerably higher at all Cypermethrin doses evaluated. | [132] |
| Siolim, Goa | Chordate | Channa punctatus (Bloch) | % tail DNA | The gills, kidneys, and lymphocytes | 1.48, 0.74, and 0.59 ppm | 0, 1, 3, 7, 15, 22 and 29 days (in vivo) Sub lethal (96 h) | Malathion | Up to day 3, DNA damage was seen in all of the tissues and increased as a function of concentration, followed by a nonlinear decline as exposure time increased. | [133] |
| Local outlets, India | Chordate | Clarias batrachus | TL | Erythrocytes | 2,4-D 25–75 ppm Butachlor 1–2.5 ppm | 48, 72, and 96 h | Herbicides: 2,4-D and Butachlor | The greatest tail length observed at the highest concentration and longer period of 2, 4-D (9.59 mm) and Butachlor indicated a concentration-related and time-dependent increase. | [134] |
| Toulouse, France’s Paul Sabatier University’s Department of Developmental Biology | Chordate | Xenopus laevis; Pleurodeles Waltl | tDNA, TL, OTM | Erythrocytes | Captan (fungicide) 2–125 mg/L; 62.5 mg/L | 1–12 days | Captan | From the first day of exposure, comet assay revealed that captan was genotoxic. | [135] |
| Toulouse, France’s Paul Sabatier University’s Department of Developmental Biology | Chordate | Xenopus laevis; Pleurodeles waltl | tDNA, TL, OTM | Erythrocytes | Cd 2–50 mg/L | 1–12 days | Cadmium | Comparing the findings of comet assay and micronuclei test, genotoxic effects were taken into account, along with the Cadmium concentrations used and the exposure duration. According to the comet assay, Cd was genotoxic from the first day of exposure. | [136] |
| Sweden | Chordate | Oncorhynchus mykiss | Tail DNA | Erythrocytes | 0.5% | 7 days | Algal extracts (Polysiphonia fucoides) | Results revealed substantial increases in DNA single-strand breaks equivalent to those caused by an in vivo exposure to 20 mg/kg B[a]P, but no changes in double-strand breaks or apoptotic cells were noted. | [137] |
| Uttar Pradesh, India | Chordate | Channapunctatus | %Tail DNA | Gill and kidney tissues | 4–10 ppb | 24–96 h | Endosulfan (pesticide) | In both tissues, there were dose-dependent reactions to DNA damage. Gill cells >kidney cells. | [138] |
| Byford, Stavanger, Norway | Chordate | Symphodus mellops | %Tail DNA | Erythrocytes | 2 mg/L | 7 days | Styrene | Fish and mussel blood cells’ comet tail DNA content was increased by styrene by 1.8 and 1.6 times, respectively | [139] |
4.12. In Vivo and In Vitro Comet Assay
4.13. Critical Factors Influencing Comet Assay Performance
4.14. The Comet Assay Modifications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pollack, N.; Cunningham, A.R.; Rosenkranz, H.S. Environmental persistence of chemicals and their carcinogenic risks to humans. Mutat. Res. 2003, 528, 81–91. [Google Scholar] [CrossRef]
- Zubair, M.; Ahmad, M.; Saleemi, M.K.; Gul, S.T.; Ahmad, M.; Martyniuk, C.J.; Ullah, Q.; Umar, S. Sodium arsenite toxicity on hematology indices and reproductive parameters in Teddy goat bucks and their amelioration with vitamin C. Environ. Sci. Pollut. Res. 2020, 27, 15223–15232. [Google Scholar] [CrossRef]
- Naz, S.; Mansouri, B.; Chatha, A.M.M.; Ullah, Q.; Abadeen, Z.U.; Khan, M.Z.; Khan, A.; Saeed, S.; Bhat, R.A. Water quality and health risk assessment of trace elements in surface water at Punjnad Headworks, Punjab, Pakistan. Environ. Sci. Pollut. Res. 2022, 29, 61457–61469. [Google Scholar] [CrossRef]
- Frenzilli, G.; Nigro, M.; Lyons, B.P. The Comet assay for the evaluation of genotoxic impact in aquatic environments. Mutat. Res. 2009, 681, 80–92. [Google Scholar] [CrossRef]
- Evenson, D.P.; Kasperson, K.; Wixon, R.L. Analysis of sperm DNA fragmentation using flow cytometry and other techniques. Soc. Reprod. Fertil. Suppl. 2007, 65, 93–113. [Google Scholar]
- Møller, P.; Azqueta, A.; Boutet-Robinet, E.; Koppen, G.; Bonassi, S.; Milić, M.; Gajski, G.; Costa, S.; Teixeira, J.P.; Costa Pereira, C.; et al. Minimum Information for Reporting on the Comet Assay (MIRCA): Recommendations for describing comet assay procedures and results. Nat. Protoc. 2020, 15, 3817–3826. [Google Scholar] [CrossRef]
- Lee, R.F.; Steinert, S. Use of the single cell gel electrophoresis/comet assay for detecting DNA damage in aquatic (marine and freshwater) animals. Mutat. Res. 2003, 544, 43–64. [Google Scholar] [CrossRef]
- Møller, P.; Loft, S.; Ersson, C.; Koppen, G.; Dusinska, M.; Collins, A. On the search for an intelligible comet assay descriptor. Front. Genet. 2014, 5, 217. [Google Scholar] [CrossRef]
- Azqueta, A.; Gutzkow, K.B.; Brunborg, G.; Collins, A.R. Towards a more reliable comet assay: Optimising agarose concentration, unwinding time and electrophoresis conditions. Mutat. Res. 2011, 724, 41–45. [Google Scholar] [CrossRef]
- Azqueta, A.; Muruzabal, D.; Boutet-Robinet, E.; Milic, M.; Dusinska, M.; Brunborg, G.; Møller, P.; Collins, A.R. Technical recommendations to perform the alkaline standard and enzyme-modified comet assay in human biomonitoring studies. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 843, 24–32. [Google Scholar] [CrossRef]
- Ostling, O.; Johanson, K.J. Microelectrophoretic study of radiation-induced DNA damages in individual mammalian cells. Biochem. Biophys. Res. Commun. 1984, 123, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P.; Durand, R.E. Heterogeneity in radiation-induced DNA damage and repair in tumor and normal cells measured using the “comet” assay. Radiat. Res. 1990, 122, 86–94. [Google Scholar] [CrossRef]
- Burlinson, B.; Tice, R.R.; Speit, G.; Agurell, E.; Brendler-Schwaab, S.Y.; Collins, A.R.; Escobar, P.; Honma, M.; Kumaravel, T.S.; Nakajima, M.; et al. Fourth International Workgroup on Genotoxicity testing: Results of the in vivo Comet assay workgroup. Mutat. Res. 2007, 627, 31–35. [Google Scholar] [CrossRef]
- Gedik, C.M.; Ewen, S.W.; Collins, A.R. Single-cell gel electrophoresis applied to the analysis of UV-C damage and its repair in human cells. Int. J. Radiat. Biol. 1992, 62, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R.; Duthie, S.J.; Dobson, V.L. Direct enzymic detection of endogenous oxidative base damage in human lymphocyte DNA. Carcinogenesis 1993, 14, 1733–1735. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. The use of bacterial repair endonucleases in the comet assay. Methods Mol. Biol. 2011, 691, 137–147. [Google Scholar] [CrossRef]
- Speit, G.; Haupter, S.; Schütz, P.; Kreis, P. Comparative evaluation of the genotoxic properties of potassium bromate and potassium superoxide in V79 Chinese hamster cells. Mutat. Res. 1999, 439, 213–221. [Google Scholar] [CrossRef]
- Smith, C.C.; O’Donovan, M.R.; Martin, E.A. hOGG1 recognizes oxidative damage using the comet assay with greater specificity than FPG or ENDOIII. Mutagenesis 2006, 21, 185–190. [Google Scholar] [CrossRef]
- McKelvey-Martin, V.J.; Green, M.H.; Schmezer, P.; Pool-Zobel, B.L.; De Méo, M.P.; Collins, A. The single cell gel electrophoresis assay (comet assay): A European review. Mutat. Res. 1993, 288, 47–63. [Google Scholar] [CrossRef]
- Binková, B.; Lewtas, J.; Mísková, I.; Rössner, P.; Cerná, M.; Mrácková, G.; Peterková, K.; Mumford, J.; Meyer, S.; Srám, R. Biomarker studies in northern Bohemia. Environ. Health. Perspect. 1996, 104 (Suppl. S3), 591–597. [Google Scholar] [CrossRef]
- Møller, P.; Knudsen, L.E.; Frentz, G.; Dybdahl, M.; Wallin, H.; Nexø, B.A. Seasonal variation of DNA damage and repair in patients with non-melanoma skin cancer and referents with and without psoriasis. Mutat. Res. 1998, 407, 25–34. [Google Scholar] [CrossRef]
- Hartmann, A.; Plappert, U.; Raddata, K.; Grünert-Fuchs, M.; Speit, G. Does physical activity induce DNA damage? Mutagenesis 1994, 9, 269–272. [Google Scholar] [CrossRef]
- Duthie, S.J.; Dobson, V.L. Dietary flavonoids protect human colonocyte DNA from oxidative attack in vitro. Eur. J. Nutr. 1999, 38, 28–34. [Google Scholar] [CrossRef]
- Pool-Zobel, B.L.; Bub, A.; Müller, H.; Wollowski, I.; Rechkemmer, G. Consumption of vegetables reduces genetic damage in humans: First results of a human intervention trial with carotenoid-rich foods. Carcinogenesis 1997, 18, 1847–1850. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Wakuri, S.; Sakamoto, K.; Tanaka, N. The photogenotoxicity of titanium dioxide particles. Mutat. Res. 1997, 394, 125–132. [Google Scholar] [CrossRef]
- Sasaki, Y.F.; Sekihashi, K.; Izumiyama, F.; Nishidate, E.; Saga, A.; Ishida, K.; Tsuda, S. The comet assay with multiple mouse organs: Comparison of comet assay results and carcinogenicity with 208 chemicals selected from the IARC monographs and U.S. NTP Carcinogenicity Database. Crit. Rev. Toxicol. 2000, 30, 629–799. [Google Scholar] [CrossRef]
- Albertini, R.J.; Anderson, D.; Douglas, G.R.; Hagmar, L.; Hemminki, K.; Merlo, F.; Natarajan, A.T.; Norppa, H.; Shuker, D.E.; Tice, R.; et al. IPCS guidelines for the monitoring of genotoxic effects of carcinogens in humans. International Programme on Chemical Safety. Mutat. Res. 2000, 463, 111–172. [Google Scholar] [CrossRef]
- Hartmann, A.; Agurell, E.; Beevers, C.; Brendler-Schwaab, S.; Burlinson, B.; Clay, P.; Collins, A.; Smith, A.; Speit, G.; Thybaud, V.; et al. Recommendations for conducting the in vivo alkaline Comet assay. 4th International Comet Assay Workshop. Mutagenesis 2003, 18, 45–51. [Google Scholar] [CrossRef]
- Burlinson, P.; Studholme, D.; Cambray-Young, J.; Heavens, D.; Rathjen, J.; Hodgkin, J.; Preston, G.M. Pseudomonas fluorescens NZI7 repels grazing by C. elegans, a natural predator. ISME J. 2013, 7, 1126–1138. [Google Scholar] [CrossRef]
- Collins, A.R.; Dobson, V.L.; Dusinská, M.; Kennedy, G.; Stĕtina, R. The comet assay: What can it really tell us? Mutat. Res. 1997, 375, 183–193. [Google Scholar] [CrossRef]
- Gedik, C.M.; Collins, A. Establishing the background level of base oxidation in human lymphocyte DNA: Results of an interlaboratory validation study. FASEB J. 2005, 19, 82–84. [Google Scholar] [CrossRef]
- Møller, P.; Friis, G.; Christensen, P.H.; Risom, L.; Plesner, G.; Kjaersgaard, J.; Vinzents, P.; Loft, S.; Jensen, A.; Tved, M. Intra-laboratory comet assay sample scoring exercise for determination of formamidopyrimidine DNA glycosylase sites in human mononuclear blood cell DNA. Free Radic. Res. 2004, 38, 1207–1214. [Google Scholar] [CrossRef]
- Møller, P.; Möller, L.; Godschalk, R.W.; Jones, G.D. Assessment and reduction of comet assay variation in relation to DNA damage: Studies from the European Comet Assay Validation Group. Mutagenesis 2010, 25, 109–111. [Google Scholar] [CrossRef]
- Neri, M.; Milazzo, D.; Ugolini, D.; Milic, M.; Campolongo, A.; Pasqualetti, P.; Bonassi, S. Worldwide interest in the comet assay: A bibliometric study. Mutagenesis 2015, 30, 155–163. [Google Scholar] [CrossRef]
- Koppen, G.; Azqueta, A.; Pourrut, B.; Brunborg, G.; Collins, A.R.; Langie, S.A.S. The next three decades of the comet assay: A report of the 11th International Comet Assay Workshop. Mutagenesis 2017, 32, 397–408. [Google Scholar] [CrossRef]
- Gajski, G.; Žegura, B.; Ladeira, C.; Novak, M.; Sramkova, M.; Pourrut, B.; Del Bo, C.; Milić, M.; Gutzkow, K.B.; Costa, S.; et al. The comet assay in animal models: From bugs to whales—(Part 2 Vertebrates). Mutat. Res. Rev. Mutat. Res. 2019, 781, 130–164. [Google Scholar] [CrossRef]
- Jha, A.N. Genotoxicological studies in aquatic organisms: An overview. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 552, 1–17. [Google Scholar] [CrossRef]
- Kumaravel, T.S.; Vilhar, B.; Faux, S.P.; Jha, A.N. Comet Assay measurements: A perspective. Cell Biol. Toxicol. 2009, 25, 53–64. [Google Scholar] [CrossRef]
- Dixon, D.R.; Pruski, A.M.; Dixon, L.R.; Jha, A.N. Marine invertebrate eco-genotoxicology: A methodological overview. Mutagenesis 2002, 17, 495–507. [Google Scholar] [CrossRef]
- Lorenzo, Y.; Costa, S.; Collins, A.R.; Azqueta, A. The comet assay, DNA damage, DNA repair and cytotoxicity: Hedgehogs are not always dead. Mutagenesis 2013, 28, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Committee on Mutagenicity of Chemicals in Food, Consumer Products and the Environment (COM). Guidance on a Strategy for Testing of Chemicals for Mutagenicity; Department of Health: London, UK, 2000.
- Møller, P. The alkaline comet assay: Towards validation in biomonitoring of DNA damaging exposures. Basic. Clin. Pharmacol. Toxicol. 2006, 98, 336–345. [Google Scholar] [CrossRef]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications, and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Bajpayee, M.; Parmar, D. Comet assay: A reliable tool for the assessment of DNA damage in different models. Cell. Biol. Toxicol. 2009, 25, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Møller, P. The comet assay: Ready for 30 more years. Mutagenesis 2018, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Afanasieva, K.; Zazhytska, M.; Sivolob, A. Kinetics of comet formation in single-cell gel electrophoresis: Loops and fragments. Electrophoresis 2010, 31, 512–519. [Google Scholar] [CrossRef]
- Glei, M.; Schneider, T.; Schlörmann, W. Comet assay: An essential tool in toxicological research. Arch. Toxicol. 2016, 90, 2315–2336. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. The Use of Bacterial Repair Endonucleases in the Comet Assay. In Drug Safety Evaluation: Methods and Protocols; Gautier, J.-C., Ed.; Springer: New York, NY, USA, 2017; pp. 173–184. [Google Scholar]
- Møller, P.; Jantzen, K.; Løhr, M.; Andersen, M.H.; Jensen, D.M.; Roursgaard, M.; Danielsen, P.H.; Jensen, A.; Loft, S. Searching for assay controls for the Fpg- and hOGG1-modified comet assay. Mutagenesis 2018, 33, 9–19. [Google Scholar] [CrossRef]
- Townsend, T.A.; Parrish, M.C.; Engelward, B.P.; Manjanatha, M.G. The development and validation of EpiComet-Chip, a modified high-throughput comet assay for the assessment of DNA methylation status. Environ. Mol. Mutagen. 2017, 58, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Sykora, P.; Witt, K.L.; Revanna, P.; Smith-Roe, S.L.; Dismukes, J.; Lloyd, D.G.; Engelward, B.P.; Sobol, R.W. Next generation high throughput DNA damage detection platform for genotoxic compound screening. Sci. Rep. 2018, 8, 2771. [Google Scholar] [CrossRef]
- Henriksson, S.; Shaposhnikov, S.; Nilsson, M.; Collins, A. Study of gene-specific DNA repair in the comet assay with padlock probes and rolling circle amplification. Toxicol. Lett. 2011, 202, 142–147. [Google Scholar] [CrossRef]
- Rojas, E.; Lorenzo, Y.; Haug, K.; Nicolaissen, B.; Valverde, M. Epithelial cells as alternative human biomatrices for comet assay. Front. Genet. 2014, 5, 386. [Google Scholar] [CrossRef]
- Enciso, J.M.; Gutzkow, K.B.; Brunborg, G.; Olsen, A.K.; López de Cerain, A.; Azqueta, A. Standardisation of the in vitro comet assay: Influence of lysis time and lysis solution composition on the detection of DNA damage induced by X-rays. Mutagenesis 2018, 33, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Gajski, G.; Žegura, B.; Ladeira, C.; Pourrut, B.; Del Bo, C.; Novak, M.; Sramkova, M.; Milić, M.; Gutzkow, K.B.; Costa, S.; et al. The comet assay in animal models: From bugs to whales—(Part 1 Invertebrates). Mutat. Res. Rev. Mutat. Res. 2019, 779, 82–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Park, K.; Hong, J.; Choi, J. Ecotoxicological evaluation of octachlorostyrene in fourth instar larvae of Chironomus riparius (Diptera, Chironomidae). Environ. Toxicol. Chem. 2008, 27, 1118–1127. [Google Scholar] [CrossRef]
- Deutschmann, B.; Kolarevic, S.; Brack, W.; Kaisarevic, S.; Kostic, J.; Kracun-Kolarevic, M.; Liska, I.; Paunovic, M.; Seiler, T.B.; Shao, Y.; et al. Longitudinal profile of the genotoxic potential of the River Danube on erythrocytes of wild common bleak (Alburnus alburnus) assessed using the comet and micronucleus assay. Sci. Total. Environ. 2016, 573, 1441–1449. [Google Scholar] [CrossRef]
- Naz, S.; Hussain, R.; Ullah, Q.; Chatha, A.M.M.; Shaheen, A.; Khan, R.U. Toxic effect of some heavy metals on hematology and histopathology of major carp (Catla catla). Environ. Sci. Pollut. Res. 2021, 28, 6533–6539. [Google Scholar] [CrossRef]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Bhagat, J. Combinations of genotoxic tests for the evaluation of group 1 IARC carcinogens. J. Appl. Toxicol. 2018, 38, 81–99. [Google Scholar] [CrossRef]
- Verschaeve, L.; Gilles, J. Single cell gel electrophoresis assay in the earthworm for the detection of genotoxic compounds in soils. Bull. Environ. Contam. Toxicol. 1995, 54, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Langie, S.; Azqueta, A.; Collins, A. The comet assay: Past, present, and future. Front. Genet. 2015, 6, 266. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.M.K.; Shyama, S.K.; Sonaye, B.S.; Naik, U.R.; Kadam, S.B.; Bipin, P.D.; D’Costa, A.; Chaubey, R.C. Evaluation of γ-radiation-induced DNA damage in two species of bivalves and their relative sensitivity using comet assay. Aquat. Toxicol. 2014, 150, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Wei, L.; Cao, J.; Qiu, L.; Jiang, X.; Li, P.; Song, Q.; Zhou, H.; Han, Q.; Diao, X. Oxidative stress, DNA damage and antioxidant enzyme activities in the pacific white shrimp (Litopenaeus vannamei) when exposed to hypoxia and reoxygenation. Chemosphere 2016, 144, 234–240. [Google Scholar] [CrossRef]
- Ge, W.; Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Oxidative stress and DNA damage induced by imidacloprid in zebrafish (Danio rerio). J. Agric. Food Chem. 2015, 63, 1856–1862. [Google Scholar] [CrossRef]
- Xu, K.; Tang, Z.; Liu, S.; liao, Z.; Xia, H.; Liu, L.; Wang, Z.; Qi, P. Effects of low concentrations copper on antioxidant responses, DNA damage and genotoxicity in thick shell mussel Mytilus coruscus. Fish Shellfish. Immunol. 2018, 82, 77–83. [Google Scholar] [CrossRef]
- Banni, M.; Negri, A.; Dagnino, A.; Jebali, J.; Ameur, S.; Boussetta, H. Acute effects of benzo[a]pyrene on digestive gland enzymatic biomarkers and DNA damage on mussel Mytilus galloprovincialis. Ecotoxicol. Environ. Saf. 2010, 73, 842–848. [Google Scholar] [CrossRef]
- Riva, C.; Binelli, A.; Cogni, D.; Provini, A. Evaluation of DNA damage induced by decabromodiphenyl ether (BDE-209) in hemocytes of Dreissena polymorpha using the comet and micronucleus assays. Environ. Mol. Mutagen. 2007, 48, 735–743. [Google Scholar] [CrossRef]
- Dixon, D.R.; Pruski, A.M.; Dixon, L.R.J. The effects of hydrostatic pressure change on DNA integrity in the hydrothermal-vent mussel Bathymodiolus azoricus: Implications for future deep-sea mutagenicity studies. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2004, 552, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.; Khan, M.N.; Qureshi, N.A.; Chaudhry, A.S. Monitoring DNA damage in gills of freshwater mussels (Anodonta anatina) exposed to heavy metals. Pak. J. Zool. 2017, 49, 305–311. [Google Scholar] [CrossRef]
- Vasanthi, L.A.; Revathi, P.; Babu Rajendran, R.; Munuswamy, N. Detection of metal induced cytopathological alterations and DNA damage in the gills and hepatopancreas of green mussel Perna viridis from Ennore Estuary, Chennai, India. Mar. Pollut. Bull. 2017, 117, 41–49. [Google Scholar] [CrossRef]
- Coughlan, B.M.; Hartl, M.G.J.; O’Reilly, S.J.; Sheehan, D.; Morthersill, C.; van Pelt, F.N.A.M.; O’Halloran, J.; O’Brien, N.M. Detecting genotoxicity using the Comet assay following chronic exposure of Manila clam Tapes semidecussatus to polluted estuarine sediments. Mar. Pollut. Bull. 2002, 44, 1359–1365. [Google Scholar] [CrossRef]
- Abdus Sallam, M.; Zubair, M.; Tehseen Gul, S.; Ullah, Q.; Idrees, M. Evaluating the protective effects of vitamin E and selenium on hematology and liver, lung and uterus histopathology of rabbits with cypermethrin toxicity. Toxin Rev. 2020, 39, 236–241. [Google Scholar] [CrossRef]
- Hartl, M.G.; Coughlan, B.M.; Sheehan, D.; Mothersill, C.; van Pelt, F.N.; O’Reilly, S.J.; Heffron, J.J.; O’Halloran, J.; O’Brien, N.M. Implications of seasonal priming and reproductive activity on the interpretation of Comet assay data derived from the clam, Tapes semidecussatus Reeves 1864, exposed to contaminated sediments. Mar. Environ. Res. 2004, 57, 295–310. [Google Scholar] [CrossRef]
- Gwo, J.C.; Wu, C.Y.; Chang, W.S.; Cheng, H.Y. Evaluation of damage in Pacific oyster (Crassostrea gigas) spermatozoa before and after cryopreservation using comet assay. Cryo letters 2003, 24, 171–180. [Google Scholar]
- Gielazyn, M.L.; Ringwood, A.H.; Piegorsch, W.W.; Stancyk, S.E. Detection of oxidative DNA damage in isolated marine bivalve hemocytes using the comet assay and formamidopyrimidine glycosylase (Fpg). Mutat. Res. 2003, 542, 15–22. [Google Scholar] [CrossRef]
- Large, A.T.; Shaw, J.P.; Peters, L.D.; McIntosh, A.D.; Webster, L.; Mally, A.; Chipman, J.K. Different levels of mussel (Mytilus edulis) DNA strand breaks following chronic field and acute laboratory exposure to polycyclic aromatic hydrocarbons. Mar. Environ. Res. 2002, 54, 493–497. [Google Scholar] [CrossRef]
- Lewis, C.; Galloway, T. Genotoxic damage in polychaetes: A study of species and cell-type sensitivities. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2008, 654, 69–75. [Google Scholar] [CrossRef]
- Palmqvist, A.; Rasmussen, L.J.; Forbes, V.E. Influence of biotransformation on trophic transfer of the PAH, fluoranthene. Aquat. Toxicol. 2006, 80, 309–319. [Google Scholar] [CrossRef]
- Buffet, P.E.; Zalouk-Vergnoux, A.; Châtel, A.; Berthet, B.; Métais, I.; Perrein-Ettajani, H.; Poirier, L.; Luna-Acosta, A.; Thomas-Guyon, H.; Risso-de Faverney, C.; et al. A marine mesocosm study on the environmental fate of silver nanoparticles and toxicity effects on two endobenthic species: The ragworm Hediste diversicolor and the bivalve mollusc Scrobicularia plana. Sci. Total. Environ. 2014, 470–471, 1151–1159. [Google Scholar] [CrossRef]
- Bach, L.; Palmqvist, A.; Rasmussen, L.J.; Forbes, V.E. Differences in PAH tolerance between Capitella species: Underlying biochemical mechanisms. Aquat. Toxicol. 2005, 74, 307–319. [Google Scholar] [CrossRef]
- Essawy, A.E.; Sherif, S.S.E.; Osman, G.Y.; Morshedy, R.M.E.; Al-Nasser, A.S.; Sheir, S.K. Immune responses, DNA damage and ultrastructural alterations of gills in the marine mussel Lithophaga lithophaga exposed to CuO nanoparticles. Environ. Sci. Pollut. Res. 2022, 29, 15800–15815. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sharma, A.; Mishra, M.; Mishra, R.K.; Chowdhuri, D.K. Heat shock proteins in toxicology: How close and how far? Life Sci. 2010, 86, 377–384. [Google Scholar] [CrossRef]
- Pauls, S.U.; Alp, M.; Bálint, M.; Bernabò, P.; Čiampor Jr, F.; Čiamporová-Zaťovičová, Z.; Finn, D.S.; Kohout, J.; Leese, F.; Lencioni, V.; et al. Integrating molecular tools into freshwater ecology: Developments and opportunities. Freshw. Biol. 2014, 59, 1559–1576. [Google Scholar] [CrossRef]
- Martínez-Paz, P.; Morales, M.; Martínez-Guitarte, J.L.; Morcillo, G. Genotoxic effects of environmental endocrine disruptors on the aquatic insect Chironomus riparius evaluated using the comet assay. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2013, 758, 41–47. [Google Scholar] [CrossRef]
- Yao, C.-L.; Somero, G.N. The impact of acute temperature stress on hemocytes of invasive and native mussels (Mytilus galloprovincialis and Mytilus californianus): DNA damage, membrane integrity, apoptosis and signaling pathways. J. Exp. Biol. 2012, 215, 4267–4277. [Google Scholar] [CrossRef]
- Cheng, C.H.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Jie, Y.K.; Guo, Z.X. Oxidative stress, cell cycle arrest, DNA damage and apoptosis in the mud crab (Scylla paramamosain) induced by cadmium exposure. Chemosphere 2021, 263, 128277. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Huang, Y.; Yan, G.; Yin, H.; Huang, Z. DNA damage, immunotoxicity, and neurotoxicity induced by deltamethrin on the freshwater crayfish, Procambarus clarkii. Environ. Toxicol. 2020, 36, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Knapik, L.F.O.; Ramsdorf, W. Ecotoxicity of malathion pesticide and its genotoxic effects over the biomarker comet assay in Daphnia magna. Environ. Monit. Assess. 2020, 192, 264. [Google Scholar] [CrossRef]
- Pérez-Cadahía, B.; Laffon, B.; Pásaro, E.; Méndez, J. Evaluation of PAH bioaccumulation and DNA damage in mussels (Mytilus galloprovincialis) exposed to spilled Prestige crude oil. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 138, 453–460. [Google Scholar] [CrossRef]
- Button, M.; Jenkin, G.R.T.; Bowman, K.J.; Harrington, C.F.; Brewer, T.S.; Jones, G.D.D.; Watts, M.J. DNA damage in earthworms from highly contaminated soils: Assessing resistance to arsenic toxicity by use of the Comet assay. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2010, 696, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Nagarajappa; Ganguly, A.; Goswami, U. DNA damage in male gonad cells of Green mussel (Perna viridis) upon exposure to tobacco products. Ecotoxicology 2006, 15, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.; Carvalho, L.; Sá, N.; Silva, V.; Beraldini, N.; Souza, V.; Conceição, M. Genotoxic effects of the water-soluble fraction of heavy oil in the brackish/freshwater amphipod Quadrivisio aff. lutzi (Gammaridea) as assessed using the comet assay. Ecotoxicology 2013, 22, 642–655. [Google Scholar] [CrossRef] [PubMed]
- El-Bibany, A.H.; Bodnar, A.G.; Reinardy, H.C. Comparative DNA Damage and Repair in Echinoderm Coelomocytes Exposed to Genotoxicants. PLOS ONE 2014, 9, e107815. [Google Scholar] [CrossRef]
- Buschini, A.; Carboni, P.; Martino, A.; Poli, P.; Rossi, C. Effects of temperature on baseline and genotoxicant-induced DNA damage in haemocytes of Dreissena polymorpha. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2003, 537, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Pavlica, M.; Klobucar, G.I.; Mojas, N.; Erben, R.; Papes, D. Detection of DNA damage in haemocytes of zebra mussel using comet assay. Mutat. Res. 2001, 490, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Manerikar, R.S.; Apte, A.A.; Ghole, V.S. In vitro and in vivo genotoxicity assessment of Cr(VI) using comet assay in earthworm coelomocytes. Environ. Toxicol. Pharmacol. 2008, 25, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Choi, J. Genotoxic effects of nonylphenol and bisphenol A exposure in aquatic biomonitoring species: Freshwater crustacean, Daphnia magna, and aquatic midge, Chironomus riparius. Bull. Environ. Contam. Toxicol. 2009, 83, 463–468. [Google Scholar] [CrossRef]
- Bernabò, P.; Gaglio, M.; Bellamoli, F.; Viero, G.; Lencioni, V. DNA damage and translational response during detoxification from copper exposure in a wild population of Chironomus riparius. Chemosphere 2017, 173, 235–244. [Google Scholar] [CrossRef]
- Eskandari, S.; Mozdarani, H.; Mashinchian Moradi, A.; Shahhosseiny, M. Cytogenetic damage induced by crude oil in Anodonta cygnea (mollusca, bivalvia) assessed by the comet assay and micronucleus test. Int. J. Mar. Sci. Eng. 2012, 2, 215–224. [Google Scholar]
- Taban, I.C.; Bechmann, R.K.; Torgrimsen, S.; Baussant, T.; Sanni, S. Detection of DNA damage in mussels and sea urchins exposed to crude oil using comet assay. Mar. Environ. Res. 2004, 58, 701–705. [Google Scholar] [CrossRef]
- Siu, W.H.; Cao, J.; Jack, R.W.; Wu, R.S.; Richardson, B.J.; Xu, L.; Lam, P.K. Application of the comet and micronucleus assays to the detection of B[a]P genotoxicity in haemocytes of the green-lipped mussel (Perna viridis). Aquat. Toxicol. 2004, 66, 381–392. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Ternjej, I.; Stanković, I.; Kerovec, M.; Kopjar, N. Application of the comet assay and detection of DNA damage in haemocytes of medicinal leech affected by aluminium pollution: A case study. Environ. Pollut. 2009, 157, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Banni, M.; Sforzini, S.; Arlt, V.M.; Barranger, A.; Dallas, L.J.; Oliveri, C.; Aminot, Y.; Pacchioni, B.; Millino, C.; Lanfranchi, G.; et al. Assessing the impact of Benzo[a]pyrene on Marine Mussels: Application of a novel targeted low density microarray complementing classical biomarker responses. PLOS ONE 2017, 12, e0178460. [Google Scholar] [CrossRef] [PubMed]
- Barranger, A.; Akcha, F.; Rouxel, J.; Brizard, R.; Maurouard, E.; Pallud, M.; Menard, D.; Tapie, N.; Budzinski, H.; Burgeot, T.; et al. Study of genetic damage in the Japanese oyster induced by an environmentally-relevant exposure to diuron: Evidence of vertical transmission of DNA damage. Aquat. Toxicol. 2014, 146, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Rank, J.; Jensen, K. Comet assay on gill cells and hemocytes from the blue mussel Mytilus edulis. Ecotoxicol. Environ. Saf. 2003, 54, 323–329. [Google Scholar] [CrossRef]
- Mitchelmore, C.L.; Birmelin, C.; Livingstone, D.R.; Chipman, J.K. Detection of DNA strand breaks in isolated mussel (Mytilus edulis L.) digestive gland cells using the “Comet” assay. Ecotoxicol. Environ. Saf. 1998, 41, 51–58. [Google Scholar] [CrossRef]
- Nacci, D.E.; Cayula, S.; Jackim, E. Detection of DNA damage in individual cells from marine organisms using the single cell gel assay. Aquat. Toxicol. 1996, 35, 197–210. [Google Scholar] [CrossRef]
- Belpaeme, K.; Cooreman, K.; Kirsch-Volders, M. Development and validation of the in vivo alkaline comet assay for detecting genomic damage in marine flatfish. Mutat. Res. 1998, 415, 167–184. [Google Scholar] [CrossRef]
- Sumathi, M.; Kalaiselvi, K.; Palanivel, M.; Rajaguru, P. Genotoxicity of textile dye effluent on fish (Cyprinus carpio) measured using the comet assay. Bull. Environ. Contam. Toxicol. 2001, 66, 407–414. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Hyun, C.-K. Comparative evaluation of the alkaline comet assay with the micronucleus test for genotoxicity monitoring using aquatic organisms. Ecotoxicol. Environ. Saf. 2006, 64, 288–297. [Google Scholar] [CrossRef]
- Singh, N.P.; Danner, D.B.; Tice, R.R.; Brant, L.; Schneider, E.L. DNA damage and repair with age in individual human lymphocytes. Mutat. Res. 1990, 237, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Cavas, T. In vivo genotoxicity evaluation of atrazine and atrazine-based herbicide on fish Carassius auratus using the micronucleus test and the comet assay. Food Chem. Toxicol. 2011, 49, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Nagarani, N.; Devi, V.J.; Kumaraguru, A.K. Identification of DNA damage in marine fish Therapon jarbua by comet assay technique. J. Environ. Biol. 2012, 33, 699–703. [Google Scholar] [PubMed]
- Mohanty, G.; Mohanty, J.; Nayak, A.K.; Mohanty, S.; Dutta, S.K. Application of comet assay in the study of DNA damage and recovery in rohu (Labeo rohita) fingerlings after an exposure to phorate, an organophosphate pesticide. Ecotoxicology 2011, 20, 283–292. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, H.; Liu, X. Low levels of cadmium exposure induce DNA damage and oxidative stress in the liver of Oujiang colored common carp Cyprinus carpio var. color. Fish. Physiol. Biochem. 2011, 37, 97–103. [Google Scholar] [CrossRef]
- Arunachalam, K.D.; Annamalai, S.K.; Kuruva, J.K. In-Vivo Evaluation of Hexavalent Chromium Induced DNA Damage by Alkaline Comet Assay and Oxidative Stress in Catla catla. Am. J. Environ. Sci. 2013, 9, 470–482. [Google Scholar] [CrossRef]
- Kousar, S.; Javed, M. Diagnosis of metals induced DNA damage in fish using comet assay. Pak. Vet. J. 2015, 35, 168–172. [Google Scholar]
- Paravani, E.V.; Simoniello, M.F.; Poletta, G.L.; Casco, V.H. Cypermethrin induction of DNA damage and oxidative stress in zebrafish gill cells. Ecotoxicol. Environ. Saf. 2019, 173, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Hong, X.; Yan, S.; Li, X.; Wu, H.; Lin, A.; Yang, W. Neonicotinoids caused oxidative stress and DNA damage in juvenile Chinese rare minnows (Gobiocypris rarus). Ecotoxicol. Environ. Saf. 2020, 197, 110566. [Google Scholar] [CrossRef]
- Rabelo, J.C.S.; Hanusch, A.L.; de Jesus, L.W.O.; Mesquita, L.A.; Franco, F.C.; Silva, R.A.; Sabóia-Morais, S.M.T. DNA damage induced by cylindrospermopsin on different tissues of the biomonitor fish Poecilia reticulata. Environ. Toxicol. 2021, 36, 1125–1134. [Google Scholar] [CrossRef]
- Morin, B.; Filatreau, J.; Vicquelin, L.; Barjhoux, I.; Guinel, S.; Leray-Forget, J.; Cachot, J. Detection of DNA damage in yolk-sac larvae of the Japanese Medaka, Oryzias latipes, by the comet assay. Anal. Bioanal. Chem. 2011, 399, 2235–2242. [Google Scholar] [CrossRef] [PubMed]
- Klobucar, G.I.; Pavlica, M.; Erben, R.; Papes, D. Application of the micronucleus and comet assays to mussel Dreissena polymorpha haemocytes for genotoxicity monitoring of freshwater environments. Aquat. Toxicol. 2003, 64, 15–23. [Google Scholar] [CrossRef]
- Dasgupta, S.; Cao, A.; Mauer, B.; Yan, B.; Uno, S.; McElroy, A. Genotoxicity of oxy-PAHs to Japanese medaka (Oryzias latipes) embryos assessed using the comet assay. Environ. Sci. Pollut. Res. Int. 2014, 21, 13867–13876. [Google Scholar] [CrossRef]
- Kosmehl, T.; Krebs, F.; Manz, W.; Braunbeck, T.; Hollert, H. Differentiation between bioavailable and total hazard potential of sediment-induced DNA fragmentation as measured by the comet assay with Zebrafish embryos. J. Soils Sediments 2007, 7, 377–387. [Google Scholar] [CrossRef]
- Woo, S.; Kim, S.; Yum, S.; Yim, U.H.; Lee, T.K. Comet assay for the detection of genotoxicity in blood cells of flounder (Paralichthys olivaceus) exposed to sediments and polycyclic aromatic hydrocarbons. Mar. Pollut. Bull. 2006, 52, 1768–1775. [Google Scholar] [CrossRef]
- Ku-Centurión, M.; González-Marín, B.; Calderón-Ezquerro, M.C.; Martínez-Valenzuela, M.C.; Maldonado, E.; Calderón-Segura, M.E. DNA Damage Assessment in Zebrafish Embryos Exposed to Monceren(®) 250 SC Fungicide Using the Alkaline Comet Assay. Zebrafish 2016, 13, 442–448. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.; Raimundo, C.C.M.; Dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Poletta, G.L.; Larriera, A.; Kleinsorge, E.; Mudry, M.D. Genotoxicity of the herbicide formulation Roundup (glyphosate) in broad-snouted caiman (Caiman latirostris) evidenced by the Comet assay and the Micronucleus test. Mutat. Res. 2009, 672, 95–102. [Google Scholar] [CrossRef]
- de Andrade, V.M.; de Freitas, T.R.; da Silva, J. Comet assay using mullet (Mugil sp.) and sea catfish (Netuma sp.) erythrocytes for the detection of genotoxic pollutants in aquatic environment. Mutat. Res. 2004, 560, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Simoniello, M.F.; Gigena, F.; Poletta, G.; Loteste, A.; Kleinsorge, E.; Campana, M.; Scagnetti, J.; Parma, M.J. Alkaline comet assay for genotoxic effect detection in neotropical fish Prochilodus lineatus (Pisces, Curimatidae). Bull. Environ. Contam. Toxicol. 2009, 83, 155–158. [Google Scholar] [CrossRef]
- Kumar, R.; Nagpure, N.S.; Kushwaha, B.; Srivastava, S.K.; Lakra, W.S. Investigation of the genotoxicity of malathion to freshwater teleost fish Channa punctatus (Bloch) using the micronucleus test and comet assay. Arch. Environ. Contam. Toxicol. 2010, 58, 123–130. [Google Scholar] [CrossRef]
- Ateeq, B.; Abul Farah, M.; Ahmad, W. Detection of DNA damage by alkaline single cell gel electrophoresis in 2,4-dichlorophenoxyacetic-acid- and butachlor-exposed erythrocytes of Clarias batrachus. Ecotoxicol. Environ. Saf. 2005, 62, 348–354. [Google Scholar] [CrossRef]
- Mouchet, F.; Gauthier, L.; Mailhes, C.; Ferrier, V.; Devaux, A. Comparative evaluation of genotoxicity of captan in amphibian larvae (Xenopus laevis and Pleurodeles waltl) using the comet assay and the micronucleus test. Environ. Toxicol. 2006, 21, 264–277. [Google Scholar] [CrossRef]
- Mouchet, F.; Gauthier, L.; Baudrimont, M.; Gonzalez, P.; Mailhes, C.; Ferrier, V.; Devaux, A. Comparative evaluation of the toxicity and genotoxicity of cadmium in amphibian larvae (Xenopus laevis and Pleurodeles waltl) using the comet assay and the micronucleus test. Environ. Toxicol. 2007, 22, 422–435. [Google Scholar] [CrossRef]
- Del Barga, I.; Frenzilli, G.; Scarcelli, V.; Nigro, M.; Malmvärn, A.; Asplund, L.; Förlin, L.; Sturve, J. Effects of algal extracts (Polysiphonia fucoides) on rainbow trout (Oncorhynchus mykiss): A biomarker approach. Mar. Environ. Res. 2006, 62, S283–S286. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Nagpure, N.S.; Kumar, R.; Sharma, S.; Srivastava, S.K.; Verma, M.S. Genotoxicity evaluation of acute doses of endosulfan to freshwater teleost Channa punctatus (Bloch) by alkaline single-cell gel electrophoresis. Ecotoxicol. Environ. Saf. 2006, 65, 56–61. [Google Scholar] [CrossRef]
- Mamaca, E.; Bechmann, R.K.; Torgrimsen, S.; Aas, E.; Bjørnstad, A.; Baussant, T.; Floch, S.L. The neutral red lysosomal retention assay and Comet assay on haemolymph cells from mussels (Mytilus edulis) and fish (Symphodus melops) exposed to styrene. Aquat. Toxicol. 2005, 75, 191–201. [Google Scholar] [CrossRef]
- Scalon, M.C.; Rechenmacher, C.; Siebel, A.M.; Kayser, M.L.; Rodrigues, M.T.; Maluf, S.W.; Rodrigues, M.A.; Silva, L.B. Evaluation of Sinos River water genotoxicity using the comet assay in fish. Braz. J. Biol. 2010, 70, 1217–1222. [Google Scholar] [CrossRef]
- Sahlmann, A.; Wolf, R.; Holth, T.F.; Titelman, J.; Hylland, K. Baseline and oxidative DNA damage in marine invertebrates. J. Toxicol. Environ. Health A 2017, 80, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, M.; Orzechowska, H.; Kędziorski, A.; Sawczyn, T.; Doleżych, B. DNA damage in grasshoppers’ larvae—Comet assay in environmental approach. Chemosphere 2014, 96, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Kolarević, S.; Knežević-Vukčević, J.; Paunović, M.; Kračun, M.; Vasiljević, B.; Tomović, J.; Vuković-Gačić, B.; Gačić, Z. Monitoring of DNA damage in haemocytes of freshwater mussel Sinanodonta woodiana sampled from the Velika Morava River in Serbia with the comet assay. Chemosphere 2013, 93, 243–251. [Google Scholar] [CrossRef]
- Kolarević, S.; Gačić, Z.; Kostić, J.; Sunjog, K.; Kračun-Kolarević, M.; Paunović, M.; Knežević-Vukčević, J.; Vuković-Gačić, B. Impact of Common Cytostatics on DNA Damage in Freshwater Mussels Unio pictorum and Unio tumidus. CLEAN-Soil, Air, Water 2016, 44, 1471–1476. [Google Scholar] [CrossRef]
- Buschini, A.; Martino, A.; Gustavino, B.; Monfrinotti, M.; Poli, P.; Rossi, C.; Santoro, M.; Dörr, A.J.; Rizzoni, M. Comet assay and micronucleus test in circulating erythrocytes of Cyprinus carpio specimens exposed in situ to lake waters treated with disinfectants for potabilization. Mutat. Res. 2004, 557, 119–129. [Google Scholar] [CrossRef]
- Marčić, M.M.; DoleNec, Z.; Mrakovčić, M. Assessment of DNA integrity in erythrocytes of Cobitis elongata affected by water pollution: The alkaline comet assay study. Folia Zool. 2008, 57, 120–130. [Google Scholar]
- Bonisoli-Alquati, A.; Voris, A.; Mousseau, T.A.; Møller, A.P.; Saino, N.; Wyatt, M.D. DNA damage in barn swallows (Hirundo rustica) from the Chernobyl region detected by use of the comet assay. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2010, 151, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Guilherme, S.I.; Barroso, C.M.; Verschaeve, L.; Pacheco, M.G.; Mendo, S.A. Evaluation of DNA damage induced by environmental exposure to mercury in Liza aurata using the comet assay. Arch. Environ. Contam. Toxicol. 2010, 58, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Rajaguru, P.; Suba, S.; Palanivel, M.; Kalaiselvi, K. Genotoxicity of a polluted river system measured using the alkaline comet assay on fish and earthworm tissues. Environ. Mol. Mutagen. 2003, 41, 85–91. [Google Scholar] [CrossRef]
- Turan, F.; Ergenler, A. Assessment of DNA Damage by Comet Assay in Trachinotus ovatus Cells from Mersin Bay in the Northeastern Mediterranean. Nat. Eng. Sci. 2019, 4, 25–31. [Google Scholar]
- Erismis, U.C.; Ciğerci İ, H.; Konuk, M. Evaluation of DNA damage in Eurasian marsh frogs (Pelophylax ridibundus) by comet assay for determination of possible pollution in the different lakes in central Anatolia, Turkey. Bull. Environ. Contam. Toxicol. 2013, 90, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, Z.; Ternjej, I.; Stanković, I.; Ivković, M.; Zelježić, D.; Mladinić, M.; Kopjar, N. Assessment of genotoxic potency of sulfate-rich surface waters on medicinal leech and human leukocytes using different versions of the Comet assay. Ecotoxicol. Environ. Saf. 2011, 74, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Rank, J.; Lehtonen, K.K.; Strand, J.; Laursen, M. DNA damage, acetylcholinesterase activity and lysosomal stability in native and transplanted mussels (Mytilus edulis) in areas close to coastal chemical dumping sites in Denmark. Aquat. Toxicol. 2007, 84, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Laffon, B.; Rábade, T.; Pásaro, E.; Méndez, J. Monitoring of the impact of Prestige oil spill on Mytilus galloprovincialis from Galician coast. Environ. Int. 2006, 32, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Frenzilli, G.; Bocchetti, R.; Annarumma, F.; Scarcelli, V.; Fattorini, D.; Nigro, M. Time-course variations of oxyradical metabolism, DNA integrity and lysosomal stability in mussels, Mytilus galloprovincialis, during a field translocation experiment. Aquat. Toxicol. 2004, 68, 167–178. [Google Scholar] [CrossRef]
- Nigro, M.; Falleni, A.; Barga, I.D.; Scarcelli, V.; Lucchesi, P.; Regoli, F.; Frenzilli, G. Cellular biomarkers for monitoring estuarine environments: Transplanted versus native mussels. Aquat. Toxicol. 2006, 77, 339–347. [Google Scholar] [CrossRef]
- Halldórsson, H.P.; Ericson, G.; Svavarsson, J. DNA strand breakage in mussels (Mytilus edulis L.) deployed in intertidal and subtidal zone in Reykjavík harbour. Mar. Environ. Res. 2004, 58, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Rank, J.; Jensen, K.; Jespersen, P.H. Monitoring DNA damage in indigenous blue mussels (Mytilus edulis) sampled from coastal sites in Denmark. Mutat. Res. 2005, 585, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Frenzilli, G.; Scarcelli, V.; Barga, I.D.; Nigro, M.; Förlin, L.; Bolognesi, C.; Sturve, J. DNA damage in eelpout (Zoarces viviparus) from Göteborg harbour. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 552, 187–195. [Google Scholar] [CrossRef]
- Russo, C.; Rocco, L.; Morescalchi, M.A.; Stingo, V. Assessment of environmental stress by the micronucleus test and the Comet assay on the genome of teleost populations from two natural environments. Ecotoxicol Environ Saf. 2004, 57, 168–174. [Google Scholar] [CrossRef]
- Winter, M.J.; Day, N.; Hayes, R.A.; Taylor, E.W.; Butler, P.J.; Chipman, J.K. DNA strand breaks and adducts determined in feral and caged chub (Leuciscus cephalus) exposed to rivers exhibiting variable water quality around Birmingham, UK. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 552, 163–175. [Google Scholar] [CrossRef]
- Wirzinger, G.; Weltje, L.; Gercken, J.; Sordyl, H. Genotoxic damage in field-collected three-spined sticklebacks (Gasterosteus aculeatus L.): A suitable biomonitoring tool? Mutat. Res. 2007, 628, 19–30. [Google Scholar] [CrossRef]
- Huang, J.; Li, Q.; Sun, D.; Lu, Y.; Su, Y.; Yang, X.; Wang, H.; Wang, Y.; Shao, W.; He, N.; et al. Biosynthesis of silver and gold nanoparticles by novel sundried Cinnamomum camphora leaf. Nanotechnology 2007, 18, 105104. [Google Scholar] [CrossRef]
- Purschke, M.; Jacobi, H.; Witte, I. Differences in genotoxicity of H(2)O(2) and tetrachlorohydroquinone in human fibroblasts. Mutat. Res. 2002, 513, 159–167. [Google Scholar] [CrossRef]
- Kennelly, J.C.; Lane, M.P.; Barker, J.A.; Barber, G.; Tinwell, H.; Gallagher, J.E.; Pool-Zobel, B.; Schmezer, P.; Ashby, J. Genotoxic activity of 1-chloromethylpyrene in stomach epithelium in vivo: Insensitivity of the stomach scintillation UDS assay. Carcinogenesis 1993, 14, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Collins, A.R. The essential comet assay: A comprehensive guide to measuring DNA damage and repair. Arch. Toxicol. 2013, 87, 949–968. [Google Scholar] [CrossRef] [PubMed]
- Ersson, C.; Möller, L. The effects on DNA migration of altering parameters in the comet assay protocol such as agarose density, electrophoresis conditions and durations of the enzyme or the alkaline treatments. Mutagenesis 2011, 26, 689–695. [Google Scholar] [CrossRef]
- Danson, S.; Ranson, M.; Denneny, O.; Cummings, J.; Ward, T.H. Validation of the comet-X assay as a pharmacodynamic assay for measuring DNA cross-linking produced by the novel anticancer agent RH1 during a phase I clinical trial. Cancer Chemother. Pharmacol. 2007, 60, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Ngo, L.P.; Owiti, N.A.; Swartz, C.; Winters, J.; Su, Y.; Ge, J.; Xiong, A.; Han, J.; Recio, L.; Samson, L.D.; et al. Sensitive CometChip assay for screening potentially carcinogenic DNA adducts by trapping DNA repair intermediates. Nucleic Acids Res. 2020, 48, e13. [Google Scholar] [CrossRef] [PubMed]
- Al-Salmani, K.; Abbas, H.H.; Schulpen, S.; Karbaschi, M.; Abdalla, I.; Bowman, K.J.; So, K.K.; Evans, M.D.; Jones, G.D.; Godschalk, R.W.; et al. Simplified method for the collection, storage, and comet assay analysis of DNA damage in whole blood. Free. Radic. Biol. Med. 2011, 51, 719–725. [Google Scholar] [CrossRef]
- Cortés-Gutiérrez, E.I.; Dávila-Rodríguez, M.I.; Fernández, J.L.; López-Fernández, C.; Gosálbez, A.; Gosálvez, J. New application of the comet assay: Chromosome-comet assay. J. Histochem. Cytochem. 2011, 59, 655–660. [Google Scholar] [CrossRef]


| Sr. No. | Procedure | Advantages | Limitations |
|---|---|---|---|
| 1. | TUNEL | Identifies single- and double-strand breaks in DNA by fluorescing its free ends Rapid, easy to perform, highly sensitive | False positives from cells involved in DNA repair or gene transcription as well as necrotic cells Difficult to differentiate apoptotic from necrotic cells |
| 2. | Polymerase chain reaction (PCR) | Gene-specific DNA damage is easily quantified Shown to be more cost-effective with selective use than culture and staining | Cannot quantify and recognize the kind of damage Its specificity is potentially low |
| 3. | Immunological assay | Requires less amount of DNA, rapid, yields improved precision | Needs costly equipment |
| 4. | Micronucleus assay | Easy to carry out the procedure, less time consuming Reliable assay to assess the induction of chromosomal aberrations | Low sensitivity, do take direct observation of DNA False positive and false negative results |
| 5. | Halo assay | Identify change in DNA organization in individual cell | Low sensitivity |
| 6. | High-performance liquid chromatography (HPLC) | Sensitive and specific for measuring oxidative DNA damage and thymidine dimer levels | Early elution property of liquid chromatography |
| 7. | Gas chromatography-mass spectroscopy (GC-MS) | Sensitive to point out oxidative DNA damage | Overestimation of damage |
| 8. | Flow cytometry (FCM) | Detects DNA damage exclusively in apoptotic cells | Sensitivity is low compared to FISH |
| 9. | Fluorescence in situ hybridization (FISH) | Non-isotope labeling and detection method Greater sensitivity than FCM | Well-equipped laboratory facility |
| Animal Specie | Type of Sample (Tissue/Cells) | Sampling Location | Descriptor | Agent | Response | Reference |
|---|---|---|---|---|---|---|
| Hyphessobrycon leutkenii | Blood | Sinos river. Sites: Towns of Caraá, Parobé and Sapucaia do Sul. Sampling Seasons included Spring, Summer, Autumn and Winter. | % Tail DNA | Water contaminated with metals, i.e., lead, chromium, copper, nickel, zinc, etc. | Metal accumulation and DNA damage index: no remarkable difference in all three sites: At site 1, spring > fall. At site 2, spring > autumn, winter and summer. | [140] |
| Ciona intestinalis Asteriasrubens Carcinusmaenas Mytilus edulis Gadus morhua | Lymphocytes | Outer Oslofjord in the vicinity of Drøbak, Southern Norway | DNA strand breaks | Hydrogen peroxide (H2O2) (25 or 250 μM) | Asterias rubens (34%) > Carcinusmaenas (25%) > Mytilus edulis (22%) > Ciona intestinalis (17%) > Gadus morhua(7%). | [141] |
| Chorthippus brunneus | Brain cells | Southern Poland. Site 1: Olkusz, Site 2: Szopienice, and Site 3: Pilica | TL, %TDNA, and OTM | Hydrogen peroxide (20 µM) A single 10 min application | DNA damage in adults: Site 2 > Site 1 and Site 3. DNA damage in larvae: Site 1 = Site 3, Site 2 > Site 1 and Site 3 | [142] |
| Sinanodonta woodiana | Hemolymph | Velika Morava River, Serbia. Site 1 and Site 2. Sampling season: Spring, Summer, Autumn and Winter. | OTM | Water contaminated with phosphates and increased concentrations of zinc, copper, and nickel. | OTM in Summer > Winter, Spring and Autumn | [143] |
| Unio pictorum Unio tumidus | Haemocytes | The Sava River the Sava River Sava river, Europe | % Tail DNA | Water contaminated with Etoposide (ETO), Vincristine sulfate (VIN), and cisplatin (CP). Sampling Period: 72 h for in vivo and 22 Hours for in vitro | ETO (40 and 100 μM) > VIN (0.04 and 0.1 µM) > CP (4 μM) post treatment with hydrogen peroxide. | [144] |
| Cyprinus carpio | Erythrocytes | Trasimeno Lake, Italy. Sampling Periods: October 2000, February 2001, and June 2001 | TM | High levels of bromide and total organic carbon (TOC) in surface water | DNA migration of populations samples in June 2001 > October 2000 and February 2001. | [145] |
| Cobitis elongata | Erythrocytes | Sava (Site 1 and Site 2) and Kupa Rivers, Slovenia | TL, %TDNA, and OTM | Polluted water | Sava 1 site > Sava 2 site > Kupa River | [146] |
| Hirundo rustica | Blood cells | Dytiaku, Pysky, Vesniane and Ghovtnere | % Tail DNA | Radionuclides released by the accident at the Fukushima-Daiichi Nuclear Power Plant (FDNPP) Nest samples 479–143,349 Bq/kg. External exposure (0.15 and 4.9 mGy) | The genetic damage in nestlings did not increase with exposure to radioactive pollution. But, with higher levels of radioactive contamination, fewer barn swallows were present, and a smaller proportion of them were juveniles, indicating reduced survival rates, reproduction, or fledging. | [147] |
| Liza aurata | Blood cells | Laranjo basin, Portugal. Reference site and LAR (Contaminated Site) | TD%, TL, and TM | Mercury contaminated site | Patterns of DNA lesions in Liza aurata for both Reference and contaminated fish, the following order was found: Summer and Autumn > Winter > Spring | [148] |
| Fish (Cyprinus carpio) and earthworms (Eisenia foetida) | Erythrocytes, liver, and kidney cells | Noyal River, India | DNA length:width ratio of the DNA mass | Water and sediment collected from the site polluted with industrial effluent and sewage | The coelomocytes of earthworms subjected to sediment samples had a considerably greater mean DNA length:width ratio | [149] |
| Trachinotus ovatus | Liver and gill tissues | Mersin Bay in the North-eastern Mediterranean, Turkey | Tail length, tail intensity, and tail migration | Gamma Irradiation | Both liver and gill tissues have medium damage | [150] |
| Eurasian Marsh Frogs (Pelophylax ridibundus) | Venous blood | Central Anatolia, Turkey. Site 1: drinking water supply, Akdeğirmen Dam Lake in Afyonkarahisar city. Site 2: Sinanpaşa pond, near Sinanpaşa town, used as an artificial reservoir for gathering the town’s domestic waste and sewage water. Site 3: Emre Lake is bordered by intensive agriculture crops, primarily vegetable and fruit crops, and is located close to Doger town. Site 4: Porsuk Dam Lake, which is on the Porsuk River, the primary supply of drinking water for the city of Eskişehir. | TL, %TDNA, and OTM | Water is polluted with heavy metals, i.e., Arsenic, Mercury, Ammonia, etc. | Order of locations for measuring the amount of DNA damage. Site-4 > Site-2 > Site-3 > Site-1. The chemical pollution level in the contaminated water bodies was Site 4 > Site 2 > Site 3 > Site 1. Regarding the level of Phosphate, Site 2 level was slightly higher than Site 4. | [151] |
| Hirudo verbana | Haemocytes | Lake Njivice, Slovenia | TL, %TDNA, and TM | Contaminated water and sediments | The findings showed that excessive aluminum compounds in specimens could damage DNA. Results indicated that both Lake Njivice water and sediment samples showed high potential for DNA damage. | [152] |
| Mytilus edulis | Haemolymph and gill cells | Chemical dumping sites, Denmark | TM | Water contaminated by anthropogenic runoff from chemical waste disposal plants | Chromium, nickel, and cadmium were shown to have a strong positive correlation with DNA damage in the haemolymph and gill cells of Mytilus edulis. The results indicated a response to pollution in all the biomarkers applied at the suspected areas, but the results were inconsistent. | [153] |
| Mytilus galloprovincialis | Gill cells | Galician coast, North West of Spain. Sampling Period: 11 month (August 2003-June 2004, nine samplings) Site 1: Lira. Site 2: Ancoradoiro beaches. | TL | Seawater contamination with prestige oil spill, using Total Polycyclic Aromatic Hydrocarbons (TPAH) as a biomarker. | In the initial samplings, TPAH levels in seawater were quite elevated; afterward, TAPH level declined to 200 ng/L, comparable to reference seawater. The TPAH levels in mussel tissues varied significantly. It started to rise again in January 2004, most likely due to unfavorable weather that disturbed the sea floor and spread oil that had collected in sediments. | [154] |
| Mytilus galloprovincialis | Gill cells | Genova harbor, Italy | %tDNA | Polluted water with Reactive Oxygen Species as a biomarker | The findings for individual antioxidants and TOSC (total oxyradical scavenging capacity) demonstrated a biphasic trend, over the first two weeks of exposure to the polluted site, there was no change or increase, and in the second half of the trial, there was a serious depletion. | [155] |
| Mytilus galloprovincialis | Gill cells | River estuarine, Cecina, Italy | %tDNA | Heavy metal pollution | Increase in MN (micro nucleus) frequency in native > transplanted or reference mussel | [156] |
| Mytilus edulis | Haemocytes, gill cells | Reykjavik harbour(PAH contaminated site) and Hvalfjörethur, Iceland (reference site) | %tDNA | PAH (Polycyclic aromatic hydrocarbon) contamination in water | For both gill cells and haemocytes, DNA damage in mussels deployed in Reykjavík harbour > the reference site. DNA damage from ReykjavÍk harbour in Intertidal mussels > subtidal mussels. | [157] |
| Mytilus edulis | Haemocytes, gill cells | Denmark coast, Northern Europe | %tDNA | Sediments and water polluted with heavy metals | DNA damage in Gill cells > haemolymph cells. DNA damage in Mussels from contaminated sites > unpolluted coastal locations. | [158] |
| Zootoca vivipara | Erythrocytes | Göteborg harbour, Sweden | %tDNA | Polycyclic aromatic hydrocarbon and bunker oil spill | In all the polluted areas, fish erythrocytes’ DNA was found to be damaged three weeks after the oil spill. | [159] |
| Gambusia holbrooki | Erythrocytes | Sarno River, Astroni natural reserve (negative control) Italy | TL | Mutagenic agents in the polluted water of Sarno river | MN (micro nucleus) and DNA movement in Sarno River > Astroni. | [160] |
| Mugil sp. Netuma sp. | Erythrocytes | Two Brazilian estuaries, Brazil | %Damage frequency (DF), Image length (IL), Damage index (DI) | Methyl Methanesulfonate (MMS) 2, 4, and 8 × 10−5 M for 1 for 2, 6, and 24 h at 25 and 37 °C. | Rising temperatures caused an elevation in the level of DNA damage that was already present, as well as an increase in MMS-induced DNA damage, and a clear dose/time response to methyl methanesulfonate therapy, i.e., trend of DNA damage in hours is 24 > 6 > 2 > 1. | [131] |
| Leuciscus cephalus | Liver cells | Three rivers: the rivers Blythe, Cole, and Tame, United Kingdom | %tlDNA | Polycyclic aromatic hydrocarbons (PAHs), Polychlorinated Biphenyls (PCBs), and Organ chlorine pesticides (OCPs) as well as heavy metals. | In polluted site, order of DNA damage: Tame > Cole > Blyth. DNA damage in Feral animals > Cage-held animals. | [161] |
| Gasterosteus Aculeatus | Erythrocytes | Three places in Northern Germany with various quantities of sewage-treatment effluent: Site 1: Ableitergraben, Site 2: KraakerMuhlenbach, Site 3: Haubach. Sampling periods: 1st in April/May 2002, 2nd in June/July 2002. | %tDNA | Water polluted with sewage-treatment effluents. | At the Ableitergraben median % tail DNA showed a trend as: May > July > August. At the Kraaker Muhlenbach: April > June. At the Haubach, sticklebacks had the second-lowest median tail DNA percentage in this research, with 6.33%. | [162] |
| Buffo raddei | Erythrocytes, liver cells | Lanzhou Region, China | %tDNA | Oil/phenol pollution in water | Dose-response relationship strength in Liver cells > Erythrocytes | [163] |
| Dreissena polymorpha | Haemocytes | Northern Croatia Five monitoring sites: 1 site from River Drava, 4 sites were in the river Sava (1) Zagreb, (2) Oborovo, (3) Sisak, and (4) Lukave) | TM and percentage of migrated DNA | Water polluted with genotoxic chemicals | The order of DNA tail moment and percentage of migrated DNA is given as: Oborovo and Sisak > Lukavec > Zagreb | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, N.; Naz, S.; Ma, Y.; Ullah, Q.; Khan, M.Z.; Wang, J.; Lu, X.; Luosang, D.-Z.; Tabassum, S.; Chatha, A.M.M.; et al. An Overview of Comet Assay Application for Detecting DNA Damage in Aquatic Animals. Agriculture 2023, 13, 623. https://doi.org/10.3390/agriculture13030623
Jiang N, Naz S, Ma Y, Ullah Q, Khan MZ, Wang J, Lu X, Luosang D-Z, Tabassum S, Chatha AMM, et al. An Overview of Comet Assay Application for Detecting DNA Damage in Aquatic Animals. Agriculture. 2023; 13(3):623. https://doi.org/10.3390/agriculture13030623
Chicago/Turabian StyleJiang, Nan, Saima Naz, Yulin Ma, Qudrat Ullah, Muhammad Zahoor Khan, Jiaqi Wang, Xuan Lu, Dun-Zhu Luosang, Sadia Tabassum, Ahmad Manan Mustafa Chatha, and et al. 2023. "An Overview of Comet Assay Application for Detecting DNA Damage in Aquatic Animals" Agriculture 13, no. 3: 623. https://doi.org/10.3390/agriculture13030623
APA StyleJiang, N., Naz, S., Ma, Y., Ullah, Q., Khan, M. Z., Wang, J., Lu, X., Luosang, D.-Z., Tabassum, S., Chatha, A. M. M., & Basang, W.-D. (2023). An Overview of Comet Assay Application for Detecting DNA Damage in Aquatic Animals. Agriculture, 13(3), 623. https://doi.org/10.3390/agriculture13030623