
Breeding for Biotic Stress Resistance in Pea


Abstract
:1. Introduction
2. State of the Art by Groups of Diseases and Pests
2.1. Ascochyta Blight
2.2. Mildews
2.3. Rusts
2.4. Wilts and Root Rots
2.5. Root Parasitic Nematodes
2.6. Broomrapes
2.7. Bacterial Blight
2.8. Viruses
2.9. Insect Pests

{kind=link}
{kind=link}
| Biotic Stress | Pathogen | Gene/QTL | Effect | Linkage Group | Resistance Type | Reference |
|---|---|---|---|---|---|---|
| Aerial fungi or oomycete | ||||||
| Ascochyta blight | Peyronellaea pinodes | Dp1.1, Dp1.2, Dp1.3, | Minor to moderate | Chr2LGI | Incomplete | [6,7,11] |
| MpII.1, | Chr6LGII | |||||
| Dp3.1, Dp3.2, Dp3.3, Dp3.4, | Chr5LGIII | |||||
| Dp3.5, Dp3.6, Dp3.7, Dp3.8, | ||||||
| Dp3.9, MpIV.1 | Chr4LGIV | |||||
| Dp5.1, Dp5.2, Dp5.3, | Chr3LGV | |||||
| Dp6.1, Dp6.2, Dp6.3, Dp6.4, | Chr1LGVI | |||||
| Dp7.1, Dp7.2, Dp7.3 | Chr7LGVII | |||||
| Powdery mildew | Erysiphe pisi | er1, | Major | Chr1LGVI | Incomplete | [22,23] |
| er2, | Major | Chr5LGIII | ||||
| Er3 | Major | Chr4LGIV | ||||
| Downy mildew | Peronospora viciae f. sp. pisi | 3552605, | Chr1LGVI | Complete | [39,41] | |
| 3559062, | Chr3LGV | |||||
| 5943381 | Chr6LGII | |||||
| Rpv | Major | Chr2LGI | ||||
| rpv-1 | Minor | |||||
| rpv-2 | Minor | |||||
| Rust | Uromyces pisi | UpDSII, | Major | Incomplete | [46] | |
| UpDSIV, | Major | |||||
| UpDSIV.2 | Minor | |||||
| Uromyces viciae-fabae | Ruf | Major | Incomplete | [44] | ||
| Soilborne fungi or oomycete | ||||||
| Fusarium root rot | Fusarium solani f. sp. pisi | Fsp-Ps2.1, | Chr6LGII | Incomplete | [68,69] | |
| Fsp-Ps6.1, | Chr1LGVI | |||||
| Fsp-Ps3.1, Fsp-Ps3.2, Fsp-Ps3.3, | Chr5LGIII | |||||
| Fsp-4.1, | Chr7LGVII | |||||
| Fsp-Ps7.1 | ||||||
| F. graminearum | Fg-Ps3.1, Fg-s3.2, | Minor | Chr5LGIII | Incomplete | [71] | |
| Fg-Ps4.1, Fg-s4.2, | Moderate | Chr4LGIV | ||||
| Fg-Ps5.1 | Minor | Chr3LGV | ||||
| Fusarium wilt | F. oxysporum. f. sp. pisi race 1 | Fw | Major | Chr5LGIII | Complete | [56,57,58] |
| F. oxysporum. f. sp. pisi race 2 | Fnw 3.1, Fnw 3.2, | Minor | Chr5LGIII | Complete | [60] | |
| Fnw 4.1 | Major | Chr4LGIV | ||||
| F. oxysporum. f. sp. pisi race 5 | Fwf | Major | Chr6LGII | Complete | [55] | |
| Common root rot | Aphanomyces euteiches | Ae-Ps1.1, Ae-Ps1.2 | Minor | Chr2LGI | Incomplete | [74,75,76] |
| Ae-Ps2.1, Ae-Ps2.2 | Minor | Chr6LGII | ||||
| Ae-Ps3.1, Ae-Ps3.2 | Minor | Chr5LGIII | ||||
| Ae-Ps4-4, Ae-Ps4.5 | Minor | Chr4LGIV | ||||
| Ae-Ps5.1, | Minor | Chr3LGV | ||||
| Ae-Ps6.1, | Minor | Chr1LGVI | ||||
| Ae-Ps7.6 | Major | Chr7LG7 | ||||
| Bacteria | ||||||
| Pea blight | Pseudomonas syringae pv. pisi | Ppi1, | Chr1LGVI | complete | [108,109,110] | |
| Ppi2, | Chr7LGVII | |||||
| Ppi3, Ppi4, | Chr6LGII, | |||||
| Ppi8 | Chr5LGIII | |||||
| Pseudomonas syringae pv. syringae | PsBB1-Psy | Minor | Chr6LGII | complete | [111,112] | |
| Psy1, PsBB3-Psy, PsBB4-Psy | Major | Chr5LGIII | ||||
| Psy2, | Minor | Chr1LGVI | ||||
| PsBB5-Psy, PsBB6-Psy | Minor | Chr7LG7 | ||||
| Viruses | ||||||
| Pea Seed-borne Mosaic Virus | PSbMV | sbm-1, sbm-3, sbm-4 | Major | Chr1LGVI | Complete | [114] |
| sbm-2 | Chr6LGII | |||||
| Pea Enation Mosaic Virus | PEMV | En | Major | Chr5LGIII | Complete | [121] |
| Pea common Mosaic | PMV | mo | Major | Chr6LGII | Complete | [119] |
| virus | ||||||
| Parasitic Plant | ||||||
| Broomrape | Orobanche crenata | Nºbr03-1, | Chr2LGI | Moderate | Partial | [102,103] |
| Nºbr03-2, PsOcr3 | Chr5LGIII | Minor | ||||
| Nºbr03-3, | Chr3LGV | Moderate | ||||
| Nºbr04, PsOcr2, | Chr1LGVI | Moderate | ||||
| PsOcr1 | Chr4LGIV | Major | ||||
| Insect Pest | ||||||
| Pea weevil | Bruchus pisorum | BpSI.I, | Chr2LGI | Moderate | Partial | [128] |
| BpSI.II, | Chr6LGII | |||||
| BpSI.III, BpLD.I | Chr4LGIV | |||||
| Pea aphid | Acyrthosiphon pisum | ApI, | Chr7LG7 | Minor | Partial | [131,133] |
| ApII, | Chr3LGV | Minor | ||||
| ApIII, | Chr5LGIII | Minor | ||||
| ApIV.1, ApIV.2 | Chr6LGII | Minor | ||||
| ApV.1, ApV.2, ApV.3, | Chr1LGVI | Major | ||||
| ApRVII | Chr7LG7 | Major | ||||
3. Germplasm Resources for Tolerance Traits
4. Generating Novel Variations for Pest and Disease Resistance
4.1. Induced Mutagenesis
4.2. Transgenic Technology
4.3. Gene Editing
5. Understanding the Genetic Makeup of Plant Traits Imparting Resistance
5.1. Phenotyping and Phenomics
5.2. Genetic Mapping
6. Genomic Selection
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rubiales, D.; González-Bernal, M.J.; Warkentin, T.; Bueckert, R.; Vaz Patto, M.C.; McPhee, K.; McGee, R.; Smýkal, P. Advances in pea breeding. In Achieving Sustainable Cultivation of Vegetables; Hochmuth, G., Ed.; Burleig Dodds Science Publishing Limited: Cambridge, UK, 2019; pp. 575–606. [Google Scholar]
- Smýkal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.H.; Flavell, A.J.; Ford, R.; Hýbl, M.; Macas, J.; Neumann, P.; et al. Pea (Pisum sativum L.) in the Genomic Era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://faostat.fao.org (accessed on 1 May 2023).
- Khan, T.N.; Timmerman-Vaughan, G.M.; Rubiales, D.; Warkentin, T.D.; Siddique, K.H.M.; Erskine, W.; Barbetti, M.J. Didymella pinodes and its management in field pea: Challenges and opportunities. Field Crops Res. 2013, 148, 61–77. [Google Scholar] [CrossRef]
- Prioul-Gervais, S.; Deniot, G.; Receveur, E.M.; Frankewitz, A.; Fourmann, M.; Rameau, C.; Pilet-Nayel, M.L.; Baranger, A. Candidate genes for quantitative resistance to Ascochyta pinodes in pea (Pisum sativum L.). Theor. Appl. Genet. 2007, 114, 971–984. [Google Scholar] [CrossRef]
- Carrillo, E.; Satovic, Z.; Aubert, G.; Boucherot, K.; Rubiales, D.; Fondevilla, S. Identification of quantitative trait loci and candidate genes for specific cellular resistance responses against Didymella pinodes in pea. Plant Cell Rep. 2014, 33, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Timmerman-Vaughan, G.M.; Moya, L.; Frew, T.J.; Murray, S.R.; Crowhurst, R. Ascochyta blight disease of pea (Pisum sativum L.): Defence-related candidate genes associated with QTL regions and identification of epistatic QTL. Theor. Appl. Genet. 2016, 129, 879–896. [Google Scholar] [CrossRef]
- Jha, A.B.; Gali, K.K.; Tar’an, B.; Warkentin, T.D. Fine mapping of QTLs for Ascochyta blight resistance in pea using heterogeneous inbred families. Front. Plant Sci. 2017, 8, 765. [Google Scholar] [CrossRef]
- Martins, L.B.; Balint-Kurti, P.; Reberg-Horton, S.C. Genome-wide association study for morphological traits and resistance to Peryonella pinodes in the USDA pea single plant plus collection. G3 Genes|Genomes|Genet. 2022, 12, jkac168. [Google Scholar] [CrossRef]
- Lee, R.C.; Grime, C.R.; O’Driscoll, K.; Khentry, Y.; Farfan-Caceres, L.M.; Tahghighi, H.; Kamphuis, L.G. Field Pea (Pisum sativum) germplasm screening for seedling Ascochyta Blight resistance and genome-wide association studies reveal loci associated with resistance to Peyronellaea pinodes and Ascochyta koolunga. Phytopathology 2023, 113, 265–276. [Google Scholar] [CrossRef]
- Boutet, G.; Lavaud, C.; Lesné, A.; Miteul, H.; Pilet-Nayel, M.-L.; Andrivon, D.; Lejeune-Hénaut, I.; Baranger, A. Five regions of the pea genome co-control partial resistance to D. pinodes, tolerance to frost, and some architectural or phenological traits. Genes 2023, 14, 1399. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Fondevilla, S. Future prospects for ascochyta blight resistance breeding in cool season food legumes. Front. Plant Sci. 2012, 3, 27. [Google Scholar] [CrossRef]
- Castillejo, M.A.; Susín, R.; Madrid, E.; Fernández-Aparicio, M.; Jorrín, J.V.; Rubiales, D. Two-dimensional gel electrophoresis-based proteomic analysis of the Medicago truncatula-rust (Uromyces striatus) interaction. Ann. Appl. Biol. 2010, 157, 243–257. [Google Scholar] [CrossRef]
- Kiba, A.; Miyake, C.; Toyoda, K.; Ichinose, Y.; Yamada, T.; Shiraishi, T. Superoxide generation in extracts from isolated plant cell walls is regulated by fungal signal molecules. Phytopathology 1997, 87, 846–852. [Google Scholar] [CrossRef]
- Carrillo, E.; Rubiales, D.; Pérez-de-Luque, A.; Fondevilla, S. Characterization of mechanisms of resistance against Didymella pinodes in Pisum spp. Eur. J. Plant Pathol. 2013, 135, 761–767. [Google Scholar] [CrossRef]
- Tran, H.S.; You, M.P.; Barbetti, M.J. Expression of defence-related genes in stems and leaves of resistant and susceptible field pea (Pisum sativum) during infection by Phoma koolunga. Plant Pathol. 2018, 67, 156–166. [Google Scholar] [CrossRef]
- Fondevilla, S.; Küster, H.; Krajinski, F.; Cubero, J.I.; Rubiales, D. Identification of genes differentially expressed in a resistant reaction to Mycosphaerella pinodes in pea using microarray technology. BMC Genom. 2011, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Osuna, D.; Colville, L.; Lorenzo, O.; Graeber, K.; Küster, H.; Leubner-Metzger, G.; Kranner, I. Transcriptome-Wide Mapping of Pea Seed Ageing Reveals a Pivotal Role for Genes Related to Oxidative Stress and Programmed Cell Death. PLoS ONE 2013, 8, e78471. [Google Scholar] [CrossRef] [PubMed]
- Fondevilla, S.; Rotter, B.; Krezdorn, N.; Jüngling, R.; Winter, P.; Rubiales, D. Identification of genes involved in resistance to Didymella pinodes in pea by deepSuperSAGE transcriptome profiling. Plant Mol. Biol. Rep. 2014, 32, 258–269. [Google Scholar] [CrossRef]
- Fondevilla, S.; Krezdorn, N.; Rubiales, D.; Rotter, B.M.; Winter, P. Transcriptomic analysis in a whole pea recombinant inbred line population segregating for resistance to Peyronellaea pinodes identifies the key factors and expressional markers for resistance to this pathogen. Sci. Rep. 2022, 12, 18159. [Google Scholar] [CrossRef]
- Castillejo, M.A.; Fondevilla, S.; Fuentes-Almagro, C.; Rubiales, D. Quantitative analysis of target peptides related to resistance against Ascochyta blight (Peyronellaea pinodes) in pea. J. Prot. Res. 2020, 19, 1000–1012. [Google Scholar] [CrossRef]
- Fondevilla, S.; Rubiales, D. Powdery mildew control in pea. A review. Agron. Sustain. Devel. 2012, 32, 401–409. [Google Scholar] [CrossRef]
- Rana, C.; Sharma, A.; Rathour, R.; Bansuli; Banyal, D.K.; Rana, R.S.; Sharma, P. In vivo and in vitro validation of powdery mildew resistance in garden pea genotypes. Sci. Rep. 2023, 13, 2243. [Google Scholar] [CrossRef] [PubMed]
- Devi, J.; Mishra, G.P.; Sagar, V.; Kaswan, V.; Dubey, R.K.; Singh, P.M.; Sharma, S.K.; Behera, T.K. Gene-Based resistance to Erysiphe species causing powdery mildew disease in peas (Pisum sativum L.). Genes 2022, 13, 316. [Google Scholar] [CrossRef] [PubMed]
- Fondevilla, S.; Carver, T.L.W.; Moreno, M.T.; Rubiales, D. Macroscopic and histological characterisation of genes er1 and er2 for powdery mildew resistance in pea. Eur. J. Plant Pathol. 2006, 115, 309–321. [Google Scholar] [CrossRef]
- Iglesias-García, R.; Rubiales, D.; Fondevilla, S. Penetration resistance to Erysiphe pisi in pea mediated by er1 gene is associated with protein cross-linking but not with callose apposition or hypersensitive response. Euphytica 2015, 201, 381–387. [Google Scholar] [CrossRef]
- Mohapatra, C.; Chand, R.; Navathe, S.; Sharma, S. Histo-chemical and biochemical analysis reveals association of er1 mediated powdery mildew resistance and redox balance in pea. Plant Physiol. Biochem. 2016, 106, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Humphry, M.; Reinstaedler, A.; Ivanov, S.; Bisseling, T.O.N.; Panstruga, R. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol. Plant Pathol. 2011, 12, 866–878. [Google Scholar] [CrossRef]
- Rispail, N.; Rubiales, D. Genome-wide identification and comparison of legume MLO gene family. Sci. Rep. 2016, 6, 32673. [Google Scholar] [CrossRef]
- Sun, S.; Deng, D.; Wu, W.; He, Y.; Luo, G.; Du, C.; Duan, C.; Zhu, Z. Molecular characterizations of the er1 alleles conferring resistance to Erysiphe pisi in three chinese pea (Pisum sativum L.) landraces. Int. J. Mol. Sci. 2022, 10, 12016. [Google Scholar] [CrossRef]
- Fondevilla, S.; Chattopadhyay, C.; Khare, N.; Rubiales, D. Erysiphe trifolii is able to overcome er1 and Er3, but not er2, resistance genes in pea. Eur. J. Plant Pathol. 2013, 136, 557–563. [Google Scholar] [CrossRef]
- Fondevilla, S.; González-Bernal, M.J.; Omri-BenYoussef, N.; Rubiales, D. Development of real-time PCR assays to quantify Erysiphe pisi and Erysiphe trifolii and its implementation for monitoring their relative prevalence in pea crops in Spain and Tunisia. Agronomy 2012, 12, 334. [Google Scholar] [CrossRef]
- Bhosle, S.M.; Marathe, N.; Bheri, M.; Makandar, R. Detection of putative pathogenicity and virulence genes of Erysiphe pisi using genome-wide in-silico search and their suppression by er2 mediated resistance in garden pea. Microb. Pathog. 2019, 136, 103680. [Google Scholar] [CrossRef] [PubMed]
- Bhosle, S.M.; Makandar, R. Comparative transcriptome of compatible and incompatible interaction of Erysiphe pisi and garden pea reveals putative defense and pathogenicity factors. FEMS Microbiol. Ecol. 2021, 97, fiab006. [Google Scholar] [CrossRef]
- Curto, M.; Camafeita, E.; López, J.A.; Maldonado, A.M.; Rubiales, D.; Jorrín, J.V. A proteomic approach to study pea (Pisum sativum) responses to powdery mildew (Erysiphe pisi). Proteomics 2006, 6, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Bheri, M.; Bhosle, S.M.; Makandar, R. Shotgun proteomics provides an insight into pathogenesis-related proteins using anamorphic stage of the biotroph, Erysiphe pisi pathogen of garden pea. Microbiol. Res. 2019, 222, 25–34. [Google Scholar] [CrossRef]
- Fondevilla, S.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of a new gene for resistance to Erysiphe pisi Syd. in pea. Breed. Sci. 2007, 57, 181–184. [Google Scholar] [CrossRef]
- Cobos, M.J.; Satovic, Z.; Rubiales, D.; Fondevilla, S. Er3 gene, conferring resistance to powdery mildew in pea, is located in pea LGIV. Euphytica 2018, 214, 203. [Google Scholar] [CrossRef]
- Davidson, J.A.; Krysinska-Kaczmarek, M.; Kimber, R.B.E.; Ramsey, M.D. Screening field pea germplasm for resistance to downy mildew (Peronospora viciae) and powdery mildew (Erysiphe pisi). Australas. Plant Pathol. 2004, 33, 413–417. [Google Scholar] [CrossRef]
- Amey, R.C.; Schleicher, T.; Slinn, J.; Lewis, M.; Macdonald, H.; Neill, S.J.; Spencer-Phillips, P.T.N. Proteomic analysis of a compatible interaction between Pisum sativum (pea) and the downy mildew pathogen Peronospora viciae. In The Downy Mildews-Genetics, Molecular Biology and Control; Lebeda, A., Spencer-Phillips, P.T.N., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 41–55. [Google Scholar] [CrossRef]
- Alemu, A.; Brantestam, A.K.; Chawade, A. Unraveling the genetic basis of key agronomic traits of wrinkled vining pea (Pisum sativum L.) for sustainable production. Front. Plant Sci. 2022, 13, 844450. [Google Scholar] [CrossRef]
- Chand, R.; Srivastava, C.P.; Singh, B.D.; Sarode, S.B. Identification and characterization of slow rusting components in pea (Pisum sativum L.). Genet. Res. Crop Evol. 2006, 53, 219–224. [Google Scholar] [CrossRef]
- Barilli, E.; Sillero, J.C.; Fernández-Aparicio, M.; Rubiales, D. Identification of resistance to Uromyces pisi (Pers.) Wint. in Pisum spp. germplasm. Field Crops Res. 2009, 114, 198–203. [Google Scholar] [CrossRef]
- Vijayalakshmi, S.; Yadav, K.; Kushwaha, C.; Sarode, S.B.; Srivastava, C.P.; Chand, R.; Singh, B.D. Identification of RAPD markers linked to the rust (Uromyces fabae) resistance gene in pea (Pisum sativum). Euphytica 2005, 144, 265–274. [Google Scholar] [CrossRef]
- Barilli, E.; Zatovic, S.; Rubiales, D.; Torres, A.M. Mapping of quantitative trait loci controlling partial resistance against rust incited by Uromyces pisi (Pers.) Wint. in a Pisum fulvum L. intraspecific cross. Euphytica 2010, 175, 151–159. [Google Scholar] [CrossRef]
- Barilli, E.; Cobos, M.J.; Carrillo, E.; Kilian, A.; Carlin, J.; Rubiales, D. A high-density integrated DArTseq SNP-based genetic map of Pisum fulvum and identification of QTLs controlling rust resistance. Front. Plant Sci. 2018, 9, 167. [Google Scholar] [CrossRef]
- Osuna-Caballero, S.; Rispail, N.; Barilli, E.; Rubiales, D. Identification and characterization of novel sources of resistance to rust caused by Uromyces pisi in Pisum spp. Plants 2022, 11, 2268. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Kushwaha, C.; Shikha, K.; Chand, R.; Mishra, G.P.; Dikshit, H.K.; Devi, J.; Aski, M.S.; Kumar, S.; Gupta, S.; et al. Rust (Uromyces viciae-fabae Pers. de-Bary) of pea (Pisum sativum L.): Present status and future resistance breeding opportunities. Genes 2023, 14, 374. [Google Scholar] [CrossRef]
- Barilli, E.; Rubiales, D.; Castillejo, M. Comparative proteomic analysis of BTH and BABA-induced resistance in pea (Pisum sativum) toward infection with pea rust (Uromyces pisi). J. Prot. 2012, 75, 5189–5205. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, C.; Chand, R.; Singh, A.K.; Kumar, M.; Srivastava, C.P. Differential induction of β-1,3-glucanase gene in expression of partial resistance to rust (Uromyces fabae (Pers.) de-Bary) in pea (Pisum sativum L.). Russ. J. Plant Physiol. 2018, 65, 697–701. [Google Scholar] [CrossRef]
- McPhee, K.E.; Tullu, A.; Kraft, J.M.; Muehlbauer, F.J. Resistance to fusarium wilt races 2 in the Pisum core collection. J. Am. Soc. Hort. Sci. 1998, 124, 28–31. [Google Scholar] [CrossRef]
- Bani, M.; Rubiales, D.; Rispail, N. A detailed evaluation method to identify sources of quantitative resistance to Fusarium oxysporum f. sp. pisi race 2 within a Pisum spp. germplasm collection. Plant Pathol. 2012, 61, 532–542. [Google Scholar] [CrossRef]
- Grajal-Martín, M.J.; Muehlbauer, F.J. Genomic location of the Fw gene for resistance to Fusarium wilt race 1 in peas. J. Hered. 2002, 93, 291–293. [Google Scholar] [CrossRef]
- Porter, L.D.; Kraft, J.M.; Grünwald, N.J. Release of pea germplasm with Fusarium resistance combined with desirable yield and anti-lodging traits. J. Plant Regist. 2014, 2, 137–139. [Google Scholar] [CrossRef]
- Coyne, C.J.; Inglis, D.A.; Whitehead, S.J.; McClendon, M.T.; Muehlbauer, F.J. Chromosomal location of Fwf, the Fusarium wilt race 5 resistance gene in Pisum sativum. Pisum Genet. 2000, 32, 20–22. [Google Scholar]
- McClendon, M.T.; Inglis, D.A.; McPhee, K.E.; Coyne, C.J. DNA markers linked to Fusarium wilt race 1 resistance in pea. J. Am. Soc. Hort. Sci. 2002, 127, 602–607. [Google Scholar] [CrossRef]
- Okubara, P.A.; Keller, K.E.; McClendon, M.T.; Inglis, D.A.; Mcphee, K.E.; Coyne, C.J. Y15_999fw, a dominant scar marker linked to the fusarium wilt race 1 (fw) resistance gene in pea. Pisum Genet. 2005, 37, 32–35. [Google Scholar]
- Jain, S.; Weeden, N.F.; Kumar, A.; Chittem, K.; McPhee, K. Functional codominant marker for selecting the Fw gene conferring resistance to fusarium wilt race 1 in pea. Crop Sci. 2015, 55, 2639–2646. [Google Scholar] [CrossRef]
- Neumann, S.; Xue, A.G. Reactions of field pea cultivars to four races of Fusarium oxysporum f. sp pisi. Can. J. Plant Sci. 2003, 83, 377–379. [Google Scholar] [CrossRef]
- Mc Phee, K.E.; Inglis, D.A.; Gundersen, B.; Coyne, C.J. Mapping QTL for Fusarium wilt race 2 partial resistance in pea (Pisum sativum). Plant Breed. 2012, 131, 300–306. [Google Scholar] [CrossRef]
- Bani, M.; Pérez-de-Luque, A.; Rubiales, D.; Rispail, N. Physical and chemical barriers in root tissues contribute to quantitative resistance to Fusarium oxysporum f. sp. pisi in pea. Front. Plant Sci. 2018, 9, 199. [Google Scholar] [CrossRef]
- Bani, M.; Cimmino, A.; Evidente, A.; Rubiales, D.; Rispail, N. Pisatin involvement in the variation of inhibition of Fusarium oxysporum f. sp. pisi spore germination by root exudates of Pisum spp. germplasm. Plant Pathol. 2018, 67, 1046–1054. [Google Scholar] [CrossRef]
- Catillejo, M.A.; Bani, M.; Rubiales, D. Understanding pea resistance mechanisms in response to Fusarium oxysporum through proteomic analysis. Phytochemistry 2015, 115, 44–58. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Huber, D.; Basnyat, P.; Zentner, R.P. Impact of agronomic practices on populations of Fusarium and other fungi in cereal and noncereal crop residues on the Canadian Prairies. Soil Tillage Res. 2008, 100, 60–71. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Coffman, V.A.; Kraft, J.M. Sources of partial resistance to Fusarium root rot in the Pisum core collection. Plant Dis. 2003, 87, 1197–1200. [Google Scholar] [CrossRef] [PubMed]
- Bodah, E.T.; Porter, L.D.; Chaves, B.; Dhingra, A. Evaluation of pea accessions and commercial cultivars for fusarium root rot resistance. Euphytica 2016, 208, 63–72. [Google Scholar] [CrossRef]
- Feng, J.; Hwang, R.; Chang, K.F.; Conner, R.L.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; McLaren, D.L.; Xue, A.G. Identification of microsatellite markers linked to quantitative trait loci controlling resistance to Fusarium root rot in field pea. Can. J. Plant Sci. 2013, 91, 199–204. [Google Scholar] [CrossRef]
- Coyne, C.J.; Pilet-Nayel, M.; McGee, R.J.; Porter, L.D.; Smýkal, P.; Grünwald, N.J. Identification of QTL controlling high levels of partial resistance to Fusarium solani f. sp. pisi in pea. Plant Breed. 2015, 134, 446–453. [Google Scholar] [CrossRef]
- Coyne, C.J.; Porter, L.D.; Boutet, G.; Ma, Y.; McGee, R.J.; Lesné, A.; Baranger, A.; Pilet-Nayel, M.-L. Confirmation of Fusarium root rot resistance QTL Fsp-Ps 2.1 of pea under controlled conditions. BMC Plant Biol. 2019, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Williamson-Benavides, B.A.; Sharpe, R.M.; Nelson, G.; Bodah, E.T.; Porter, L.D.; Dhingra, A. Identification of root rot resistance QTLs in pea using Fusarium solani f. sp. pisi-responsive differentially expressed genes. Front. Genet. 2021, 12, 629267. [Google Scholar] [CrossRef]
- Wu, L.; Fredua-Agyeman, R.; Strelkov, S.E.; Chang, K.-F.; Hwang, S.-F. Identification of quantitative trait loci associated with partial resistance to fusarium root rot and wilt caused by Fusarium graminearum in field pea. Front. Plant Sci. 2022, 12, 784593. [Google Scholar] [CrossRef]
- Wu, L.; Fredua-Agyeman, R.; Strelkov, S.E.; Chang, K.-F.; Hwang, S.-F. Identification of novel genes associated with partial resistance to Aphanomyces root rot in field pea by BSR-Seq analysis. Int. J. Mol. Sci. 2022, 23, 9744. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.; Muehlbauer, F.; McGee, R.; Kraft, J.; Baranger, A.; Coyne, C. Quantitative trait loci for partial resistance to aphanomyces root rot in pea. Theor. Appl. Genet. 2002, 106, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Lavaud, C.; Lesné, A.; Piriou, C.; Le Roy, G.; Boutet, G.; Moussart, A.; Poncet, C.; Delourme, R.; Baranger, A.; Pilet-Nayel, M.-L. Validation of QTL for resistance to Aphanomyces euteiches in different pea genetic backgrounds using near-isogenic lines. Theor. Appl. Genet. 2015, 128, 2273–2288. [Google Scholar] [CrossRef] [PubMed]
- Desgroux, A.; L’Anthoëne, V.; Roux-Duparque, M.; Rivière, J.-P.; Aubert, G.; Tayeh, N.; Moussart, A.; Mangin, P.; Vetel, P.; Piriou, C.; et al. Genome-wide association mapping of partial resistance to Aphanomyces euteiches in pea. BMC Genom. 2016, 17, 124. [Google Scholar] [CrossRef]
- Desgroux, A.; Baudais, V.N.; Aubert, V.; Le Roy, G.; De Larambergue, H.; Miteul, H.; Aubert, G.; Boutet, G.; Duc, G.; Baranger, A.; et al. Comparative genome-wide-association mapping identifies common loci controlling root system architecture and resistance to Aphanomyces euteiches in pea. Front. Plant Sci. 2018, 8, 2195. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, M.; Fariello, M.I.; Navier, H.; Hajri, A.; Badis, Y.; Miteul, H.; Samac, D.A.; Dumas, B.; Baranger, A.; Jacquet, C.; et al. A local score approach improves GWAS resolution and detects minor QTL: Application to Medicago truncatula quantitative disease resistance to multiple Aphanomyces euteiches isolates. Heredity 2019, 123, 517–531. [Google Scholar] [CrossRef]
- Hosseini, S.; Elfstrand, M.; Heyman, F.; Funck Jensen, D.; Karlsson, M. Deciphering common and specific transcriptional immune responses in pea towards the oomycete pathogens Aphanomyces euteiches and Phytophthora pisi. BMC Genom. 2015, 16, 627. [Google Scholar] [CrossRef]
- Lavaud, C.; Baviere, M.; Le Roy, G.; Hervé, M.R.; Moussart, A.; Delourme, R.; Pilet-Nayel, M.L. Single and multiple resistance QTL delay symptom appearance and slow down root colonization by Aphanomyces euteiches in pea near isogenic lines. BMC Plant Biol. 2016, 16, 166. [Google Scholar] [CrossRef]
- Quillévéré-Hamard, A.; Le Roy, G.; Moussart, A.; Baranger, A.; Andrivon, D.; Pilet-Nayel, M.L.; Le May, C. Genetic and pathogenicity diversity of Aphanomyces euteiches populations from pea-growing regions in France. Front. Plant Sci. 2018, 9, 1673. [Google Scholar] [CrossRef]
- Sharma-Poudyal, D.; Paulitz, T.C.; Porter, L.D.; Sharma-Poudyal, D. Characterization and pathogenicity of Rhizoctonia and Rhizoctonia-like spp. from pea crops in the Columbia basin of Oregon and Washington. Plant Dis. 2015, 99, 604–613. [Google Scholar] [CrossRef]
- Muehlbauer, F.J.; Kraft, J.M. Evidence of heritable resistance to Fusarium solani f. sp. pisi and Pythium ultimum in peas. Crop Sci. 1973, 13, 34–36. [Google Scholar] [CrossRef]
- Schroeder, K.L.; Martin, F.N.; de Cock, A.W.A.M.; Lévesque, C.A.; Spies, C.F.J.; Okubara, P.A.; Paulitz, T.C. Molecular detection and quantification of Pythium species: Evolving taxonomy, new tools, and challenges. Plant Dis. 2013, 97, 4–20. [Google Scholar] [CrossRef]
- Harveson, R.M.; Pasche, J.S.; Porter, L.; Chen, W.; Burrows, M. Compendium of Pea Diseases and Pests, 3rd ed.; American Phytopathological Society: St. Paul, MN, USA, 2021; ISBN 978-0-89054-655-0. [Google Scholar]
- Veronico, P.; Melillo, M.T.; Saponaro, C.; Leonetti, P.; Picardi, E.; Jones, J.T. A polygalacturonase-inhibiting protein with a role in pea defence against the cyst nematode Heterodera goettingiana. Mol. Plant Pathol. 2011, 12, 275–287. [Google Scholar] [CrossRef]
- Sharma, A.; Haseeb, A.; Abuzar, S. Screening of field pea (Pisum sativum) selections for their reactions to root-knot nematode (Meloidogyne incognita). J. Zhejiang Univ. Sci. B 2006, 7, 209–214. [Google Scholar] [CrossRef]
- Thompson, J.P.; Reen, R.A.; Clewett, T.G.; Sheedy, J.G.; Kelly, A.M.; Gogel, B.J.; Knights, E.J. Hybridisation of Australian chickpea cultivars with wild Cicer spp. increases resistance to root-lesion nematodes (Pratylenchus thornei and P. neglectus). Australas. Plant Pathol. 2011, 40, 601–611. [Google Scholar] [CrossRef]
- Upadhaya, A.; Yan, G.; Pasche, J. Reproduction ability and growth effect of pin nematode, Paratylenchus nanus, with selected field pea cultivars. Plant Dis. 2019, 103, 2520–2526. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in parasitic weeds management in legume crops. A review. Agron. Sustain. Dev. 2012, 32, 433–449. [Google Scholar] [CrossRef]
- Rubiales, D.; Moreno, M.T.; Sillero, J.C. Search for resistance to crenate broomrape (Orobanche crenata Forsk.) in pea germplasm. Genet. Resour. Crop Evol. 2005, 52, 853–861. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Pérez-de-Luque, A.; Castillejo, M.A.; Prats, E.; Sillero, J.C.; Rispail, N.; Fondevilla, S. Breeding approaches for crenate broomrape (Orobanche crenata Forsk.) management in pea (Pisum sativum L.). Pest Manag. Sci. 2009, 65, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de-Luque, A.; Jorrín, J.V.; Cubero, J.I.; Rubiales, D. Orobanche crenata resistance and avoidance in pea (Pisum spp.) operate at different developmental stages of the parasite. Weed Res. 2005, 45, 379–387. [Google Scholar] [CrossRef]
- Rispail, N.; Dita, M.A.; González-Verdejo, C.; Pérez-de-Luque, A.; Castillejo, M.A.; Prats, E.; Román, B.; Jorrín, J.V.; Rubiales, D. Plant resistance to parasitic plants: Molecular approaches to an old foe. New Phytol. 2007, 173, 703–712. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Moreno, M.T.; Rubiales, D. Host plant resistance against broomrapes (Orobanche spp.): Defence reactions and mechanisms of resistance. Ann. Appl. Biol. 2008, 152, 131–141. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Rubiales, D. Differential response of pea (Pisum sativum) to Orobanche crenata, Orobanche foetida and Phelipanche aegyptiaca. Crop Prot. 2011, 31, 27–30. [Google Scholar] [CrossRef]
- Fondevilla, S.; Flores, F.; Emeran, A.A.; Kharrat, M.; Rubiales, D. High productivity of dry pea genotypes resistant to crenate broomrape in Mediterranean environments. Agron. Sustain. Dev. 2017, 37, 61. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Fernández-Aparicio, M. Development of pea breeding lines with resistance to Orobanche crenata derived from pea landraces and wild Pisum spp. Agronomy 2021, 11, 36. [Google Scholar] [CrossRef]
- Rubiales, D.; Osuna-Caballero, S.; González-Bernal, M.J.; Cobos, M.J.; Flores, F. Pea breeding lines adapted to autumn sowings in broomrape prone Mediterranean environments. Agronomy 2021, 11, 769. [Google Scholar] [CrossRef]
- Rubiales, D. Can we breed for durable resistance to broomrapes? Phytopathol. Mediterr. 2018, 57, 170–185. [Google Scholar]
- Wohor, Z.O.; Rispail, N.; Rubiales, D. Characterization of a Pisum spp. germplasm for resistance to Fusarium oxysporum and Orobanche crenata. In Proceedings of the 9th International Conference on Legume Genome and Genetics, Dijon, France, 13–17 May 2019. [Google Scholar]
- Valderrama, M.R.; Román, B.; Satovic, Z.; Rubiales, D.; Cubero, J.I.; Torres, A.M. Locating quantitative trait loci associated with Orobanche crenata resistance in pea. Weed Res. 2004, 44, 323–328. [Google Scholar] [CrossRef]
- Fondevilla, S.; Fernández-Aparicio, M.; Satovic, Z.; Emeran, A.A.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of quantitative trait loci for specific mechanisms of resistance to Orobanche crenata Forsk. in pea (Pisum sativum L.). Mol. Breed. 2010, 25, 259–272. [Google Scholar] [CrossRef]
- Delvento, C.; Arcieri, F.; Marcotrigiano, A.R.; Guerriero, M.; Fanelli, V.; Dellino, M.; Curci, P.L.; Bouwmeester, H.; Lotti, C.; Ricciardi, L.; et al. High-density linkage mapping and genetic dissection of resistance to broomrape (Orobanche crenata Forsk.) in pea (Pisum sativum L.). Front. Plant Sci. 2023, 14, 1216297. [Google Scholar] [CrossRef]
- Dita, M.A.; Die, J.V.; Román, B.; Krajinski, F.; Küster, H.; Moreno, M.T.; Cubero, J.I.; Rubiales, D. Gene expression profiling of Medicago truncatula roots in response to the parasitic plant Orobanche crenata. Weed Res. 2009, 49, 66–80. [Google Scholar] [CrossRef]
- Castillejo, M.A.; Maldonado, A.M.; Dumas-Gaudot, E.; Fernández-Aparicio, M.; Susín, R.; Rubiales, D.; Jorrín, J.V. Differential expression proteomics to investigate responses and resistance to Orobanche crenata in Medicago truncatula. BMC Genom. 2009, 10, 294. [Google Scholar] [CrossRef]
- Castillejo, M.A.; Amiour, N.; Dumas-Gaudot, E.; Rubiales, D.; Jorrín, J.V. A proteomic approach to studying plant response to crenate broomrape (Orobanche crenata) in pea (Pisum sativum). Phytochemistry 2004, 65, 1817–1828. [Google Scholar] [CrossRef]
- Rodda, M.S.; Kant, P.; Lindbeck, K.D.; Gnanasambandam, A.; Hollaway, G.J. A high-throughput glasshouse-based screening method to evaluate bacterial blight resistance in field pea (Pisum sativum). Australas. Plant Pathol. 2015, 44, 515–526. [Google Scholar] [CrossRef]
- Hunter, P.J.; Ellis, N.; Taylor, J.D. Association of dominant loci for resistance to Pseudomonas syringae pv. pisi with linkage groups II, VI and VII of Pisum sativum. Theor. Appl. Genet. 2001, 103, 129–135. [Google Scholar] [CrossRef]
- Martín-Sanz, A.; de la Vega, M.P.; Murillo, J.; Caminero, C. Genetic, biochemical and pathogenic diversity of Pseudomonas syringae pv. pisi strains. Plant Pathol. 2012, 61, 1063–1072. [Google Scholar] [CrossRef]
- Martín-Sanz, A.; Aparicio, T.; Santana, J.C.; García, P.; Winter, P.; Caminero, C.; de la Vega, M.P. Mapping genes for resistance to bacterial blight (Pseudomonas syringae pv. pisi) in pea and identification of genes involved in resistance by DeepsuperSAGE transcriptome profiling. Euphytica 2016, 210, 375–392. [Google Scholar] [CrossRef]
- Sudheesh, S.; Rodda, M.; Kennedy, P.; Verma, P.; Leonforte, A.; Cogan, N.O.I.; Materne, M.; Forster, J.W.; Kaur, S. Construction of an integrated linkage map and trait dissection for bacterial blight resistance in field pea (Pisum sativum L.). Mol. Breed. 2015, 35, 185. [Google Scholar] [CrossRef]
- Fondevilla, S.; Martín-Sanz, A.; Satovic, Z.; Fernández-Romero, M.D.; Rubiales, D.; Caminero, C. Identification of quantitative trait loci involved in resistance to Pseudomonas syringae pv. syringae in pea (Pisum sativum L.). Euphytica 2012, 186, 805–812. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Taylor, J.D. Resistance to bacterial blight (Pseudomonas syringae pv. pisi) in Spanish pea (Pisum sativum) landraces. Euphytica 2001, 118, 305–311. [Google Scholar] [CrossRef]
- Gao, Z.; Eyers, S.; Thomas, C.; Ellis, N.; Maule, A. Identification of markers tightly linked to sbm recessive genes for resistance to Pea seed-borne mosaic virus. Theor. Appl. Genet. 2004, 109, 488–494. [Google Scholar] [CrossRef]
- Smýkal, P.; Šafářová, D.; Navrátil, M.; Dostalová, R. Marker assisted pea breeding: eIF4E allele specific markers to Pea seed-borne mosaic virus (PSbMV) resistance. Mol. Breed. 2010, 26, 425–438. [Google Scholar] [CrossRef]
- Hjulsager, C.K.; Lund, O.S.; Johansen, I.E. A new pathotype of Pea seed-borne mosaic virus explained by properties of the P3-6k1 and viral genome-linked protein (VPg)-coding regions. Mol. Plant-Micr. Inter. 2002, 15, 169–171. [Google Scholar] [CrossRef]
- Swisher Grimm, K.D.; Porter, L.D. Development and validation of KASP markers for the identification of Pea seed-borne mosaic virus pathotype P1 resistance in Pisum sativum. Plant Dis. 2020, 104, 1824–1830. [Google Scholar] [CrossRef] [PubMed]
- Swisher Grimm, K.D.; Porter, L.D. KASP markers reveal established and novel sources of resistance to Pea Seedborne Mosaic Virus in pea genetic resources. Plant Dis. 2021, 105, 2503–2508. [Google Scholar] [CrossRef]
- Dirlewanger, E.; Isaac, P.; Ranade, S.; Belajouza, M.; Cousin, R.; Devienne, D. Restriction fragment length polymorphism analysis of loci associated with disease resistance genes and developmental traits in Pisum sativum L. Theor. Appl. Genet. 1994, 88, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Baggett, J.R.; Hampton, R.O. Inheritance of viral bean leaf roll tolerance in peas. J. Am. Soc. Hort. Sci. 1991, 116, 728–731. [Google Scholar] [CrossRef]
- Jain, S.; Weeden, N.F.; Porter, L.D.; Eigenbrode, S.D.; McPhee, K. Finding Linked Markers to En for Efficient Selection of Pea Enation Mosaic Virus Resistance in Pea. Crop Sci. 2013, 53, 2392–2399. [Google Scholar] [CrossRef]
- Teshome, A.; Mendesil, E.; Geleta, M.; Andargie, D.; Anderson, P.; Rämert, B.; Seyoum, E.; Hillbur, Y.; Dagne, K.; Bryngelsson, T. Screening the primary gene pool of field pea (Pisum sativum L. subsp. sativum) in Ethiopia for resistance against pea weevil (Bruchus pisorum L.). Genet. Res. Crop Evol. 2015, 62, 525–538. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Carrillo-Perdomo, E.; Flores, F.; Rubiales, D. Identification and multi-environment validation of resistance to pea weevil (Bruchus pisorum) in Pisum germplasm. J. Pest Sci. 2017, 91, 505–514. [Google Scholar] [CrossRef]
- Aryamanesh, N.; Zeng, Y.; Byrne, O.; Hardie, D.C.; Al-Subhi, A.M.; Khan, T.; Siddique, K.H.M.; Yan, G. Identification of genome regions controlling cotyledon, pod wall/seed coat and pod wall resistance to pea weevil through QTL mapping. Theor. Appl. Genet. 2014, 127, 489–497. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Rubiales, D. Flower and pod source influence on pea weevil (Bruchus pisorum) oviposition capacity and preference. Front. Plant Sci. 2019, 10, 491. [Google Scholar] [CrossRef]
- Byrne, O.M.; Hardie, D.C.; Khan, T.N.; Speijers, J.; Yan, G. Genetic analysis of pod and seed resistance to pea weevil in a Pisum sativum × P. fulvum interspecific cross. Australian J. Agric. Res. 2018, 59, 854–862. [Google Scholar] [CrossRef]
- Sari, H.; Sari, D.; Eker, T.; Aydinoglu, B.; Canci, H.; Ikten, C.; Gokturk, R.S.; Zeybek, A.; Bakir, M.; Smykal, P.; et al. Inheritance and expressivity of neoplasm trait in crosses between the domestic pea (Pisum sativum subsp. sativum) and tall wild pea (Pisum sativum subsp. elatius). Agronomy 2020, 10, 1869. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Barilli, E.; Cobos, M.J.; Kilian, A.; Carling, J.; Rubiales, D. Identification of quantitative trait loci (QTL) controlling resistance to pea weevil (Bruchus pisorum) in a high-density integrated DArTseq SNP-based genetic map of pea. Sci. Rep. 2020, 10, 33. [Google Scholar] [CrossRef]
- Ali, K.; van den Louw, S.M.; Swart, W.J. Components and mechanisms of resistance in selected field pea Pisum sativum lines to the pea aphid Acyrthosiphon pisum (Homoptera: Aphididae). Int. J. Trop. Insect. Sci. 2005, 25, 114–121. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Rubiales, D. Identification and characterization of antixenosis and antibiosis to pea aphid (Acyrthosiphum pisum) in Pisum spp. germplasm. Ann. Appl. Biol. 2018, 3, 268–281. [Google Scholar] [CrossRef]
- Ollivier, R.; Glory, I.; Cloteau, R.; Le Gallic, J.-F.; Denis, G.; Morlière, S.; Miteul, H.; Rivière, J.-P.; Lesné, A.; Klein, A.; et al. A major-effect genetic locus, ApRVII, controlling resistance against both adapted and non-adapted aphid biotypes in pea. Theor. Appl. Genet. 2022, 135, 1511–1528. [Google Scholar] [CrossRef]
- Rahman, M.M.; Porter, L.D.; Ma, Y.; Coyne, C.J.; Zheng, P. Resistance in pea (Pisum sativum) genetic resources to the pea aphid, Acyrthosiphon pisum. Entomol. Exp. Appl. 2023, 171, 435–448. [Google Scholar] [CrossRef]
- Barilli, E.; Carrillo-Perdomo, E.; Cobos, M.J.; Kilian, A.; Carling, J.; Rubiales, D. Identification of potential candidate genes controlling pea aphid tolerance in a Pisum fulvum high-density integrated DArTseq SNP-based genetic map. Pest Manag. Sci. 2020, 76, 1731–1742. [Google Scholar] [CrossRef]
- Carrillo, E.; Rubiales, D.; Castillejo, M.A. Proteomic analysis of pea (Pisum sativum L.) response during compatible and incompatible interactions with the pea aphid (Acyrthosiphon pisum H.). Plant Mol. Biol. Rep. 2014, 32, 697–718. [Google Scholar] [CrossRef]
- Maxted, N.; Ambrose, M. Peas (Pisum L.). Plant Genetic Resources of Legumes in the Mediterranean; Maxted, N., Bennett, S.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 181–190. [Google Scholar]
- Weeden, N.F. Domestication of Pea (Pisum sativum L.). The case of the abyssinian pea. Front. Plant Sci. 2018, 9, 515. [Google Scholar] [CrossRef]
- Rispail, N.; Wohor, O.Z.; Osuna-Caballero, S.; Barilli, E.; Rubiales, D. Genetic diversity and population structure of a wide Pisum spp. core collection. Int. J. Mol. Sci. 2023, 24, 2470. [Google Scholar] [CrossRef]
- Bogdanova, V.S.; Kosterin, O.E.; Yadrikhinskiy, A.K. Wild peas vary in their cross-compatibility with cultivated pea (Pisum sativum subsp. sativum L.) depending on alleles of a nuclear–cytoplasmic incompatibility locus. Theor. Appl. Genet. 2014, 127, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Kosterin, O.E.; Bogdanova, V.S. Reciprocal compatibility within the genus Pisum L. as studied in F1 hybrids: 4. Crosses within P. sativum L. subsp. elatius (Bieb.) Aschers. et Graebn. Genet. Resour. Crop Evol. 2021, 68, 2565–2590. [Google Scholar] [CrossRef]
- Smykal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef]
- Thudi, M.; Palakurthi, R.; Schnable, J.C.; Chitikineni, A.; Dreisigacker, S.; Mace, E.; Srivastava, R.K.; Satyavathi, C.T.; Odeny, D.; Tiwari, V.K.; et al. Genomic resources in plant breeding for sustainable agriculture. J. Plant Physiol. 2021, 257, 153351. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Annicchiarico, P.; Vaz-Patto, M.C.; Julier, B. Legume breeding for the agroecological transition of global agri-food systems: A European perspective. Front. Plant Sci. 2021, 12, 782574. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Gudi, S.; Amandeep, U.; Upadhyay, P.; Shekhawat, P.K.; Nayak, G.; Goyal, L.; Kumar, D.; Kumar, P.; Kamboj, A.; et al. Unlocking the hidden variation from wild repository for accelerating genetic gain in legumes. Front. Plant Sci. 2022, 13, 1035878. [Google Scholar] [CrossRef]
- Parida, S.K.; Mondal, N.; Yadav, R.; Vishwakarma, H.; Rana, J.C. Mining legume germplasm for genetic gains: An Indian perspective. Front. Genet. 2023, 14, 996828. [Google Scholar] [CrossRef]
- Durieu, P.; Ochatt, S.J. Efficient intergeneric fusion of pea (Pisum sativum L.) and grass pea (Lathyrus sativus L.) protoplasts. J. Exp. Bot. 2020, 51, 1237–1242. [Google Scholar]
- Sharma, A.; Plaha, P.; Rathour, R.; Katoch, V.; Singh, Y.; Khalsa, G.S. Induced mutagenesis for improvement of garden pea. Int. J. Veg. Sci. 2009, 16, 60–72. [Google Scholar] [CrossRef]
- Sinjushin, A.; Semenova, E.; Vishnyakova, M. Usage of morphological mutations for improvement of a garden pea (Pisum sativum): The experience of breeding in russia. Agronomy 2022, 12, 544. [Google Scholar] [CrossRef]
- Deng, D.; Sun, S.; Wu, W.; Xiang, C.; Duan, C.; Yu, D.; Wu, X.; Zhu, Z. Disease resistance and molecular variations in irradiation induced mutants of two pea cultivars. Int. J. Mol. Sci. 2022, 23, 8793. [Google Scholar] [CrossRef]
- Pereira, G.; Marques, C.; Ribeiro, R.; Formiga, S.; Dâmaso, M.; Tavares Sousa, M.; Farinhó, M.; Leitão, J.M. Identification of DNA markers linked to an induced mutated gene conferring resistance to powdery mildew in pea (Pisum sativum L.). Euphytica 2009, 171, 327–335. [Google Scholar] [CrossRef]
- Santo, T.; Rashkova, M.; Alabaça, C.; Leitão, J. The ENU-induced powdery mildew resistant mutant pea (Pisum sativum L.) lines S(er1mut1) and F(er1mut2) harbour early stop codons in the PsMLO1 gene. Mol. Breed. 2013, 32, 723–727. [Google Scholar] [CrossRef]
- Sharma, A.; Rathour, R.; Plaha, P.; Katoch, V.; Khalsa, G.S.; Patial, V.; Singh, Y.; Pathania, N.K. Induction of Fusarium wilt (Fusarium oxysporum f. sp. pisi) resistance in garden pea using induced mutagenesis and in vitro selection techniques. Euphytica 2010, 173, 345–356. [Google Scholar] [CrossRef]
- Gritton, E.T.; Hagedorn, D.J. Mutation breeding for pea (Pisum sativum L.) root rot resistance. Agron. Abstr. 1979, 62. [Google Scholar]
- Kurowska, M.; Daszkowska-Golec, A.; Gruszka, D.; Marzec, M.; Szurman, M.; Szarejko, I.; Maluszynski, M. TILLING: A shortcut in functional genomics. J. Appl. Genet. 2011, 52, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Domoney, C.; Knox, M.; Moreau, C.; Ambrose, M.; Palmer, S.; Smith, P.; Christodoulou, V.; Isaac, P.G.; Hegarty, M.; Blackmore, T.; et al. Exploiting a fast neutron mutant genetic resource in Pisum sativum (pea) for functional genomics. Funct. Plant Biol. 2013, 40, 1261–1270. [Google Scholar] [CrossRef]
- Ludvíková, M.; Griga, M. Pea transformation: History, current status and challenges. Czech J. Genet. Plant Breed. 2022, 58, 127–161. [Google Scholar] [CrossRef]
- Negawo, A.T.; Aftabi, M.; Jacobsen, H.-J.; Altosaar, I.; Hassan, F.S. Insect resistant transgenic pea expressing cry1Ac gene product from Bacillus thuringiensis. Biol. Contr. 2013, 67, 293–300. [Google Scholar] [CrossRef]
- Timmerman-Vaughan, G.M.; Pither-Joyce, M.D.; Cooper, P.A.; Russell, A.C.; Goulden, D.S.; Butler, R.; Grant, J.E. Partial resistance of transgenic peas to alfalfa mosaic virus under greenhouse and field conditions. Crop Sci. 2001, 41, 846–853. [Google Scholar] [CrossRef]
- Jones, A.L.; Johansen, E.I.; Bean, S.J.; Bach, I.; Maule, A.L. Specificity of resistance to pea seed-borne mosaic potyvirus in transgenic peas expressing the viral replicase (NIb) gene. J. Gen. Vir. 1998, 79, 3129–3137. [Google Scholar] [CrossRef] [PubMed]
- Chowrira, G.M.; Cavileer, T.D.; Gupta, S.K.; Lurquin, P.F.; Berger, P.H. Coat protein resistance to pea enation mosaic virus in transgenic Pisum sativum L. Transgenic Res. 1998, 7, 265–271. [Google Scholar] [CrossRef]
- Kahlon, J.G.; Jacobsen, H.J.; Chatterton, S.; Hassan, F.; Bowness, R.; Hall, L.M. Lack of efficacy of transgenic pea (Pisum sativum L.) stably expressing antifungal genes against Fusarium spp. in three years of confined field trials. GM Crops Food 2018, 9, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.L.; Schroeder, H.E.; Bateman, K.S.; Chrispeels, M.J.; Armstrong, E.; Higgins, T.J. Bean alpha-amylase inhibitor 1 in transgenic peas (Pisum sativum) provides complete protection from pea weevil (Bruchus pisorum) under field conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 3820–3825. [Google Scholar] [CrossRef]
- Prescott, V.E.; Campbell, P.M.; Moore, A.; Mattes, J.; Rothenberg, M.E.; Foster, P.S.; Higgins, T.J.V.; Hogan, S.P. Transgenic expression of bean alpha amylase inhibitor in pea results in altered structure and immunogenicity. J. Agric. Food Chem. 2005, 16, 9023–9030. [Google Scholar] [CrossRef]
- Pandey, P.K.; Bhowmik, P.; Kagale, S. Optimized methods for random and targeted mutagenesis in field pea (Pisum sativum L.). Front. Plant Sci. 2022, 13, 995542. [Google Scholar] [CrossRef]
- Bhowmik, P.; Konkin, D.; Polowick, P.; Hodgins, C.L.; Subedi, M.; Xiang, D.; Yu, B.; Patterson, N.; Rajagopalan, N.; Babic, V.; et al. CRISPR/Cas9 gene editing in legume crops: Opportunities and challenges. Legume Sci. 2021, 3, e96. [Google Scholar] [CrossRef]
- Li, G.; Liu, R.; Xu, R.; Varshney, R.K.; Ding, H.; Li, M.; Yan, X.; Huang, S.; Li, J.; Wang, D.; et al. Development of an Agrobacterium-mediated CRISPR/Cas9 system in pea (Pisum sativum L.). Crop J. 2023, 11, 132–139. [Google Scholar] [CrossRef]
- Tayed, N.; Aubert, G.; Pilet-Nayel, M.L.; Lejeune-Hénaut, I.; Warkentin, T.D.; Burstin, J. Genomic tools in pea breeding programs: Status and perspectives. Front. Plant Sci. 2015, 6, 1037. [Google Scholar]
- Pandey, A.K.; Rubiales, D.; Wang, Y.; Fang, P.; Sun, T.; Liu, N.; Xu, P. Omics resources and omics-enabled approaches for achieving high productivity and improved quality in pea (Pisum sativum L.). Theor. Appl. Genet. 2021, 134, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.K.; Kumar, J.; Gupta, D.S.; Lamichaney, A.; Naik, S.J.S.; Singh, A.K.; Dixit, G.P.; Gupta, S.; Toklu, F. Genomics enabled breeding strategies for major biotic stresses in pea (Pisum sativum L.). Front. Plant Sci. 2022, 13, 861191. [Google Scholar] [CrossRef] [PubMed]
- Kreplak, J.; Madoui, M.-A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Liu, R.; Luo, Y.; Hu, S.; Wang, D.; Wang, C.; Pandey, M.K.; Ge, S.; Xu, Q.; Li, N.; et al. Improved pea reference genome and pan-genome highlight genomic features and evolutionary characteristics. Nat. Genet. 2022, 54, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Bari, M.A.A.; Fonseka, D.; Stenger, J.; Zitnick-Anderson, K.; Atanda, S.A.; Worral, H.; Piche, L.; Kim, J.; Morales, M.; Johnson, J.; et al. A greenhouse-based high-throughput phenotyping platform for identification and genetic dissection of resistance to Aphanomyces root rot in field pea. Plant Phenome J. 2022, 6, e20063. [Google Scholar] [CrossRef]
- Humplík, J.F.; Lazár, D.; Fürst, T.; Husičková, A.; Hýbl, M.; Spíchal, L. Automated integrative high-throughput phenotyping of plant shoots: A case study of the cold-tolerance of pea Pisum sativum L. Plant Met. 2015, 11, 20. [Google Scholar] [CrossRef]
- Nguyen, G.N.; Norton, S.L.; Rosewarne, G.M.; James, L.E.; Slater, A.T. Automated phenotyping for early vigour of field pea seedlings in a controlled environment by colour imaging technology. PLoS ONE 2018, 13, e0207788. [Google Scholar] [CrossRef]
- Roth, L.; Streit, B. Predicting cover crop biomass by lightweight UAS-based RGB and NIR photography: An applied photogrammetric approach. Precis. Agric. 2018, 19, 93–114. [Google Scholar] [CrossRef]
- Zhang, C.; McGee, R.J.; Vandemark, G.J.; Sankaran, S. Crop performance evaluation of chickpea and dry pea breeding lines across seasons and locations using phenomics data. Front. Plant Sci. 2021, 12, 640259. [Google Scholar] [CrossRef]
- Marzougui, A.; McGee, R.J.; Van Vleet, S.; Sankaran, S. Remote sensing for field pea yield estimation: A study of multi-scale data fusion approaches in phenomics. Front. Plant Sci. 2023, 14, 1111575. [Google Scholar] [CrossRef] [PubMed]
- Rispail, N.; Rubiales, D. Rapid and efficient estimation of pea resistance to the soil-borne pathogen Fusarium oxysporum by infrared imaging. Sensors 2015, 15, 3988–4000. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Caballero, S.; Olivoto, T.; Jiménez-Vaquero, M.A.; Rubiales, D.; Rispail, N. RGB image-based method for phenotyping rust disease progress in pea leaves using R. Plant Methods 2023, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Divyanth, L.G.; Marzougui, A.; González-Bernal, M.J.; McGee, R.J.; Rubiales, D.; Sankaran, S. Evaluation of effective class-balancing techniques for CNN-based assessment of aphanomyces root rot resistance in pea (Pisum sativum L.). Sensors 2022, 22, 7237. [Google Scholar] [CrossRef]
- Holdsworth, W.L.; Gazave, E.; Cheng, P.; Myers, J.R.; Gore, M.A.; Coyne, C.J.; McGee, R.J.; Mazourek, M. A community resource for exploring and utilizing genetic diversity in the USDA pea single plant plus collection. Hortic. Res. 2017, 4, 17017. [Google Scholar] [CrossRef] [PubMed]
- Trněný, O.; Brus, J.; Hradilová, I.; Rathore, A.; Das, R.R.; Kopecký, P.; Coyne, C.J.; Reeves, P.; Richards, C.; Smýkal, P. Molecular evidence for two domestication events in the pea crop. Genes 2018, 9, 535. [Google Scholar] [CrossRef]
- Sudheesh, S.; Lombardi, M.; Leonforte, A.; Cogan, N.O.I.; Materne, M.; Forster, J.W.; Kaur, S. Consensus genetic map construction for field pea (Pisum sativum L.), trait dissection of biotic and abiotic stress tolerance and development of a diagnostic marker for the er1 powdery mildew resistance gene. Plant Mol. Biol. Rep. 2014, 33, 1391–1403. [Google Scholar] [CrossRef]
- Ghafoor, A.; McPhee, K. Marker assisted selection (MAS) for developing powdery mildew resistant pea cultivars. Euphytica 2012, 186, 593–607. [Google Scholar] [CrossRef]
- Rai, R.; Singh, A.K.; Singh, B.D.; Joshi, A.K.; Chand, R.; Srivastava, C.P. Molecular mapping for resistance to pea rust caused by Uromyces fabae (Pers.) de-Bary. Theor. Appl. Genet. 2011, 123, 803–813. [Google Scholar] [CrossRef]
- Kwon, S.J.; Smýkal, P.; Hu, J.; Wang, M.; Kim, S.J.; McGee, R.J.; McPhee, K.; Coyne, C.J. User-friendly markers linked to Fusarium wilt race 1 resistance Fw gene for marker-assisted selection in pea. Plant Breed. 2013, 132, 642–648. [Google Scholar] [CrossRef]
- Scott, M.F.; Ladejobi, O.; Amer, S.; Bentley, A.R.; Biernaskie, J.; Boden, S.A.; Clark, M.; Dell’acqua, M.; Dixon, L.E.; Filippi, C.V.; et al. Multi-parent populations in crops: A toolbox integrating genomics and genetic mapping with breeding. Heredity 2020, 125, 396–416. [Google Scholar] [CrossRef]
- Ellis, N.; Hattori, C.; Cheema, J.; Donarski, J.; Charlton, A.; Dickinson, M.; Venditti, G.; Kaló, P.; Szabó, Z.; Kiss, G.B.; et al. NMR metabolomics defining genetic variation in pea seed metabolites. Front. Plant Sci. 2018, 9, 1022. [Google Scholar] [CrossRef] [PubMed]
- Leonova, T.; Popova, V.; Tsarev, A.; Henning, C.; Antonova, K.; Rogovskaya, N.; Vikhnina, M.; Baldensperger, T.; Soboleva, A.; Dinastia, E.; et al. Does protein glycation impact on the drought-related changes in metabolism and nutritional properties of mature pea (Pisum sativum L.) seeds? Int. J. Mol. Sci. 2020, 21, 567. [Google Scholar] [CrossRef]
- Burstin, J.; Salloignon, P.; Chabert-Martinello, M.; Magnin-Robert, J.-B.; Siol, M.; Jacquin, F.; Chauveau, A.; Pont, C.; Aubert, G.; Delaitre, C.; et al. Genetic diversity and trait genomic prediction in a pea diversity panel. BMC Genom. 2015, 16, 105. [Google Scholar] [CrossRef]
- Tayeh, N.; Klein, A.; Le Paslier, M.-C.; Jacquin, F.; Houtin, H.; Rond, C.; Chabert-Martinello, M.; Magnin-Robert, J.-B.; Marget, P.; Aubert, G.; et al. Genomic prediction in pea: Effect of marker density and training population size and composition on prediction accuracy. Front. Plant Sci. 2015, 6, 941. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Nazzicari, N.; Pecetti, L.; Romani, M.; Ferrari, B.; Wei, Y.; Brummer, E.C. GBS-Based genomic selection for pea grain yield under severe terminal drought. Plant Genome 2017, 10. [Google Scholar] [CrossRef]
- Bari, M.A.A.; Zheng, P.; Viera, I.; Worral, H.; Szwiec, S.; Ma, Y.; Main, D.; Coyne, C.J.; McGee, R.J.; Bandillo, N. Harnessing genetic diversity in the USDA pea germplasm collection through genomic prediction. Front. Genet. 2021, 12, 707754. [Google Scholar] [CrossRef]
- Atanda, S.A.; Steffes, J.; Lan, Y.; Al Bari, M.A.; Kim, J.-H.; Morales, M.; Johnson, J.P.; Saludares, R.; Worral, H.; Piche, L.; et al. Multi-trait genomic prediction improves selection accuracy for enhancing seed mineral concentrations in pea. Plant Genome 2022, 15, e20260. [Google Scholar] [CrossRef]
- Carpenter, M.A.; Goulden, D.S.; Woods, C.J.; Thomson, S.J.; Kenel, F.; Frew, T.J.; Cooper, R.D.; Timmerman-Vaughan, G.M. Genomic selection for ascochyta blight resistance in pea. Front. Plant Sci. 2018, 9, 1878. [Google Scholar] [CrossRef]
- Zhao, H.; Pandey, B.R.; Khansefid, M.; Khahrood, H.V.; Sudheesh, S.; Joshi, S.; Kant, S.; Kaur, S.; Rosewarne, G.M. Combining NDVI and bacterial blight score to predict grain yield in field pea. Front. Plant Sci. 2022, 13, 923381. [Google Scholar] [CrossRef]
- Osuna-Caballero, S.; Rispail, N.; Nazzicari, N.; Annicchiarico, P.; Rubiales, D. Predicción genómica para resistencia a roya en guisante. In Proceedings of the Congreso de Mejora Genética de Plantas, Pontevedra, Spain, 19–22 September 2022; p. 72. [Google Scholar]
- Mobini, S.H.; Warkentin, T.D. A simple and efficient method of in vivo rapid generation technology in pea (Pisum sativum L.). Vitro Cel. Develop. Biol.—Plant 2016, 52, 530–536. [Google Scholar] [CrossRef]
- Cazzola, F.; Bermejo, C.J.; Guindon, M.F.; Cointry, E. Speed breeding in pea (Pisum sativum L.), an efficient and simple system to accelerate breeding programs. Euphytica 2020, 216, 178. [Google Scholar] [CrossRef]
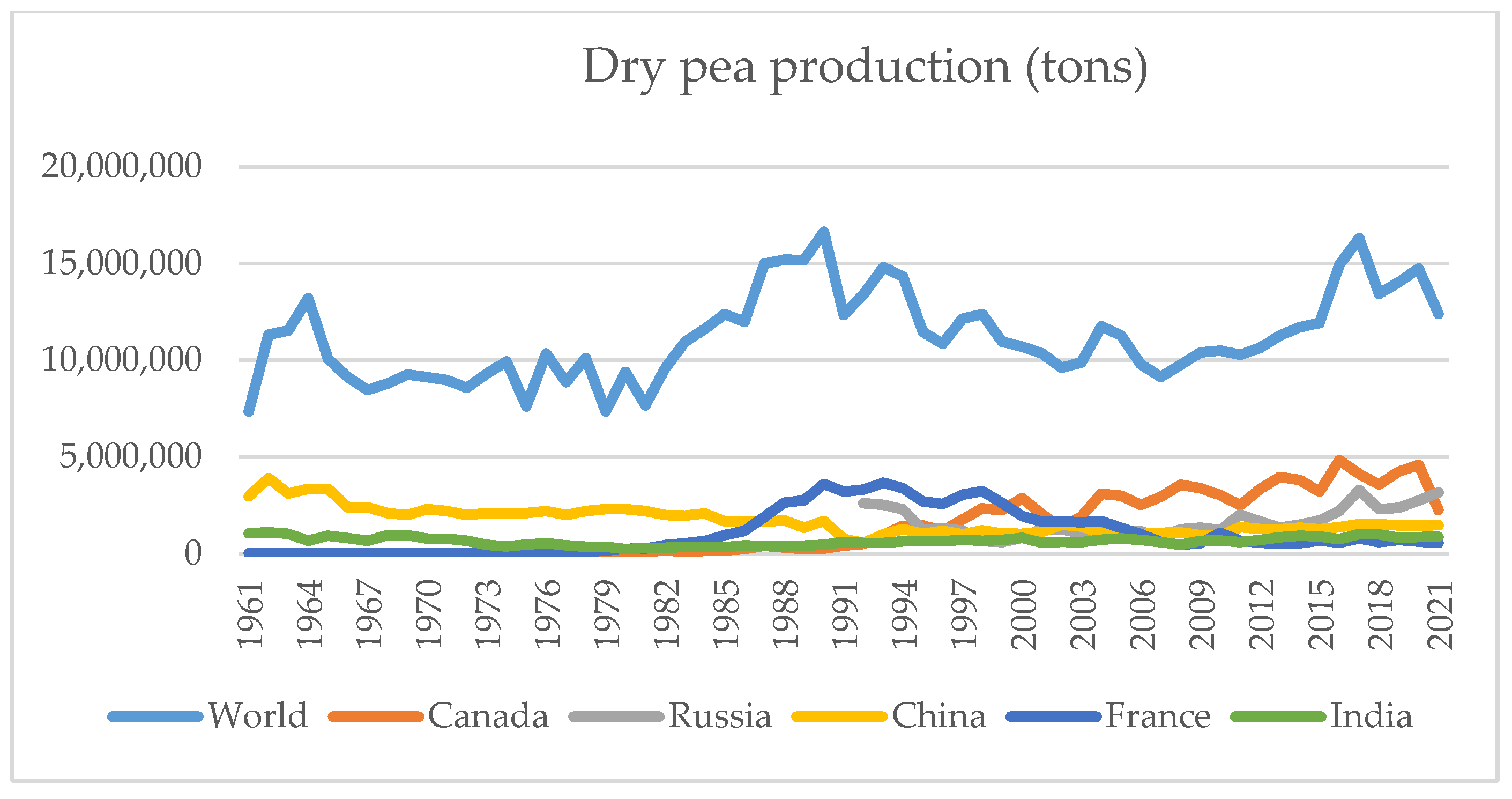
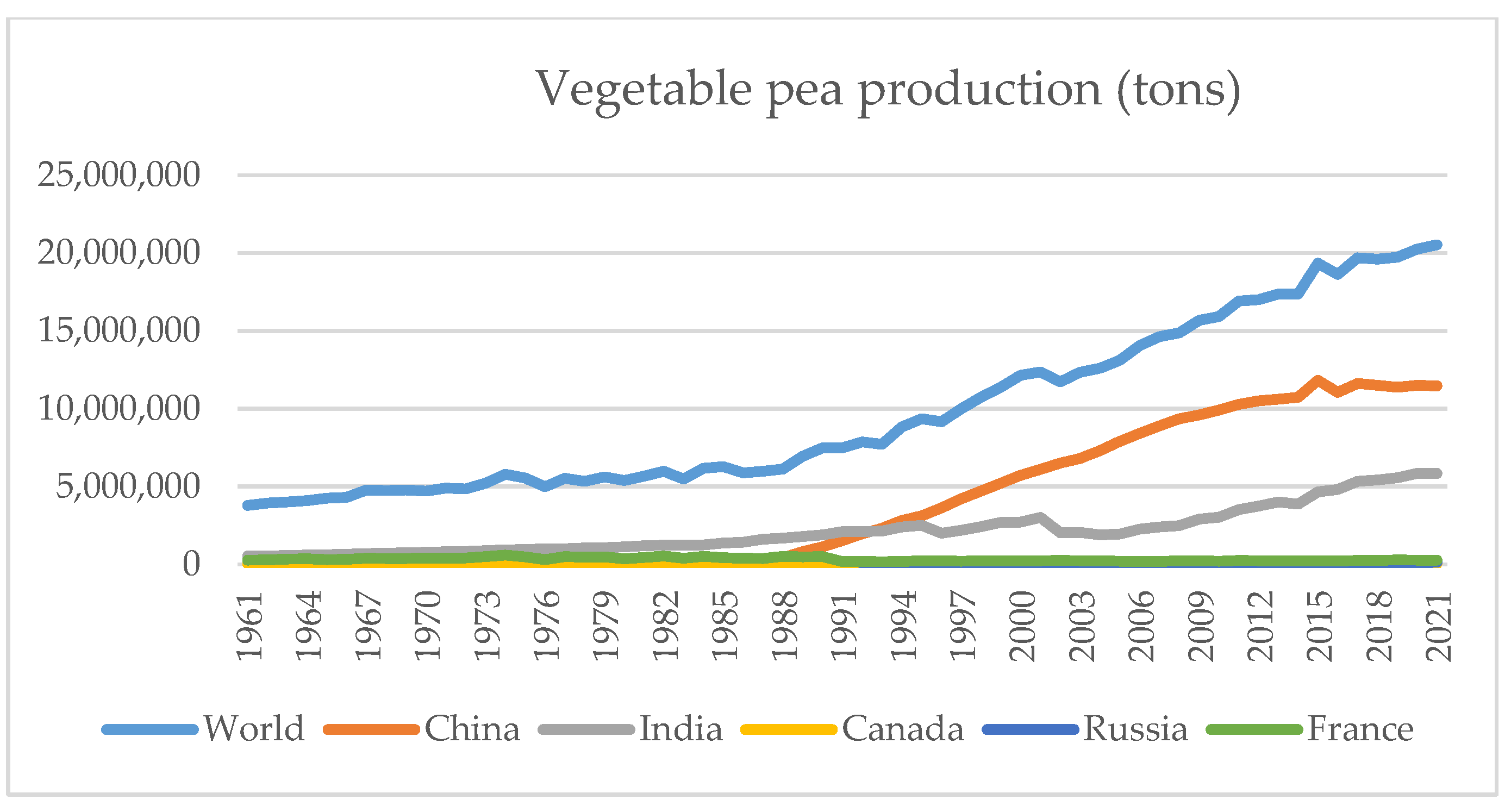
| Biotic Stress | Pathogen Species | Source of Infection | Organ | Distribution |
|---|---|---|---|---|
| Aerial fungi | ||||
| Ascochyta blight complex | Ascochyta pisi Lib. | Infected crop debris, seedborne, ascospores, and conidia | Leaves, stems, pods, and seeds | Europe and North America |
| A. pinodes Berk. and Blox. | Infected crop debris, seedborne, ascospores, and conidia | Leaves, stems, pods, and seeds | Worldwide | |
| Phoma medicaginis var. pinodella (L.K. Jones) Boerema | Infected crop debris, seedborne, ascospores, and conidia | Leaves, stems, pods, and seeds | Worldwide | |
| P. koolunga Davidson | Infected crop debris, seedborne, ascospores, and conidia | Leaves, stems, pods, and seeds | Australia | |
| P. glomerata [(Corda) (Wollenw. and Hochapfel)] | Infected crop debris, seedborne, ascospores, and conidia | Leaves, stems, pods, and seeds | Australia | |
| Powdery mildew | Erysiphe pisi (DC.) | Infected crop debris and conidia | Leaves, stems, and pods | Worldwide climates with warm, dry days and cool nights |
| E. trifolii (Grev.) | Infected crop debris and conidia | Leaves, stems, and pods | USA, India, Spain, and Tunisia | |
| Downy mildew | Peronospora viciae (Berk.) Caspary f.sp. pisi Sidow. | Infected crop debris, oospores, and conidia | Leaves, stems, pods, and seeds | Cool and wet weather conditions |
| Rust | Uromyces pisi (Pers.) Wint. | Infected debris of Euphorbia cyparissias L. and urediospores | Leaves, stems, and occasionally pods | Temperate regions |
| U. viciae-fabae (Pers.) de Bary | Infected crop debris, aeciospores, and urediospores | Leaves, stems, and occasionally pods | Tropical and sub-tropical regions, e.g., India, China | |
| Soilborne diseases | ||||
| Fusarium wilt | Fusarium oxysporum f.sp. pisi (W.C. Snyder and H.N. Hansen) | Infected crop debris, chlamydospores, and micro- and macroconidia | Roots, xylem vessels, and seeds | Worldwide, in both dry and wet field conditions |
| Fusarium root rot complex | Fusarium solani f. sp. pisi (W.C. Snyder and H.N. Hansen) | Infected crop debris, chlamydospores, and micro- and macroconidia | Roots and seeds | Worldwide (mainly in the Pacific North-West regions) |
| F. graminearum Schw. | Infected crop and cereal debris, ascospores, and micro- and macroconidia | Roots and seeds | Canada, USA, and Europe | |
| F. avenaceum (Fries) Saccardo | Infected crop debris and ascospores | Roots and seeds | Canada, USA, and Europe | |
| Common root rot | Aphanomyces euteiches (Drechsler) | Infected soil and crop debris, oospores, and zoospores | Roots, stems, and leaves | Worldwide, temperate, and wet areas |
| Rhizoctonia root rot | Rhizoctonia solani Kühn | Infected soil and sclerotia | Roots, stems, and leaves | Temperate and subarctic areas |
| Bacteria | ||||
| Pea blight | Pseudomonas syringae pv. pisi Sackett | Infected seeds and crop debris | Leaves, stems, pods, and seeds | Areas with cool and wet weather |
| Viruses | ||||
| Pea Seed-borne Mosaic Virus | PSbMV | Infected seeds and aphids | Leaves, stems, pods, and seeds | Worldwide |
| Pea Enation Mosaic Virus | PEMV | Infected aphids (infected seeds in a small proportion) | Leaves, stems, pods, and seeds | USA, Europe, Africa, and India |
| Nematodes | ||||
| Cyst nematodes | Heterodera goettingiana Liebscher | Infected soil and roots with eggs | Roots | Worldwide |
| Root-knot nematodes | Meloidogyne incognita (Kofoid and White) Chitwood | Infected soil and roots with eggs | Roots | Europe |
| Root lesion nematodes | Pratylenchus neglectus Rensch | Infected soil and roots with eggs | Roots | Worldwide |
| P. thornei Sher and Allen | Infected soil and roots with eggs | Roots | Worldwide | |
| Parasitic plants | ||||
| Broomrapes | Orobanche crenata Forskal | Infested soil with seeds | Roots | Mediterranean basin |
| Insect pests | ||||
| Pea weevil | Bruchus pisorum L. | Infested seeds | Pods and seeds | Worldwide |
| Pea aphid | Acyrthosiphon pisum H. | Infested soil and crop debris with eggs, parthenogenetic individuals | Leaves, stems, pods, and seeds | Temperate areas |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubiales, D.; Barilli, E.; Rispail, N. Breeding for Biotic Stress Resistance in Pea. Agriculture 2023, 13, 1825. https://doi.org/10.3390/agriculture13091825
Rubiales D, Barilli E, Rispail N. Breeding for Biotic Stress Resistance in Pea. Agriculture. 2023; 13(9):1825. https://doi.org/10.3390/agriculture13091825
Chicago/Turabian StyleRubiales, Diego, Eleonora Barilli, and Nicolas Rispail. 2023. "Breeding for Biotic Stress Resistance in Pea" Agriculture 13, no. 9: 1825. https://doi.org/10.3390/agriculture13091825