
Response of Strawberry Fruit Yield, Soil Chemical and Microbial Properties to Anaerobic Soil Disinfestation with Biochar and Rice Bran


 ,
, 

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Soil Sampling
2.4. Detection of Soil-Borne Pathogens
2.5. Analysis of Soil Physicochemical Properties
2.6. Strawberry Plant Growth and Yield
2.7. High-Throughput Gene Sequencing
2.8. Statistical Analysis
3. Results
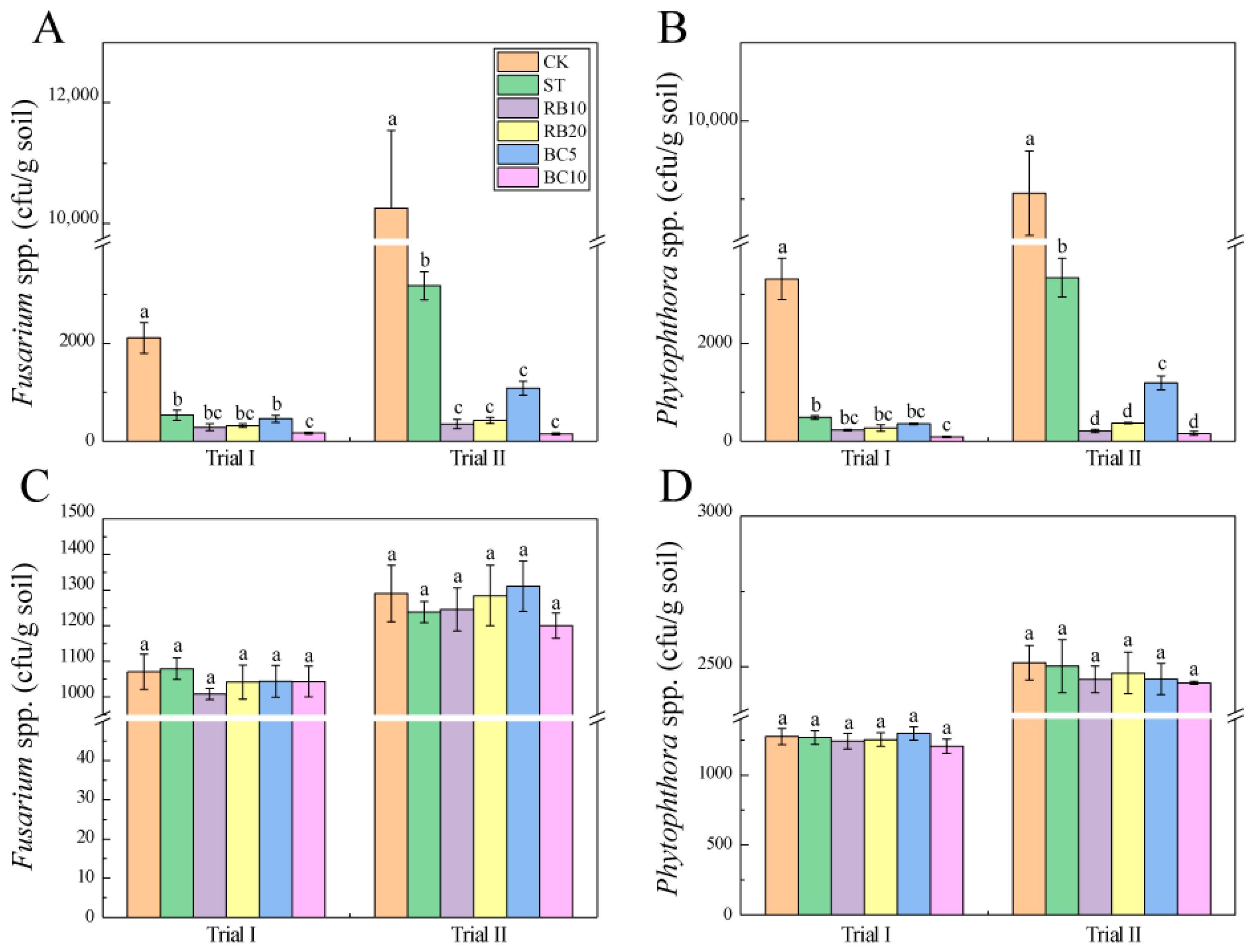
3.1. Soil Pathogens Control
3.2. Physicochemical Properties of Soil
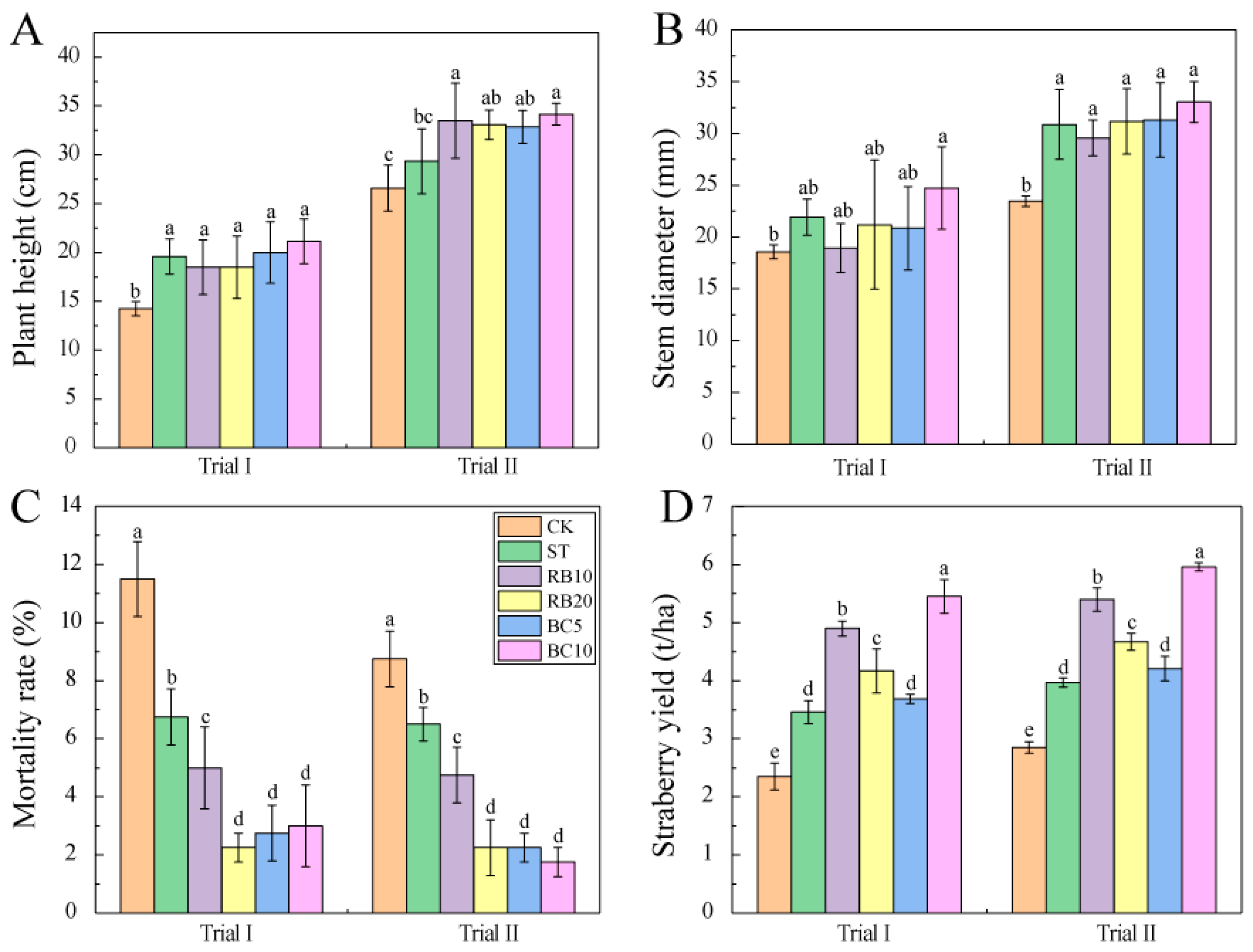
3.3. Strawberry Growth and Yield
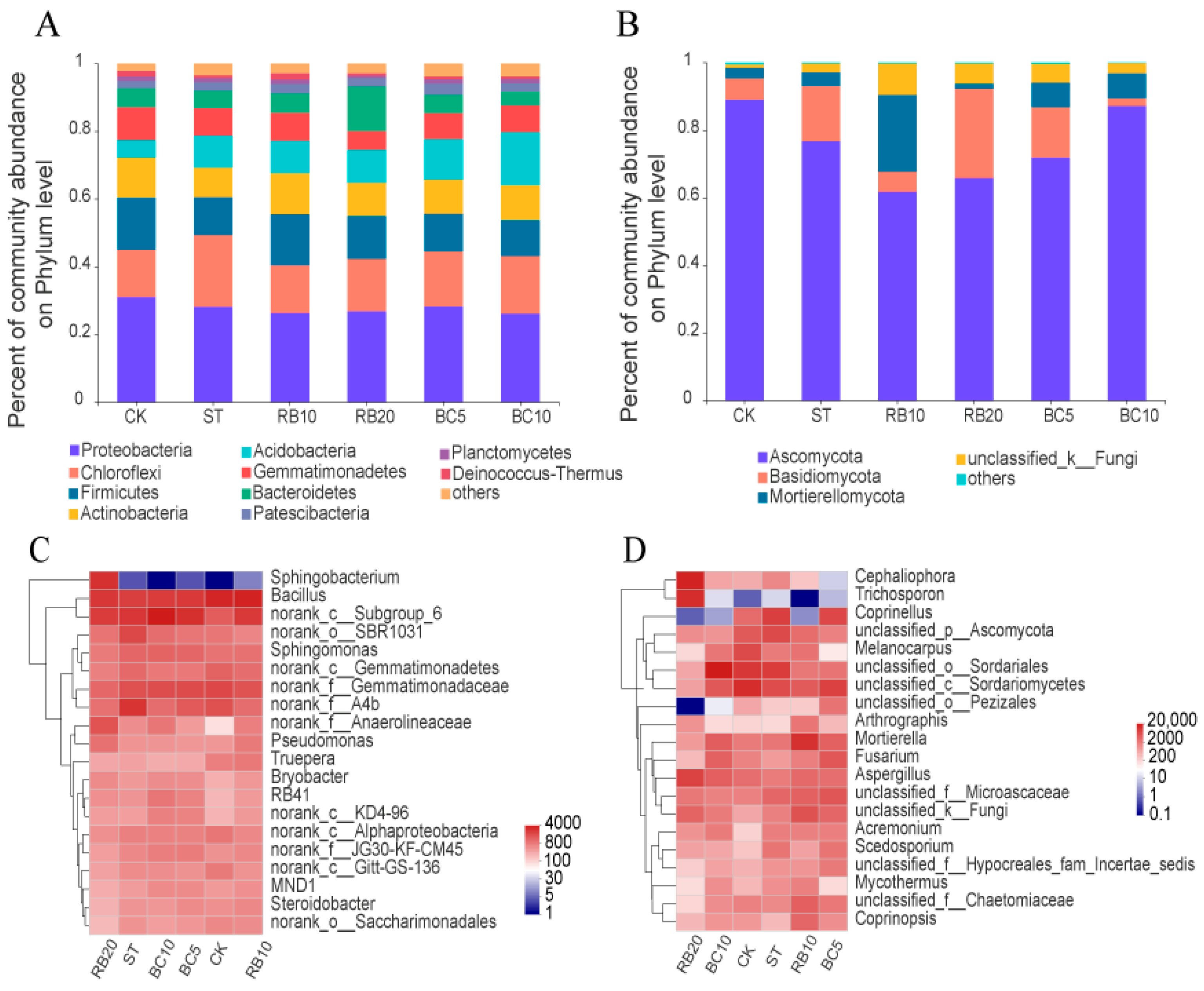
3.4. Changes to the Taxonomic Composition of Soil Microbial Communities
3.4.1. Base Pair Length and Rarefaction Curves
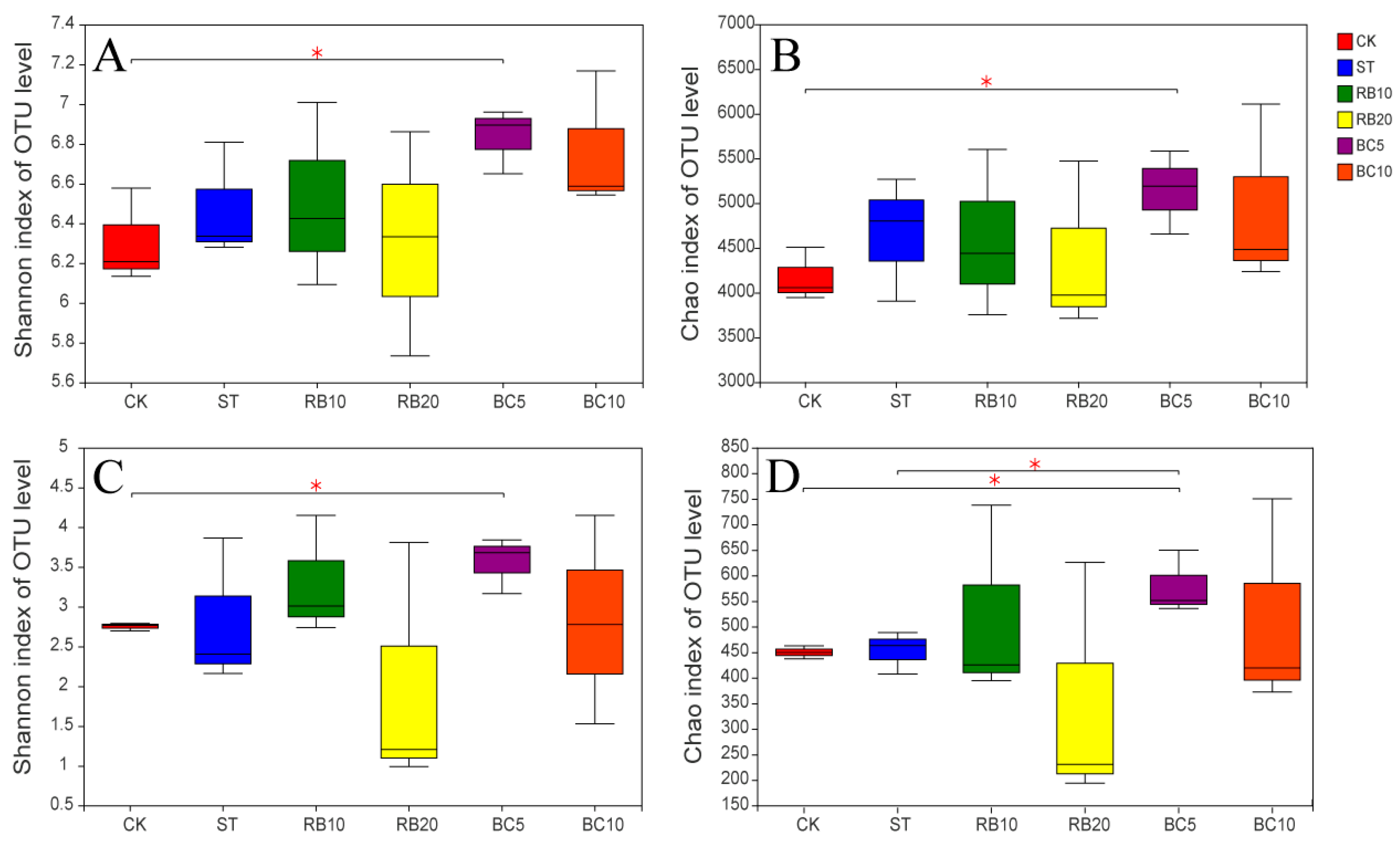
3.4.2. Alpha Diversity
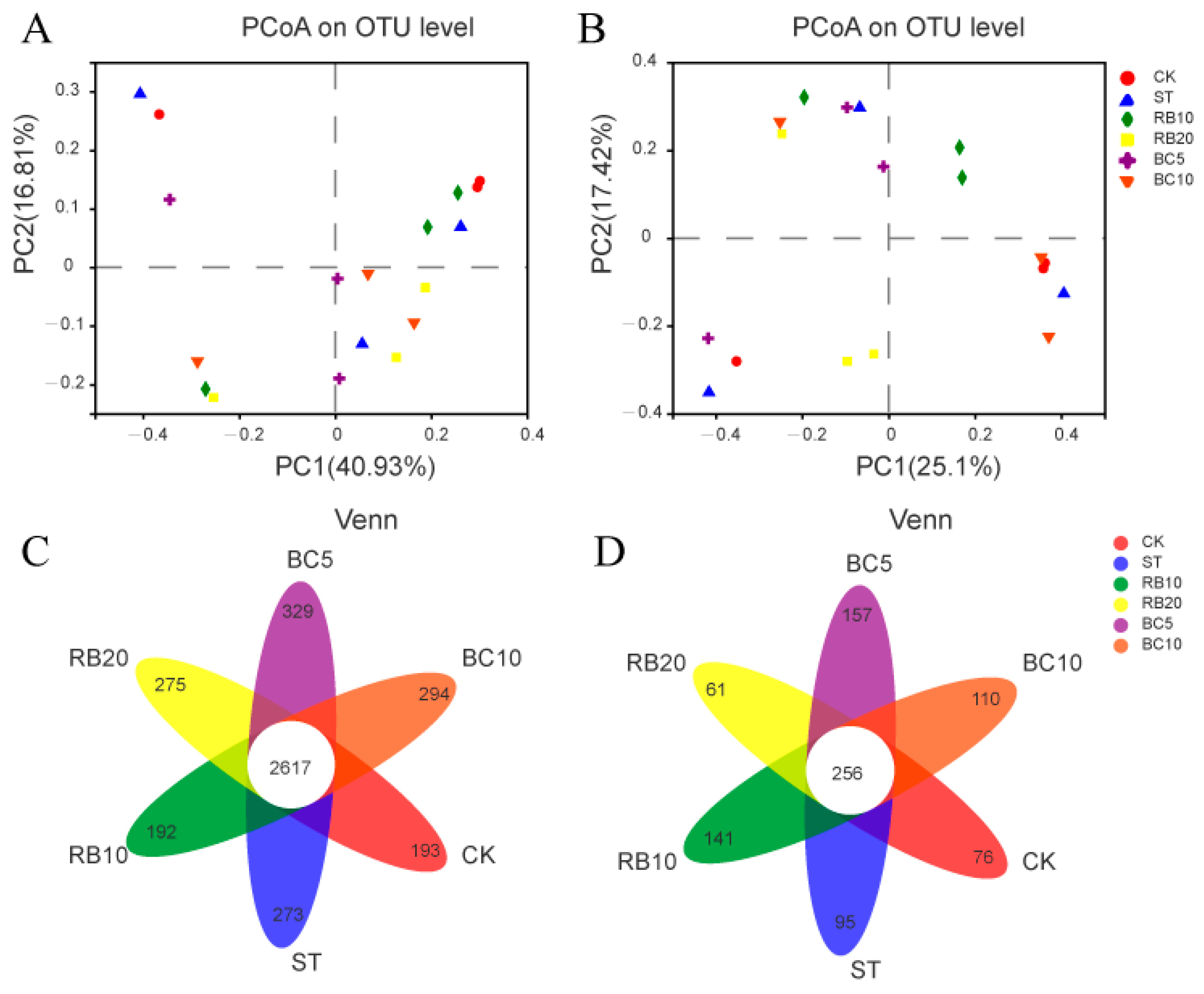
3.4.3. Beta Diversity and Venn Diagram
3.4.4. Changes in Soil Microbial Communities
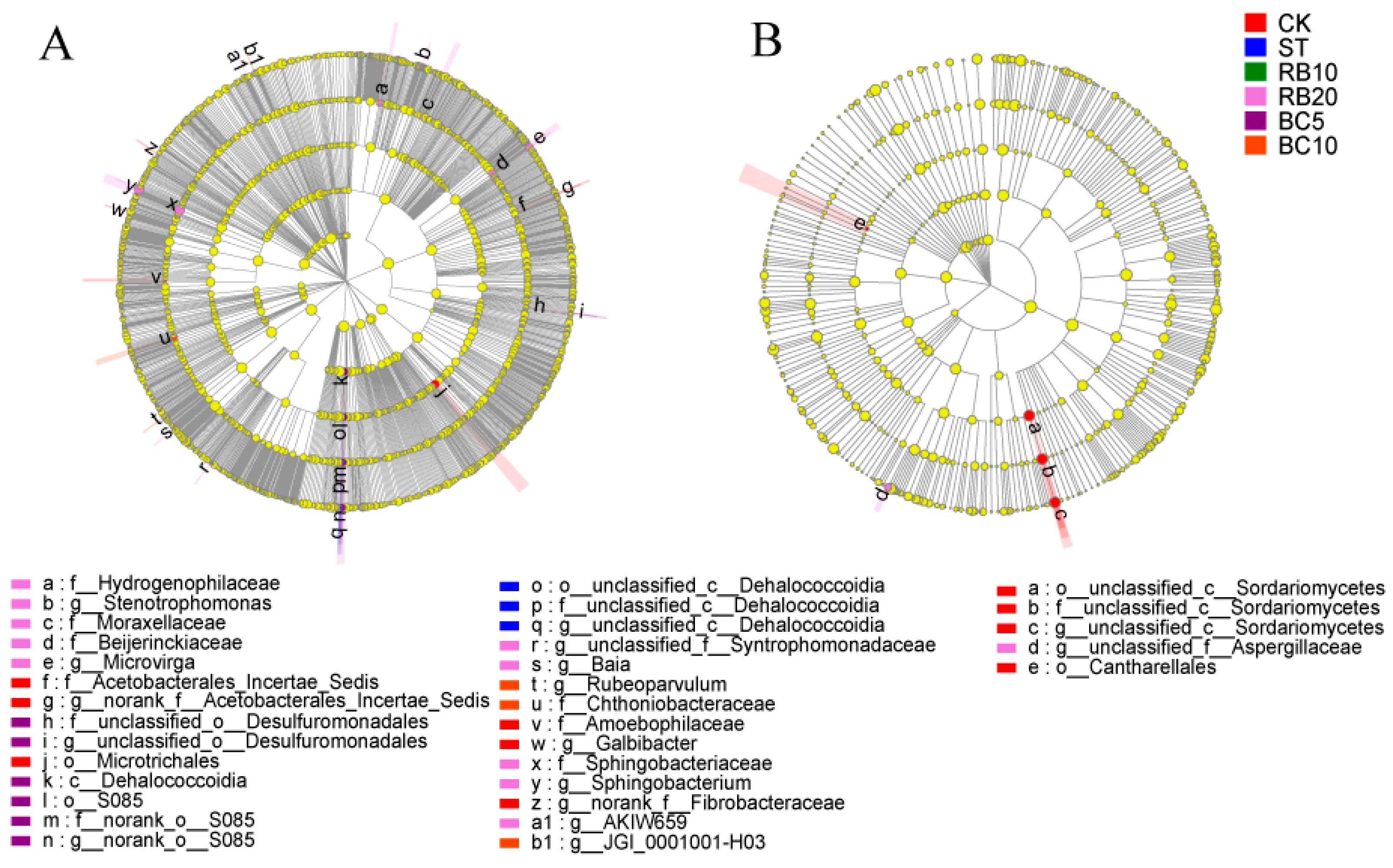
3.5. Biomarker Analysis of Soil Microbial Communities
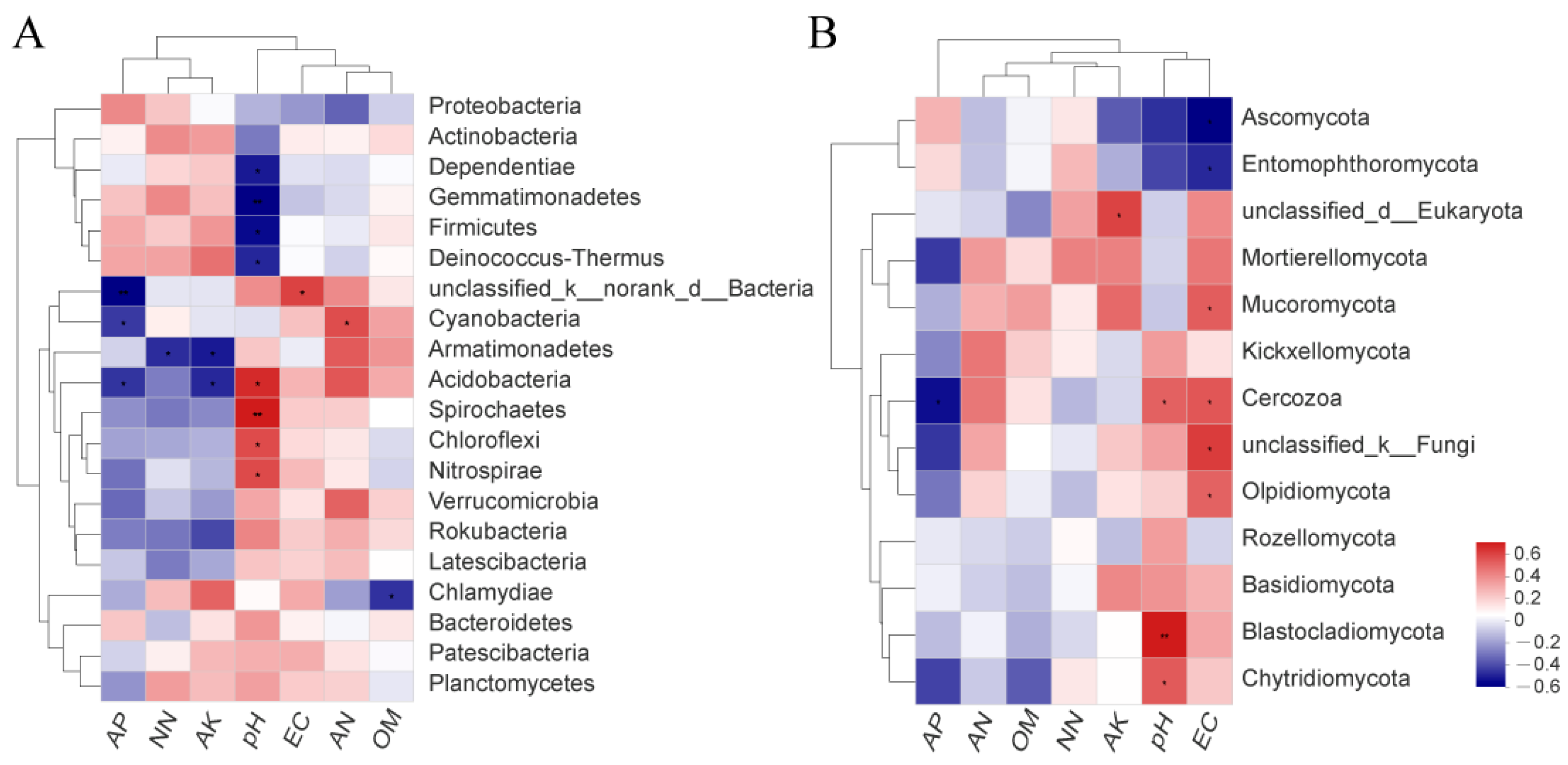
3.6. Correlation between Soil Physicochemical Properties and Microorganisms
4. Discussion
4.1. Effects of ASD on Soil Pathogens
4.2. Effects of ASD on Soil Physicochemical Properties
4.3. Effects of ASD on Strawberry Plant Growth and Fruit Yield
4.4. Effects of ASD on Microbial Taxonomic Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, F.; Ren, W.; Fang, X.; Chen, L.; Zha, X. Residues, dissipation, and safety evaluation of pymetrozine-clothianidin mixture in strawberry. Environ. Sci. Pollut. Res. Int. 2021, 28, 22641–22650. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.; Zhang, D.Q.; Ren, L.R.; Song, Z.X.; Li, Q.J.; Wu, J.J.; Fang, W.S.; Huang, B.; Yan, D.D.; Li, Y.; et al. Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. Pollut. 2021, 283, 117160. [Google Scholar] [CrossRef] [PubMed]
- Iapichino, G.; Prinzivalli, C.; D’Anna, F. Soil solarization as an alternative to methyl bromide fumigation for annual strawberry production in a Mediterranean Area. J. Sustain. Agr. 2008, 32, 365–375. [Google Scholar] [CrossRef]
- He, Y.; Chen, J.; Tang, C.; Deng, Q.; Guo, L.; Cheng, Y.; Li, Z.; Wang, T.; Xu, J.; Gao, C. Genetic diversity and population structure of fusarium commune causing strawberry root rot in southcentral China. Genes 2022, 13, 899. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, M.Y.; Zhao, S.C. Research progress of strawberry verticillium wilt. Anhui Agric. Sci. Bull. 2021, 27, 111–114. [Google Scholar]
- Li, Q.J.; Zhang, D.Q.; Song, Z.X.; Ren, L.R.; Jin, X.; Fang, W.S.; Yan, D.D.; Li, Y.; Wang, Q.X.; Cao, A.C. Organic fertilizer activates soil beneficial microorganisms to promote strawberry growth and soil health after fumigation. Environ. Pollut. 2022, 295, 118653. [Google Scholar] [CrossRef]
- Li, H.; Yang, H.Y.; Calderón-Urrea, A.; Li, Y.P.; Zhang, L.P.; Yu, Y.L.; Ma, J.Y.; Shi, G.Y. Microbial Fertilization Improves Soil Health When Compared to Chemical Fumigation in Sweet Lily. J. Fungi 2022, 8, 847. [Google Scholar] [CrossRef]
- Wang, C.; Ma, X.H.; Wang, G.; Li, G.T.; Zhu, K. Implication of O2 dynamics for both N2O and CH4 emissions from soil during biological soil disinfestation. Sci. Rep. 2021, 11, 6590. [Google Scholar] [CrossRef]
- Utsala, S.; Mary, E.D.; Bonnie, H.O.; David, M.B. Anaerobic soil disinfestation reduces germination and affects colonization of Sclerotium rolfsii sclerotia. Phytopathology 2018, 108, 342–351. [Google Scholar]
- Murphy, C.M.; Weller, D.L.; Reiter, M.S.; Bardsley, C.A.; Eifert, J.; Ponder, M.; Rideout, S.L.; Strawn, L.K. Anaerobic soil disinfestation, amendment-type, and irrigation regimen influence Salmonella survival and die-off in agricultural soils. J. Appl. Microbiol. 2022, 132, 2342–2354. [Google Scholar] [CrossRef]
- Song, Z.X.; Yan, D.D.; Fang, W.S.; Wang, Q.X.; Li, Y.; Xu, J.; Jin, X.; Cao, A.C. Anaerobic soil disinfestation technology, action mechanisms and influencing factors. J. Plant Prot. 2022, 49, 994–1003. [Google Scholar]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soil-borne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef]
- Song, Z.X.; Yan, D.D.; Fang, W.S.; Huang, B.; Wang, X.L.; Zhang, D.Q.; Zhu, J.H.; Liu, J.; Ouyang, C.B.; Li, Y.; et al. Maltose and totally impermeable film enhanced suppression of anaerobic soil disinfestation on soil-borne pathogens and increased strawberry yield. Sustainability 2020, 12, 5456. [Google Scholar] [CrossRef]
- Cai, Z.C.; Zhang, J.B.; Huang, X.Q.; Zhu, T.B.; Wen, T. Application of reductive soil disinfestation to suppress soil-borne pathogens. Acta Pedol. Sin. 2015, 52, 469–476. [Google Scholar]
- Patwa, D.; Bordoloi, U.; Dubey, A.A.; Ravi, K.; Sekharan, S.; Kalita, P. Energy-efficient biochar production for thermal backfill applications. Sci. Total Environ. 2022, 833, 155253. [Google Scholar] [CrossRef]
- Zheng, X.M.; Xu, W.H.; Dong, J.; Yang, T.; Shangguan, Z.C.; Qu, J.; Li, X.; Tan, X.F. The effects of biochar and its applications in the microbial remediation of contaminated soil: A review. J. Hazard. Mater. 2022, 438, 129557. [Google Scholar] [CrossRef] [PubMed]
- Ginebra, M.; Muñoz, C.; Calvelo-Pereira, R.; Doussoulin, M.; Zagal, E. Biochar impacts on soil chemical properties, greenhouse gas emissions and forage productivity: A field experiment. Sci. Total Environ. 2022, 806, 150465. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chen, Y.P.; Su, C.C.; Ma, J.F.; Wu, J.H.; Wang, K.B.; Wang, Y.; Jiang, Y. Microbial characteristics response to the soil quality of newly created farmland on the Loess Plateau. Environ. Sci. Pollut. Res. Int. 2021, 28, 42290–42299. [Google Scholar] [CrossRef]
- Mehrabi, Z.; McMillan, V.E.; Clark, I.M.; Canning, G.; Hammond-Kosack, K.E.; Preston, G.; Hirsch, P.R.; Mauchline, T.H. Pseudomonas spp. diversity is negatively associated with suppression of the wheat take-all pathogen. Sci. Rep. 2016, 6, 29905. [Google Scholar] [CrossRef]
- Zhang, D.Q.; Yan, D.D.; Fang, W.S.; Huang, B.; Wang, X.L.; Wang, X.N.; Zhu, J.H.; Liu, J.; Ouyang, C.B.; Li, Y.; et al. Chloropicrin alternated with biofumigation increases crop yield and modifies soil bacterial and fungal communities in strawberry production. Sci. Total Environ. 2019, 675, 615–622. [Google Scholar] [CrossRef]
- Momma, N.; Kobara, Y.; Momma, M. Fe2+ and Mn2+, potential agents to induce suppression of Fusarium oxysporum for biological soil disinfestation. J. Gen. Plant Pathol. 2011, 77, 331–335. [Google Scholar] [CrossRef]
- Komada, H. Development of a selective medium for quantitative isolation of Fusarium oxysporum from natural soil. Rev. Plant Prot. Res. 1975, 8, 114–125. [Google Scholar]
- Masago, H.; Yoshikawa, M.; Fukada, M.; Nakanishi, N. Selective inhibition of pythium spp. on a medium for direct isolation of phytophthora spp. from soils and plants. Phytopathology 1977, 77, 425–428. [Google Scholar] [CrossRef]
- Oka, Y. Mechanisms of nematode suppression by organic soil amendments—A review. Appl. Soil Ecol. 2010, 44, 101–115. [Google Scholar] [CrossRef]
- Runia, W.T.; Molendijk, L.P.G. Physical methods for soil disinfestation in intensive agriculture: Old methods and new approaches. Acta Hortic. 2010, 883, 249–258. [Google Scholar] [CrossRef]
- Overbeek, L.V.; Runia, W.; Kastelein, P.; Molendijk, L. Anaerobic disinfestation of tare soils contaminated with Ralstonia solanacearum biovar 2 and Globodera pallida. Eur. J. Plant Pathol. 2014, 138, 323–330. [Google Scholar] [CrossRef]
- Song, Z.X.; Massart, S.; Yan, D.D.; Cheng, H.Y.; Eck, M.; Berhal, C.; Ouyang, C.B.; Li, Y.; Wang, Q.X.; Cao, A.C. Composted Chicken Manure for Anaerobic Soil Disinfestation Increased the Strawberry Yield and Shifted the Soil Microbial Communities. Sustainability 2020, 12, 6313. [Google Scholar] [CrossRef]
- Li, Q.J.; Fang, W.S.; Zhang, D.Q.; Yan, D.D.; Li, Y.; Wang, Q.X.; Cao, A.C. Effects of dazomet fumigation combined with biochar and zeolite on soil microecology and soil-borne pathogens. Plant Prot. 2022, 48, 150–157. [Google Scholar]
- Huang, X.Q.; Wen, T.; Meng, L.; Zhang, J.B.; Zhu, T.; Cai, Z.C. Anaerobic Soil Disinfestation on Fusarium oxysporum. Soils 2014, 46, 851–855. [Google Scholar]
- Drury, C.F.; Findlay, W.I.; McKenney, D.J. Oxygen inhibition of denitrification in chloroform fumigated and non-fumigated soil. Soil Biol. Biochem. 2021, 23, 711–715. [Google Scholar] [CrossRef]
- Wu, C.R.; Huang, F.; Gao, Y.; Mao, Y.H.; Cai, K.Z. Effect of biological disinfestation on soil improvement, Ralstonia solanacearum suppression and tomato growth. Chin. J. Eco-Agr. 2017, 25, 1173–1180. [Google Scholar]
- Tian, J.H.; Kuang, X.; Tang, M.T.; Chen, X.D.; Huang, F.; Cai, Y.X.; Cai, K.Z. Biochar application under low phosphorus input promotes soil organic phosphorus mineralization by shifting bacterial phoD gene community composition. Sci. Total Environ. 2021, 779, 146556. [Google Scholar] [CrossRef]
- Dai, Z.G.; Lu, J.W.; Lu, M.X.; Yang, W.B.; Fan, X.P. Effect of rape straw rate on physicochemical properties of soil surface solution under waterlogged incubation. J. Soil Water Conserv. 2010, 24, 200–204. [Google Scholar]
- Zhu, T.B.; Zhang, J.B.; Cai, Z.C. Effects of organic material amendment on vegetable soil nitrate content and nitrogenous gases emission under flooding condition. Chin. J. Appl. Ecol. 2012, 23, 109–114. [Google Scholar]
- Liu, Y.N.; He, G.S.; Wei, J.Y.; Jia, H.J.; Huang, C.J.; Cai, Y.X.; Cai, K.Z.; Pan, B.G.; Wang, W. Effects of anaerobic soil disinfestation on the quality and bacterial community of tobacco-growing soils. Chin. Tobacco Sci. 2019, 40, 39–46. [Google Scholar]
- Hewavitharana, S.S.; Mazzola, M. Carbon source-dependent effects of anaerobic soil disinfestation on soil microbiome and suppression of rhizoctonia solani AG-5 and pratylenchus penetrans. Phytopathology 2016, 106, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, U.; Augé, R.M.; Butler, D.M. A meta-analysis of the impact of anaerobic soil disinfestation on pest suppression and yield of horticultural crops. Front. Plant Sci. 2016, 7, 1254. [Google Scholar] [CrossRef]
- Butler, D.M.; Kokalis-Burelle, N.; Albano, J.P.; McCollum, T.G.; Muramoto, J.; Shennan, C.; Rosskopf, E.N. Anaerobic soil disinfestation (ASD) combined with soil solarization as a methyl bromide alternative: Vegetable crop performance and soil nutrient dynamics. Plant Soil 2014, 378, 365–381. [Google Scholar] [CrossRef]
- Wang, X.Y.; Duan, Y.; Zhang, J.; Ciampitti, I.A.; Cui, J.W.; Qiu, S.J.; Xu, X.P.; Zhao, S.C.; He, P. Response of potato yield, soil chemical and microbial properties to different rotation sequences of green manure-potato cropping in North China. Soil Till. Res. 2022, 217, 105273. [Google Scholar] [CrossRef]
- Feng, X.Y.; Wang, Q.L.; Sun, Y.H.; Zhang, S.W.; Wang, F.Y. Microplastics change soil properties, heavy metal availabilit and bacterial community in a Pb-Zncontaminated soil. J. Hazard. Mater. 2022, 424, 127364. [Google Scholar] [CrossRef]
- Bauld, J.; Brock, T.D. Ecological studies of Chloroflexis, a gliding photosynthetic bacterium. Arch. Microbiol. 1973, 92, 267–284. [Google Scholar] [CrossRef]
- Hausmann, B.; Pelikan, C.; Herbold, C.W.; Köstlbacher, S.; Albertsen, M.; Eichorst, S.A.; Glavina, D.R.T.; Huemer, M.; Nielsen, P.H.; Rattei, T.; et al. Peatland Acidobacteria with a dissimilatory sulfur metabolism. ISME J. 2018, 12, 1729–1742. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Y.; Deng, J.B.; Chi, S.L.; Wang, W.Z.; Xu, L.S.; Huang, Q.Z.; Zhang, Y.M.; Yu, X.M.; Xu, J.; Chen, Y.C.; et al. Sustainability assessment of topsoil ecology in Chongqing, China based on the application of livestock and poultry manure. J. Clean. Prod. 2022, 358, 131969. [Google Scholar] [CrossRef]
- Zampieri, B.D.B.; Pinto, A.B.; Schultz, L.; Oliveira, M.A.D.; Oliveira, A.J.F. Diversity and distribution of heavy metal-resistant bacteria in polluted sediments of the araça bay, são sebastião (SP), and the relationship between heavy metals and organic matter concentrations. Microb. Ecol. 2021, 72, 582–594. [Google Scholar] [CrossRef]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes, as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Liu, W.R.; Zeng, D.; She, L.; Su, W.X.; He, D.C.; Wu, G.Y.; Ma, X.R.; Jiang, S.; Jiang, C.H.; Ying, G.G. Comparisons of pollution characteristics, emission situations, and mass loads for heavy metals in the manures of different livestock and poultry in China. Sci. Total Environ. 2020, 734, 139023. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glasshouses Soil | Sand % | Clay % | Silt % | Ammonium Nitrogen (mg/kg) | Nitrate Nitrogen (mg/kg) | Available Phosphorus (mg/kg) | Available Potassium (mg/kg) | Organic Matter (g/kg) | pH (1:2.5) | Electrical Conductivity (μS/cm) |
|---|---|---|---|---|---|---|---|---|---|---|
| Trial I | 60 | 3 | 37 | 10.4 | 188.4 | 1102.3 | 982.8 | 23.6 | 7.9 | 895.4 |
| Trial II | 57 | 6 | 37 | 6.6 | 146.9 | 734.9 | 470.9 | 16.9 | 7.5 | 954.3 |
| Treatment | AN (mg/kg) | NN (mg/kg) | AP (mg/kg) | AK (mg/kg) | OM (g/kg) | pH (1:2.5) | EC (μS/cm) |
|---|---|---|---|---|---|---|---|
| CK | 6.92 ± 0.40 f | 282.60 ± 5.44 a | 944 ± 20.55 a | 605 ± 29.57 b | 14.40 ± 0.47 c | 6.54 ± 0.21 c | 228 ± 3.06 d |
| ST | 12.18 ± 0.60 e | 87.55 ± 7.39 d | 882 ± 57.29 ab | 469 ± 41.68 cd | 14.58 ± 0.55 c | 7.41 ± 0.27 ab | 1142 ± 56.82 b |
| RB10 | 18.22 ± 0.72 b | 165.32 ± 12.67 b | 801 ± 34.64 bc | 710 ± 57.83 a | 18.30 ± 1.20 a | 7.15 ± 0.21 b | 1372 ± 33.47 a |
| RB20 | 14.89 ± 0.43 c | 86.46 ± 7.97 d | 914 ± 24.34 a | 408 ± 22.11 de | 18.22 ± 0.56 a | 7.38 ± 0.25 ab | 614 ± 59.02 c |
| BC5 | 13.82 ± 0.44 d | 138.98 ± 4.10 c | 669 ± 32.52 d | 523 ± 41.00 c | 16.40 ± 0.36 b | 7.73 ± 0.29 a | 1338 ± 115.76 a |
| BC10 | 21.87 ± 0.45 a | 98.34 ± 2.87 d | 774 ± 85.91 c | 381 ± 34.65 e | 19.52 ± 0.87 a | 7.22 ± 0.40 ab | 596 ± 84.07 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Z.; Yan, D.; Fang, W.; Zhang, D.; Jin, X.; Li, Y.; Wang, Q.; Wang, G.; Li, Q.; Cao, A. Response of Strawberry Fruit Yield, Soil Chemical and Microbial Properties to Anaerobic Soil Disinfestation with Biochar and Rice Bran. Agriculture 2023, 13, 1466. https://doi.org/10.3390/agriculture13071466
Song Z, Yan D, Fang W, Zhang D, Jin X, Li Y, Wang Q, Wang G, Li Q, Cao A. Response of Strawberry Fruit Yield, Soil Chemical and Microbial Properties to Anaerobic Soil Disinfestation with Biochar and Rice Bran. Agriculture. 2023; 13(7):1466. https://doi.org/10.3390/agriculture13071466
Chicago/Turabian StyleSong, Zhaoxin, Dongdong Yan, Wensheng Fang, Daqi Zhang, Xi Jin, Yuan Li, Qiuxia Wang, Guirong Wang, Qingjie Li, and Aocheng Cao. 2023. "Response of Strawberry Fruit Yield, Soil Chemical and Microbial Properties to Anaerobic Soil Disinfestation with Biochar and Rice Bran" Agriculture 13, no. 7: 1466. https://doi.org/10.3390/agriculture13071466
APA StyleSong, Z., Yan, D., Fang, W., Zhang, D., Jin, X., Li, Y., Wang, Q., Wang, G., Li, Q., & Cao, A. (2023). Response of Strawberry Fruit Yield, Soil Chemical and Microbial Properties to Anaerobic Soil Disinfestation with Biochar and Rice Bran. Agriculture, 13(7), 1466. https://doi.org/10.3390/agriculture13071466