
The Effect of Glutamine as Feed Additive on Selected Parameters of the Nonspecific Immune Response in Pigs
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Blood Samples Collection
2.3. Determination of Selected Immune Response Parameters
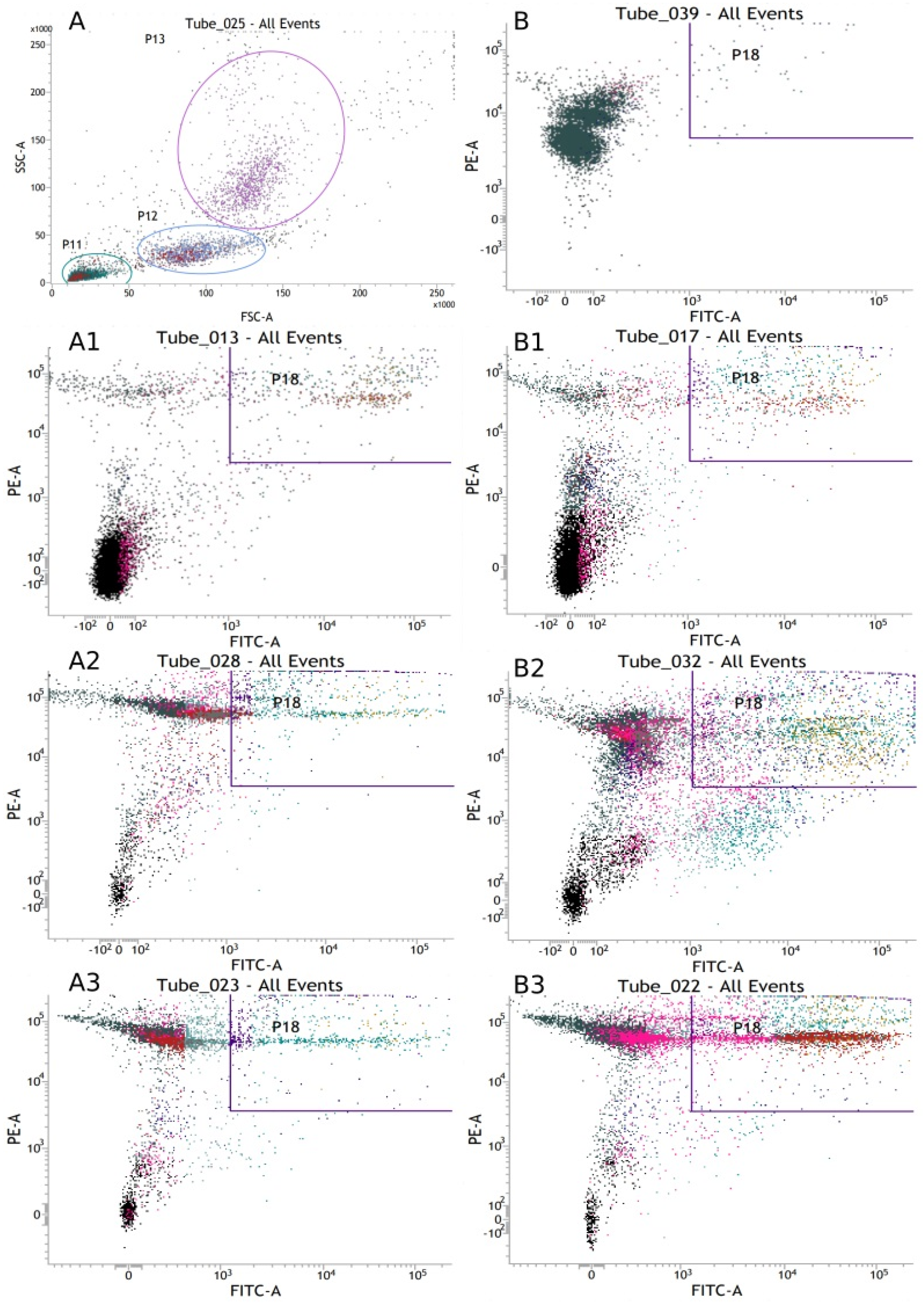
2.4. Assessment of Phagocytosis
2.5. Respiratory Burst of Granulocytes and Monocytes
2.6. Determination of the Bacteriolytic Activity of Lysozyme (LZM) in the Peripheral Blood of Pigs
2.7. Assay of IL-1β, IL-6, IL-10 and TNF-α in Pig Serum
2.8. Assay of L-Glutamine and Glutamate in Pig Serum
2.9. Statistical Analysis
3. Results
3.1. Determination of Selected Immune Response Parameters
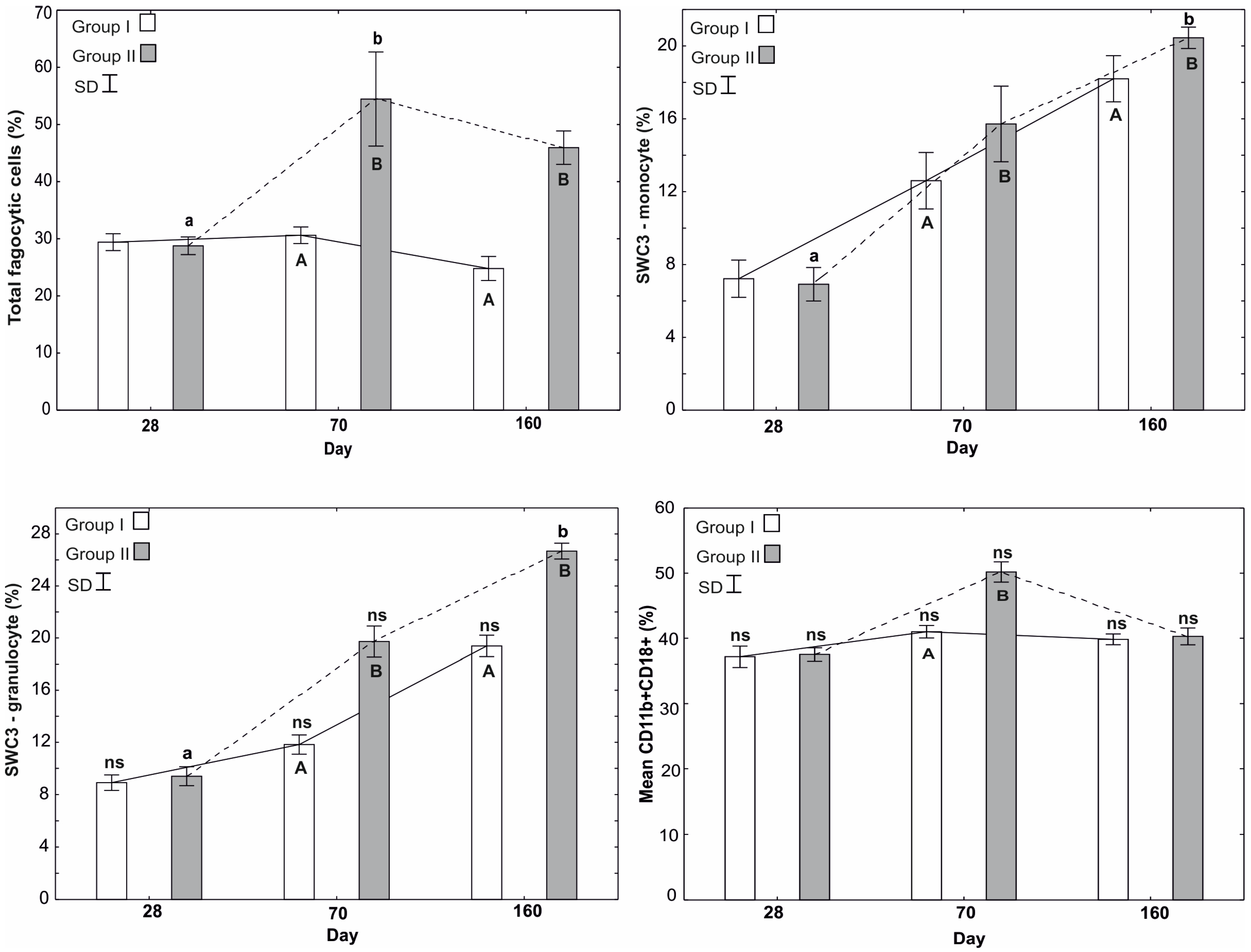
3.1.1. Mean Total Phagocyte Count
3.1.2. Percentages of SWC3-Monocytes and SWC3-Granulocytes
3.1.3. Mean Percentage of CD11b+CD18+
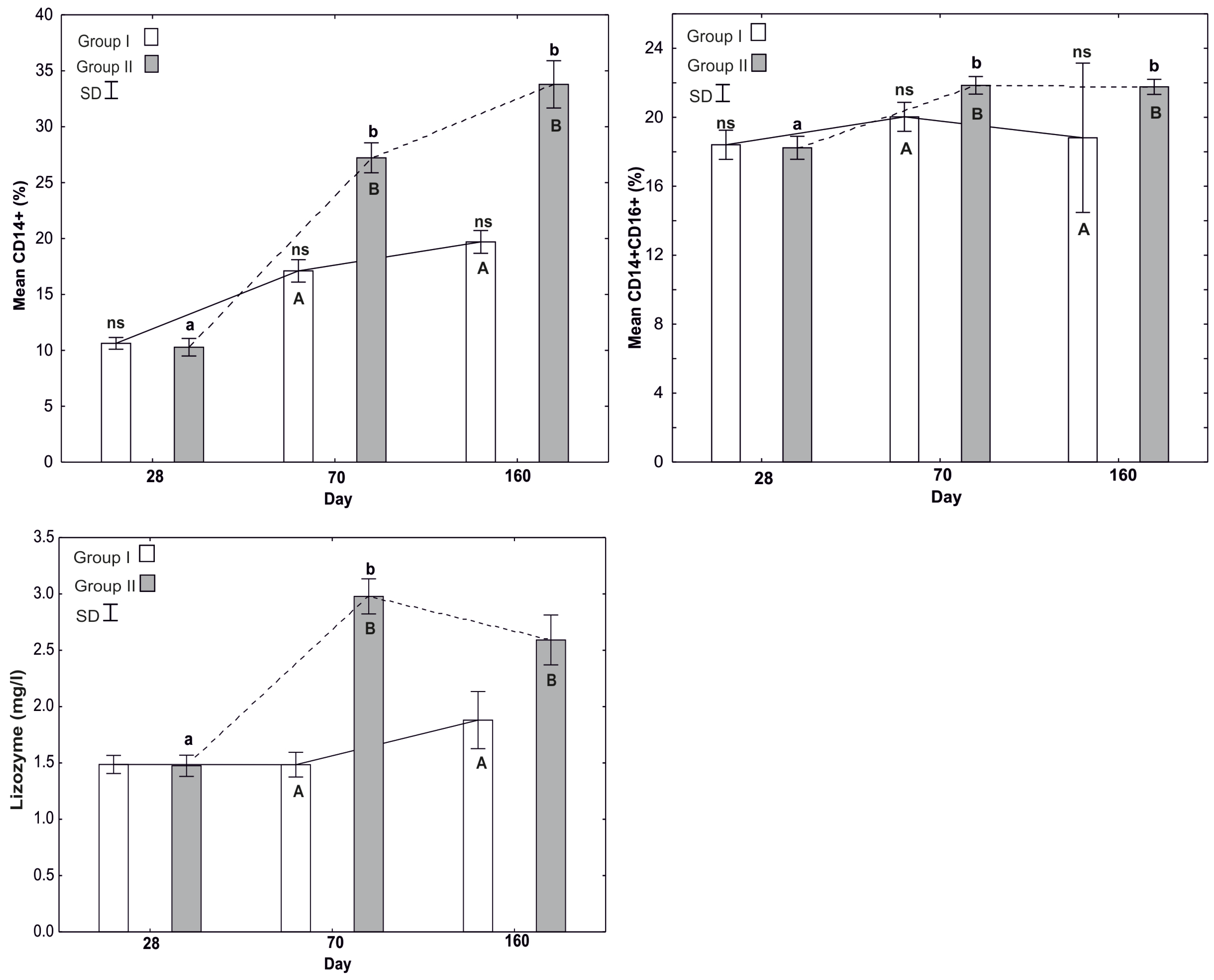
3.1.4. Mean Percentage of CD14+
3.1.5. Mean Percentage of CD14+CD16+
3.1.6. Oxidative Burst of Granulocytes and Monocytes
3.1.7. Mean Fluorescence Intensity of Peripheral Blood Phagocytic Cells
3.2. Determination of Bacteriolytic Activity of Lysozyme (LZM) in the Peripheral Blood of Pigs
3.3. Serum Concentrations of IL-1β, IL-6, IL-10, and TNF-α
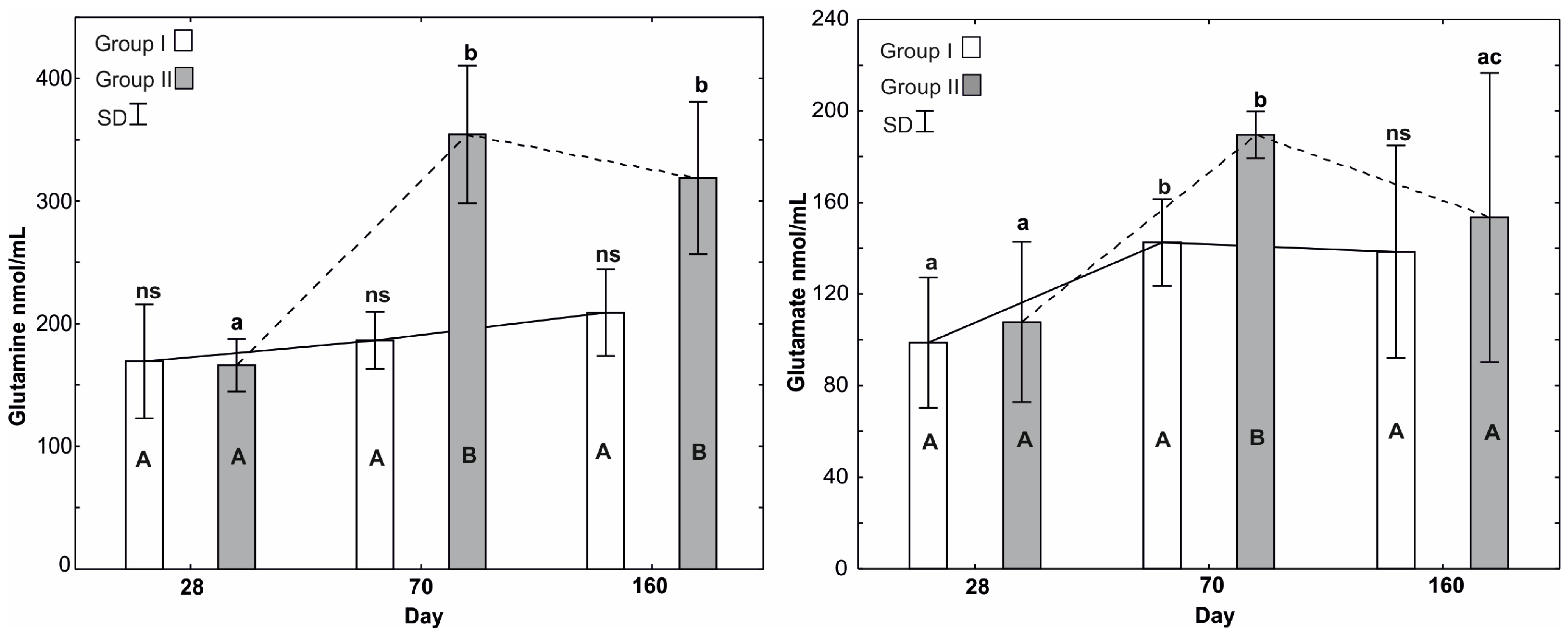
3.4. Mean Glutamine and Glutamate Levels
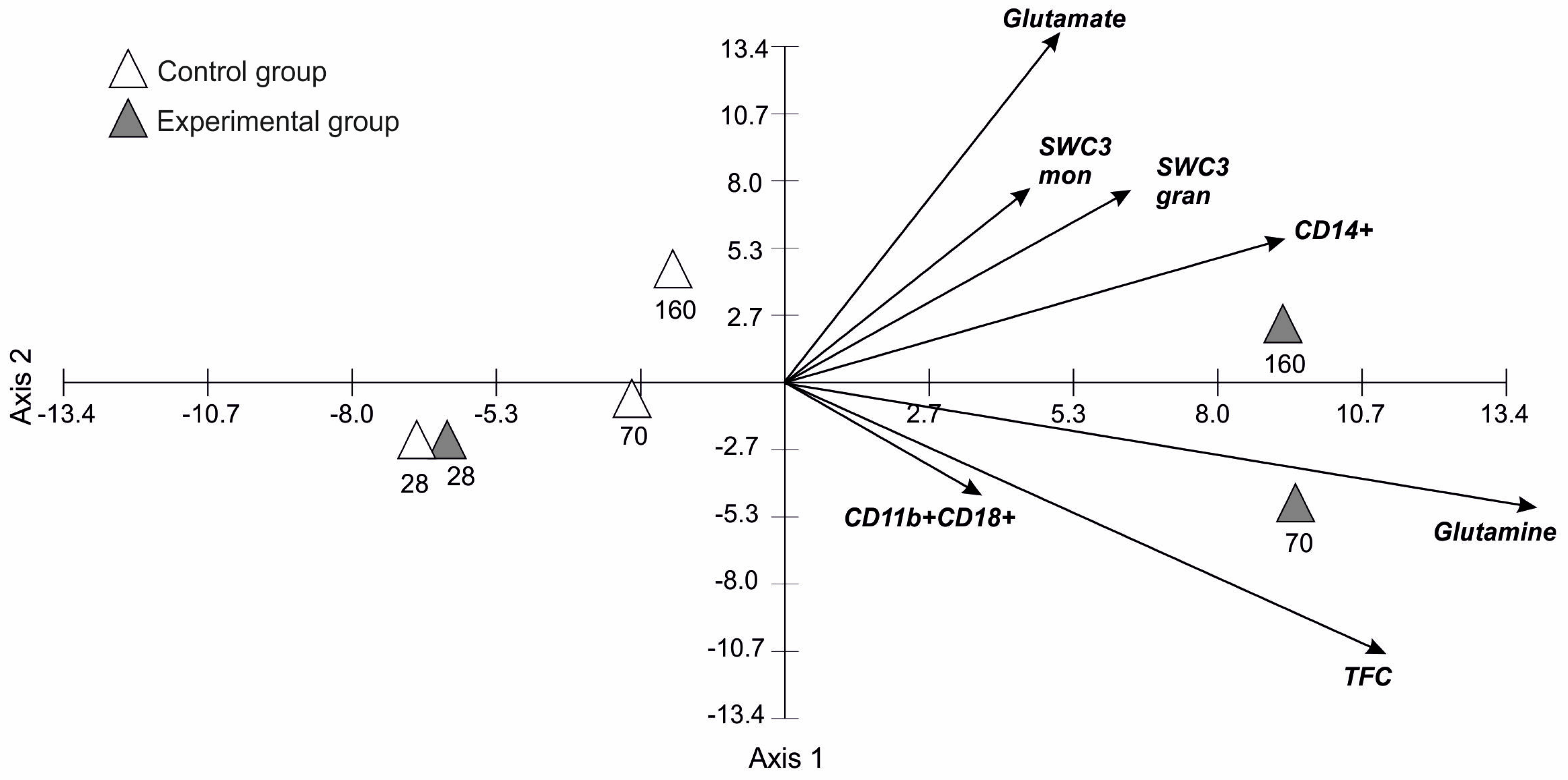
3.5. Principal Component Analysis for Blood Parameters in Time
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tokach, M.D.; Goodband, B.D.; O’Quinn, T.G. Performance-enhancing technologies in swine production. Anim. Front. 2016, 6, 15–21. [Google Scholar] [CrossRef]
- Blavi, L.; Solà-Oriol, D.; Llonch, P.; López-Vergé, S.; Martín-Orúe, S.M.; Pérez, J.F. Management and feeding strategies in early life to increase piglet performance and welfare around weaning: A Review. Animals 2021, 11, 302. [Google Scholar] [CrossRef]
- He, J.; Feng, G.D.; Ao, X.; Li, Y.F.; Qian, H.X.; Liu, J.B.; He, Z.Z. Effects of L-glutamine on growth performance, antioxidant ability, immunity and expression of genes related to intestinal health in weanling pigs. Livest. Sci. 2016, 189, 102–109. [Google Scholar] [CrossRef]
- Zou, T.D.; Deng, C.X.; Wang, Z.R.; Ye, Y.L.; You, J.M. Dietary alanyl-glutamine improves growth performance of weaned piglets through maintaining intestinal morphology and digestion-absorption function. Animal 2019, 13, 1826–1833. [Google Scholar] [CrossRef]
- Liao, S.F. Invited Review: Maintain or improve piglet gut health around weanling: The fundamental effects of dietary amino acids. Animals 2021, 11, 1110. [Google Scholar] [CrossRef]
- Newsholme, P.; Curi, R.; Curi, T.C.P.; Murphy, C.J.; Garcia, C.; de Melo, M.P. Glutamine metabolism by lymphocytes, macrophages, and neutrophils: Its importance in health and disease. J. Nutr. Biochem. 1999, 10, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Procopio, J.; Lima, M.M.R.; Pithon-Curi, T.C.; Curi, R. Glutamine and glutamate—Their central role in cell metabolism and function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef]
- Alverdy, J.C. Effects of glutamine-supplemented diets on immunology of the gut. J. Parenter. Enteral Nutr. 1990, 14, 109–113. [Google Scholar] [CrossRef]
- Ji, F.J.; Wang, L.X.; Yang, H.S.; Hu, A.; Yin, Y.L. Review: The roles and functions of glutamine on intestinal health and performance of weaning pigs. Animal 2019, 13, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Duttlinger, A.W.; Kpodo, K.R.; Schinckel, A.P.; Richert, B.T.; Johnson, J.S. Effects of increasing dietary L-glutamine to replace antibiotics on pig health and performance following weaning and transport. Transl. Anim. Sci. 2020, 4, txaa157. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional intervention for the intestinal development and health of weaned pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Coeffier, M.; Miralles-Barrachina, O.; Le Pessot, F.; Lalaude, O.; Daveau, M.; Lavoinne, A.; Lerebours, E.; Dechelotte, P. Influence of glutamine on cytokine production by human gut in vitro. Cytokine 2001, 13, 148–154. [Google Scholar] [CrossRef]
- Field, C.J.; Johnson, I.R.; Schley, P.D. Nutrients and their role in host resistance to infection. J. Leukoc. Biol. 2002, 71, 16–32. [Google Scholar] [CrossRef]
- Hsu, C.B.; Lee, J.W.; Huang, H.J.; Wang, C.H.; Lee, T.T.; Yen, H.T.; Yu, B. Effects of supplemental glutamine on growth performance, plasma parameters and LPS-induced immune response of weaned barrows after castration. Asian Australas. J. Anim. Sci. 2012, 25, 674–681. [Google Scholar] [CrossRef]
- Singleton, K.D.; Wischmeyer, P.E. Glutamine attenuates inflammation and NF-κB activation via Cullin-1 deneddylation. Biochem. Biophys. Res. Commun. 2008, 373, 445–449. [Google Scholar] [CrossRef]
- Wu, L.; Wang, W.; Yao, K.; Zhou, T.; Yin, J.; Li, T.; Yang, L.; He, L.; Yang, X.; Zhang, H.; et al. Effects of dietary arginine and glutamine on alleviating the impairment induced by deoxynivalenol stress and immune relevant cytokines in growing pigs. PLoS ONE 2013, 8, e69502. [Google Scholar] [CrossRef]
- Yu, I.T.; Wu, J.F.; Yang, P.C.; Liu, C.Y.; Lee, D.N.; Yen, H.T. Roles of glutamine and nucleotides in combination in growth, immune responses and FMD antibody titres of weaned pigs. Anim. Sci. 2002, 75, 379–385. [Google Scholar] [CrossRef]
- Wasinski, F.; Gregnani, M.F.; Ornellas, F.H.; Bacurau, A.V.N.; Câmara, N.O.; Araujo, R.C.; Bacurau, R.F. Lymphocyte glucose and glutamine metabolism as targets of the anti-inflammatory and immunomodulatory effects of exercise. Mediators Inflamm. 2014, 2014, 326803. [Google Scholar] [CrossRef]
- Wells, S.M.; Kew, S.; Yaqoob, P.; Wallace, F.A.; Calder, P.C. Dietary glutamine enhances cytokine production by murine macrophages. Nutrition 1999, 15, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.R.; Skomiał, J. (Eds.) Polish Pig Feeding Recommendations. In The Nutritional Recommendations and Nutritional Value of Pig Feed; Kielanowski Institute of Animal Physiology and Nutrition Polish Academy of Sciences: Jabłonna, Poland, 2020; pp. 1–125. (In Polish) [Google Scholar]
- Laskowska, E.; Jarosz, Ł.S.; Grądzki, Z. Effect of the EM Bokashi® Multimicrobial Probiotic Preparation on the Non-specific Immune Response in Pigs. Probiotics Antimicro. Prot. 2019, 11, 1264–1277. [Google Scholar] [CrossRef]
- Sinkora, M.; Sinkorova, J.; Holtmeier, W. Development of γδ thymocyte subsets during prenatal and postnatal ontogeny. Immunology 2005, 115, 544–555. [Google Scholar] [CrossRef]
- Hankiewicz, J.J.; Świerczek, E.E. Studies on serum and urinary lysozyme. Pol. Arch. Intern. Med. 1974, 51, 591–597. [Google Scholar]
- Kovach, W. MVSP—A Multivariate Statistical Package for Windows, version 3.1; Kovach Computing Services: Pentraeth, UK, 1999. [Google Scholar]
- Pithon-Curi, T.C.; De Melo, M.P.; Curi, R. Glucose and glutamine utilization by rat lymphocytes, monocytes and neutrophils in culture: A comparative study. Cell Biochem. Funct. 2004, 22, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immune 2010, 32, 593–604. [Google Scholar] [CrossRef]
- Liang, M.; Wang, X.; Yuan, Y.; Zhou, Q.; Tong, C.; Jiang, W. Different effect of glutamine on macrophage tumor necrosis factor-alpha release and heat shock protein 72 expression in vitro and in vivo. Acta Biochim. Biophys. Sin. 2009, 41, 171–177. [Google Scholar] [CrossRef]
- Wallace, C.; Keast, D. Glutamine and macrophage function. Metabolism 1992, 41, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Xia, Y.; Chen, S.; Wu, G.; Bazer, F.W.; Zhou, B.; Tan, B.; Zhu, G.; Deng, J.; Yin, Y. Glutamine metabolism in macrophages: A novel target for obesity/type 2 diabetes. Adv. Nutr. 2019, 10, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Di Giancamillo, A.; Domeneghini, C.; Paratte, R.; Dell’Orto, V.; Bontempo, V. Oral feeding with L-glutamine and nucleotides: Impact on some GALT (gut associated lymphoid tissue) parameters and cell proliferation/death rates in weaning piglets. Ital. J. Anim. Sci. 2003, 2, 364–366. [Google Scholar]
- Rao, R.; Samak, G. Role of glutamine in protection of intestinal epithelial tight junctions. J. Epithel. Biol. Pharmacol. 2012, 5, 47–54. [Google Scholar] [PubMed]
- Shan, Y.; Shan, A.; Li, J.; Zhou, C. Dietary supplementation of arginine and glutamine enhances the growth and intestinal mucosa development of weaned piglets. Livest. Sci. 2012, 150, 369–373. [Google Scholar] [CrossRef]
- Spittler, A.; Winkler, S.; Gotzinger, P.; Oehler, R.; Willhiem, M.; Tempfer, C.; Weigel, G.; Fugger, R.; Boltz-Nitulescu, G.; Roth, E. Influence of glutamine on the phenotype and function of human monocytes. Blood 1995, 86, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Parry-Billings, M.; Evans, J.C.; Calder, P.C.; Newsholme, E.A. Does glutamine contribute to immunosuppression after burns? Lancet 1990, 336, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Curi, T.C.; De Melo, M.P.; De Azevedo, R.B.; Zorn, T.M.; Curi, R. Glutamine utilization by rat neutrophils: Presence of phosphate-dependent glutaminase. Am. J. Physiol. Cell Physiol. 1997, 273, 1124–1129. [Google Scholar] [CrossRef]
- Spittler, A.; Holzer, S.; Oehler, R.; Boltz-Nitulescu, G.; Roth, E. A glutamine deficiency impairs the function of cultured human monocytes. Clin. Nutr. 1997, 16, 97–99. [Google Scholar] [CrossRef]
- Furukawa, S.; Saito, H.; Inoue, T.; Matsuda, T.; Fakatsu, K.; Han, I.; Ikeda, S.; Hidemura, A. Supplemental glutamine augments phagocytosis and reactive oxygen intermediate production by neutrophils and monocytes from postoperative patients in vitro. Nutrition 2000, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Vega-Lopez, M.A.; Bailey, M.; Telemo, E.; Stokes, C.R. Effect of early weaning on the development of immune cells in the pig small intestine. Vet. Immunol. Immunopathol. 1995, 44, 319–327. [Google Scholar] [CrossRef]
- Bain, C.C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef]
- Xing, S.; Zhang, B.; Lin, M.; Zhou, P.; Li, J.; Zhang, L.; Gao, F.; Zhou, G. Effects of alanyl-glutamine supplementation on the small intestinal mucosa barrier in weaned piglets. Asian Australas. J. Anim. Sci. 2017, 30, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Kallio, R.; Aalto, H.; Takala, A.; Ohtonen, P.; Collan, J.; Siitonen, S.; Joensuu, H.; Syrjala, H.; Repo, H. Expression of CD11b/CD18 adhesion molecules on circulating phagocytes-a novel aid to diagnose infection in patients with cancer. Support Care Cancer 2008, 16, 1389–1396. [Google Scholar] [CrossRef] [PubMed]
- May, R.C.; Machesky, L.M. Phagocytosis and the actin cytoskeleton. J. Cell Sci. 2001, 114, 1061–1077. [Google Scholar] [CrossRef]
- Fairbairn, L.; Kapetanovic, R.; Beraldi, D.; Sester, D.P.; Tuggle, C.K.; Archibald, A.L.; Hume, D.A. Comparative analysis of monocyte subsets in the pig. J. Immunol. 2013, 190, 6389–6396. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, S.; Revilla, C.; Alvarez, B.; Alonso, F.; Ezquerra, A.; Domínguez, J. Phenotypic and functional heterogeneity of porcine blood monocytes and its relation with maturation. Immunology 2005, 114, 63–71. [Google Scholar] [CrossRef]
- Fabriek, B.O.; Dijkstra, C.D.; van den Berg, T.K. The macrophage scavenger receptor CD163. Immunobiology 2005, 210, 153–160. [Google Scholar] [CrossRef]
- Zhao, C.; Tan, Y.C.; Wong, W.C.; Sem, X.; Zhang, H.; Han, H.; Ong, S.M.; Wong, K.L.; Yeap, W.H.; Sze, S.K.; et al. The CD14+/lowCD16+ monocyte subset is more susceptible to spontaneous and oxidant-induced apoptosis than the CD14+CD16− subset. Cell Death Dis. 2010, 1, e95. [Google Scholar] [CrossRef]
- Ziegler-Heitbrock, L. The CD14+ CD16+ blood monocytes: Their role in infection and inflammation. J. Leukoc. Biol. 2007, 81, 584–592. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, Y.; Dong, W.; Zhu, G.; Wu, S.; Bao, W. CD14 in the TLRs signaling pathway is associated with the resistance to E. coli F18 in Chinese domestic weaned piglets. Sci. Rep. 2016, 6, 24611. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Kiarie, E.; Bhandari, S.K.; Zhang, G.; Krause, D.O. Weaned pig responses to Escherichia coli K88 oral challenge when receiving a lysozyme supplement. J. Anim. Sci. 2012, 90, 252–260. [Google Scholar] [CrossRef]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed]
- Yassad, A.; Lavoinne, A.; Bion, A.; Daveau, M.; Husson, A. Glutamine accelerates interleukin-6 production by rat peritoneal macrophages in culture. FEBS Lett. 1997, 413, 81–84. [Google Scholar] [CrossRef]
- Murphy, C.J.; Newsholme, P. The importance of glutamine metabolism in murine macrophages and human monocytes to L-arginine biosynthesis and rates of nitrite or urea production. Clin. Sci. 1998, 95, 397–407. [Google Scholar] [CrossRef]
- De Moreno de LeBlanc, A.; Chaves, S.; Carmuega, E.; Weill, R.; Antoine, J.; Perdigon, G. Effect of long-term continuous consumption of fermented milk containing probiotic bacteria on mucosal immunity and the activity of peritoneal macrophages. Immunobiology 2008, 213, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Zoumpopoulou, G.; Foligne, B.; Christodoulou, K.; Grangette, C.; Pot, B.; Tsakalidou, E. Lactobacillus fermentum ACA-DC 179 displays probiotic potential in vitro and protects against trinitrobenzene sulfonic acid (TNBS)-induced colitis and Salmonella infection in murine models. Int. J. Food Microbiol. 2008, 121, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Nowland, T.L.; Plush, K.J.; Barton, M.; Kirkwood, R.N. Development and function of the intestinal microbiome and potential implications for pig production. Animals 2019, 9, 76. [Google Scholar] [CrossRef]
- Pie, S.; Lalles, J.P.; Blazy, F.; Laffitte, J.; Séve, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [PubMed]
- Pardo, A.L.; Poveda, A.P.; da Silva, C.; dos Santos, A.; Venâncio, E.; Arantes, V.; Nogueira, E. Effect of L-glutamine levels in piglets diets challenged with Escherichia coli lipopolysacharides. Revista MVZ Córdoba 2014, 19, 4328–4337. [Google Scholar] [CrossRef]
- Yi, G.F.; Carroll, J.A.; Allee, G.L.; Gaines, A.M.; Kendall, D.C.; Ursy, J.L.; Toride, Y.; Izuru, S. Effect of glutamine and spray-dried plasma on growth performance, small intestinal morphology, and immune responses of Escherichia coli K88+-challenged weaned pigs. J. Anim. Sci. 2005, 83, 634–643. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Murdoch, G.K.; Johnson, I.R.; Madsen, K.L.; Field, C.J. Glutamine supplementation improves intestinal barrier function in a weaned piglet model of Escherichia coli infection. Brit. J. Nutr. 2011, 106, 870–877. [Google Scholar] [CrossRef]
- Cabrera, R.A.; Ursy, J.L.; Arellano, C.; Nogueira, E.T.; Kutschenko, M.; Moeser, A.J. Effects of creep feeding and supplemental glutamine or glutamine plus glutamate (Aminogut) on pre- and post-weaning growth performance and intestinal health of piglets. J. Anim. Sci. Biotech. 2013, 4, 29. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Phase 0 Preweaning, 7–28 Days of Age | Phase 1 Prestarter, 28–42 Days of Age about 13–20 kg Body Weight | Phase 2 Starter, 42–70 Days of Age about 20–35 kg Body Weight | Phase 3 Grower, 71–98 Days of Age about 35–60 kg Body Weight | Phase 4 Finisher, 98–160 Days of Age about 60–100 kg Body Weight) | ||
|---|---|---|---|---|---|---|---|
| Experimental Period | |||||||
| Ingredient (%) | All animals | Group I | Group II | Group I | Group II | All animals | All animals |
| Wheat | 24.82 | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Barley | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 38.42 | 39.04 |
| Triticale | - | 10.0 | 10.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Corn | 19.0 | 27.62 | 27.12 | 25.36 | 24.86 | - | - |
| Soybean meal | 10.0 | 16.0 | 16.0 | 18.0 | 18.0 | 14.0 | 12.0 |
| Wheat bran | - | - | - | 10.0 | 10.0 | 12.0 | 15.0 |
| Soybean oil | 4.0 | 3.3 | 3.3 | 3.0 | 3.0 | 2.5 | 1.5 |
| Skimmed milk powder | 12.0 | 10.0 | 10.0 | - | - | - | - |
| Dried whey | 10.0 | 5.0 | 5.0 | - | - | - | - |
| Fish meal | 8.0 | - | - | - | - | - | - |
| Sodium chloride | 0.1 | 0.15 | 0.15 | 0.32 | 0.32 | 0.28 | 0.26 |
| Phosphate 1-Ca | - | 0.28 | 0.28 | 0.50 | 0.50 | 0.30 | 0.15 |
| Limestone | 0.8 | 1.20 | 1.20 | 1.50 | 1.50 | 1.40 | 1.2 |
| Vitamin/mineral premix | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| L-Lysine | 0.37 | 0.48 | 0.48 | 0.45 | 0.45 | 0.34 | 0.23 |
| DL-Methionine | 0.24 | 0.25 | 0.25 | 0.17 | 0.17 | 0.11 | 0.03 |
| L-Threonine | 0.17 | 0.22 | 0.22 | 0.20 | 0.20 | 0.15 | 0.09 |
| L-Glutamine | - | - | 0.5 | - | 0.5 | - | - |
| Item | Phase 0 Preweaning, 7–28 Days of Age | Phase 1 Prestarter, 28–42 Days of Age about 13–20 kg Body Weight | Phase 2 Starter, 42–70 Days of Age about 20–35 kg Body Weight | Phase 3 Grower, 71–98 Days of Age about 35–60 kg Body Weight | Phase 4 Finisher, 98–160 Days of Age about 60–100 kg Body Weight) |
|---|---|---|---|---|---|
| Experimental Period | |||||
| Metabolizable Energy (MJ) | 14.2 | 13.9 | 13.4 | 13.0 | 12.6 |
| Crude protein | 200.0 | 180.0 | 170.0 | 160.0 | 156.0 |
| Dry matter | 894.0 | 888.0 | 880.0 | 880.0 | 880.0 |
| Crude fiber | 20.2 | 24.0 | 35.0 | 43.0 | 45.0 |
| Crude ash | 58.7 | 52.0 | 53.2 | 52.8 | 49.5 |
| Ether extract | 60.6 | 52.8 | 52.3 | 44.2 | 35.4 |
| N-free extractives | 554.5 | 579.2 | 569.5 | 580.0 | 594.1 |
| Phosphorus (digestible) | 4.0 | 3.0 | 2.7 | 2.5 | 2.2 |
| Calcium | 8.5 | 7.6 | 7.6 | 6.8 | 5.6 |
| Lysine | 16.1 | 14.2 | 12.2 | 10.5 | 9.1 |
| Methionine + Cystine | 9.8 | 8.5 | 7.4 | 6.6 | 5.7 |
| Threonine | 10.0 | 8.8 | 7.6 | 6.8 | 5.8 |
| Tryptophan | 2.8 | 2.5 | 2.4 | 2.5 | 2.4 |
| Day | Mean Fluorescence Intensity (%) | |
|---|---|---|
| Group I | Group II | |
| 28 | 188.75 ± 22.2 a | 183.47 ± 23.1 a |
| 70 | 156.87 ± 15.4 bA | 299.02 ± 23.1 bB |
| 160 | 170.36 ± 10.6 bA | 287.24 ± 12.8 bB |
| Day | Oxidized Cells (%) E. coli Stimulation | |
|---|---|---|
| Group I | Group II | |
| 28 | 49.23 ± 6.3 a | 47.9 ± 7.8 a |
| 70 | 59.62 ± 7.2 aA | 82.14 ± 11.2 bB |
| 160 | 56.1 ± 9.3 aA | 72.4 ± 6.5 bB |
| IL-1β | IL-6 | TNF-α | IL-10 | |||||
|---|---|---|---|---|---|---|---|---|
| Day | I | II | I | II | I | II | I | II |
| 28 | 34.91 ± 2.25 a | 34.22 ± 3.82 a | 41.82 ± 4.53 a | 44.12 ± 5.11 a | 27.98 ± 6.54 a | 26.64 ± 3.22 a | 45.25 ± 6.54 a | 43.21 ± 8.16 a |
| 70 | 45.18 ± 6.11 aA | 93.11 ± 7.38 bB | 61.35 ± 7.25 aA | 127.42 ± 11.62 bB | 38.16 ± 9.25 aA | 112.62 ± 11.08 bB | 41.13 ± 6.22 a | 46.72 ± 9.25 a |
| 160 | 51.21 ± 5.14 bA | 88.74 ±6.72 bB | 129.12 ± 13.21 b | 135.42 ± 10.29 b | 47.23 ± 12.33 bA | 84.94 ± 11.13 cB | 58.23 ± 11.14 aA | 75.36 ± 7.14 bB |
| Variables | Axis 1 | Axis 2 | Axis 3 | Axis 4 |
|---|---|---|---|---|
| Eigenvalues | 267.35 | 55.07 | 8.26 | 1.29 |
| Percentage | 80.53 | 16.59 | 2.49 | 0.39 |
| Cumulative Percentages | 80.53 | 97.12 | 99.60 | 99.99 |
| Lysozyme | 0.04 | −0.01 | 0.02 | −0.13 |
| Total Fagocytic Cells | 0.66 | −0.64 | −0.25 | −0.12 |
| SWC3_mon | 0.27 | 0.46 | 0.36 | 0.11 |
| SWC3_gran | 0.38 | 0.45 | −0.09 | −0.72 |
| Mean CD11b+CD18+ | 0.22 | −0.27 | 0.88 | −0.06 |
| Mean CD14+ | 0.55 | 0.34 | −0.16 | 0.55 |
| Mean CD14+CD16+ | 0.10 | −0.02 | 0.01 | 0.37 |
| Glutamate | 0.34 | 0.92 | −0.02 | 0.05 |
| Glutamine | 0.92 | −0.33 | −0.03 | −0.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarosz, Ł.S.; Tomaszewska, E.; Marek, A.; Hejdysz, M.; Burmańczuk, A.; Ciszewski, A.; Nowaczewski, S.; Grądzki, Z.; Batorski, M.; Świątkiewicz, M.; et al. The Effect of Glutamine as Feed Additive on Selected Parameters of the Nonspecific Immune Response in Pigs. Agriculture 2023, 13, 912. https://doi.org/10.3390/agriculture13040912
Jarosz ŁS, Tomaszewska E, Marek A, Hejdysz M, Burmańczuk A, Ciszewski A, Nowaczewski S, Grądzki Z, Batorski M, Świątkiewicz M, et al. The Effect of Glutamine as Feed Additive on Selected Parameters of the Nonspecific Immune Response in Pigs. Agriculture. 2023; 13(4):912. https://doi.org/10.3390/agriculture13040912
Chicago/Turabian StyleJarosz, Łukasz S., Ewa Tomaszewska, Agnieszka Marek, Marcin Hejdysz, Artur Burmańczuk, Artur Ciszewski, Sebastian Nowaczewski, Zbigniew Grądzki, Maciej Batorski, Małgorzata Świątkiewicz, and et al. 2023. "The Effect of Glutamine as Feed Additive on Selected Parameters of the Nonspecific Immune Response in Pigs" Agriculture 13, no. 4: 912. https://doi.org/10.3390/agriculture13040912
APA StyleJarosz, Ł. S., Tomaszewska, E., Marek, A., Hejdysz, M., Burmańczuk, A., Ciszewski, A., Nowaczewski, S., Grądzki, Z., Batorski, M., Świątkiewicz, M., & Rysiak, A. (2023). The Effect of Glutamine as Feed Additive on Selected Parameters of the Nonspecific Immune Response in Pigs. Agriculture, 13(4), 912. https://doi.org/10.3390/agriculture13040912