
Plant Height and Stem Diameter of Solanum quitoense Lamarck Improved with Applications of AMF and Biostimulants
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Biological Material Used
2.3. Physicochemical Analysis of Soil
2.4. Isolation of Mycorrhizal Fungal Spores
2.5. Treatments and Inoculation of Symbiotic Microorganisms
2.6. Variables
2.6.1. Plant Growth
2.6.2. Leaf Area Growth
2.6.3. Mycorrhizal Colonization in the Roots
2.6.4. Foliar Analysis
2.7. Experimental Design and Statistical Analysis
3. Results
3.1. Soil Analysis
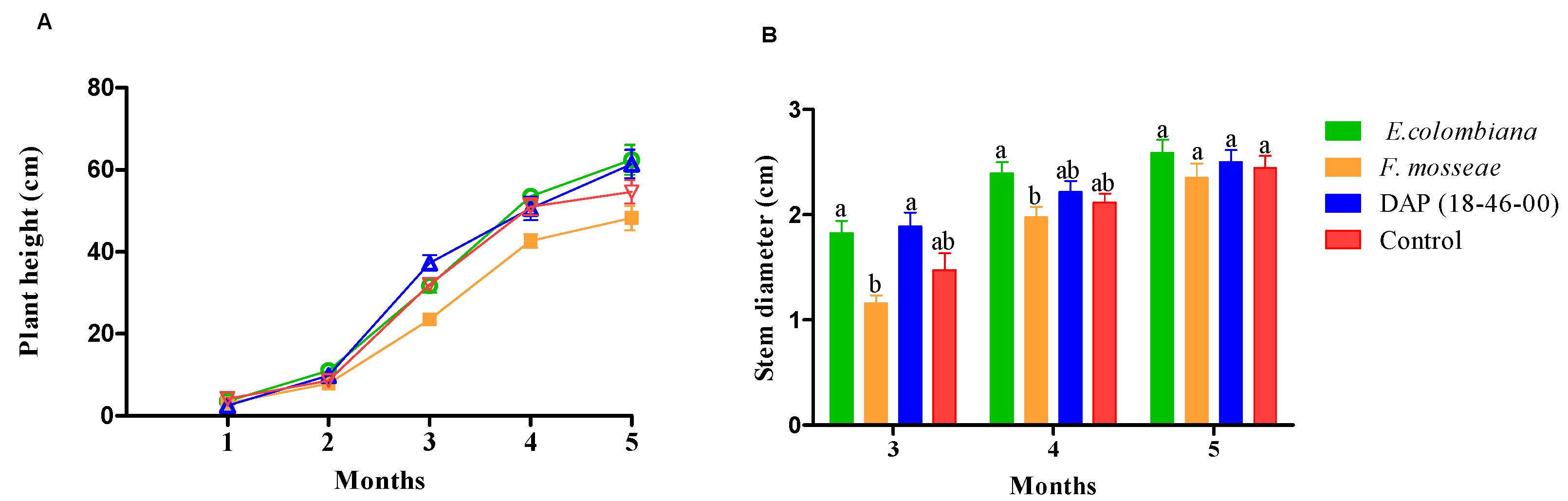
3.2. Plant Height and Stem Diameter
3.3. Number of Leaves and the Leaf Area
3.4. Mycorrhizal Colonization
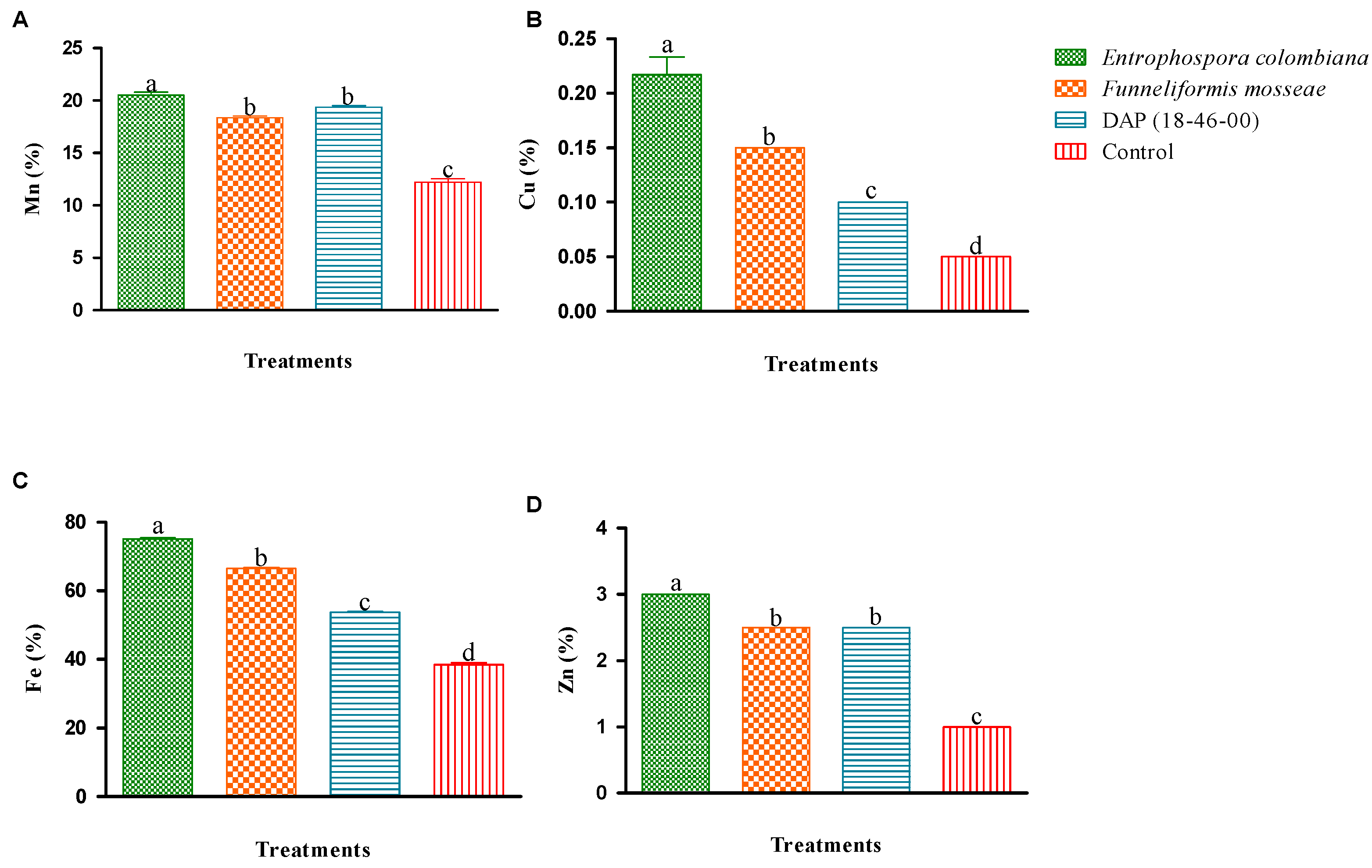
3.5. Foliar Analysis
4. Discussion
4.1. Soil Analysis
4.2. Plant Height and Stem Diameter
4.3. Number of Leaves and the Leaf Area
4.4. Mycorrhizal Colonization
4.5. Foliar Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.R.; Morales, J.A.R.; de la Torre, D.D.J.P.; Pérez, M.A.F.; Morales, P.M.; García, A.M.; Medina, J.F.A. Diversidad y estructura arbórea de los árboles de sombra asociados a Coffea arabica L. en el Soconusco, Chiapas. Rev. Mex. Cienc. For. 2022, 13, 4–27. [Google Scholar] [CrossRef]
- Baldi, E. Soil-plant interaction: Effects on plant growth and soil biodiversity. Agronomy 2021, 11, 2378. [Google Scholar] [CrossRef]
- Bhale, S.A.; Bansode, A.; Singh, S. Multifactorial Role of Arbuscular Mycorrhizae in Agroecosystem. In Fungi and their Role in Sustainable Development: Current Perspectives; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 2005–2020. [Google Scholar] [CrossRef]
- Devi, S.H.; Bhupenchandra, I.; Sinyorita, S.; Chongtham, S.; Devi, E.L. Mycorrhizal fungi and sustainable agriculture. In Nitrogen in Agriculture−Physiological, Agricultural and Ecological Aspects; Ohyama, T., Inubushi, K., Eds.; IntechOpen: London, UK, 2021; pp. 1–19. [Google Scholar] [CrossRef]
- Agnolucci, M.L.; Avio, A.; Pepe, A.; Turrini, C.; Cristani, P.; Bonini, P.; Giovannetti, M. Bacteria associated with a commercial mycorrhizal inoculum: Community composition and multifunctional activity as assessed by illumina sequencing and culture-dependent tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef] [PubMed]
- Lara-Capistran, L.; Zulueta-Rodriguez, R.; Murillo-Amador, B.; Preciado-Rangel, P.; Verdecia-Acosta, D.M.; Hernandez-Montiel, L.G. Biodiversity of AM fungi in Coffee cultivated on eroded soil. Agronomy 2021, 11, 567. [Google Scholar] [CrossRef]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Abarenkov, K. Regional-scale in-depth analysis of soil fungal diversity reveals strong pH and plant species effects in Northern Europe. Front. Microbiol. 2020, 11, 1953. [Google Scholar] [CrossRef]
- Janowski, D.; Leski, T. Factors in the distribution of mycorrhizal and soil fungi. Diversity 2022, 14, 1122. [Google Scholar] [CrossRef]
- El-Sawah, A.M.; El-Keblawy, A.; Ali, D.F.I.; Ibrahim, H.M.; El-Sheikh, M.A.; Sharma, A.; Alhaj Hamoud, Y.; Shaghaleh, H.; Brestic, M.; Skalicky, M.; et al. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- Duarte-Alvarado, D.; Lagos-Burbano, T.C.; Lagos-Santander, L.K.; Benavides-Cardona, C.A. General and specific combining abilities in a diallel cross of lulo (Solanum quitoense Lam.). Rev. Colomb. Cienc. Hortícolas 2021, 15, e11976. [Google Scholar] [CrossRef]
- Andrade-Cuvi, M.J.; Guijarro-Fuertes Figueroa, J.L. Evaluación fisicoquímica y antioxidante de naranjilla (Solanum quitoense Lam.) durante la maduración. Rev. Iberoam. Tecnol. Postcosecha 2021, 22, 145–164. [Google Scholar]
- Ramírez, F.; Kallarackal, J.; Davenport, T.L. Lulo (Solanum quitoense Lam.) reproductive physiology: A review. Sci. Hortic. 2018, 238, 163–176. [Google Scholar] [CrossRef]
- Daunay, M.C.; Rousselle-Bourgeois, F.; Lester, R.N.; Peron, J.Y. Known and less known Solanum species for fresh market. Int. Symp. Solanacea Fresh Mark. 1995, 412, 293–305. [Google Scholar] [CrossRef]
- Chiarini, F.E.; Barboza, G.E. Fruit anatomy of species of Solanum sect. Acanthophora (Solanaceae). In Flora-Morphology, Distribution, Functional Ecology of Plants; Elsevier: Amsterdam, The Netherlands, 2009; Volume 204, pp. 146–156. [Google Scholar] [CrossRef]
- González-Espinosa, M.; Meave, J.A.; Ramírez-Marcial, N.; Toledo-Aceves, T.; Lorea-Hernández, F.G.; Ibarra-Manríquez, G. Los bosques de niebla de México: Conservación y restauración de su componente arbóreo. Ecosistemas 2012, 21, 36–54. [Google Scholar]
- Muñoz, J.A.; Rodríguez, L.F.; Bermúdez, L.T. Análisis de competitividad del sistema de producción de lulo (Solanum quitoense Lam.) en tres municipios de Nariño. Rev. Colomb. Cienc. Hortícolas 2013, 7, 173–185. [Google Scholar]
- Lim, T.K. Solanum quitoense. In Edible Medicinal and Non-Medicinal Plants; Springer: Dordrecht, The Netherlands, 2013; Volume 6, pp. 419–423. [Google Scholar] [CrossRef]
- Varela, J.C.; Velásquez, J.C.; de Tafur, M.S.M. Respuesta fisiológica del lulo (Solanum quitoense Lam.) a diferentes concentraciones de N, P, K, Ca y Mg en la solución nutritiva. Acta Agron. 2002, 51, 53–59. [Google Scholar] [CrossRef]
- Di Barbaro, G.; Andrada, H.; González Basso, V.; Alurralde, A.L.; Del Valle, E.; Brandán de Weht, C. Micorrizas arbusculares y hongos septados oscuros nativos en topinambur (Helianthus tuberosus L.) en Catamarca, Argentina. Rev. Cienc. Agric. 2017, 34, 98–106. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the ideotype mycorrhizal symbionts for the production of healthy food. Front. Plant Sci. 2018, 9, 1089. [Google Scholar] [CrossRef]
- Arocha-Rodríguez, M.D.L.C.; Pérez-Ortega, E.; Fernández-Suárez, K.; Haesaert, G. Efecto del pH del medio de cultivo en el crecimiento pre simbiótico de Rhizoglomus irregulare. Cultiv. Trop. 2019, 40, 1–12. [Google Scholar]
- Ferlian, O.; Biere, A.; Bonfante, P.; Buscot, F.; Eisenhauer, N.; Fernandez, I.; Martinez-Medina, A. Growing research networks on mycorrhizae for mutual benefits. Trends Plant Sci. 2018, 23, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loor, M.J.B.; Zambrano, L.J.S. Evaluación de la infectividad de comunidades de hongos micorrízicos arbusculares. Pro Sci. Rev. Prod. Cienc. Investig. 2019, 3, 1–5. [Google Scholar]
- Peñuelas-Rubio, O.; Trejo-Aguilar, D.; Cervantes-Gámez, R.G.; Maldonado-Mendoza, I.E. Análisis microbiómico de hongos micorrízicos arbusculares asociados a papaya, inoculada con consorcios de dos ecosistemas naturales. Sci. Fungorum 2021, 51, e1292. [Google Scholar] [CrossRef]
- Raya-Montaño, Y.A.; Apáez-Barrios, P.; Aguirre- Paleo, S.; Vargas -Sandoval, M.; Paz-Da Silva, R.; Lara-Chávez, M.B.N. Identificación de hongos micorrizógenos arbusculares en huertos de aguacate de Uruapan, Michoacán. Rev. Mex. Cienc. Agrícolas 2019, 10, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Acosta, E.; Trejo-Aguilar, D.; Rivera-Fernández, A.; Ferrera-Cerrato, R. La micorriza arbuscular como biofertilizante en cultivo de café. Terra Latinoam. 2020, 38, 613–628. [Google Scholar] [CrossRef]
- Vázquez-Hernández, M.V.; Arévalo-Galarza, M.D.L.; Jaen-Contreras, D.; Escamilla-García, J.L.; Luna-Esquivel, G. Calidad y almacenamiento de frutos de papaya procedentes de plantas inoculadas con Glomus mosseae. Rev. Mex. Cienc. Agrícolas 2020, 11, 1163–1170. [Google Scholar] [CrossRef]
- SMN-CONAGUA. Servicio Meteorológico Nacional. Comisión Nacional del Agua. 2022. Available online: https://smn.conagua.gob.mx/es/ (accessed on 21 October 2022).
- Cadena-Iñiguez, J.; Soto-Hernández, M.; Arévalo-Galarza, M.; Avendaño-Arrazate, C.H.; Aguirre-Medina, J.F.; Ruiz-Posadas, L.D.M. Caracterización bioquímica de variedades domesticadas de chayote Sechium edule (Jacq.) Sw. comparadas con parientes silvestres. Revista Chapingo. Ser. Hortic. 2011, 17, 45–55. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of Soil Analysis, Part 2. Agronomy Monograph 9; Page, A.L., Miller, H., Keeny, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 56–59. [Google Scholar]
- Boschetti, G.; Quintero, C.; Diaz-Zorita, M.; Barraco, M. Determinación del fósforo disponible en el suelo por el método de Bray. Argentina. Actividad del comité de química de la AACS. Inf. Agron. Cono Sur 2003, 17, 6–9. [Google Scholar]
- Chapman, H.D. Cation exchange capacity. In Methods of Soil Analysis. Part 2. Agronomy Monograph 9; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 891–901. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Directions for making mechanical analyses of soils by the hydrometer method. Soil Sci. 1936, 42, 225–230. [Google Scholar] [CrossRef]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- INVAM. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. Species Descriptions from Reference Cultures; West Virginia University: Morgantown, WV, USA, 2016; Available online: https://invam.ku.edu/species-descriptions (accessed on 23 January 2023).
- Steiner, A.A. Selective compacity of tomato plants for ions in a nutrient solution. In Proceedings of the 3d International Congress on Soilless Culture, Sassari, Cerdeña, Italy, 7–12 May 1973; pp. 43–54. [Google Scholar]
- Schenck, N.C.; Pérez, Y. Manual for the Identification of VA Mycorrhizal Fungi, 3rd ed.; Synergistic Publications: Gainesville, FL, USA, 1990; 286p. [Google Scholar]
- Sieverding, E. Manual de Métodos para la Investigación de la Micorriza Vesículo-Arbuscular en el Laboratorio; Centro Internacional de Agricultura Tropical: Cali, Colombia, 1983; p. 121. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Chapman, H.D.; Pratt, P.F. Métodos de Análisis para Suelo, Plantas y Agua; Trillas: Mexico city, Mexico, 1973. [Google Scholar]
- Duarte Alvarado, D.E.; Lagos Burbano, T.C.; Vallejo Cabrera, F.A.; y Lagos Santander, L.K. Agronomic evaluation of lulo introductions Solanum quitoense Lamarck. Acta Agron. 2021, 70, 66–72. [Google Scholar] [CrossRef]
- Gómez-Merino, F.C.; Trejo-Téllez, L.I.; García-Albarado, J.C.; Cadeña-Íñiguez, J. Lulo (Solanum quitoense [Lamarck.]) as new landscape crop in the Mexican agro-ecosystem. Rev. Mex. Cienc. Agrícolas 2014, 5, 1741–1753. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Santander, L.K.; Lagos-Burbano, T.C.; Duarte-Alvarado, D.E.; Criollo-Escobar, H.; Angulo-Ramos, N.F. Evaluación del rendimiento y calidad del fruto de parentales e híbridos de lulo de Castilla. Rev. UDCA Act. Div. Cient. 2019, 22, e1344. [Google Scholar] [CrossRef] [Green Version]
- Medina, C.C.I.; Lobo, A.M.; Martínez, B.E. State of knowledge review on the productive function of lulo (Solanum quitoense Lam.) in Colombia. Rev. Corpoica-Cienc. Tecnol. Agropecu. 2009, 10, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Coronado, A.C.M.; Fagua, A.D.P.R.; Coronado, Y.M. Morphological Characterization of Lulo (Solanum quitoense Lam.) in the Municipality for Pachavita, Boyacá. Acta Biol. Colomb. 2019, 24, 291–298. [Google Scholar] [CrossRef]
- Ramírez-Builes, V.H.; Duque, N.N. Respuesta del lulo La Selva (Solanum quitoense x Solanum hirtum) a la aplicación de fermentados aeróbicos tipo bocashi y fertilizante químico. Acta Agron. 2010, 59, 155–161. [Google Scholar]
- Casierra-Posada, F.; Peña-Olmos, J.; Peñaloza, J.; Roveda, G. Influencia de la sombra y de las micorrizas sobre el crecimiento de plantas de lulo (Solanum quitoense Lam.). Rev. UDCA Actual. Divulg. Cienti. 2013, 16, 61–70. [Google Scholar]
- Miranda, D.; Fischer, G.; Ulrichs, C. The influence of arbuscular mycorrhizal colonization on the growth parameters of cape gooseberry (Physalis peruviana L.) plants grown in a saline soil. J. Soil Sci. Plant Nutr. 2011, 11, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Baltazar-Bernal, O.; Jaén-Contreras, D. Hongos micorrízicos arbusculares y fertilización en Heliconia psittacorum L. f.× H. spathocircinata cv. tropics. Rev. Fitotec. Mex. 2020, 43, 45–52. [Google Scholar] [CrossRef]
- Sánchez, L.R.F.; Espinosa, R.R.; Cárdenas, J.V.M.; Puig, A.C.; Hernández, A.T. Utilización de cepas eficientes de hongos micorrízicos arbusculares en el desarrollo de portainjertos de aguacate en un sustrato suelo-cachaza. Cultiv. Trop. 2011, 32, 23–29. [Google Scholar]
- Díaz-Leguizamón, J.J.; Chingaté-Cruz, O.F.; Sánchez-Reinoso, A.D.; Restrepo-Díaz, H. The effect of foliar applications of a bio-stimulant derived from algae extract on the physiological behavior of lulo seedlings (Solanum quitoense cv. Septentrionale). Cienc. Investig. Agrar. 2016, 43, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Otiniano, C.C.; Hidalgo Rodríguez, J.E.M.; Vera Vega, M.Á.; Pedro Huaman, J.J.; Rodríguez Seminario, C.E.; Chaman Medina, M.E. Efecto del NaCl y micorrizas (Rhizophagus irregularis) en el crecimiento de “tomate” Solanum lycopersicum L. (Solanaceae). Arnaldoa 2021, 28, 675–692. [Google Scholar]
- Dennis, F.G., Jr.; Herner, R.C.; Camacho, S. Naranjilla: A potential cash crop for the small farmer in Latin America. In Proceedings of the X African Symposium on Horticultural Crops, Addis Ababa, Ethiopia, 16–21 January 1984; Volume 158, pp. 475–481. [Google Scholar] [CrossRef]
- Almanza-Fandiño, M.T.A. Management of Bombus atratus Bumblebees to pollinate lulo (Solanum quitoense L), a Native Fruit from the Andes of Colombia; Cuvillier Verlag: Göttingen, Germany, 2007; Volume 50, pp. 50–112. [Google Scholar]
- Flórez, S.L.; Miranda, D.; Chaves, B. Nutrient dynamic in the vegetative growth phase of lulo (Solanum quitoense Lam.) in response to NaCl salinity. Agron. Colomb. 2008, 26, 205–216. [Google Scholar]
- Flórez-Velasco, N.; Balaguera-López, H.E.; Restrepo-Díaz, H. Effects of foliar urea application on lulo (Solanum quitoense cv. septentrionale) plants grown under different waterlogging and nitrogen conditions. Sci. Hortic. 2015, 186, 154–162. [Google Scholar] [CrossRef]
- Aranguren, Y.A.; Castellanos, L.; Escalante, J.C. Hongos formadores de micorrizas arbusculares (HMA) en frutales de Colombia y su comparación con investigaciones internacionales: Arbuscular mycorrhizal fungi (AMF) in Colombian fruit trees and their comparability with international research. Cienc. Tecnol. Agropecu. 2020, 5, 27–37. [Google Scholar]
- Carreón-Abud, Y.; Vega-Fraga, M.; Gavito, M.E. Interaction of arbuscular mycorrhizal inoculants and chicken manure in avocado rootstock production. J. Soil Sci. Plant Nutr. 2015, 15, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Arias-Mota, R.M.; Romero Fernández, A.D.J.; Bañuelos Trejo, J.; Cruz Elizondo, Y.D.L. Inoculation of phosphorus solubilizing fungi and arbuscular mycorrhizae in tomato plants. Rev. Mex. Cienc. Agric. 2019, 10, 1747–1757. [Google Scholar] [CrossRef] [Green Version]
- González, O.; Osorio, W. Determinación de la dependencia micorrizal del lulo. Acta Biol. Colomb. 2008, 13, 163–174. [Google Scholar]
- Vázquez-Hernández, M.V.; Arévalo-Galarza, L.; Jaen-Contreras, D.; Escamilla-García, J.L.; Mora-Aguilera, A.; Hernández-Castro, E.; Téliz-Ortiz, D. Effect of Glomus mosseae and Entrophospora colombiana on plant growth, production, and fruit quality of ‘Maradol’papaya (Carica papaya L.). Sci. Hortic. 2011, 128, 255–260. [Google Scholar] [CrossRef]
- Yang, G.H.; Yang, L.T.; Jiang, H.X.; Li, Y.; Wang, P.; Chen, L.S. Physiological impacts of magnesium-deficiency in Citrus seedlings: Photosynthesis, antioxidant system and carbohydrates. Trees 2012, 26, 1237–1250. [Google Scholar] [CrossRef]
- Parra-Coronado, A.; Ardila-Roa, G.H.; Restrepo-Díaz, H. The physiological response of lulo plants (Solanum quitoense var. Septentrionale) to soil and foliar applications of nutrients. Int. J. Fruit Sci. 2015, 15, 148–160. [Google Scholar] [CrossRef]
- Cavallaro Júnior, M.L.; Trani, P.E.; Passos, F.A.; Kuhn Neto, J.; Tivelli, S.W. Produtividade de rúcula e tomate em função da adubação N e P orgânica e mineral. Bragantia 2009, 68, 347–356. [Google Scholar] [CrossRef]
- Schroeder, M.A.; Martínez, G.C. Concentraciones foliares de microelementos en Pimiento (Capsicum annuum L.). Agrotecnia 2004, 13, 17–21. [Google Scholar] [CrossRef]
- Loué, A. Los Microelementos en Agricultura; Mundi Prensa: Madrid, Spain, 1986; 166p. [Google Scholar]
- Herrera, J.G.A.; Guerrero, M.J.; Fischer, G. Aspectos de la fisiología y el cultivo del lulo (Solanum quitoense LAM.) en Colombia: Una revisión. Rev. Investig. Agrar. Ambient. 2022, 13, 131–148. [Google Scholar] [CrossRef]
- Bolívar, M.I.V.; Medellín, L.A.C.; Trujillo, M.M.P. Efecto de las deficiencias de algunos nutrientes en plantas de lulo (Solanum quitoense var. quitoense) en etapa de vivero. Rev. Fac. Cienc. Básicas 2009, 5, 64–81. [Google Scholar] [CrossRef]
- Cabezas, M.; Botía, T.; Medina, L.M. Determinación de síntomas de deficiencia inducida de nutrimentos en lulo (Solanum quitoense Lam.). In IV Seminario Nacional de Frutales de Clima Frío Moderado; Corpoica: Mosquera, Colombia, 2003; pp. 176–181. Available online: http://hdl.handle.net/20.500.12324/13648 (accessed on 11 February 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Content (mg kg−1) | Interpretation |
|---|---|---|
| N | 14.3 | Low |
| P | 11.3 | Low |
| K | 63.0 | Low |
| Ca | 3010 | High |
| Mg | 570 | High |
| Fe | 52.12 | Very high |
| Zn | 1.45 | Low |
| Mn | 57.30 | Very high |
| Cu | 3.04 | Very high |
| CEC | 14.64 cmol kg−1 | Low |
| Texture | ||
| Organic matter | 4.02 (%) | Rich |
| Arena | 17.16 (%) | Clay soil |
| Limo | 13.08 (%) | |
| Clay | 69.76 (%) | |
| pH | 6.39 | Slightly acidic |
| Treatment | Hyphae (%) | Vesicles (%) | Arbuscules (%) | Colonization Total (%) |
|---|---|---|---|---|
| Entrophospora colombiana | 63.20 a | 32.10 a | 36.50 a | 70.50 a |
| Funneliformis mosseae | 53.12 b | 10.70 b | 24.78 b | 59.30 b |
| Diammonium phosphate (DAP), native colonization | 59.70 a | 14.70 b | 28.70 b | 63.40 a |
| Control (native colonization) | 21.36 c | 1.47 c | 0.98 c | 9.74 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olguín-Hernández, A.L.; Arévalo-Galarza, M.d.L.; Cadena-Iñiguez, J.; Jaén-Contreras, D.; Peña-Valdivia, C.B. Plant Height and Stem Diameter of Solanum quitoense Lamarck Improved with Applications of AMF and Biostimulants. Agriculture 2023, 13, 1420. https://doi.org/10.3390/agriculture13071420
Olguín-Hernández AL, Arévalo-Galarza MdL, Cadena-Iñiguez J, Jaén-Contreras D, Peña-Valdivia CB. Plant Height and Stem Diameter of Solanum quitoense Lamarck Improved with Applications of AMF and Biostimulants. Agriculture. 2023; 13(7):1420. https://doi.org/10.3390/agriculture13071420
Chicago/Turabian StyleOlguín-Hernández, Ana Laura, Ma. de Lourdes Arévalo-Galarza, Jorge Cadena-Iñiguez, David Jaén-Contreras, and Cecilia B. Peña-Valdivia. 2023. "Plant Height and Stem Diameter of Solanum quitoense Lamarck Improved with Applications of AMF and Biostimulants" Agriculture 13, no. 7: 1420. https://doi.org/10.3390/agriculture13071420
APA StyleOlguín-Hernández, A. L., Arévalo-Galarza, M. d. L., Cadena-Iñiguez, J., Jaén-Contreras, D., & Peña-Valdivia, C. B. (2023). Plant Height and Stem Diameter of Solanum quitoense Lamarck Improved with Applications of AMF and Biostimulants. Agriculture, 13(7), 1420. https://doi.org/10.3390/agriculture13071420