

Resistance of Sorghum Genotypes to Ergot (Claviceps Species)
by
 , ,
, ,
Dejene Kebede
1,2,3,*,
Isaac Onziga Dramadri
1,2,
Patrick Rubaihayo
1,
Thomas Odong
1 and
Richard Edema
1,2 1
Department of Agricultural Production, College of Agricultural and Environmental Sciences, Makerere University, Kampala P.O. Box 7062, Uganda
2
Makerere University Regional Centre for Crop Improvement, College of Agricultural and Environmental Sciences, Kampala P.O. Box 7062, Uganda
3
Ethiopian Institute of Agricultural Research, Addis Ababa P.O. Box 2003, Ethiopia
*
Author to whom correspondence should be addressed.
Agriculture 2023, 13(5), 1100; https://doi.org/10.3390/agriculture13051100
Submission received: 15 February 2023
/
Revised: 9 March 2023
/
Accepted: 19 May 2023
/
Published: 21 May 2023
(This article belongs to the Special Issue Crop Improvement through Conventional and Molecular Approaches)
Abstract
:Ergot is an important disease affecting sorghum when susceptible cultivars are grown and environmental conditions are favorable. Resistance breeding to ergot is, therefore, of paramount importance to provide effective, sustainable, and environmentally friendly management options. This study was conducted to assess the response of a global collection of sorghum genotypes to ergot. A total of 358 sorghum accessions were evaluated under field conditions. Data collection was conducted based on ergot incidence, severity, and agronomic traits, and the results revealed highly significant differences among genotypes (p < 0.01) for all traits evaluated. A mean disease incidence range of 23.8% to 69.4% was observed across test environments. Of the screened genotypes, 23, 52, and 2 genotypes showed resistant reactions during 2019, 2020, and 2021, respectively. Disease incidence and severity showed a moderate correlation with days to 50% flowering and pollen quantity, indicating their importance in ergot infection. Resistant genotypes E313, E111, E225, E200, E351, E352, E353, and E354 were identified from this study. These genotypes could be exploited as a resistance source in breeding for resistance to ergot.
1. Introduction
Sorghum is one of the five major cultivated cereal crops after wheat, maize, rice, and barley. Globally, it is grown on over 40.07 million hectares, producing 57.89 million tons of grains annually [1]. In the USA, Australia, Europe, and China, sorghum is used mainly as animal feed and forage and for industrial purposes, including ethanol production, whereas, in Africa, the Middle East, Asia, and Central America, it is a principal source of food for millions of people [2]. In Sub-Saharan Africa, 26 million tons of sorghum is produced annually, with over 100 million people depending on it as a source of food [3]. The major sorghum-producing countries in Africa include Nigeria, Sudan, Ethiopia, Burkina Faso, and Niger which account for more than 70% of Africa’s total production [4]. In Sub-Saharan Africa, sorghum is primarily produced by smallholder farmers and in arid to semi-arid environments with very little or no capital inputs, such as fertilizers, pesticides, or irrigation [5]. In Uganda, sorghum is adapted to a wide range of environments and is mainly produced in the southwestern highlands of Kabale and Ntungamo districts and the lowland areas of northern and eastern regions [6]. It is the third most important cereal crop grown after maize and rice [7]. It is used for making a wide variety of nutritious traditional foods such as semi-leavened bread, dumplings, and fermented and non-fermented porridge, and in the brewery industries for making beers for the local and export markets [6].
Sorghum production is constrained by a wide variety of biotic and abiotic stresses and socio-economic factors [8]. Leaf blight (Helminthosporium turcicum), anthracnose (Colletotrichum graminicolum), grain mold (Fusarium moniliforme), rust (Puccinia purpura), head smut (Sporisorium reilianum), downy mildew (Sclerospora sorghi), and ergot or sugary disease (Claviceps species) are among the various biotic stresses reducing sorghum productivity [9]. The incidence and severity of these diseases vary depending on the season, with the wet season recording higher disease incidence and severity compared to the dry season [10].
Ergot is a threat to sorghum production and causes significant yield loss worldwide [11]. According to Kazungu et al. (2023) [12], ergot is one of the most damaging sorghum diseases in Africa. In Zimbabwe and India, a yield loss of 20–80% had been reported in hybrid seed production due to ergot [13,14]. A report from southern Africa, Mexico, and the United States indicated that a yield loss of up to 80% and 50% was observed in hybrid sorghum seed and grain production, respectively [15,16]. Honeydew-contaminated grain has reduced feed intake and causes toxicity when fed to livestock [17]. Ergot-infected seeds exhibited lower germination and seedling emergence and could also increase the incidence and severity of several grain molds such as cerebella species, Curvularia species, Fusarium species, Alternaria species, and Cladosporium species [11].
Ergot is a serious problem, particularly in hybrid seed production that utilizes male-sterile parents [11]. However, when weather conditions are favorable for infection or when self-pollination is delayed or inhibited due to different environmental and genetic factors, all types of sorghum germplasm, fertile or male-sterile sorghum become susceptible to ergot [18]. Ergot infects unfertilized ovaries and competes with the pollen grain for germination in the stigma, and then colonizes the ovule [19]. According to Dahlberg et al. (2001) [20], certain characteristics of sorghum flowers, such as the small size of stigmas and short duration of stigma, receptivity, including particularly less aperture at glume tip, short exposure time of stigma before pollination, and rapid stigma drying after pollination, are important in reducing ergot infection [11].
Chemical control strategies using different fungicides can be used in combination with cultural practices or independently to reduce the incidence and severity of ergot [21]. However, in low-cash-value crops such as sorghum, disease management through chemical methods is often too expensive for farmers, and chemicals have a potential risk to human and animal health and are environmentally unsafe [2]. The use of resistant genotypes is a viable strategy for the efficient management and control of ergot disease in sorghum [21]. The objective of this study was to assess the response of a large number of sorghum genotypes and identify genotypes with resistance to ergot.
2. Materials and Methods
2.1. Experimental Site
The experiment was carried out at Makerere University Agricultural Research Institute Kabanyolo (MUARIK) during the first rain season (April–July) of 2019 and 2021 and the second rain season (September–December) of 2020 and at National Semi-Arid Resources Research Institute (NaSARRI) during the first rain season of 2019. MUARIK is located at 0°28′ N and 32°37′ E with an altitude of 1200 m above sea level and deep ferrallitic soils with a pH range of 5.2 to 6.0 [22]. The NaSARRI site is located at 01°32′ N, 033°27′ E with an altitude of 1140 m above sea level and is characterized by dry, warmer climatic conditions [23]. Weather data of NaSARRI and MUARIK for the study periods (2019–2021) are summarized in Supplementary Table S1.
2.2. Plant Materials
A total of 358 sorghum accessions, comprising 336 accessions from the sorghum association panel (SAP) and an additional 22 released genotypes, were evaluated for resistance to ergot. The male-sterile ATX623 was included as a susceptible check during 2020 evaluation at MUARIK but was not planted in other seasons due to shortage of seeds. The SAP included accessions from all major races of cultivated sorghum and geographic centers of diversity in sub-Saharan Africa and Asia, as well as from the United States [24].
2.3. Experimental Design and Field Layout
The experiment was planted using an alpha-lattice design with two replications. Each genotype was planted on a plot area consisting of two rows with a plot length of 5 m. A spacing of 1.5 m and 2 m between plots and replications were used, respectively. A spacing of 0.75 m between rows and 0.15 m within rows was used, and fields were weeded three times.
2.4. Data Collection and Analysis
In 2019, planting was carried out in April, and ergot resistance traits were collected in July. In 2020, planting was conducted on 31 August, and data on ergot resistance traits were collected in December for three consecutive weeks: 14 weeks after planting, 15 weeks after planting, and the final score was collected 16 weeks after planting. Likewise, in 2021, planting was carried out in April, and data on ergot infection was recorded in July for three consecutive weeks (14WAP, 15WAP, and 16WAP). Ergot severity (ES) was collected based on a 1–5 scale visual scoring as described by Musabyimana et al. (1995) [25] (Table 1).
Ergot incidence (EI) was estimated by counting all the individual plants with ergot disease symptoms in each plot and computed using the following formula.
Other parameters such as days to 50% flowering (DTF), pollen quantity (PQ), hundred seed weight (g) (HSW), seedling vigour (SV), plant height (PH), and panicle length (PL) were also collected. The HSW was measured by weighing a random sample of 100 seeds from each plot. Pollen quantity was measured following the procedures used by Parh et al. (2006) [26]. When 50% of plants had flowered in each plot, five heads per plot were flicked once and rated from 1 to 10 (1 for no visible pollen and 10 for a copious quantity of visible pollen) by observing the density of the resultant clouds of pollen. Plant height was measured from the base of the plant to the tip of the head from randomly selected five plants. Panicle length was measured from the base to the tip of the head. Seedling vigour was recorded on a 1–5 scale where 1 = healthy and vigorous and 5 = weak and diseased plants. Days to 50% flowering were recorded as the number of days from planting to when 50% of plants shaded the pollen [27].
Data collected on disease incidence and severity were transformed using Arcsine and Square root transformation methods, respectively [9], and analyzed using the R statistical package for Windows V-4.1.1. Mean separation was achieved using Least Significant Difference (LSD) at a 0.05 level of significance. Coefficient of variations, Standard error, and Pearson’s correlation analysis were carried out using Agricolae and Corplot package in R statistical package for Windows V-4.1.1 [28].
3. Results
3.1. Analysis of Variance
A separate analysis of variance was carried out across location and season trials. The analysis of variance across location trials revealed highly significant (p < 0.01) differences between locations for ergot incidence, severity, and DTF. The mean square due to genotype was highly significant (p < 0.01) for all traits, and the genotype by location interaction mean square was highly significant (p < 0.01) for all traits except DTF (Table 2).
The results of the analysis of variance for ergot incidence and severity at 14WAP, 15WAP, and 16WAP, HSW, PQ, DTF, PH, PL, and SV across seasons are presented in Table 3. The mean squares for the season were highly significant (p < 0.01) for ergot incidence at 14WAP and 16WAP, ergot severity at 14WAP, 15WAP, and 16WAP, HSW, DTF, PQ, PH, PL, and SV. Genotype mean square was highly significant (p < 0.01) for all recorded traits. The mean square due to genotype by seasons interaction also showed highly significant (p < 0.01) variation for all recorded traits except HSW (Table 3).
3.2. Response of Genotypes to Ergot Infection
The reaction of selected highly resistant, resistant, moderately resistant, susceptible, and highly susceptible genotypes evaluated for resistance to ergot at MUARIK and NaSARRI in the 2019, 2020, and 2021 seasons are presented in Table 4.
3.2.1. Disease Incidence
The responses of sorghum genotypes to ergot infection under natural infection conditions resulted in varying levels of resistance based on disease incidence and severity scores across seasons and locations. Genotypes showed varying levels of infections for ergot at MUARIK during the 2019, 2020, and 2021 seasons evaluations, whereas at NaSARRI, 19.7% (67 genotypes) of the genotypes had no ergot infection. Disease incidence varied between 8.5–100%, 6.0–100%, and 8.0–100% in 2019, 2020, and 2021, respectively, at MUARIK and from no disease to 100% at NaSARRI. Across environments, the mean ergot incidence was 59.6%, 69.4%, and 66.8% during 2019, 2020, and 2021, respectively, at MUARIK and 23.8% at NaSARRI.
Genotypes E351, E225, E133, E354, E033, E336, E355, and E352 showed lower disease incidence ranging between 8.0–23.5% across all environments (Table 4). The least disease incidence was recorded by genotypes E111 (8.5% incidence), E225 (9.0%), and E109 (9.0%) in 2019, E113 (6.0%) in 2020, and E351 (8.0%) in 2021. Out of the total screened genotypes, 38, 61, and 35 genotypes had 100% disease incidence at MUARIK in the 2019, 2020, and 2021 seasons, respectively. Conversely, only 10 genotypes showed 100% disease incidence at NaSARRI. Genotypes E248, E183, E094, E148, E325, E106, and E165 recorded 100% disease incidence consistently across the three seasons at MUARIK and manifested from 78.2–95.9% incidence at NaSARRI. The susceptible check ATX623 exhibited 100% disease incidence.
3.2.2. Disease Severity
Substantial variability was observed for disease severity scores among the evaluated genotypes. In 2019, the disease severity score at MUARIK varied between 1.5–5.0, with a mean severity score of 3.7. Genotypes E352, E353, E351, E011, E030, E013, E002, E228, E051, E319, E086, E354, E236, E133, E131, E141, E334, E328, E200, E313, E109, E225, and E111 showed resistant responses, with severity scores ranging from 1.5–2.0 (Table 4). The remaining 26.5%, 42.4%, and 24.1% of genotypes had moderately resistant, susceptible, and highly susceptible responses, respectively. However, no genotypes showed a highly resistant response (Figure 1).
A low disease severity score was observed at NaSARRI, ranging from 1.0–5.0, with a mean severity score of 1.9. Out of the total screened genotypes, 48.5% were rated as resistant, and 23.5% were rated as moderately resistant. The remaining 6.5% and 1.8% showed susceptible and highly susceptible responses, respectively. Overall, 19.7% of the total genotypes were not infected with ergot.
In 2020, the disease severity score varied from 1.0–5.0 at 14WAP and 1.5–5.0 at 15WAP and 16WAP. Out of the total evaluated genotypes, 52 genotypes had resistant responses (Figure 1). Genotypes E111, E297, E200, E032, E035, E066, E324, E268, E358, E313, E030, E034, E007, E226, E290, E279, E161, E036, E011, E351, E033, E273, E156, E30, E136, E181, E225, and E336 were among the resistant genotypes with disease severity scores ranging between 1.5–2.0. Furthermore, 49.8%, 28.3%, and 5.7% of the screened genotypes showed moderate resistant, susceptible, and highly susceptible reactions, respectively. The susceptible check ATX623 was among the highly susceptible genotypes, with a mean severity score of 5.0.
Ergot severity scores varied from 1.7–4.5 at 14WAP and from 1.8–5.0 at 15WAP and 16 WAP during the 2021 field evaluation. Only two genotypes, E351 and E354, had resistant responses to ergot infection with disease severity scores of 1.8 and 1.9, respectively. The other 54.8%, 33.8%, and 10.8% of evaluated genotypes showed moderately resistant, susceptible, and highly susceptible responses, respectively.
3.2.3. Disease Severity vs. PQ and DTF
The effect of pollen quantity on disease severity for selected resistant, moderately resistant, and susceptible genotypes is presented in Figure 2. In general, genotypes with good pollen quantity exhibited lower ergot infection. Genotypes with delayed flowering showed higher disease infection.
3.3. Variability for Agronomic Traits
The mean and ranges of all traits evaluated at MUARIK and NaSARRI during 2019, 2020, and 2021 are presented in Supplementary Table S2. The mean HSW varied between 1.7–4.7 g, 1.5–5.6 g, and 1.1–4.7 g in the 2019, 2020, and 2021 seasons, respectively. The highest mean HSW was recorded by genotypes E081 (4.7 g), E151 (4.5 g), and E186 (4.4 g) in 2019. In 2020, genotypes E186 (5.6 g), E196 (5.0 g), and E294 (4.9 g) recorded the highest HSW. Furthermore, E294 (4.6 g), E151 (4.7 g), and E128 (4.7 g) recorded the highest mean hundred seed weight in 2021. The mean PQ score ranged between 4.5–8.5, 5.3–8.8, and 5.5–8.0 in the 2019, 2020, and 2021 seasons, respectively, at MUARIK and 5.0–8.0 at NaSARRI. DTF varied from 59 to 102 days, with an average of 78 days across all environments. Plant height varied between 62.1–284 cm in 2020 and 38.0–245 cm in 2021, while PL ranged between 9.5–55.9 cm in 2020 and 8.7–54.3 cm in 2021 at MUARIK. Moreover, the SV score varied between 1.0–5.0, with an average performance of 2.8 across seasons.
3.4. Correlation between Resistance to Ergot and Agronomic Traits
Correlation analyses between ergot disease incidence, severity, and agronomic traits were significant for most traits (Table 5). A highly significant correlation was observed between ergot incidence and severity (r = 0.6, p < 0.001). Ergot incidence showed a significant positive correlation with days to 50% flowering (r = 0.24, p < 0.01) and seedling vigour (r = 0.27, p < 0.01) and a significant negative correlation with pollen quantity (r = −0.16, p < 0.05) and plant height (r = −0.32, p < 0.01), respectively. Ergot severity showed a significant negative correlation with seedling vigour (r = −0.24, p < 0.01), pollen quantity (r = −0.18, p < 0.05), and plant height (r = −0.28, p < 0.01) (Table 5).
4. Discussion
Substantial variability was observed among genotypes, locations, and seasons for ergot incidence, severity, and agronomic traits. There were highly significant differences among genotypes indicating the existence of considerable variations in their responses to ergot infection that could be attributed to genetic differences among the genotypes. Parh et al. (2006) [26] observed moderate genetic variability for resistance to ergot among a population of recombinant inbred lines and their parents. Variation among full-sib families for resistance to ergot was also observed by Mirdita et al. (2008) [29]. The results showed sorghum genotypes, including E313, E111, E225, E200, E351, E352, E353, and E354, with relatively lower ergot incidence and severity consistently across environments, suggesting that these genotypes possessed resistant genes and could be useful in breeding for resistance to ergot. Bogo et al. (2004) [15] screened sweet sorghum genotypes for resistance to ergot in Brazil and reported that out of the 47 accessions, 5 never showed any signs of infection and exhibited a very high degree of seed set.
The observed significant variations between locations and seasons for ergot incidence and severity indicated that the evaluation of sorghum genotypes for resistance to ergot should be conducted in more than one environment if carried out under natural infestation conditions. The prevalence of ergot infection was high at MUARIK during all three seasons indicating that MUARIK can be considered a hotspot for ergot and therefore affirming natural infestation conditions in this location and, therefore, could be used for screening of sorghum genotypes for resistance to ergot. The successful ergot infection observed at MUARIK could be due to the ready availability of inoculums (conidia) from ergot-infected male-sterile and wild sorghum genotypes and grasses in the surrounding fields. According to Miedaner and Geiger (2015) [11], ergot infection under natural infestation was mainly caused by secondary inoculums or conidia from ergot-infected wild grasses and sorghums. The higher relative humidity and relatively lower minimum temperature that occurred during flowering at MUARIK assisted in successful ergot infection. The importance of humid, rainy, and cool weather conditions during flowering for ergot infection was reported by Cisneros-lópez1 et al. (2010) [30]. The low ergot infection level observed at NaSARRI was due to unfavorable environmental conditions that hinder ergot infections. A similar result was observed by Kodisch et al. (2020) [31], who reported a low ergot infection rate in Poland due to dry and warm environmental conditions.
A highly significant negative correlation observed between ergot incidence and severity with pollen quantity attested the importance of pollen trait in ergot infection, and the higher pollen quantity score observed in the resistant genotypes suggested that high pollen availability played a significant role in reducing ergot infection through competition with ergot to germinate on stigma. This was in tandem with Parh et al. (2006) [26], who reported a linear relationship between successful pollen production and resistance to ergot in some sorghum genotypes. Ergot severity and incidence showed a significant positive correlation with days to 50% flowering, with late flowering genotypes showing higher severity which was attributed to the amount of the pathogen inoculum increasing at a later stage and poor pollen viability in late flowering genotypes due to higher humidity and moisture. Late-maturing sorghum genotypes were reported to have higher ergot severity [21].
5. Conclusions
Significant variations were observed among genotypes in response to ergot disease infection and agronomic traits. Among the evaluated genotypes, E313, E111, E225, E200, E351, E352, E353, and E354 were found to be resistant to ergot disease infection. These genotypes could be utilized as a source of resistance in a sorghum program to breed for resistance to ergot.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture13051100/s1, Table S1: Rain fall, relative humidity, minimum and maximum temperature of MUARIK and NaSARRI in 2019, 2020 and 2021; Table S2: Mean and ranges of traits evaluated at MUARIK and NaSARRI during 2019, 2020 and 2021.
Author Contributions
Conceptualization, D.K. and I.O.D.; methodology, D.K. and I.O.D.; software, D.K. and I.O.D.; validation, D.K., I.O.D. and P.R.; formal analysis, D.K.; investigation, D.K.; resources, I.O.D. and R.E.; data curation, D.K. and I.O.D.; writing—original draft preparation, D.K.; writing—review and editing, D.K., I.O.D., P.R., T.O. and R.E.; visualization, D.K., I.O.D. and P.R.; supervision, D.K.; project administration, I.O.D. and R.E.; funding acquisition, T.O. and R.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank SCIFSA and MaRCCI staff for their unrestricted support during the study period.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Food and Agriculture Organization of the United Nations. Statistics Division, Rome, Italy. Available online: https://www.fao.org/faostat/en/ (accessed on 20 May 2022).
- Caroline, O.; Evans, O.; Samuel, G. Screening of Selected Sorghum Genotypes for Resistance to Covered Kernel Smut Disease in Western Kenya. J. Agric. Sci. 2021, 13, 63–73. [Google Scholar] [CrossRef]
- Mofokeng, M.; Shimelis, H.; Laing, M.; Shargie, N. Sorghum [Sorghum bicolor (L.) Moench] breeding for resistance to leaf and stalk anthracnose, Colletotrichum sublineolum, and improved yield: Progress and prospects. Aust. J. Crop Sci. 2017, 11, 1078–1085. [Google Scholar] [CrossRef]
- Gebrekorkos, G.; Egziabher, Y.G.; Habtu, S. Response of Sorghum (Sorghum bicolor (L.) Moench) Varieties to Blended Fertilizer on yield, yield component and nutritional content under Irrigation in Raya Valley, Northern Ethiopia. Int. J. Agric. Biosci. 2019, 6, 153–162. Available online: www.ijagbio.com (accessed on 1 November 2019).
- Beyene, A.; Hussien, S.; Pangirayi, T.; Mark, L. Physiological mechanisms of drought tolerance in sorghum, genetic basis, and breeding methods: A review. Afr. J. Agric. Res. 2015, 10, 3029–3040. [Google Scholar] [CrossRef]
- Tenywa, M.M.; Nyamwaro, S.O.; Kalibwani, R.; Mogabo, J.; Buruchara, R.; Oluwole, F. Innovation Opportunities in Sorghum Production in Uganda. FARA Res. Rep. 2018, 2, 20. [Google Scholar]
- Njoroge, S.M.; Takan, J.P.; Letayo, E.A.; Okoth, P.S.; Ajaku, D.O.; Kumar, A.; Rathore, A.; Ojulong, H.; Manyasa, E. Survey of Fungal Foliar and Panicle Diseases of Sorghum in Important Survey of Fungal Foliar and Panicle Diseases of Sorghum in Important Agroecological Zones of Tanzania and Uganda. Plant Health Prog. 2018, 19, 265–271. [Google Scholar] [CrossRef]
- Amelework, B.A.; Shimelis, H.A.; Tongoona, P.; Laing, M.D.; Ayele, D.G. Sorghum production systems and constraints, and coping strategies under drought-prone agro-ecologies of Ethiopia. S. Afr. J. Plant Soil 2016, 33, 207–217. [Google Scholar] [CrossRef]
- Muturi, P.W.; Mgonja, M.; Rubaihayo, P. Identification of new sorghum genotypes resistant to the African and spotted stemborers. Int. J. Trop. Insect Sci. 2014, 34, 260–268. [Google Scholar] [CrossRef]
- Thakur, R.P.; Reddy, B.V.; Mathur, K. Screening Techniques for Sorghum Diseases. Information Bulletin No. 76; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2007; pp. 1–92. ISBN 978-92-9066-504-5. [Google Scholar]
- Miedaner, T.; Geiger, H.H. Biology, Genetics, and Management of Ergot (Claviceps spp.) in Rye, Sorghum, and Pearl Millet. Toxins 2015, 7, 659–678. [Google Scholar] [CrossRef]
- Kazungu, F.K.; Mundi, E.M.; Mulinge, J.M. Overview of Sorghum (Sorghum bicolor L.), its Economic Importance, Ecological Requirements and Production Constraints in Kenya. Int. J. Plant Soil Sci. 2023, 35, 62–71. [Google Scholar] [CrossRef]
- Frederickson, D.E.; Mantle, P.G.; DE Milliano, W.A.J. Susceptibility to ergot in Zimbabwe of sorghums that remained unaffected in their native climates in Ethiopia and Rwanda. Plant Pathol. 1994, 43, 27–32. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Frederickson, D.E.; Mclaren, N.W.; Odvody, G.N.; Ryley, M.J. Ergot: A New Disease Threat to Sorghum in the Americas and Australia. Plant Dis. 1998, 82, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Bogo, A.; Mantle, P.G.; Harthmann, O.E.L. Screening of sweet sorghum accessions for inhibition of secondary sporulation and saccharide measurements in honeydew of Claviceps africana. Fitopatol. Bras. 2004, 29, 86–90. [Google Scholar] [CrossRef]
- Prom, L.K.; Erpelding, J.E.; Montes-Garcia, N. Evaluation of Sorghum Germplasm from China against Claviceps africana, Causal Agent of Sorghum Ergot. Plant Health Prog. 2008, 9, 1–7. [Google Scholar] [CrossRef]
- Parh, D.K.; Jordan, D.R.; Aitken, E.A.B.; Mace, E.S.; Jun-ai, P.; McIntyre, C.L.; Godwin, I.D. QTL analysis of ergot resistance in sorghum. Theor. Appl. Genet. 2008, 117, 369–382. [Google Scholar] [CrossRef]
- Shivakumar, B.Y. Studies on Sorghum Ergot Caused by Claviceps africana Frederickson Mantle and de Milliano. Master’s Thesis, University of Agricultural Sciences, Dharwad, India, 2011. [Google Scholar]
- Mendoza-Onofre, L.E.; Hernández-Martínez, M.; Cárdenas-Soriano, E.; Ramírez-Vallejo, P. El germoplasma de sorgo tolerante al frío como fuente potencial de tolerancia al ergot (Claviceps africana Frederickson, Mantle & de Milliano). Agrociencia 2006, 40, 593–603. [Google Scholar]
- Dahlberg, J.A.; Bandyopadhyay, R.; Rooney, W.L.; Odvody, G.N.; Madera-Torres, P. Evaluation of sorghum germplasm used in US breeding programmes for sources of sugary disease resistance. Plant Pathol. 2001, 50, 681–689. [Google Scholar] [CrossRef]
- Sorghum Diseases Sorghum Ergot (Claviceps africana). Fusarium Stalk Rot (Fusarium spp.). Charcoal Rot (Macrophomina phaseolina). Rust (Puccinia purpurea). Johnson Grass Mosaic Virus|Head Smut (Sporisorium reilianum). Leaf Blight (Exserohilum turcic). Australia. Available online: https://grdc.com.au/about/who-we-are/corporate-governance/annual-reports (accessed on 22 December 2022).
- Kebede, D.; Rubaihayo, P.; Dramadri, I.; Odong, T.; Edema, R. Mechanisms associated with resistance to ergot in sorghum. Res. Agric. Vet. Sci. 2022, 6, 127–140. [Google Scholar]
- Beshir, M.M. Genetic Analysis of Dual Resistance to Anthracnose and Turcicum Leaf Blight in Sorghum. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2016. [Google Scholar]
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef]
- Musabyimana, T.; Sehene, C.; Bandyopadhyay, R. Ergot resistance in Sorghum in relation to flowering, inoculation technique and disease development. Plant Pathol. 1995, 44, 109–115. [Google Scholar] [CrossRef]
- Parh, D.K.; Jordan, D.R.; Aitken, E.A.; Gogel, B.J.; McIntyre, C.L.; Godwin, I.D. Genetic components of variance and the role of pollen traits in Sorghum ergot resistance. Crop Sci. 2006, 46, 2387–2395. [Google Scholar] [CrossRef]
- Awori, E.; Kiryowa, M.; Basirika, A.; Dradiku, F.; Kahunza, R.; Oriba, A.; Mukalazi, J. Performance of elite grain sorghum varieties in the West Nile Agro-ecological Zones. Uganda J. Agric. Sci. 2015, 16, 139–148. [Google Scholar] [CrossRef]
- RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA; Available online: http://www.rstudio.com/ (accessed on 3 December 2022).
- Mirdita, V.; Dhillon, B.S.; Geiger, H.H.; Miedaner, T. Genetic variation for resistance to ergot (Claviceps purpurea [Fr.] Tul.) among full-sib families of five populations of winter rye (Secale cereale L.). Theor. Appl. Genet. 2008, 118, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-lópez, M.E.; Mendoza-onofre, L.E.; González-hernández, V.A.; Mora-aguilera, G.; Hernández-Martínez, M.; Zavaleta-Mancera, H.A.; Córdova-Téllez, A.L. Floral traits, ergot resistance and grain yield relationships in infected male-sterile sorghum A-lines. Seed Sci. Technol. 2010, 38, 114–124. [Google Scholar] [CrossRef]
- Kodisch, A.; Wilde, P.; Schmiedchen, B.; Fromme, F.J.; Rodemann, B.; Tratwal, A.; Oberforster, M.; Wieser, F.; Schiemann, A.; Jørgensen, L.N.; et al. Ergot infection in winter rye hybrids shows differential contribution of male and female genotypes and environment. Euphytica 2020, 216, 65. [Google Scholar] [CrossRef]
Figure 1.
Summary of highly resistant, resistant, moderately resistant, susceptible, and highly susceptible genotypes in different environments. HR—highly resistant, R—resistant, MR—moderately resistant, S—susceptible, and HS—highly susceptible.
Figure 1.
Summary of highly resistant, resistant, moderately resistant, susceptible, and highly susceptible genotypes in different environments. HR—highly resistant, R—resistant, MR—moderately resistant, S—susceptible, and HS—highly susceptible.
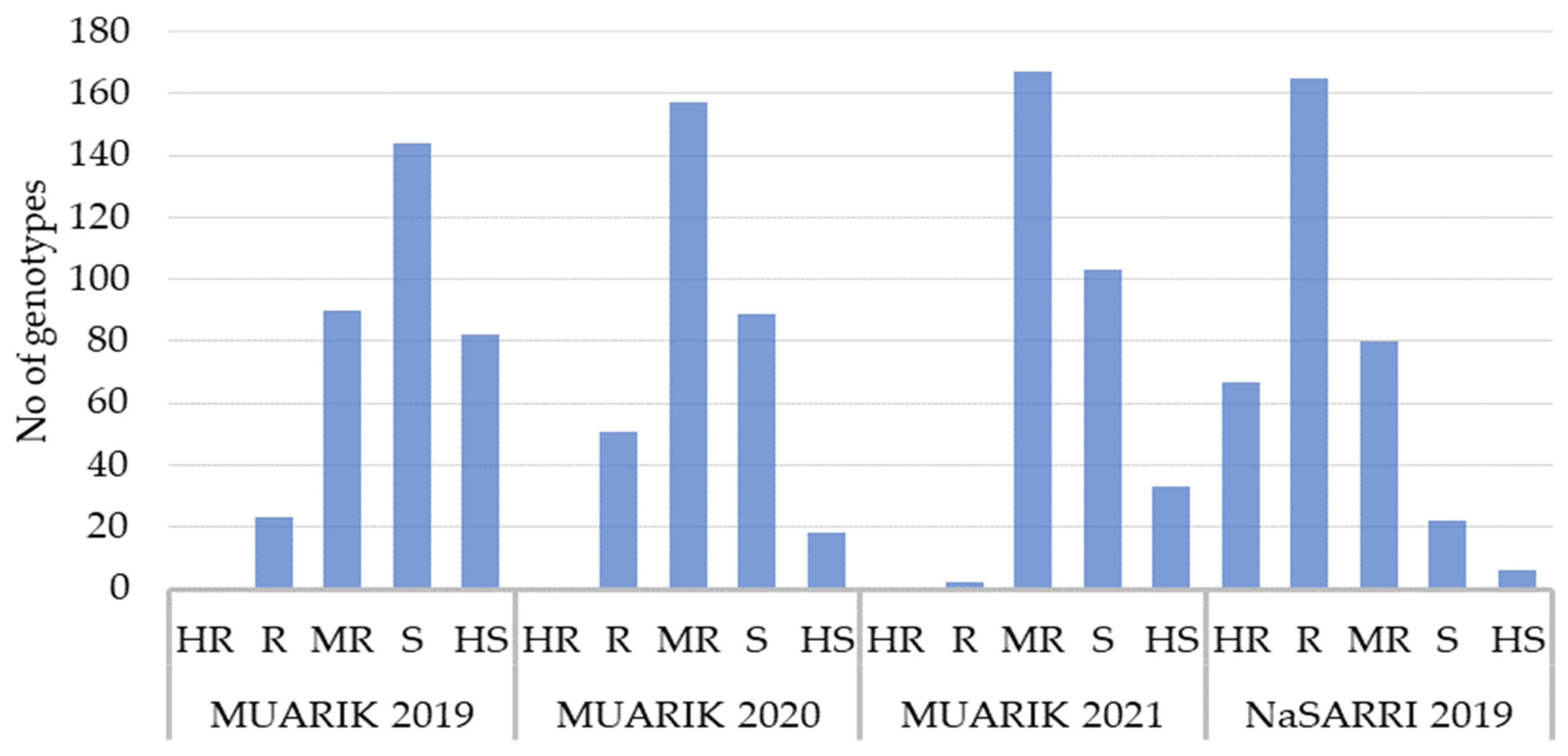
Figure 2.
Ergot severity and PQ for selected resistant, moderately resistant, and susceptible genotypes in different locations and seasons; (a), MUARIK 2019 (b), MUARIK 2020 (c), MUARIK 2021 and (d), NaSARRI 2019.
Figure 2.
Ergot severity and PQ for selected resistant, moderately resistant, and susceptible genotypes in different locations and seasons; (a), MUARIK 2019 (b), MUARIK 2020 (c), MUARIK 2021 and (d), NaSARRI 2019.


{kind=link}
{kind=link}
Table 1.
Ergot severity score and disease reactions.
| Score | Infection (%) | Disease Reaction |
|---|---|---|
| 1 | No infection | Highly resistant (HR) |
| 2 | 1–10% infection | Resistant (R) |
| 3 | 11–25% infection | Moderately resistant (MR) |
| 4 | 26–50% infection | Susceptible (S) |
| 5 | >50% infection | Highly susceptible (HS) |
Source: [25].
Table 2.
Analysis of variance for ergot incidence, severity, and agronomic traits evaluated at MUARIK and NaSARRI in 2019.
Table 2.
Analysis of variance for ergot incidence, severity, and agronomic traits evaluated at MUARIK and NaSARRI in 2019.
| SOV | DF | Incidence (%) | Severity (1–5) | DTF (Days) | Pollen Quantity (1–10) | Seedling Vigour (1–5) |
|---|---|---|---|---|---|---|
| Location | 1 | 290,823 ** | 110 ** | 3438 ** | 0.3 ns | 0.06 ns |
| Rep (Rep × Loc) | 2 | 7095 ** | 0.53 ** | 240 * | 4.5 ** | 2.6 ** |
| Genotype | 339 | 1059 ** | 0.17 ** | 225 ** | 0.7 ** | 0.9 ** |
| Genotype × Loc | 339 | 592 ** | 0.09 ** | 28.3 ns | 0.8 ** | 0.5 ** |
| Error | 664 | 166 | 0.02 | 24.8 | 0.2 | 0.4 |
*, ** = Significant at p < 0.05 and p < 0.01, respectively. SOV—source of variations, DF—degree of freedom, DTF—days to 50% flowering and ns—non-significant.
Table 3.
Mean squares for ergot incidence, severity, and agronomic traits evaluated during 2019, 2020, and 2021 seasons at MUARIK.
Table 3.
Mean squares for ergot incidence, severity, and agronomic traits evaluated during 2019, 2020, and 2021 seasons at MUARIK.
| SOV | DF | Incidence (%) | Severity (1–5) | HSW (g) | DTF (Days) | PQ (1–10) | PH (cm) | PL (cm) | SV (1–5) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 14WAP | 15WAP | 16WAP | 14WAP | 15WAP | 16WAP | ||||||||
| Season | 2 | 48,869 ** | 121 ns | 8049 ** | 26.7 ** | 32 ** | 8.4 ** | 1.66 ** | 34,828 ** | 10.6 ** | 50,662 ** | 96 ** | 615 ** |
| Rep (Rep × Season) | 3 | 3037 ** | 4261 ** | 2517 ** | 0.17 ** | 0.4 ns | 0.18 ** | 0.14 ns | 454 ** | 5.1 ** | 535 ns | 29.5 * | 1.26 ** |
| Genotype | 356 | 549 ** | 779 ** | 1115 ** | 0.09 ** | 1.1 ** | 0.15 ** | 1.15 ** | 187 ** | 0.78 ** | 4316 ** | 105 ** | 0.78 ** |
| Genotype × Season | 607 | 261 ** | 316 ** | 415 ** | 0.03 ** | 0.35 ** | 0.05 ** | 0.12 ns | 71 ** | 0.55 ** | 435 ** | 14.5 ** | 0.48 ** |
| Error | 841 | 198 | 206 | 173 | 0.02 | 0.16 | 0.014 | 0.14 | 26 | 0.19 | 193 | 7.8 | 0.32 |
*, ** = Significant at p < 0.05 and p < 0.01, respectively. SOV—source of variations, DF—degree of freedom, WAP—weeks after planting, HSW—hundred seed weight, DTF—days to 50% flowering, PQ—pollen quantity, PH—plant height, PL—panicle length SV—seedling vigour and ns—non-significant.
Table 4.
Reaction of selected genotypes to ergot infection at MUARIK and NaSARRI evaluated during 2019, 2020, and 2021 seasons.
Table 4.
Reaction of selected genotypes to ergot infection at MUARIK and NaSARRI evaluated during 2019, 2020, and 2021 seasons.
| Genotype | MUARIK | NaSARRI | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | 2021 | 2019 | |||||||||
| Incidence (%) | Severity (1–5) | RC | Incidence (%) | Severity (1–5) | RC | Incidence (%) | Severity (1–5) | RC | Incidence (%) | Severity (1–5) | RC | |
| E313 | 10.0 | 2.0 | R | 54.0 | 1.6 | R | 32.0 | 2.2 | MR | 4.4 | 1.3 | R |
| E351 | 14.0 | 1.5 | R | 14.5 | 1.8 | R | 8.0 | 1.8 | R | 0 | 1.0 | HR |
| E200 | 10.5 | 2.0 | R | 44.5 | 1.5 | R | 40.0 | 2.4 | MR | 6.0 | 1.3 | R |
| E352 | 38.5 | 1.5 | R | 22.5 | 2.0 | R | 15.5 | 2.1 | MR | 4.6 | 1.3 | R |
| E354 | 16.0 | 2.0 | R | 46.0 | 2.4 | MR | 14.5 | 1.9 | R | 6.8 | 1.8 | R |
| E138 | 28.0 | 3.0 | MR | 31.0 | 2.1 | MR | 36.0 | 2.6 | MR | 33.4 | 3.0 | MR |
| E187 | 58.0 | 2.5 | MR | 67.0 | 2.1 | MR | 54.0 | 2.8 | MR | 0 | 1.0 | HR |
| E230 | 19.0 | 3.0 | MR | 37.0 | 1.9 | MR | 67.5 | 3.0 | MR | 17.7 | 2.5 | MR |
| E304 | 41.0 | 3.0 | MR | 34.3 | 2.3 | MR | 55.5 | 2.9 | MR | 6.3 | 1.5 | MR |
| E017 | 32.0 | 3.0 | MR | 39.5 | 2.8 | MR | 46.5 | 2.9 | MR | 33.4 | 2.3 | MR |
| E163 | 54.3 | 4.0 | S | 100 | 3.8 | S | 63.5 | 3.8 | S | 76.0 | 4.0 | S |
| E213 | 82.0 | 4.0 | S | 100 | 4.5 | HS | 80 | 3.5 | S | 64.7 | 4.0 | S |
| E147 | 100 | 5.0 | HS | 75.0 | 3.5 | S | 66.5 | 3.2 | S | 100 | 4.5 | HS |
| E169 | 100 | 5.0 | HS | 71.5 | 4.0 | S | 58.5 | 3.1 | S | 100 | 5.0 | HS |
| CV | 24.0 | 10.8 | 24.8 | 16.5 | 23.1 | 12.3 | 57.0 | 22 | ||||
| LSD | 29.3 | 0.79 | 36.9 | 1.01 | 32.7 | 0.73 | 32.0 | 0.84 | ||||
| SE | 1.57 | 0.05 | 1.52 | 0.05 | 1.32 | 0.04 | 1.9 | 0.05 | ||||
RC—Reaction class, LSD—least significance difference, CV—coefficient of variation, SE—standard error, HR—highly resistant, R—resistant, MR—moderately resistant, S—susceptible, and HS—highly susceptible.
Table 5.
Pearson’s correlation between ergot incidence, severity, and agronomic traits combined across location and years.
Table 5.
Pearson’s correlation between ergot incidence, severity, and agronomic traits combined across location and years.
| Trait | Days to 50% Flowering (Days) | Severity (1–5) | Incidence (%) | Hundred Seed Weight (g) | Plant Height (cm) | Panicle Length (cm) | Seedling Vigour (1–5) |
|---|---|---|---|---|---|---|---|
| Severity (1–5) | 0.11 * | ||||||
| Incidence (%) | 0.24 ** | 0.6 ** | |||||
| Hundred seed weight (g) | −0.21 ** | −0.1 * | 0.01 ns | ||||
| Plant height (cm) | −0.11 ns | −0.28 ** | −0.32 ** | 0.09 ns | |||
| Panicle length (cm) | 0.17 ** | −0.05 ns | −0.07 ns | −0.11 ns | 0.39 ** | ||
| Seedling vigour (1–5) | 0.14 * | −0.24 ** | 0.27 ** | 0.04 ns | −0.04 ns | −0.03 ns | |
| Pollen quantity (1–10) | 0.02 ns | −0.18 ** | −0.16 ** | −0.08 ns | −0.12 * | −0.02 ns | 0.01 ns |
*—significant at p < 0.05, **—significant at p < 0.01 and ns—non-significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kebede, D.; Dramadri, I.O.; Rubaihayo, P.; Odong, T.; Edema, R. Resistance of Sorghum Genotypes to Ergot (Claviceps Species). Agriculture 2023, 13, 1100. https://doi.org/10.3390/agriculture13051100
AMA Style
Kebede D, Dramadri IO, Rubaihayo P, Odong T, Edema R. Resistance of Sorghum Genotypes to Ergot (Claviceps Species). Agriculture. 2023; 13(5):1100. https://doi.org/10.3390/agriculture13051100
Chicago/Turabian StyleKebede, Dejene, Isaac Onziga Dramadri, Patrick Rubaihayo, Thomas Odong, and Richard Edema. 2023. "Resistance of Sorghum Genotypes to Ergot (Claviceps Species)" Agriculture 13, no. 5: 1100. https://doi.org/10.3390/agriculture13051100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.