1. Introduction
The yield losses due to weeds are tremendous. For instance, according to estimates from 2021, winter wheat production in North America was impacted by weeds by more than 25% on average [
1]. For Germany, maize yield was shown to be reduced by 70% when weeds were not controlled [
2]. Thus, the efficient control of weeds is important for agricultural production to be economically successful. However, the use of herbicides is one of the most discussed issues in agricultural production practice. The toxicity and persistence of chemically synthesised substances pose a serious problem to the environment (for a review, see [
3]). Moreover, the extensive use of herbicides has fostered the evolution of resistant weeds. A classic example is the R-type of goosegrass (
Eleusine indica), which has acquired resistance to dinitroaniline herbicides due to a mutation in α-tubulin, affecting the binding site for these herbicides [
4]. The ecological concerns along with the progressive loss of efficacy have eroded the lead structures available for the development of new herbicides, stimulating the search for alternative strategies, since mechanical weed control is energy- and labour intensive and, therefore, does not represent an attractive alternative. Total herbicides are usually confined to agricultural systems using GMO crops expressing a resistance marker, which is no option for organic agriculture.
In this context, natural products that can suppress weed growth have attracted considerable attention (for a review, see [
5]). Many plant species use chemical interactions to ward off competitors, a phenomenon known as allelopathy. Such allelopathic activities might be useful for weed control (reviewed in [
6,
7]). This is possible by either growing plants in intercropping systems, or by identifying and isolating the compounds responsible for the allelopathic effect (for a review, see [
8]). For instance, the bioherbicide Avenger
®, approved by the USDA, is based on limonene, extracted from
Citrus peels and, thus, exploits the allelopathic inhibition of
Citrus trees upon their neighbours. Likewise, pelargonic acid, a saturated C
9 fatty acid from
Pelargonium species, is efficiently used against broadleaf weeds [
9]. Although they derive from natural sources, essential oils as bioherbicides must be critically assessed with respect to toxicity in the same way as their synthetic counterparts (for reviews, see [
10,
11]). Following the principle of Paracelsus, the dosage is decisive to delineate undesired collateral damage on other life forms from the desired targeted inhibition of the weed. If the effect of the bioherbicide merely relied on more or less unspecific phytotoxicity, the line towards synthetic herbicides would be relatively arbitrary and just be based upon the mode of production (extraction from natural sources versus chemical synthesis by humans). However, there are indications that some of these bioherbicides do not exert a broadband toxicity, but rather act as specific signals. For instance, there is accumulating evidence that allelopathic interactions often evoke programmed cell death in the target species (recently reviewed in [
12]). Signals differ from toxins with respect to their specificity, which is also reflected in far lower effective concentrations. Both specificity and low concentrations actually come from the target species itself—programmed cell death (PCD) is a central and necessary element of both normal plant development and resilience to stress (for a classical review, see [
13]). Allelopathic signals hijack the underlying signalling, shifting PCD out of its physiological context and thus seriously damaging the target plant.
Mints (Lamiaceae) are interesting in this regard, since they are rich in essential oils (i.e., oils that can be extracted by water steam distillation), mainly volatile monoterpenes that are synthetised in glandular trichomes on the leaves and are widely used for medicinal and cosmetic purposes, but also as spices (for a recent review, see [
14]). In addition to the genus
Mentha, the neighbouring taxa, such as
Nepeta cataria (cat mint) or
Agastache rugosa (Korean mint), are also interesting for their highly active compounds. Since it is progressively clear that
Mentha is not a monophyletic group and the taxonomic relationship with the neighbouring genera is currently under debate, in the current study, we will use the term “mint” in a pragmatic sense to describe the tribe of Menthaea. In their natural environment, mints are often very competitive, which is already indicative of their allelopathic power. In fact, essential oils from horsemint (
M. longifolia) inhibited the germination and growth of wheat [
15], as well as of chickpea [
16], while aqueous extracts of peppermint inhibited germination and photosynthetic pigments in sunflower seedlings [
17]. Likewise, the herbicidal action of mint essential oil has been shown for the pertinent weeds
Rumex and
Convolvulus upon treatment with oil from
M. longifolia [
18].
While the allelopathic activity as such has been well demonstrated, the cellular mode of action of mint essential oils has been addressed only rarely. In a previous study, we showed that the allelopathic activity of essential oil from Korean mint (
Agastache rugosa) correlated with the abundance of menthone and isomenthone in this species [
19] and targeted microtubules as central drivers of cell growth. The effect was dose-dependent and specific, since menthol, which differs from menthone only by a hydroxyl group instead of the carbonyl, required around ten times higher concentrations to exert the same effect in tobacco BY-2 cells. The rapid disassembly of microtubules was followed by cell death that was accentuated when microtubules were stabilised by the overexpression of the green fluorescent protein with tobacco tubulin α3. The microtubule effect of menthone was confirmed for seedlings of
Arabidopsis thaliana as well [
19].
Even though this study demonstrates that mint essential oil evokes a specific response that has the potential to be employed for a bioherbicidal effect, we did not explore this potential, nor did we probe for differential sensitivity of different target species as a prerequisite for use as a bioherbicide. Therefore, we address these issues in the current study, comparing essential oils from three different mints and the lead compounds found in oils extracted from them with respect to germination inhibition in cress. The study was designed in an iterative manner, testing different mints (genus
Mentha, in some experiments also cat mint,
Nepeta cataria, and Korean mint,
Agastache rugosa) that in preparatory research have been found to be endowed with allelopathic activity. Starting with a screening of oils and compounds, we funnelled down to the mode of action of
M. longifolia (since the mode of action for
A. rugosa has already been addressed) [
19]. We confirm a dose-dependent inhibition of germination in a panel of different species, including the pertinent weed,
Rumex acerifolia. We demonstrate further that the efficacy of the essential oils and the individual compounds therein depends on the target species, and that each target species has a specific profile of inhibitory oils and compounds. To obtain insight into the cellular mode of action, we test the effect of essential oil from
M. longifolia in fluorescently labelled cytoskeletal marker lines in tobacco BY-2 and find that this oil specifically disrupts actin microfilaments, while microtubules are not sensitive. Since actin is crucial for auxin transport, which in turn is pivotal for the regeneration of vegetatively propagating weeds, we test the use of
M. longifolia essential oil on bindweed (
Convolvulus arvensis), a pertinent problem in organically grown cereals. Using a slow-release carrier and two different modes of application, we demonstrate that horsemint oil can be used as bioherbicide to contain bindweed. Our study connects phenomenologically assessed sensitivity patterns with a specific cellular mode of action, and a hypothesis-driven application derived from this mode of action, which can be experimentally confirmed. We propose that allelopathic signals can hijack biological signalling in the target species, which allows their valorisation for organic agriculture.
2. Materials and Methods
2.1. Extraction of Essential Oils
The plants, which were used for oil extraction, were raised in the greenhouses of the Botanical Garden of the Karlsruhe Institute of Technology, Karlsruhe, Germany. A few days prior to extraction, the plants were exposed to sunlight to stimulate the formation of essential oils. All source plants had been verified by taxonomic identification, as well as by genetic barcoding (
Supplementary Table S1). In brief, essential oils were extracted from around 30–50 g of fresh leaf material by water steam distillation as described in [
19] and the resulting essential oils were stored at 4 °C till experimental use. The lead compounds of these oils, identified by GC-MS [
19], were used in parallel for comparison. The GC-MS data for the essential oils used in the current study are given in
Supplementary Material S1. These included menthone/isomenthone, linalool, limonene, α-pinene and β-pinene, pulegone, and carvone, all of which were purchased from Sigma-Aldrich (Deisenhofen, Germany). With the exception of menthone, which was of 90% purity (with 10% isomenthone), these compounds were of analytical grade. The solvent, n-hexane, was of hypergrade for organic trace analysis (Sigma-Aldrich, Deisenhofen, Germany).
2.2. Assays for Germination Inhibition
To assess the inhibition pattern of the mint oils, we used a panel of different target species: Arabidopsis thaliana L. ecotype Columbia, Triticum aestivum L. cv. Wiwa (Sativa Rheinau, Switzerland), Phacelia tanacetifolia (UFA Samen, Winterthur, Switzerland), Solanum lycopersicum L. Marmande (Samen Mauser Eric Schweizer AG, Thun, Switzerland), Avena sativa L. cv. Ebners Nackthafer (Botanical Garden of the Karlsruhe Institute of Technology), Rumex obtusifolia L. (FiBL, field collection, Frick), and Convolvulus arvensis L. (FiBL, field collection). To analyse the bioactivity of the essential oils and the different compounds thereof, two different approaches were used, either through the gas phase without the direct contact of the target seeds with the active agent, or, alternatively, after preceding imbibition facilitated by a detergent.
For application through the gas phase, the target species was allowed to germinate in Plexiglass boxes with covers (that were sealed with Parafilm after sowing. The seeds were sown equidistantly on humidified tissue paper. For releasing the volatile substances, a glass slide carrying a filter paper doped with either 1 μL or 10 μL of the oil sample, the respective compound, or the respective solvent was placed in the centre of the box. Each combination was tested three times in technical triplicates. The seeds were incubated in the dark at 25 °C and germination was scored after two (L. sativum, T. aestivum) to five (S. lycopersicum, Tanacetum) days.
For detergent-facilitated imbibition, the probing solution was prepared from 10 mL deionised water with 1% v/v of Tween 20 and the respective test oil or compound (between 0.10 and 0.20% v/v) and vortexed vigorously. The seeds were then sown equidistantly on filter paper in Petri dishes and complemented with 2 mL of the probing solution (revortexed again immediately prior to application). The Petri dishes were sealed using Parafilm and the seeds allowed to germinate for 5 days at 20 °C in the dark. The experiments were repeated in triplicate.
2.3. Assay for Inhibition of Regeneration in Convolvulus
To test their effect on the regeneration of the pertinent weed Convolvulus arvensis, essential oils from M. longifolia, M. spicata crispa, and A. rugosa were mixed into a slow-release alginate carrier (proprietary formulation) in a concentration of 2.5% v/v. Segments of Convolvulus runners (3 cm) were exposed in two configurations in standard field soil sampled from the site of origin (see above). In the ‘in-soil’ configuration, the compound was in direct contact with the doped carrier and covered by 2 cm of soil. In the ‘application from top’ configuration, the compound was placed above the segments and separated by a thin layer of soil, while keeping the same overall depth (2 cm). In total, 2 g of the formulation was used per pot and the experiment was conducted with nine individual pots per configuration and type of essential oil. The entire experiment was repeated twice. The growth and development of the target rhizomes were scored after two weeks at 20 °C and a cycle of 16 h light and 8 h dark. As readouts, the shoot length, number of regenerated leaves, and fresh weight were monitored.
2.4. Live-Cell Imaging of the Cytoskeleton
To assess the cellular effects of the
M. longifolia oil, we used two GFP-tagged tobacco BY-2 lines expressing GFP-tagged markers for the cytoskeleton. Microtubules could be followed by tobacco tubulin α3 with a N-terminal fusion of GFP under the control of a constitutive CaMV 35S promoter [
20], while actin filaments were visualised by the second actin-binding domain of fimbrin (AtFIM1) fused to GFP, also under the control of the Cauliflower Mosaic Virus 35S promoter [
21]. The cytoskeleton was followed in vivo by spinning disc confocal microscopy as described in [
19].
2.5. Statistical Analysis
The data from the germination assays were tested by ANOVA. Post hoc comparisons of differences among means were compared by the Least Significant Difference (LSD) test, applying different numbers for a significance threshold of p < 0.05. For the Convolvulus experiment, significant differences from the control were probed using a two-tailed Student’s t-test for significance thresholds of p < 0.05 and p < 0.01.
3. Results
3.1. Essential Horsemint Oil Has Allelopathic Activity
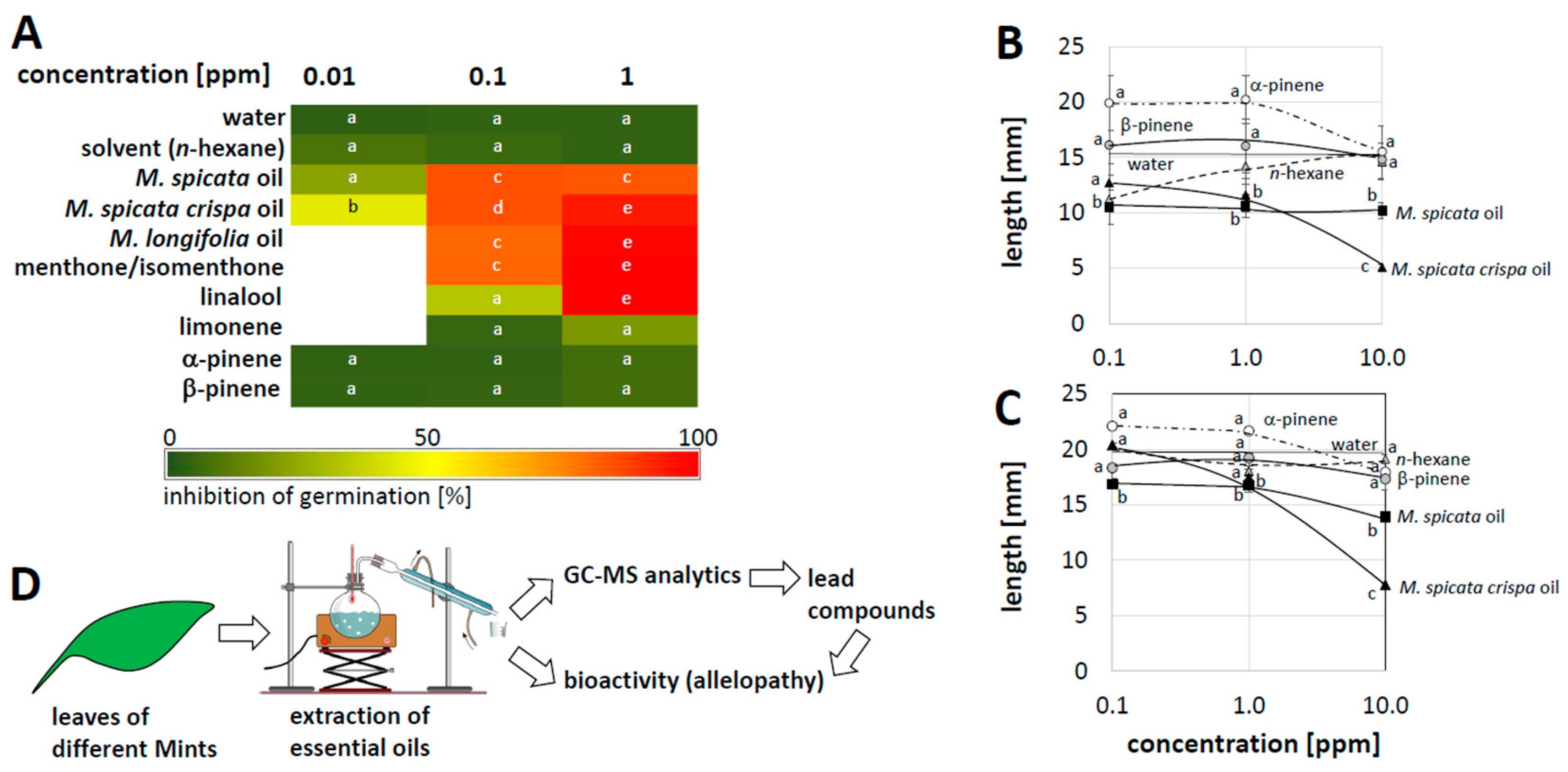
Circumstantial observations in Swiss mountain pastures (
Figure 1) show that horsemint can efficiently outcompete other species, leading to extensive clusters where no other herbaceous species remain. These observations stimulated the question of whether horsemint exerts an allelopathic activity, and, if so, whether this activity is linked with its essential oils. We addressed this using the standard cress germination assay (
Figure 2), administering the essential oils or compounds found in different mint species through the gas phase. In fact, the essential oils from all three tested mint species (
M. spicata,
M. spicata crispa,
M. longifolia) exerted a dose-dependent and strong inhibition of germination, especially for the essential oils from
M. spicata crispa and
M. longifolia (
Figure 2A). The monoterpenes menthone/isomenthone (found in
M. longifolia, accompanying high levels of menthol [
19]) and linalool (found in small amounts in several mints) were highly active as well, while limonene (which is very abundant in
M. spicata crispa, but not in
M. spicata, [
19]) produced only a weak inhibition. The same holds true for α- and β-pinene that were found in small amounts in several mints. Likewise, n-hexane, used as solvent, did not show any effect.
Thus, there is a clear allelopathic inhibition of germination in cress. This inhibition is specific, since it differs depending on the source plant, and it is dependent on the dose. Among the individual compounds, menthone/isomenthone, which is abundant in horsemint, produced a strong inhibition, while limonene, which is abundant in
M. spicata crispa, inhibited germination only mildly, contrasting with the strong inhibition by the essential oil of
M. spicata crispa. Obviously, limonene cannot account for the strong effect of this oil, while the strong effect of horsemint oil correlates with a strong effect of its main compound menthone/isomenthone, which is present in this oil, albeit not as the main component. In addition, the development of the seedlings that managed to germinate was affected, as evidenced by the reduced root and hypocotyl elongation, as exemplarily shown for oil from
M. spicata and
M. spicata crispa (
Figure 2B,C).
3.2. Effect of Horsemint Oil Depends on the Target Species
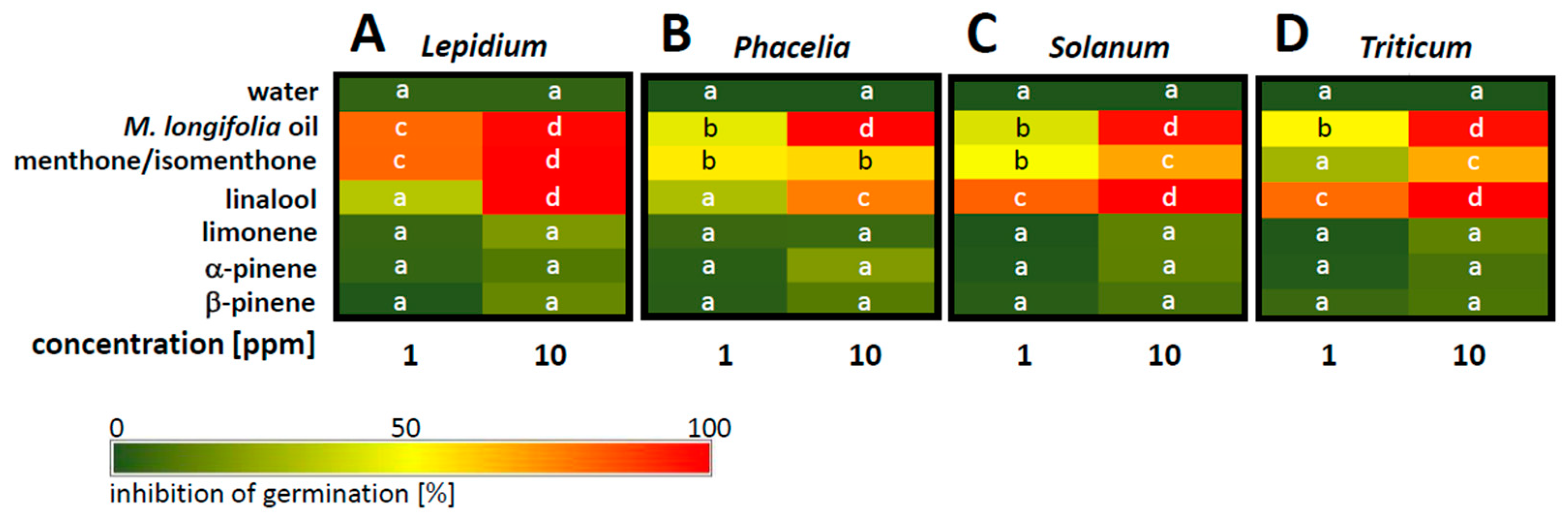
Since horsemint oil efficiently inhibited cress germination, even if just administered through the gas phase, we wondered whether this effect was general or whether it was dependent on the target species. We therefore compared the germination inhibition of
M. longifolia and the individual active compounds thereof for cress (
Figure 3A),
Phacelia tanacetifolia as a representative of legumes (
Figure 3B), tomato as a Solanaceous species (
Figure 3C), and winter wheat as a representative of cereals (
Figure 3D). While the overall pattern of inhibition was comparable at high concentrations, there were specific differences with respect to efficacy, especially at the lower concentration of 1 ppm. For instance, the essential oil of
M. longifolia, while inhibiting all target species at 10 ppm, was less efficient in
Phacelia and tomato than in winter wheat at 1 ppm, and the inhibition in winter wheat was, in turn, less pronounced than in cress. In contrast, for menthone, winter wheat was inhibited less than
Phacelia or tomato at 1 ppm. Linalool was most efficient in winter wheat and tomato, but less in cress and in
Phacelia, if administered at 1 ppm, while at 10 ppm, it inhibited all four targets. Limonene, as well as α- and β-pinene, did not show a particular effect in any of the four target species.
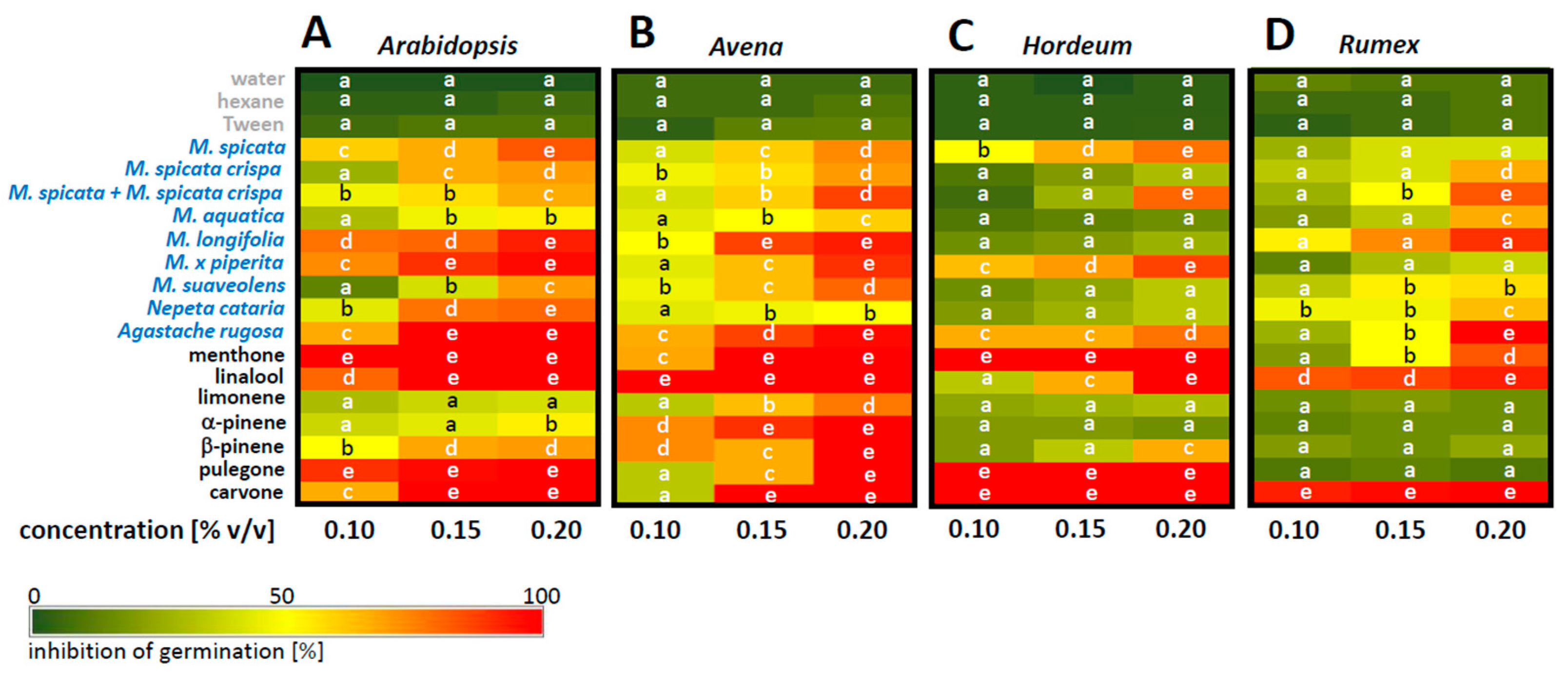
In the next step, we expanded the species spectrum as well as the spectrum of compounds. Moreover, since the effective concentration for the application through the gas phase can only be estimated based on the assumption that the compounds are equally distributed, we changed the application to the detergent-supported imbibition of the target seeds (
Figure 4). While none of the solvents (
n-hexane in the case of the pure compounds, Tween 20 for the essential oils) caused any inhibition, both the essential oils and the compounds found therein produced significant effects. These effects were not only dependent on concentration, but also on the donor species or the chemical nature of the compound, respectively. It was also dependent on the target species. These features indicate that the effect is specific and not caused by a general phytotoxicity.
With respect to the donor plants, the essential oils of M. longifolia and Agastache rugosa were generally the most potent inhibitors. M. x piperita was also effective, while M. suaveolens and M. spicata crispa only produced a weak effect. There were differences between the target species, though. For instance, the essential oil of M. longifolia efficiently suppressed the germination of Arabidopsis, Avena, and Rumex, while Hordeum remained largely unimpressed. On the other hand, Avena was blocked by the essential oil of M. x piperita, which in turn had only a low impact on Rumex. Thus, the sensitivity of a given target species is strongly dependent on the origin of the essential oil.
Similar conclusions can be drawn for the compounds found in these oils. While linalool is very potent against Arabidopsis, Avena, and Rumex, it is far less efficient in Hordeum. On the other hand, α- and β-pinene, which only produce weak inhibition in Hordeum and Rumex, are quite efficient in suppressing Avena. Menthone and carvone were the two compounds with a generally strong inhibitory effect, but even there, menthone was much less potent against Rumex, while (R)-carvone needed 0.15% v/v to efficiently suppress Avena.
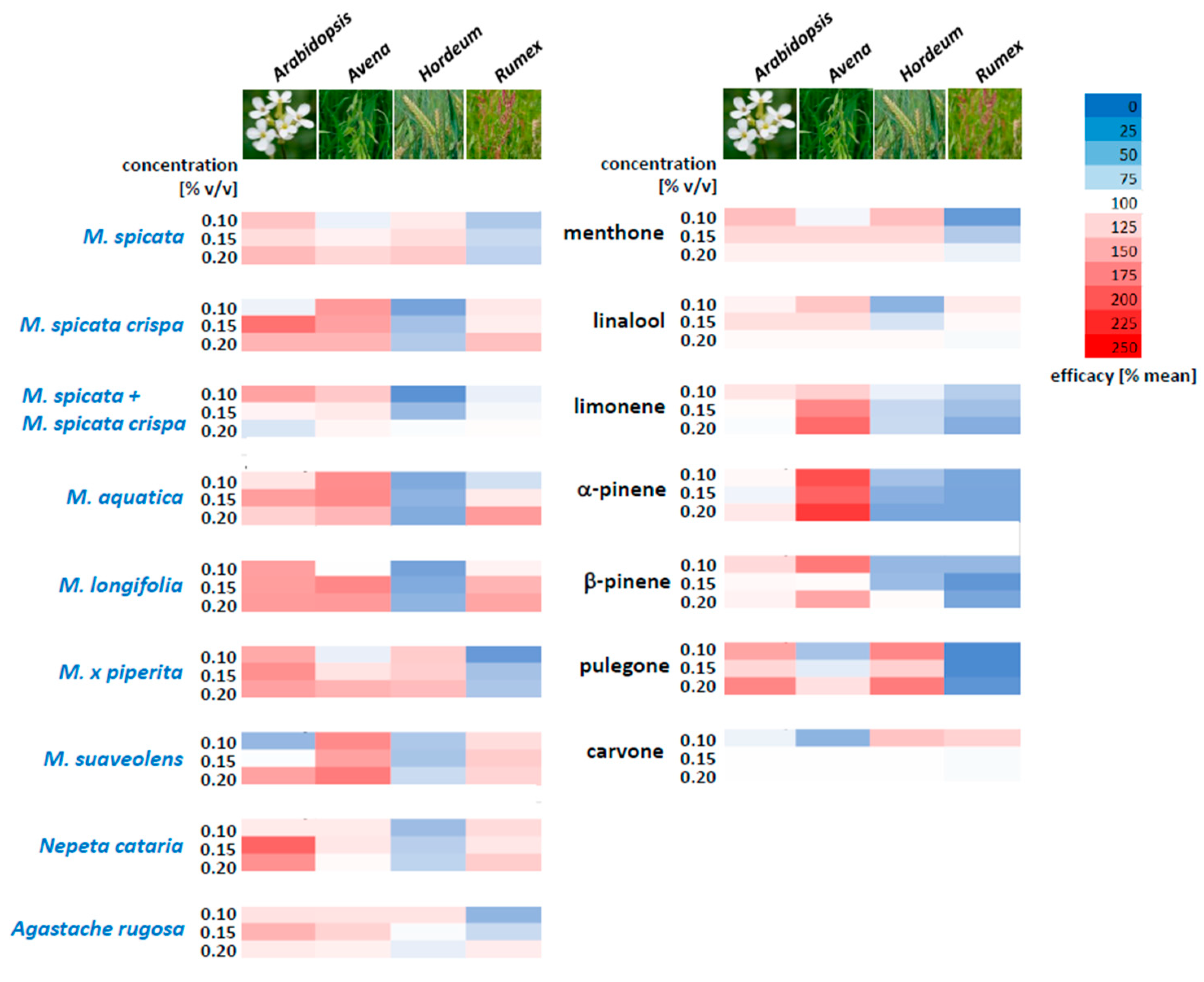
To detect whether a particular oil or a particular compound targets one species over the others, we determined the efficacy spectra (
Figure 5). For this purpose, we first determined the average inhibition for the compound of interest over the entire range of the four target species. In the next step, we related the inhibition for each individual target species to this average value. This approach revealed whether a given target species was specifically susceptible or resistant to this compound. Using this map, one can move along a column and see at a glance which compound generates a pronounced differential inhibition between two target species of interest.
Using this approach, one sees that, for instance, Arabidopsis is especially sensitive to essential oils from M. spicata crispa, M. longifolia, M. x piperita, and Nepeta cataria when compared to the other species, while Avena is also sensitive to M. suaveolens. Hordeum, which is generally quite robust, shows sensitivity to M. spicata and M. x piperita, but is not sensitive to M. longifolia, which, instead, can be used to address Rumex. With the individual compounds, the sensitivity of Avena to α-pinene, limonene, and β-pinene is noteworthy, while pulegone is of little effect, but shows good activity against Arabidopsis and Hordeum. Rumex can be tackled by carvone and also, to some extent, by linalool, which does not affect Hordeum.
3.3. Horsemint Oil Disrupts Actin Filaments, but Not Microtubules
To obtain insight into the mode of action of horsemint oil, we probed for the response of the cytoskeleton. Microtubules are not only essential for cell division driving the development of meristems in germinating seedlings, but also sustain cell axis in elongating tissues such as the distal elongation zone of the root or the hypocotyl. Actin filaments are not only essential for the elongation of root hairs, but also support the polar flow of auxin through the plant. Therefore, we exposed tobacco BY-2 cells expressing GFP-tagged markers for actin filaments and microtubules [
20,
21] to horsemint oil acting through the gas phase as described in [
19] and followed the responses using spinning disc confocal microscopy. While in cells exposed to the solvent control,
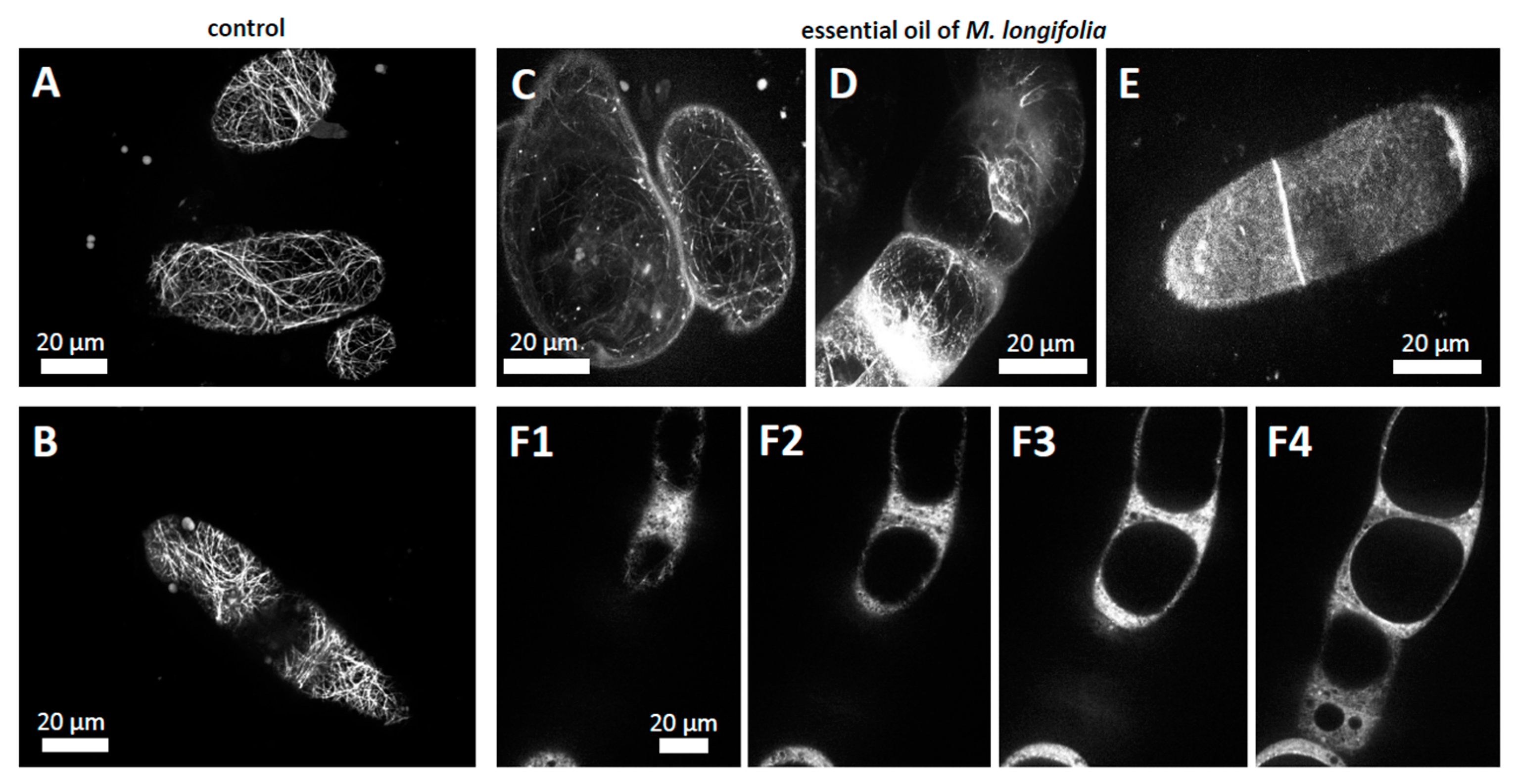
n-hexane, the actin filaments displayed a well-developed network (
Figure 6A,B) and the essential oil of
M. longifolia caused the almost complete breakdown of the actin cytoskeleton (
Figure 6C–E). The filaments were reduced to very short rod-like structures along with a soluble signal that often accumulates along the cross-walls (
Figure 6D–E). In confocal z-stacks (
Figure 6(F1–F4)), individual fluorescent
punctae could be discerned on the surface of the vacuole, probably representing actin nucleation centres.
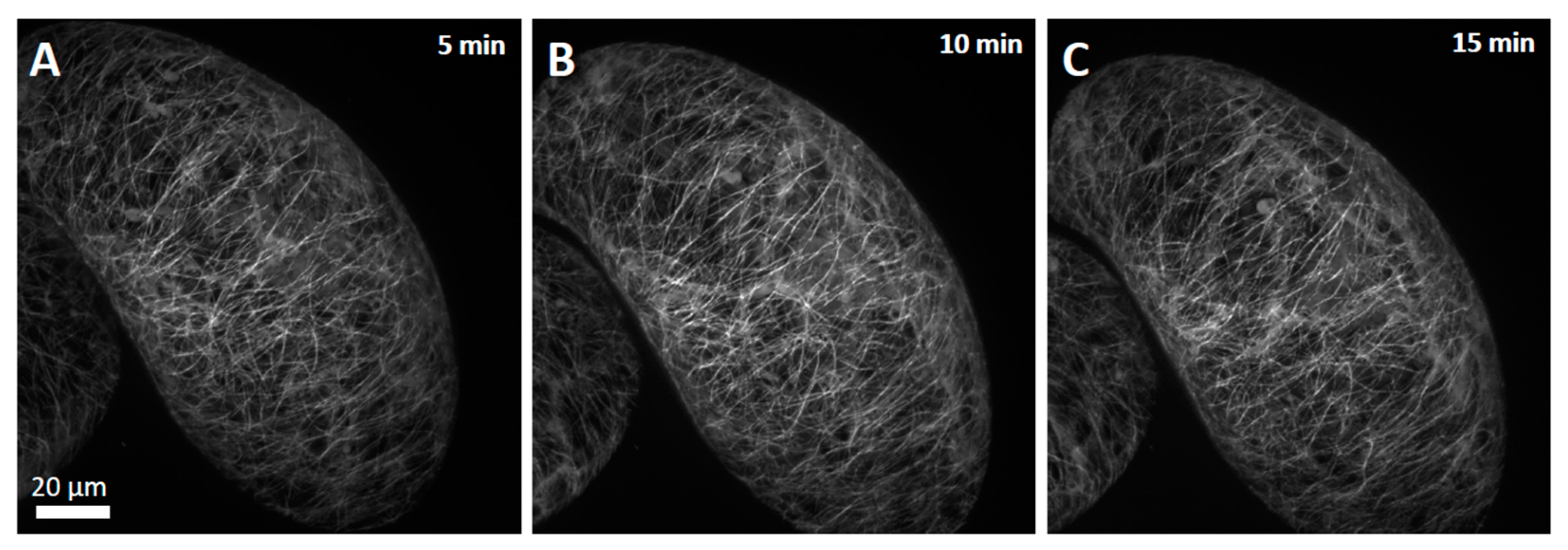
To assess whether this breakdown of the actin cytoskeleton was reflecting a general disruption of the cytoskeletal structure, we investigated the response of the microtubules in parallel. However, the microtubules that were organised in cortical arrays perpendicular to the axis of cell elongation persisted (
Figure 7), contrasting the complete collapse of the actin filaments observed under the same conditions.
3.4. Horsemint Oil Can Be Used to Control Bindweed
Since the effect of horsemint oil was specific for actin filaments, we wondered whether this could be exploited for specific weed control. Actin filaments are essential for polar auxin transport [
22], which in turn is crucial to the regeneration of adventitious roots [
23]. We wondered, therefore, whether a weed relying on vegetative regeneration could be targeted by this essential oil. For this proof-of-concept, we selected bindweed (
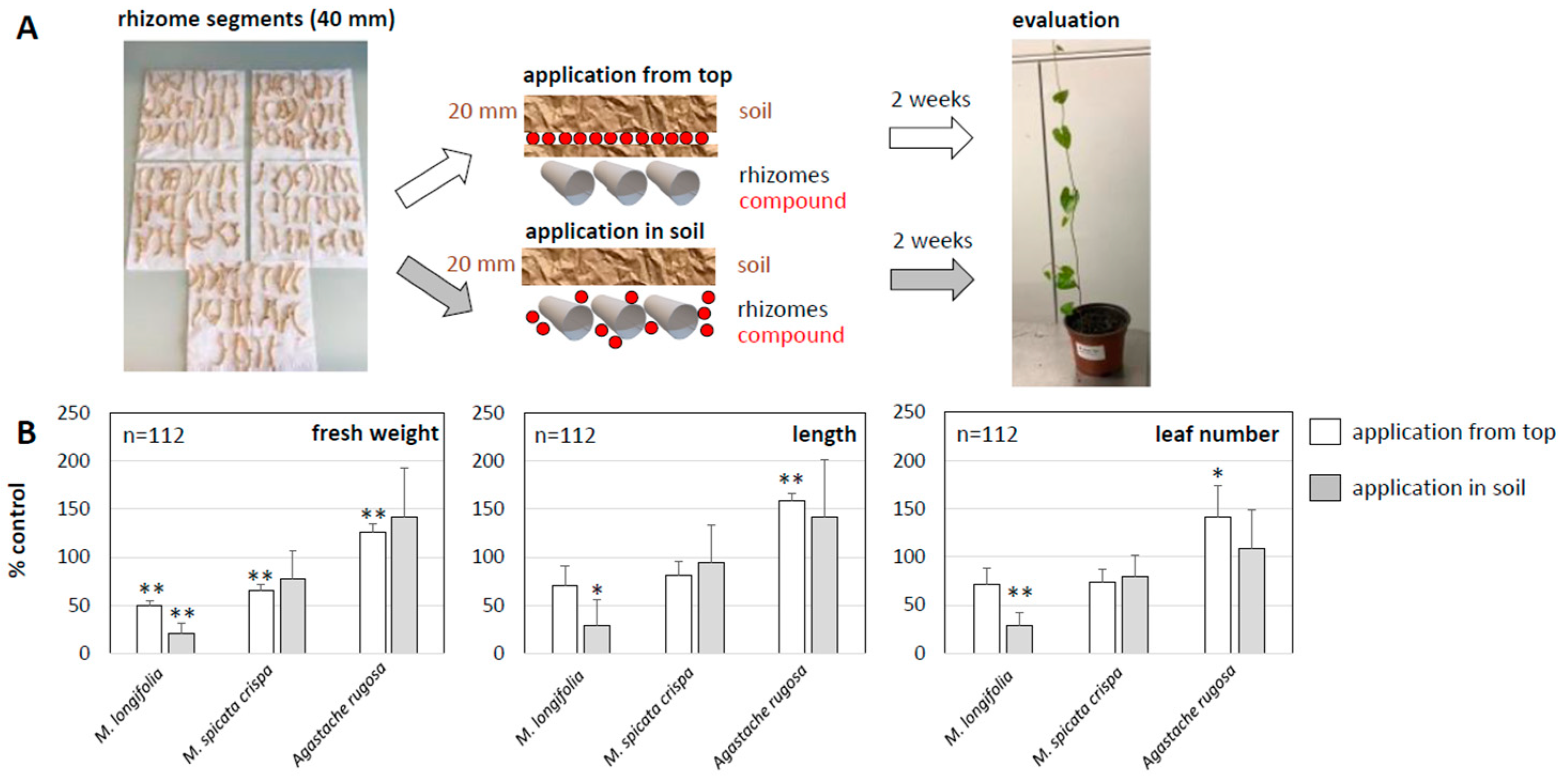
Convolvulus arvensis), a pertinent problem in organic cereals. This weed mainly invades from rhizomes that can reach several metres in length and are difficult to tackle, even using herbicides. In organic cereal culture, it has to be controlled by intensive tilling, which is not only laborious, but also contributes to erosion. We tested two modes of application of horsemint oil mixed into a slow-release carrier in a pot experiment with rhizome segments of bindweed (
Figure 8A): either the carrier was placed on top of the rhizomes and covered by soil, or, alternatively, the carrier was worked into the soil harbouring the rhizomes. The results were assessed after two weeks. To probe for specificity, we used oil from
M. spicata crispa and of
Agastache rugosa in parallel, along with a negative control, where the carrier was applied without being doped with oil.
In fact, we observed a significant and substantial inhibition of bindweed development in response to the treatment, particularly in the setup where the doped carrier had been worked into the soil (
Figure 8B). Compared to the control value, which was set to 100%, the fresh weight was reduced by a factor of four, and for the application from the top, the inhibition was weaker (a factor of two). A similarly strong inhibition was seen for the leaf number, although it was weaker for shoot length, indicating that bindweed was still partially sustaining cell elongation, but had problems forming new organs and, thus, accumulating biomass. The inhibitory effect was specific for horsemint oil, since oil from
M. spicata crispa, with exception of a reduction in fresh weight by around 40%, failed to evoke significant effects. For the essential oil of
Agastache rugosa, we even obtained values that were larger than the control values (defined as 100%). In other words, this oil caused significant stimulation, especially with respect to elongation, such that the measured values turned out to be larger than in the controls.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







