Abstract
Lentinus sajor-caju has shown potential for the bioconversion of lignocellulosic biomass in various substrates. Here, we evaluated whether L. sajor-caju affects the lignocellulosic biomass, in vitro fermentation and antioxidant properties of Astragalus membranaceus var. mongholicus (AMM) stems. The lignocellulosic biomass content and the antioxidant activity of AMM stems were determined by scanning electron microscopy, chemical component analysis, in vitro fermentation and LC-MS metabolomics after 30 days of fermentation by L. sajor-caju at 25 °C. L. sajor-caju significantly altered the rigid structure of the stems. Compared with the control condition, the lignocellulose contents were significantly reduced (p < 0.001) and improved in vitro digestibility and total volatile fatty acids. In total, 624 differential metabolites, including 201 up-regulated and 423 down-regulated, were identified in unfermented and fermented comparison groups. Correlation analysis indicated that there were strongly correlations of the total phenolic content and total antioxidant capacity. Meanwhile, the differential metabolites were primarily associated with antioxidant activity, with 4(3H)-quinazolinone, dihydrocarvyl acetate and jaceoside being the most representative. In summary, L. sajor-caju altered the composition of the cell wall of AMM stems, thereby enhancing their antioxidant activity.
1. Introduction
Astragalus membranaceus (Fisch.) Bge. var. mongholicus (Bge.) Hsiao (A. membranaceus var. mongholicus) (AMM) is a perennial, herbaceous plant from the legume family [1,2]. Its root has been widely used as a traditional Chinese medicine (Astragali Radix) and tonic food material for thousands of years [3]. AMM can reinforce cell-mediated immunity and improve antiviral activity, protect liver and stomach health, inhibit cancer and exert anti-inflammatory and anti-aging effects [4,5,6,7]. In addition, AMM contains a wide variety of bioactive constituents, such as triterpenoid saponins, flavonoids and amino acids, that have health-enhancing properties. These characteristics have led to a huge increase in the annual production of AMM roots, which are now among the major medicine and food homology materials with the largest market share.
Like the roots, AMM stems and leaves also contain a variety of bioactive constituents with health-promoting properties [8]. However, in most cases, the aerial parts (leaf and stem) of AMM are directly burned after harvesting the roots, which resulted in natural resource waste and significant environmental pollution. It has been long known that AMM can eliminate free radicals and effectively inhibit oxidation, and its supplementation in goat feedstock was reported to effectively improve the antioxidant capacity of the animals [9]. AMM stems have potential for development as functional feed. Moussaoui et al. (2010) [10] showed that Astragalus armatus has a lignin content of approximately 17%, which is higher than that in corn straw (average 12.5%) [11], but less than that in wheat straw (20.5%) or paddy straw (18.5%) [12]. This highlights the potential of utilizing AMM by-products as ruminant feed, which would effectively contribute to reducing resource waste and improving the economic efficiency of animal production.
It is difficult for ruminants to degrade lignin, but they are able to convert cellulose and hemicellulose into energy for use. The substantial degradation of lignin and the retention of cellulose and hemicellulose have the potential to improve the utilization of roughage in ruminants. Therefore, improving lignin degradation is the key to promoting the feed conversion potential of AMM by-products. Some white-rot fungi preferentially attack on lignin in a selective manner by secreting laccase, lignin peroxidase and manganese peroxide. The lignin can be decomposed to obtain phenolic and aromatic compounds with various chemical structures by white-rot fungi, and synthesize high- value (antioxidant, anti-inflammatory, antineoplastic, etc.) compounds like flavonoid, terpenes and other small molecules [13]. Lignocellulose-degrading Lentinus sajor-caju is an edible mushroom (Basidiomycetes) belonging to the white-rot fungi. We have previously shown that L. sajor-caju had increasing crude protein content (13.83%) and the best selectivity of degrading lignin (40.95%) and of the substrate to a greater extent compared with five other white-rot fungal species [14]. To promote the efficient utilization of AMM by-products to develop functional (strong antioxidant activity) feed source, in this study, we evaluated whether L. sajor-caju affects the lignocellulosic biomass, antioxidant properties and in vitro rumen digestibility of AMM stems.
2. Materials and Methods
2.1. Fungal Preparation
White-rot fungi, Lentinus sajor-caju (Fr.) Fr. CGMCC 5.592, was used for this study. It was provided by China General Microbiological Culture Collection Center (CGMCC) in Beijing, China. The fungus was placed in potato dextrose agar (PDA, Haibo, Qingdao, China) medium and incubated at 4 °C. PDA plates were inoculated with six pieces of the fungus (6 mm diameter) at 25 °C for 7 days.
2.2. Experimental Set-Up
AMM stems were collected from Tongliao County, Inner Mongolia, China (42°15′–45°59′ N, 119°14′–123°34′ E), and chopped into small pieces (1–2 cm). The 50 g AMM stems was placed in triangular flask at 65% (w/w) moisture content, sterilized by autoclave (Boxun YXQ-50A, Shanghai, China) at 121 °C during 30 min, and then stayed in a cool room temperature. A 2-cm3 inoculum from 7 days fungal cultures grown on solid PDA at 25 ± 0.5 °C was added to each triangular flask, and they were then incubated 30 days in the dark at 25 ± 0.5 °C until sampling (LP group) (Figure 1). Six replicates were used per AMM stem sample. Controls were subjected to the same experimental conditions but were not inoculated (CK group).

Figure 1.
Control and Lentinus sajor-caju-treated Astragalus membranaceus (AMM) stems after 30 days of fermentation at 25 °C. (a) Control (unincubated) AMM stems and (b) L. sajor-caju-treated AMM stems. Scanning electron micrographs of unfermented (c) and Lentinus sajor-caju-fermented (d) Astragalus membranaceus stems (scale bars: 50 μm).
2.3. Structural Characterization of AMM Stems Using Scanning Electron Microscopy
Fermented and unfermented AMM stems samples were analyzed from a macroscopic perspective by scanning electron microscopic (SEM). The sample were reduced to the small pieces (2.5 × 2.5 mm) and kept in different concentrations of ethanol, critical-point dried and attached to the sample stage. Finally, AMM stems samples were viewed using SEM MIRA3 (TESCAN, Brno, Czech Republic).
2.4. Chemical Analysis
After 30 days of fermentation, substrate samples were oven-dried at 105 °C for 48 h. After that, the samples were crushed and passed a 2 mm net mesh. Neutral detergent fiber (NDF), acid detergent fiber (ADF) and acid detergent lignin (ADL) contents were assessed, according to the methods of Van Soest [15,16]. Cellulose and hemicellulose content were calculated according to Wang et al. [14].
2.5. In Vitro Rumen Fermentation
In vitro rumen digestibility was performed according to Wang et al. [14]. Rumen contents were collected from three Horqin yellow cattle fed AMM stems and filtered through four layers of cheesecloth, mixed with the buffer solution (v/v 1:4) and purged with CO2. The 500 mg of each sample (DM basis) and 50 mL buffered rumen fluid was mixed and added to the 100 mL fermentation flask before being incubated for 48 h at 39 °C. Each sample was incubated in triplicate. After fermentation for 48 h, the sample was dried at 65 °C to constant weight and analyzed for digestibility (DM, NDF, ADF, cellulose, hemicellulose, and ADL). The rumen fluid was centrifuged at 12,000× g for 20 min at 4 °C, and then the supernatants were measured for pH using a PHS-3C pH meter (Olabo, Jinan, China). The volatile fatty acids (VFA) were determined as described by Wang et al. [14].
2.6. Antioxidant Activity
A total of 10 g of fresh samples and 100 mL methyl alcohol were mixed in a triangular flask and ultrasonically extracted at 25 °C for 4 h. All samples were centrifuged at 10,000 rpm and 4 °C for 15 min in order to separate the supernatant. The supernatant was used for antioxidant activity analysis.
The antioxidant capacity (DPPH, reducing power, FRAP and Fe2+ scavenging) and total phenolic content (TPC) were determined by using kits according to the instructions from Beijing Solarbio Science Technology Co., Ltd. (Beijing Solarbio Science Technology Co., Ltd., Beijing, China) [17,18,19,20,21].
2.7. Metabolomic Profiling Analysis
The 0.5 g samples were mixed with 1 mL extract solution (methanol:H2O = 3:1 [v/v]). The mixtures were kept in a thermostatic shaker at 30 °C at 220 rpm for 4 h and then centrifuged at 12,000 rpm at 4 °C for 15 min. The identification of differential metabolites was performed using high-performance liquid chromatography-mass spectrometry/mass spectrometry (LC–MS/MS). The sample was separated using Compass C18 (2) reversed-phase column (100 mm × 2.1 mm × 1.7 μm; Waters, Santa Clara, CA, USA) and maintained at 4 °C.
The mobile phase consisted of solvent A (0.04% acetic acid in ultrapure water) and solvent B (0.04% acetic acid in acetonitrile). The extract was injected while the gradient was 0.5 min, 5% A, 95% B; 0.5–7 min, 95% A, 5% B; 7–8 min, 5% A, 95% B; 9 min, 5% A, 95% B; 9–9.1 min, 5% A, 95% B; 12 min, 95% A, 5% B. The 2 μL sample was injected into the column and left at 40 °C. For MS analysis, the parameters for the scan range of 50–1500 (m/z), needle voltage, +3600 V/−3500 V.
2.8. Statistical Analyses
One-way analysis variance (ANOVA) were used to analyze all data by SAS v.9.0. High confidence (VIP > 1) and p-value < 0.05, a minimum of a two-fold change or a maximum (Fold Change (FC) > 2 or <0.05), were considered as differential components. Spearman rank correlation coefficients were performed by using R package (R Core Team, Vienna, Austria, v2021) for visualization.
3. Results and Discussion
3.1. Morphological Observations and Structural Characterization
The morphology and SEM images of the AMM stems in CK and LP group after 30 days of fermentation are shown in Figure 1. Compared with the AMM stems in the CK group (Figure 1a), the morphology of the fermented AMM stems in the LP group were covered with axenic L. sajor-caju white mycelium, and the stem particles were packed tightly (Figure 1b). The SEM images of the AMM stems in the CK group showed a flat, intact, and clear surface structure (Figure 1c), whereas the surface texture of the AMM stems in the LP group was rough and porous (Figure 1d). These results demonstrate that L. sajor-caju mycelia can penetrate the AMM stems and destroy the cell wall structure. This is consistent with Zhao et al., who reported that white-rot fungi significantly altered the rigid structure of straw under solid-state fermentation [20].
3.2. Lignocellulose Composition
The lignocellulose composition (DM basis) of AMM stems after fermentation with L. sajor-caju was shown (Table 1). The lower of lignocellulosic content in the LP group than that in the CK group (p < 0.001). The neutral detergent fiber (NDF), acid detergent fiber (ADF), cellulose, hemicellulose, and acid detergent lignin (ADL) contents in AMM stems were reduced by 16.38%, 12.14%, 4.27%, 23%, and 31.09%, respectively, with the LP group compared with that in the CK group.

Table 1.
The mass loss rate and lignocellulose composition (% DM basis) of Astragalus membranaceus stems following fermentation with Lentinus sajor-caju.
The concentration of lignocellulose in plants varies according to plant type. The lignin and α-cellulose contents of hardwood trees range from 20.2% to 35.5% and from 48.2% to 59.0%, respectively [22]; meanwhile, grasses contain 15% to 25% lignin by weight [23]. The lignocellulose contents in AMM stems (leguminous plants) were reported to be approximately 17% lower than those in wood [10]. This indicates that AMM stems have a good potential to be developed as a feed resource.
White-rot fungus-mediated lignin degradation is efficient, especially in the context of its application in the removal of lignin from roughage (crops and grasses) [24,25]. Thus far, white-rot fungi are efficient microorganisms for degrading roughage development and utilization relating to a wide range of agricultural by-products. Studies have found that L. sajor-caju exhibits high lignocellulose degradation potential under axenic culture conditions. For instance, the ADL degradation rate in highland barley straw was found to be 46.29% after 21 days of fermentation with L. sajor-caju [14]. Additionally, after 21 and 30 days of L. sajor-caju pretreatment, 40.95% and 38.29% of the lignin in corn straw was degraded, respectively, whereas for cellulose the proportions were 23.13% and 35.25%, respectively [26,27].
In this study, lignin, hemicellulose and cellulose loss in AMM stems was 31.09%, 23% and 4.27% after 30 days of inoculation with L. sajor-caju, respectively. L. sajor-caju preferentially degrades lignin, followed by hemicellulose and cellulose. Therefore, the content of cellulose changed little in the AMM stems with L. sajor-caju fermentation. The lignocellulosic component of banana waste (leaves and pseudostem) followed a similar pattern as with the AMM stems fermented with L. sajor-caju [28]. The reduction of cellulose and lignin contents in the banana leaves and pseudostem were 4%, 52% and 1.5%, a 23% reduction, respectively. The reduction of cellulose contents was lower when compared to lignin contents, which was consistent with the experimental results. Low-lignin lignocellulosic materials are a good source of animal feed and help improve digestibility of ruminants.
Although the lignin degradation rate was lower in AMM stems than in corn straw or highland barley straw, the lignin degradation ability of L. sajor-caju was still significant. The degradation rate of lignin is higher than that of cellulose and hemicellulose, and it further indicated that L. sajor-caju was related to the preferential selection of lignocellulosic degradation. In addition, the study been shown that with increasing fermentation duration, the degradation rate of cellulose gradually exceeds that of lignin by some white-rot fungi [25]. The loss ratio of lignocellulose by white-rot fungi is dependent on both the fermentation substrate and the duration of fermentation.
3.3. In Vitro Rumen Digestibility
The effect of fermented and unfermented of chemical composition digestibility and fermentation variables in the AMM stems is shown (Table 2). The dry matter digestibility (DMD), neutral detergent fiber digestibility (NDFD), acid detergent fiber digestibility (ADFD), cellulose digestibility (CLD), hemicellulose digestibility (HLD) and lignin digestibility (ADLD) was significantly (p < 0.001) higher in the LP group than in the CK group. With the L. sajor-caju fermentation, the pH was not significantly (p > 0.05) affected compared to the unfermented AMM stems. The propionic acid content was reduced, while the total VFA, acetic acid and butyric acid content increased (p < 0.001) after 30 days of incubation.
Table 2.
Chemical composition digestibility and fermentation variables in unfermented and fermented AMM stems by L. sajor-caju.
The increased digestibility of lignocellulose was associated with the ability of L. sajor-caju to damage the plant cell wall structure. In addition, the DMD with L. sajor-caju fermented was increased (12.52%) compared to the CK group, which was likely due to the degradations of NDF and ADF. This result further confirms that the white-rot fungi L. sajor-caju can improve fiber digestibility and feed utilization, which is consistent with the results of our previous studies on the degradation of other substrates by L. sajor-caju [14,26]. The concentration of VFA reflects the energy provided by the fermentation substrate to the animal. The more fermentable carbohydrates in the feed, the higher the in vitro digestibility [29]. In this study, there was a decreased ADL content and an increased DMD and total VFA in the AMM stems with L. sajor-caju fermentation, which could be because L. sajor-caju weakened the cell wall recalcitrance and promoted the utilization of substrate carbohydrates by rumen microorganisms. In addition, the increase of propionic acid content inhibited the activity of rumen microorganisms that produced acetic acid in rumen. L. sajor-caju treatment increased the release of energy from the AMM stems and promoted propionic acid content. Increase in the concentration of the total VFA and propionic acid, and a decrease in acetic acid in Pleurotus ostreatus fermented rice straw and wheat straw were reported by Datsomor et al. and Sufyan et al. [30,31].
3.4. The Antioxidant Activity of AMM Stem Extracts
The antioxidant capacity of the AMM stems was determined using DPPH and FRAP assays as well as by the assessment of reducing power, Fe2+ scavenging ability and TPC (Table 3). The antioxidant activity of AMM stem extracts significantly increased (p < 0.001) after 30 days in the fermented substrate relative to that in the unfermented substrate. TPC, DPPH, FRAP, reducing power and Fe2+ scavenging activity were 86.2%, 22.01%, 46.67%, 29.58% and 67.80% higher in the LP group than in the CK group, respectively. The high antioxidant activity of extracts in the LP group could be related to the structural transformation after L. sajor-caju fermentation. The above findings indicated that the L. sajor-caju-fermented AMM stems are an important source of antioxidants for roughage.
Table 3.
The antioxidant properties of Astragalus membranaceus stems following fermentation with L. sajor-caju.
Oxidative stress refers to a condition in which the oxidative reactions in cells lead to tissue damage and cell death [32]. Antioxidants can reduce oxidative stress by scavenging free radicals, thereby reducing the incidence of many diseases and improving the quality of animal products [33]. Naturally occurring antioxidants may have many advantages over synthetic antioxidants in promoting animal health and improving safety for human consumption. It is well known that L. sajor-caju, a typical Basidiomycete (mushroom), contains bioactive compounds with beneficial antioxidant properties [34].
We have demonstrated in this current study that L. sajor-caju can increase antioxidant activity, and it was found that the Fe2+ scavenging activity exhibited the greatest increase (1.68-fold) in fermented AMM stems compared with unfermented AMM stems. In addition, other antioxidant properties were also significantly increased in extracts of L. sajor-caju treatment, which is consistent with the findings of Elhusseiny et al. (2021) [35]. However, Sun et al. (2022) [36] showed that the DPPH and the FRAP were markedly decreased in fermented green tea following 3 days of fermentation with L. sajor-caju. The results of the current experiment contrast with those of Sun et al. (2022) and may be explained by differences in, among other factors, the substances used, cultivation methods and fermentation periods between the studies [37].
3.5. Metabolite Differences and Classification
3.5.1. Analysis of Differential Metabolite
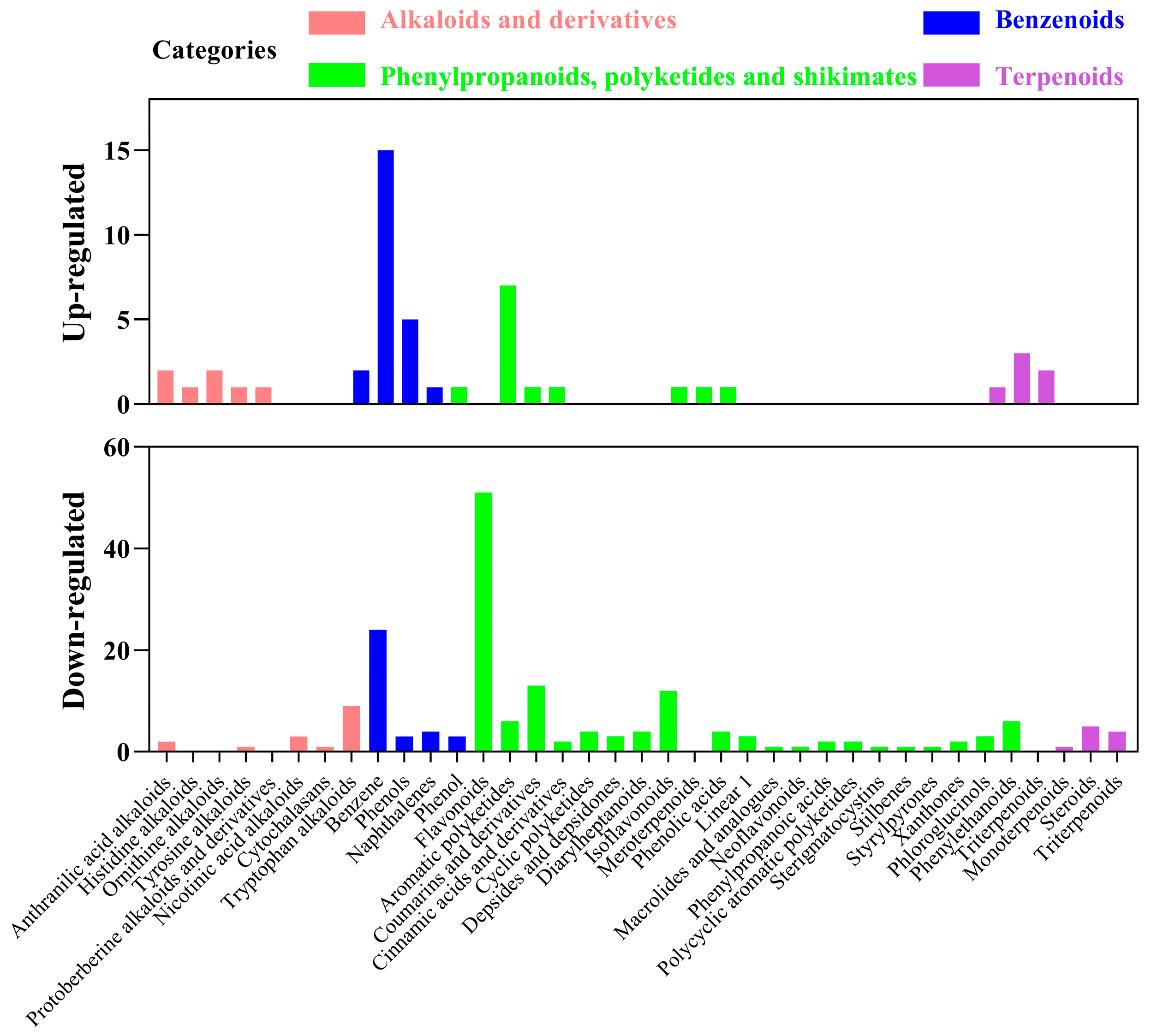
The pie charts clearly showed the significant differential abundance between differential metabolites in fermented AMM stems and unfermented AMM stems (Figure 2a). There were 624 differential metabolites, 201 up-regulated and 423 down-regulated, between the two groups. In the total of the 624 differentially abundant metabolites in the current analysis, 143 were phenylpropanoids, polyketides and shikimates (22.92%), 117 were lipids or lipid-like molecules (18.75%), 95 were organ oheterocyclic compounds (15.22%), 59 were benzenoids (9.46%), 41 were organic acids and derivatives (6.57%), 30 were alkaloids and derivatives (4.81%), 24 were organic oxygen compounds (3.84%), 19 were terpenoids (3.04%), 12 were organic nitrogen compounds (1.92%), 10 were amino acids and peptides (1.60%), 8 were carbohydrates (1.28%), 8 were fatty acids (1.28%) and 58 were other metabolites (9.29%).
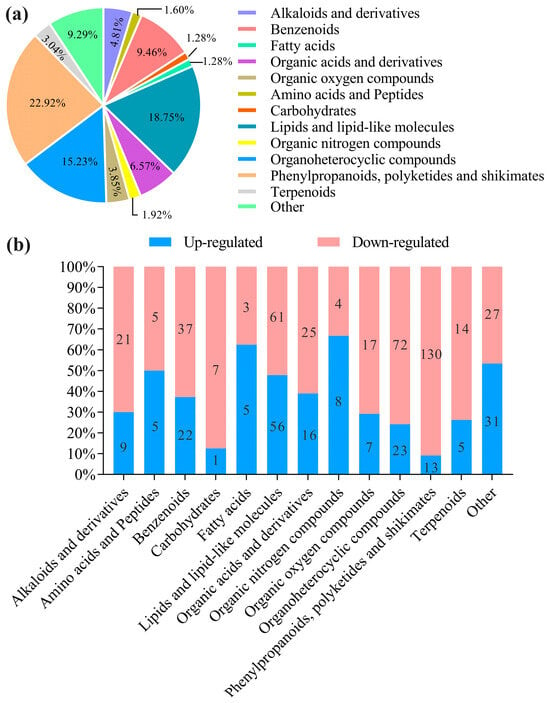
Figure 2.
Analysis of the identified metabolites. The main types of different metabolites were indicated by color codes (fold change < 0.5 or >2, p-value < 0.05) are shown in the pie charts (a). Differential metabolite up-regulated and down-regulated of the different superclass classifications (b).
Different classes of metabolites were comparatively investigated in different treatment groups. In particular, fatty acids and organic nitrogen compounds were 1.78- and two-fold more abundant in the fermented AMM stems than in unfermented AMM stems in the up-regulated metabolites, whereas phenylpropanoids, polyketides and shikimates, carbohydrates, organoheterocyclic compounds, terpenoids, organic oxygen compounds, alkaloids and derivatives, benzenoids, organic acids and derivatives and lipids or lipid-like molecules were 13-, 7-, 3.13-, 2.8-, 2.43-, 2.33-, 1.68-, 1.56- and 1.09-fold lower with fermented AMM stems than unfermented AMM stems (Figure 2b).
The large amounts of highly represented superclass in LP group was phenylpropanoids, polyketides and shikimates, while phenylpropanoids, polyketides and shikimates, lipids and lipid-like molecules and organoheterocyclic compounds were the top three represented subclasses among all metabolites. This result was in accordance with those of our previous reported for fermented of highland barley by L. sajor-caju [38].
However, although lipids and lipid-like molecules were higher than phenylpropanoids and polyketides, the result of this experiment displayed the opposite trend [39]. The intermediates of the shikimate pathway were a substrate of the phenylpropanoids biosynthesis pathway. Flavonoid compounds are synthesized by phenylpropanoids, polyketides and shikimates pathway in higher plant species [40]. Therefore, the three are grouped into one category in our study. Although, the contents of phenylpropanoids, polyketides and shikimates in fermented AMM stems was similar to phenylpropanoids and polyketides detected in fermented tea leaves [41]. However, the phenylpropanoids and polyketides were slightly lower, which is because the shikimate showed no significant difference in the fermented tea leaves. Their result also verified that microbial fermentation treatment can induce the above metabolic processes
3.5.2. Antioxidant Composition in AMM Stems Fermented and Unfermented
It is well known that alkaloids, benzenoids, phenylpropanoids, polyketides, shikimates and terpenoids pathways were closely related to antioxidant activities [42,43]. The present study was investigated the metabolite composition with antioxidant function. The antioxidant components of metabolites were divided into alkaloids and derivatives, phenylpropanoids, polyketides and shikimates, benzenoids and terpenoids (Figure 3). Despite the differences between up-regulated and down-regulated metabolites, we found that there were some commonalities between unfermented and fermented AMM stems by L. sajor-caju. Among the 30 alkaloids and derivatives were 4 anthranilic acid alkaloids (2 up-regulated and 2 down-regulated) and 2 tyrosine alkaloids (1 up-regulated and 1 down-regulated), while the others classified are only either up-regulated or down-regulated metabolites. Among the 143 phenylpropanoids, polyketides and shikimates, 58 were flavonoids (7 up-regulated and 51 down-regulated), 14 were coumarins and derivatives (1 up-regulated and 13 down-regulated), 13 were isoflavonoids (1 up-regulated and 12 down-regulated), 7 were aromatic polyketides (1 up-regulated and 6 down-regulated), 7 were phenylethanoids (1 up-regulated and 6 down-regulated), 5 were phenolic acids (1 up-regulated and 4 down-regulated), while the others classified are only either down-regulated metabolites. Among the 19 terpenoids, 3 were monoterpenoids (2 up-regulated and 1 down-regulated), while the others classified are only either up-regulated or down-regulated metabolites. The difference in the composition and content of differential metabolite might lead to different antioxidant activities. Many substances in the metabolites have displayed potent biological activities and promote animal health, such as alkaloids and phenols. Phenolic of different classes are converted and increased antioxidant activity by fungal fermentation [44]. The bioaccessibility of phenolic compounds after fermentation depends on the type of microorganisms and fermentation substrate. This view is also confirmed by research in our laboratory. In this experiment, the metabolites of L. sajor-caju-fermented AMM stems were different from those of L. sajor-caju-fermented highland barley straw [14].
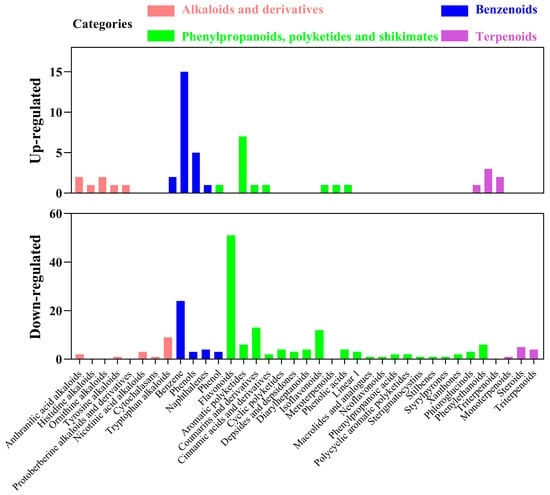
Figure 3.
Antioxidant composition among the up-regulated and down-regulated metabolites in CK.
The association between the lignin content and antioxidant indices (Fe2+ scavenging activity, TPC, FRAP, reducing power and DPPH scavenging ability) of extracts in both fermented and unfermented AMM stems was determined by Spearman rank correlation analysis (Table 4). The loss of lignin was significantly correlated with antioxidant activity in the AMM stems with fermentation by L. sajor-caju. The strongest positively correlation was between lignin losses and Fe2+ scavenging activity (r = 0.86), reducing power and TPC (r = 0.82). TPC had the strongest correlation with other antioxidant indices. In our study, the ligninolysis contents were significantly decreased, suggesting that white-rot fungi L. sajor-caju fermentation could facilitate the conversion of polyphenol compounds, thereby enhancing the biological function of AMM stems as ruminal feed.
Table 4.
Spearman rank correlation coefficient (r) between ADL loss in unfermented and fermented AMM stems with antioxidant activity by L. sajor-caju.
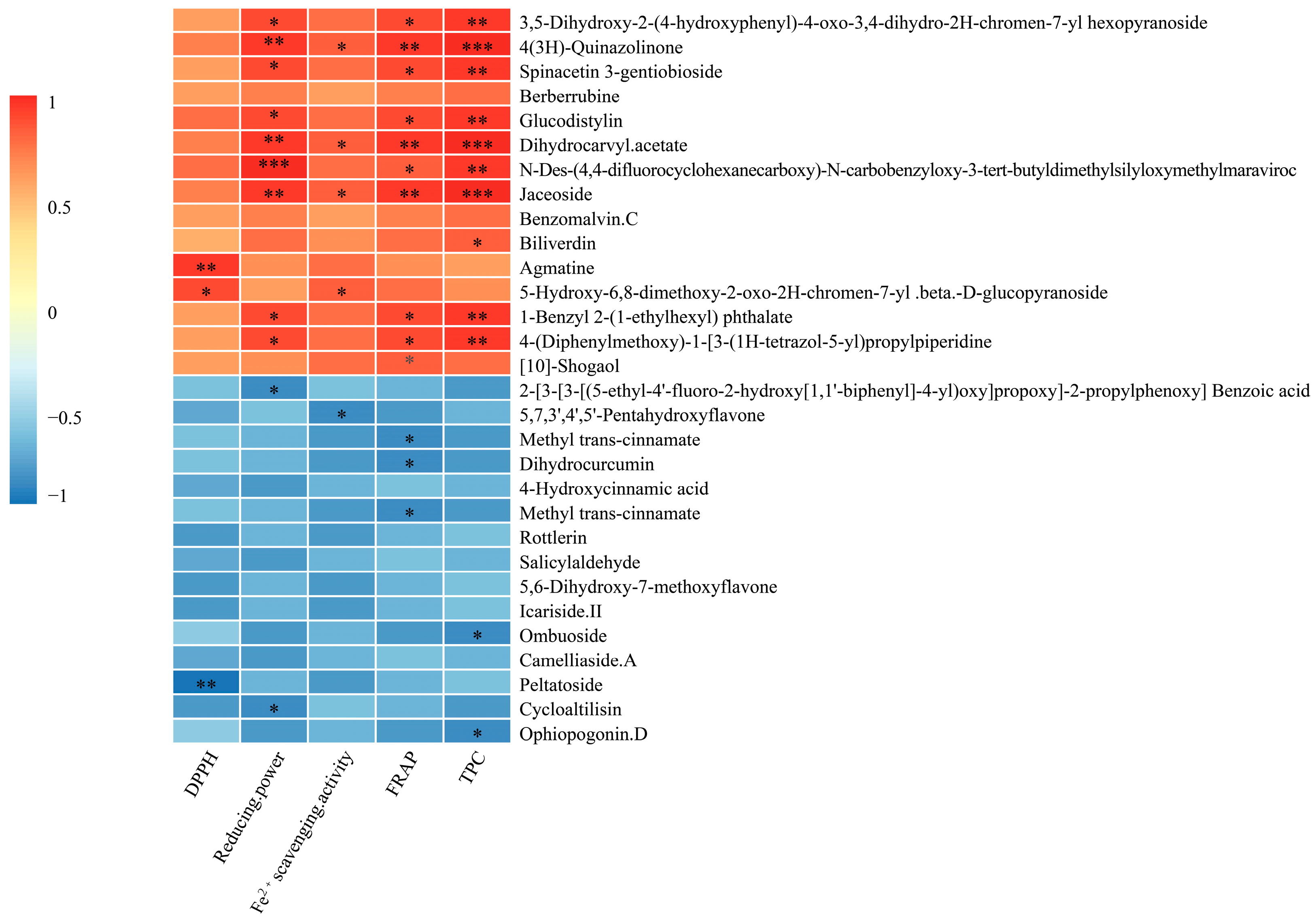
The results of the correlation analysis to analyze the relationship between the top 15 up-regulated and down-regulated antioxidant composition metabolites and the DPPH, reducing power, Fe2+ scavenging activity, FRAP and TPC are shown (Figure 4). Peltatoside was significantly (p < 0.01) negatively correlated with DPPH. There were seven metabolites with a negative correlation with antioxidant indices, among which peltatoside was significantly (p < 0.01) negatively correlated with DPPH. Strictosamide, dihydrocurcumin and methyl trans-cinnamate (p < 0.05) negatively correlated with FRAP. There was the presence of a strong correlation between the 4(3H)-quinazolinone, dihydrocarvyl acetate, jaceoside for reducing power, FRAP, Fe2+ scavenging activity and TPC. The benzenoids and substituted derivatives were significantly positively correlated with reducing power, FRAP, Fe2+ scavenging activity (p < 0.05) and TPC (p < 0.01). The results of the correlation showed that although the down-regulated metabolite FC changed greatly, it had little correlation with antioxidant activity.
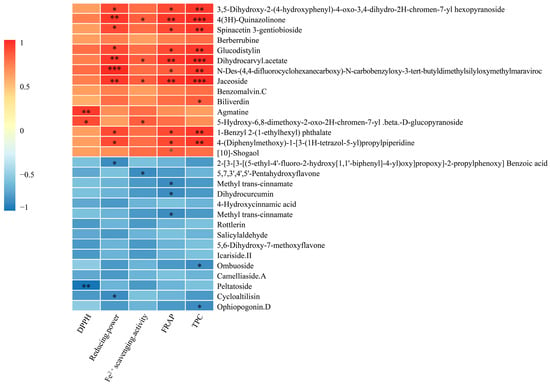
Figure 4.
Correlation analysis between the top 15 up-regulated and down-regulated antioxidant composition metabolites and antioxidant properties (* p < 0.05, ** p < 0.01, *** p < 0.001).
Correlation analysis indicated the relationships between different metabolites and antioxidant properties. In this study, 4(3H)-quinazolinone, dihydrocarvyl acetate and jaceoside were the main metabolic markers with antioxidant properties. Quinazolinone is an alkaloid heterocyclic compound found in active components of many medicinal plants [42] and which has an antioxidant activity. The 4(3H)-quinazolinone is formed by a nitrogen-containing heterocyclic linked to a carbonyl group. Ding et al. (2022) showed that the lignin β-O-4 polymer was converted to synthesize quinazolinone derivatives [45]. This may result in C-O (β-O-4) bond cleavage and a significant increase of 4(3H)-quinazolinone in AMM stems after L. sajor-caju fermentation. The dihydrocarvyl acetate is a monoterpenoid with antioxidant activity. The contents released by monoterpenoids increase when fungi grow into fermented substrate compared to unfermented substrate [46]. This result is consistent with our research results. In addition, the higher amount of monoterpenoids may result from being released from the substrate and generated by the fungi. Jaceoside is a pharmacologically effective flavone and is rich in biological activity and pharmaceutical properties [47]. In this study, jaceoside was significantly and well correlated with reducing power, FRAP and TPC. Therefore, the identification of representative differential metabolites contributes to the functional feed evaluation of AMM stems after L. sajor-caju fermentation.
4. Conclusions
In conclusion, the white-rot fungus L. sajor-caju was an effective treatment due to its selectivity for degrading lignin, its increased degradations of DMD and lignocellulose, how it improved in vitro degradation parameters and how it enhanced the antioxidant activity of fermented AMM stems. An in-depth metabolomics analysis of the antioxidant component showed that a total of 624 differential metabolites were identified between the CK group and the LP group, 243 of which were antioxidant components. The 4(3H)-quinazolinone, dihydrocarvyl acetate and jaceoside closely related with TPC, reducing power and FRAP, which were the key metabolic markers with antioxidant properties. This shows that we can potentially promote the development of novel antioxidants and functional feeds with AMM stems fermented for 30 days by L. sajor-caju.
Author Contributions
Conceptualization, C.-L.G.; formal analysis, Y.-Q.W.; investigation, L.-M.C. and L.-L.L.; methodology, H.Z. and L.-L.L.; supervision, C.-L.G.; writing-original draft, Y.-Q.W., C.-L.G. and Y.-C.L.; writing-review and editing, Y.-Q.W. and L.-L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Inner Mongolia autonomous region, China, grant number 2023LHMS03009; 2023LHMS03060; 2021LHBS03003; Basic Research Funds for Universities Directly under Inner Mongolia Autonomous Region, grant number GXKY23Z050; GXKY23Z002.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Li, R.Q.; Yin, M.Z.; Yang, M.; Chu, S.S.; Han, X.J.; Wang, M.J.; Peng, H.S. Developmental anatomy of anomalous structure and classification of commercial specifications and grades of the Astragalus membranaceus var. mongholicus. Microsc. Res. Tech. 2018, 81, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Li, J.; Lyu, X.; Chen, X.M.; Guo, S. Nutritional characterization and untargeted metabolomics of oyster mushroom produced using Astragalus membranaceus var. mongolicus stems and leaves as substrates. Front. Plant Sci. 2022, 13, 802801. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Findlay, J.A. Constituents of Astragalus membranaceus. J. Nat. Prod. 1991, 54, 810–815. [Google Scholar] [CrossRef]
- Auyeung, K.K.; Han, Q.B.; Ko, J.K. Astragalus membranaceus: A review of its protection against inflammation and gastrointestinal cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Noh, H.J.; Choi, J.; Lee, K.H.; Lee, M.H.; Lee, J.H.; Hong, Y.; Lee, S.E.; Kim, S.Y.; Kim, G.S. Anti-inflammatory cycloartane-type saponins of Astragalus membranaceus. Molecules 2013, 18, 3725–3732. [Google Scholar] [CrossRef]
- Li, S.G.; Zhang, Y.Q. Characterization and renal protective effect of a polysaccharide from Astragalus membranaceus. Carbohydr. Polym. 2009, 78, 343–348. [Google Scholar] [CrossRef]
- Cui, R.T.; He, J.C.; Wang, B.; Zhang, F.K.; Chen, G.Y.; Yin, S.S.; Shen, H. Suppressive effect of Astragalus membranaceus bunge on chemical hepatocarcinogenesis in rats. Cancer Chemother. Pharmacol. 2003, 51, 75–80. [Google Scholar] [CrossRef]
- Zhang, G. Investigation of active ingredients from roots and stem leaves of Astragalus mongholicus Bunge. Tianjin Pharm. 2010, 5, 5–7. [Google Scholar]
- Luo, Y.L.; Su, L.; Su, R.; Wang, B.H.; Liu, C.; Wang, Z.G.; Zhao, L.H.; Jin, Y. Effects of Astragalus membranaceus supplementation on oxidative stability of Cashmere goat. Food Sci. Nutr. 2020, 8, 5550–5556. [Google Scholar] [CrossRef]
- Moussaoui, Y.; Ferhi, F.; Elaloui, E.; Ben Salem, R.B.; Belgacem, M.N. Utilisation of Astragalus armatus roots in papermaking. BioResources 2011, 6, 4969–4978. [Google Scholar] [CrossRef]
- Rauf, A.; Shafeeq, A.; Shahzad, K. Delignification of corn straw using the ionic liquid triethylammonium hydrogen sulfate. Chem. Eng. Technol. 2022, 45, 1106–1113. [Google Scholar] [CrossRef]
- Sharma, R.K.; Arora, D.S. Bioprocessing of wheat and paddy straw for their nutritional up-gradation. Bioprocess. Biosyst. Eng. 2014, 37, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.T. Lignin degradation by microorganisms: A review. Biotechnol. Prog. 2022, 38, e3226. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Gou, C.L.; Chen, L.M.; Liao, Y.C.; Zhang, H.; Luo, L.L.; Ji, J.H.; Qi, Y. Solid-state fermentation with white rot fungi (Pleurotus species) improves the chemical composition of highland barley straw as a ruminant feed and enhances in vitro rumen digestibility. J. Fungi 2023, 9, 1156. [Google Scholar] [CrossRef] [PubMed]
- Goering, H.K.; Soest, P.J.V. Forage Fiber Analyses: Apparatus, Reagents, Procedures, and Some Applications. In Agricultural Handbook; Agricultural Research Service, U.S. Department of Agriculture: Washington, DC, USA, 1970; Volume 379, pp. 1–20. [Google Scholar]
- Van Soest, P.J.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Arora, D.S.; Chandra, P. Assay of antioxidant potential of two Aspergillus isolates by different methods under various physio-chemical conditions. Braz. J. Microbiol. 2010, 41, 765–777. [Google Scholar] [CrossRef]
- Chang, L.W.; Yen, W.J.; Huang, S.C.; Duh, P.D. Antioxidant activity of sesame coat. Food Chem. 2002, 78, 347–354. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chen, Y.; Jia, Y.N.; Xue, Z.H.; Chen, Z.Q.; Zhang, M.; Pharkphoom, P.; Yang, S.Y.; Chen, H.X. Chrysophyllum cainito. L alleviates diabetic and complications by playing antioxidant, antiglycation, hypoglycemic roles and the chemical profile analysis. J. Ethnopharmacol. 2021, 281, 114569. [Google Scholar] [CrossRef]
- Zhao, W.; Huang, P.P.; Zhu, Z.D.; Chen, C.; Xu, X.Q. Production of phenolic compounds and antioxidant activity via bioconversion of wheat straw by Inonotus obliquus under submerged fermentation with the aid of a surfactant. J. Sci. Food Agric. 2021, 101, 1021–1029. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrate and antioxidants by means of folin-ciocalteu reagent. Method Enzymol. 1999, 299, 152–178. [Google Scholar]
- Shinya, T.; Iwata, E.; Nakahama, K.; Fukuda, Y.; Hayashi, K.; Nanto, K.; Rosa, A.C.; Kawaoka, A. Transcriptional profiles of hybrid Eucalyptus genotypes with contrasting lignin content reveal that monolignol biosynthesis-related genes regulate wood composition. Front. Plant Sci. 2016, 7, 443. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Lu, Y.; Hu, H.; Xie, F.; Wei, X.; Fan, X. Structural characterization of lignin and its degradation products with spectroscopic methods. J. Spectrosc. 2017, 2017, 1–15. [Google Scholar] [CrossRef]
- Datsomor, O.; Gou-Qi, Z.; Miao, L. Effect of ligninolytic axenic and coculture white-rot fungi on rice straw chemical composition and in vitro fermentation characteristics. Sci. Rep. 2022, 12, 1129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.L.; Wang, F.; Fang, Y.; Zhou, D.W.; Wang, S.P.; Wu, D.Q.; Wang, L.X.; Zhong, R.Z. High-potency white-rot fungal strains and duration of fermentation to optimize corn straw as ruminant feed. Bioresour. Technol. 2020, 312, 123512. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Luo, Y.; Luo, L.L.; Zhang, H.; Liao, Y.C.; Gou, C.L. Enhancement of the nutritional value of fermented corn stover as ruminant feed using the fungi Pleurotus Spp. Sci. Rep. 2021, 11, 11961. [Google Scholar] [CrossRef]
- Ding, C.H.; Wang, X.; Li, M.X. Evaluation of six white-rot fungal pretreatments on corn stover for the production of cellulolytic and ligninolytic enzymes, reducing sugars, and ethanol. Appl. Microbiol. Biotechnol. 2019, 103, 5641–5652. [Google Scholar] [CrossRef]
- Reddy, G.V. Bioconversion of Banana Waste into Protein by Two Pleurotus Species (P. ostreatus and P. sajor-caju). Biotechnological Approach. Ph.D. Thesis, Sardar Patel University, Gujarat, India, 2001. [Google Scholar]
- Ghimire, S. Volatile Fatty Acid Production in Ruminants. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, Virginia, 2015. [Google Scholar]
- Datsomor, O.; Yan, Q.; Wang, K.; Mohamed, S.; Opoku-Mensah, L.; Zhao, G.Q.; Miao, L. Effect of ammoniated and/or basidiomycete white-rot fungi treatment on rice straw proximate composition, cell wall component, and in vitro rumen fermentation characteristics. Fermentation 2022, 8, 228. [Google Scholar] [CrossRef]
- Sufyan, A.; Khan, N.A.; AbuGhazaleh, A.; Ahmad, N.; Tang, S.; Tan, Z. Novel techniques for the mass production of nutritionally improved, fungus-treated lignocellulosic biomass for ruminant nutrition. J. Sci. Food. Agri. 2024, 4, 2215–2224. [Google Scholar] [CrossRef]
- Chakraborty, A.; Majumdar, S.; Bhowal, J. Phytochemical screening and antioxidant and antimicrobial activities of crude extracts of different filamentous fungi. Arch. Microbiol. 2021, 203, 6091–6108. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Kiani, A.; Santhiravel, S.; Holman, B.W.B.; Lauridsen, C.; Dunshea, F.R. The importance of dietary antioxidants on oxidative stress, meat and milk production, and their preservative aspects in farm animals: Antioxidant action, animal health, and product quality—Invited review. Animals 2022, 12, 3279. [Google Scholar] [CrossRef]
- Huang, M.Y.; Lin, K.H.; Lu, C.C.; Chen, L.R.; Hsiung, T.C.; Chang, W.T. The intensity of blue light-emitting diodes influences the antioxidant properties and sugar content of oyster mushrooms (Lentinus sajor-caju). Sci. Hortic. Amst. 2017, 218, 8–13. [Google Scholar] [CrossRef]
- Elhusseiny, S.M.; El-Mahdy, T.S.; Awad, M.F.; Elleboudy, N.S.; Farag, M.M.S.; Aboshanab, K.M.; Yassien, M.A. Antiviral, cytotoxic, and antioxidant activities of three edible Agaricomycetes mushrooms: Pleurotus columbinus, Pleurotus sajor-caju, and Agaricus bisporus. J. Fungi 2021, 7, 645. [Google Scholar] [CrossRef]
- Su, W.Y.; Gao, S.Y.; Zhan, S.J.; Wu, Q.; Chen, G.M.; Han, J.Z.; Lv, X.C.; Rao, P.F.; Ni, L. Evaluation of volatile profile and in vitro antioxidant activity of fermented green tea infusion with Pleurotus sajor-caju (Oyster Mushroom). Front. Nutr. 2022, 9, 865991. [Google Scholar] [CrossRef]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liao, Y.; Gou, C.; Zhang, H.; Chen, L.; Bao, Y. Effect of Lentinus sajor-caju on the chemical composition and antioxidant activity of highland barley straw under solid-state fermentation. Front. Microbiol. 2024, 15, 1365254. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, Y.; Qin, Y.; Shi, H. Influence of microbiota and metabolites on the quality of tobacco during fermentation. BMC Microb. 2020, 20, 356. [Google Scholar] [CrossRef] [PubMed]
- Rehan, M. Biosynthesis of Diverse Class Flavonoids via Shikimate and Phenylpropanoid Pathway. In Bioactive Compounds-Biosynthesis, Characterization and Applications; Zepka, L.Q., Nascimento, T.C.d., Lopes, E.J., Eds.; IntechOpen: London, UK, 2021; p. 75392. [Google Scholar]
- Zhu, W.; Wang, W.F.; Xu, W.C.; Wu, S.; Chen, W.J.; Huang, Y.Y.; Wang, S.P. Influence of thermophilic microorganism on non-volatile metabolites during high-temperature pile-fermentation of Chinese dark tea based on metabolomic analysis. Food Sci. Biotechnol. 2022, 31, 827–841. [Google Scholar] [CrossRef]
- Gan, J.; Feng, Y.; He, Z.; Li, X.; Zhang, H. Correlations between antioxidant activity and alkaloids and phenols of maca (Lepidium meyenii). J. Food Qual. 2017, 2017, 3185945. [Google Scholar] [CrossRef]
- Shao, L.; Zhao, S.; Yang, S.; Zhou, X.; Li, Y.; Li, C.; Chen, D.; Li, Z.; Ouyang, G.; Wang, Z. Design, synthesis, antibacterial evaluation, three-dimensional quantitative structure–activity relationship, and mechanism of novel quinazolinone derivatives. J. Agr. Food Chem. 2023, 9, 3939–3949. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechno. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Ding, Y.M.; Guo, T.L.; Li, Z.W.; Zhang, B.; Kühn, F.E.; Liu, C.; Zhang, J.; Xu, D.Z.; Lei, M.; Zhang, T.; et al. Transition-metal-free synthesis of functionalized quinolines by direct conversion of β-O-4 model compounds. Angew. Chem. 2022, 134, 202206284. [Google Scholar] [CrossRef]
- Mäki, M.; Mali, T.; Hellén, H.; Heinonsalo, J.; Lundell, T.; Bäck, J. Deadwood substrate and species-species interactions determine the release of volatile organic compounds by wood-decaying fungi. Fungal Ecol. 2021, 54, 101106. [Google Scholar] [CrossRef]
- Nageen, B.; Rasul, A.; Hussain, G.; Shah, M.A.; Anwar, H.; Hussain, S.M.; Uddin, M.S.; Sarfraz, I.; Riaz, A.; Selamoglu, Z. Jaceosidin: A natural flavone with versatile pharmacological and biological activities. Curr. Pharm. Des. 2021, 4, 456–466. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).