Abstract
Rubus species are affected by the gall inducer Lasioptera rubi (Diptera, Cecidomyiidae), which produces galls predominantly on the stem but sometimes also on the petiole. The presence of these neo-structures at the stem level can affect their survival and implicit production in raspberry crops. The purpose of the present study is to achieve the first detailed description of the histological and morphological peculiarities of these galls, which present great variability from these points of view. Three morphotypes were described in the galls produced in the stem, depending on the position of the larval chambers and the degree of damage to the vascular system. The histochemical investigations highlighted the presence of reducing sugars and proteins as the main nutrients in the gall tissues, as well as the presence of polyphenols and terpenoids as secondary products of metabolism. Indoleacetic acid was identified near the larval chambers and in the neighboring tissues, its presence having an important role in the changes in the morphogenesis of the secondary xylem in the stem and in inducing the appearance of meristematic nodules in the gall structure.
1. Introduction
Rubus idaeus is native to temperate regions of the Northern Hemisphere from North America (if we consider Rubus strigosus Michx. from North America as a subspecies or a variety of R. idaeus) to Europe and northern Asia but is cultivated and introduced in many temperate regions, including southern Australia, New Zealand and Brazil [1,2,3]. R. idaeus can be found in forests, forests margins, clearings, valleys, meadows, slopes, hedgerows, roadsides, and rural and urban areas, up to 2500 m altitude [4,5,6].
Being widely cultivated can sometimes make it difficult to discern a difference between native and the alien populations. Rubus idaeus is classified as the LC (least concern) status in the Global IUCN Red List [7], primarily due to the global decline of mature individuals and of the quality of the specific habitats [8], with Rubus spp. habitats generally being under pressure from agricultural intensification, the removal of hedges and bushes, silviculture, changes in species composition and invasive species [9]. R. idaeus also has the LC status in some countries: Denmark, Estonia, Germany, Norway, Switzerland, United Kingdom [8]. The appearance includes red raspberries, 1–2 m tall perennials shrubs with brown to reddish brown stems, imparipinnate leaves, usually 5–7 foliolate leaves, and terminal inflorescences as short racemes with 1–1.5 cm in diameter flowers with five white petals. The fruits are an aggregate fruit of numerous red drupelets around the receptacle, the drupelets detached from it, as one piece of fruit with a hollow inside [3,4,5].
The raspberry has been domesticated for fruit production, with references to using the fruits dating from the period of Theophrastus, Dioscorides, Aristotle, Pliny [10]. In traditional herbal medicine, roots, stems, leaves, flowers, immature fruits and ripe fruits were used, especially as an astringent, internally and externally. Raspberry vinegar was used as a home remedy for digestive upsets and sore throats, while raspberry juice prevents gastroenteritis. The volatile components of the fruits have antibacterial activity, and raspberry fruits are rich in antioxidants [10]. In the red raspberry, 10 minerals, 17 vitamins, 11 flavonoids and 8 phenolic acids have been discovered [11]. World fresh raspberry (FAO Code 0547) production in 2018 was 870.209 (947.852 in 2022) tons according to the Food and Agriculture Organization of the United Nations (FAOSTAT), which has almost doubled since 2010, with 70.3% of production in Europe and 27.4% in the Americas. The leading countries are Russia, Mexico, Serbia, Poland and the United States of America [12].
Galls represent neo-formations that appear in various plant organs and whose development is induced by various organisms—bacteria, fungi, mites, worms and insects [13]. Among insects, species of the Cecidomyiidae family induce a wide variety of galls in various plant organs. Cecidomyiidae Macquart, with more than 6650 described species, more than 1800 in Europe [14,15], and an estimated number of more than 1.8 million [16], represent maybe the most diverse animal family. Inside the Cecidomyiidae family, 51% of species are phytophagous, 39% are mycophagous, mycosaprophagous and xylophagous, and 8% are zoophagous [15].
The name “gall midges” comes from the fact that many Cecidomyiidae larvae feed inside plants, producing abnormal outgrowths of the vegetal tissue, named galls. Many species are important pests in agricultural ecosystems. In Europe, 915 species live inside plants, including galls, associated with 280 genera of host plants from 72 families [15]. Lasioptera Meigen include more than 130 species across the world [14], with most of them being located in the Old World, 31 in North America and just 3 in Australia, with the supposition that the origin of the genus is in the Old World, from where few species migrate [14]. Most species of Lasioptera are gall makers, especially in stems, and these galls are usually associated with fungal mycelia (“ambrosia galls”) [14,15]. Thirteen species of gall midge on Rubus are known worldwide, seven in the Palearctic [14]. Lasioptera rubi (Schrank) (raspberry gall midge, raspberry stem gall midge) is a Palearctic Eurosiberian species [15] producing galls on stems of R. idaeus and related species, being the only species of Lasioptera developing in Rubus.
Although many studies have examined the anatomy of galls, most have focused on galls that develop on leaves, with much fewer studies dedicated to the anatomy of stem galls.
The gall inducer determines the alteration in the characteristic morphogenetic pattern for the species and the organ of the plant that is attacked. The response reaction of the plant organism is realized rather in the sense of creating optimal conditions for the development of the parasite than a typical defense reaction against herbivory [17,18]. The anatomical and metabolic peculiarities of galls help us to better understand the way in which the specific interaction between a plant and its parasite takes place. Studying the histochemistry of insect-induced galls in plants is important because it brings additional information regarding the reallocation of nutrients inside the original and unformed tissues, but it also reveals the presence and distribution of defense produced by the host within gall tissues [18,19]. Histochemistry allows the investigation of how insects affect the plant’s biochemical processes, such as altering hormone levels, redirecting nutrients, and triggering abnormal cell growth.
The term “ambrosia” was introduced for the first time by Schmidberger in 1836 to designate a group of fungi that develop mutualistic relationships with some Scolytinae beetles [20]. Later, Neger (1908) [21] used the term “ambrosia” for the galls induced by some species of Cecidomyiidae that cultivate fungi in the larval chambers for feeding.
In this study, we aimed to achieve a detailed description of the gall inducer Lasioptera rubi in addition to morphological, structural and histochemical characterizations of the galls induced by L. rubi mainly in the stems from R. idaeus and secondarily those in the petiole (given the rarity of the latter, such as the reduced degree of damage to the health of the cultivated plant). We followed the identification of the tissues on the basis of which the neo-formed structures of the galls are built and the vascular changes that appear with the building of the galls (especially at the level of the secondary xylem). Also, the changes in the distribution pattern of the primary and secondary metabolism products were highlighted.
2. Materials and Methods
2.1. Gall Sampling, Measurement and Morphometric Analysis
In the period from 2001 to 2024, we collected galls of Lasioptera rubi from Rubus idaeus from Romania. The collecting galls sites map (Figure 1a) was created using Google Earth Pro® (7.3.6.9345 Release, Google, Santa Clara, CA, USA). We collected and measured 295 galls. Measuring was carried out with a Parkside HG10714 digital caliper (OWIM GmbH & Co. KG, Neckarsulm, Baden-Württemberg, Germany) with 0.01 mm accuracy. We measured just mature, full-growth galls with a measuring method similar to Tanasković [22] (Figure 1b).
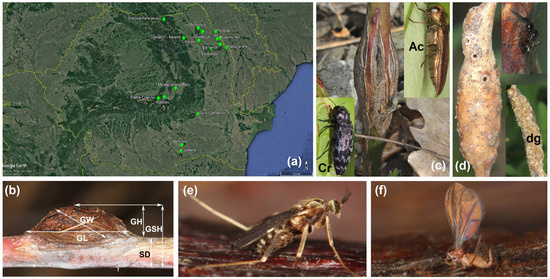
Figure 1.
Rubus idaeus, Lasioptera rubi adults, map of investigated sites, measuring scheme and alike galls: (a)—map of investigated sites, (b)—typical L. rubi gall with measuring scheme (GL—gall length along the stem, GW—gall width, perpendicular to the stem, GH—gall height, from the stem to its edge, GSH—gall height plus stem, SD—stem diameter), (c)—gall of Agrilus cuprescens (Ac); Coraebus rubi (Cr), (d)—gall of Diastrophus rubi with destroyed gall by birds (dg), (e)—adult of L. rubi, (f)—freshly emerged adult of L. rubi.
Images of the live specimens, host plant and galls were taken using a Canon 100 mm f/2.8 Macro USM lens attached to a Canon 60D digital camera (Canon Inc., Tokyo, Japan).
The following parameters were measured: gall length along the stem (GL), gall width, perpendicular to the stem (GW), gall height, from the stem to its edge (GH), gall height plus stem (GSH), stem diameter (SD) (Figure 1b). Considering the general appearance of the galls, a mathematical formula for a hemi-ellipsoid was used to calculate the volume: V = (2/3) × π × (GL/2) × GH × (GW/2).
The remaining galls not used for dissections and voucher specimens from the insects are kept in I. E. Popescu’s personal collection, University “Alexandru Ioan Cuza”, Faculty of Biology, Laboratory of Entomology, Iaşi, Romania (collection acronym: IPCO). The morphological terminology for L. rubi follows that of Möhn [23], Yukawa [24] and Thomas and Goolsby [25].
2.2. Light Microscopy
The galls (n = 50) were dissected using razor blades and micro dissecting needles under a Euromex stereomicroscope (Euromex Microscopen BV, Arnhem, The Netherlands) with a maximum magnification of 180×. Images of the dissected galls, larvae, pupa, pupal exuviae and mycelium were captured using a Canon 60D digital camera connected to a ZEISS SteREO Discovery.V20 stereomicroscope (ZEISS Microscopy, Oberkochen, Germany). Both stereomicroscopes were used for observations and descriptions of galls and insects. The images were processed with Helicon Focus 8.1.0® (Helicon Soft Ltd., Kharkov, Ukraine) and their clarity was enhanced using Adobe® Photoshop® CC (20.0.0 Release, Adobe, San Jose, CA, USA).
2.3. Histo-Anatomical Analysis
Samples of the non-galled and galled stems (n = 25) and petioles (n = 5) were preserved in 70% ethyl alcohol. Cross-sections were performed using a manual microtome and a razor blade. The sections were stained either with ruthenium red (0.05%) and iodine green [26,27], with toluidine blue at 0.05%, pH 4.7 [28], or with Astra blue 1% aqueous solution [29].
Photographs were taken with a 60D digital camera (Canon Inc., Tokyo, Japan), using an Olympus BX41 (Olympus Corporation, Tokyo, Japan) research microscope. Measurements for anatomical parameters, including xylem vessel diameter (n = 50), secondary xylem ring width (n = 25) and xylem vessel density (n = 15), were carried out using IC Measure Imaging Source software 2.0.0.286. (The Imaging Source, LLC, Charlotte, NC, USA). The measurements were carried out in 3 areas: non-galled stem, galled stem (in the vicinity of the neo-formed tissue) and unmodified galled stem (in the region of the stem that is not affected by gall induction).
2.4. Scanning Electron Microscopy (SEM)
The plant material (galled stems) was fixed and stored in 70% ethanol. Also, L. rubi larvae, extracted from galls, were fixed in 70% ethyl alcohol. Galls were sectioned under the stereomicroscope, with a razor blade. After dehydration in a graded ethanol series (80%, 90% and 100%) and acetone, the material was critical point dried with CO2 (using an EMS 850 Critical Point Dryer, Hatfield, PA, USA), mounted on stubs with carbon conductive tape, and sputter coated with a thin layer of gold (30 nm) using an sputter coater (EMS 550X Sputter Coater, Hatfield, PA, USA). Samples were examined with the Tescan Vega II SBH scanning electron microscope (TESCAN, Brno, Czech Republic), from the Electron Microscopy Laboratory, Faculty of Biology, “Alexandru Ioan Cuza” University of Iaşi, at an acceleration voltage of 30 kV.
2.5. Histochemical Analysis
Cross-sections performed through fresh galls (n = 10) immediately after harvesting were subjected to histochemical analyses.
To detect starch, Lugol reagent (1% potassium iodine–iodide solution) was applied for 5 min [30]. Total lipids were detected with a saturated Sudan Red solution in 70% ethanol for 10 min [31]. Reducing sugars were detected by Fehling’s reagent (solution A—7.9% copper sulfate and solution B—34.6% potassium sodium tartrate and 1% sodium hydroxide), followed by heating [32]. The NADI reagent for terpenes was 1% α-naphthol and 1% dimethyl-p-phenylenediamine [33]. Proteins were detected with Coomassie brilliant blue [34]. Ferric chloride 1% was used for 5 min for the phenolic compounds [34]. For lignins, acidified phloroglucinol (solution A—2% phloroglucin and solution B—25% hydrochloric acid) was applied for 5 min [30]. The presence of IAA (indoleacetic acid) was detected with the Ehrlich reagent (% p-dimethylaminno-benzaldehyde in 1 M HCl) [35]. Unstained sections were used as controls.
2.6. Statistical Analysis
Excel 2010 was used for test data entry; Analyse-it, Standard Edition (version 6.15.4) (Analyse-it Software, Ltd., Leeds, United Kingdom) was used to create graphs regarding the distribution of the values of the morphological parameters of the galls. A one-way ANOVA was used to compare differences between the means of the anatomical parameters, regarding the secondary xylem.
3. Results
3.1. Gall Inducer—Lasioptera rubi
The length of the adults of L. rubi (Figure 1e,f) is around 1.5–3 mm, with a brown and black body with silver spots and stripes. The wing length of L. rubi is 1.8–2.1 mm, 2.3–2.6 as long as broad, the R5 vein is straight and close to the C vein, not curved in the middle, the flagellum has 15–17 articles in males and 18–23 in females, the fifth article is as long as wide or slightly shorter than wide, and the palpus has four articles, where the first one is shortest and slightly longer than wide. The second tarsomeres of the fore and middle legs are shorter than the tibia and nearly as long as the tibia, or slightly longer on the hind legs. The claws are bifid on all legs, with the empodium as long or slightly shorter than the claws. The eighth to the ninth segments of the abdomen from the female ovipositor have microtrichia (cuticular external processes without innervation). The ninth and tenth segments (uromere nine and ten) have mostly ventral, nonporous sensory hairs. Dorsolateral on each side of the uromere seven there are 30–40 nonporous bristles. The superior lamella has spines (thorn-like extensions of the cuticle without innervation), uniporous sensory hairs (dorsally and apically) and 7–11 scoop-like sensilla (or fishhook-like), each one being terminally expanded containing three dendrites. Conidia are entrapped by the nonporous bristles and the spines, with uromere ten having a mycangial pouch containing the conidia, beneath the superior lamella. These structures, used for collecting and transporting the fungus, are called mycangia [25,36,37].
The L. rubi has a life cycle with one generation a year. The adults of L. rubi emerge from the overwintering galls in spring, from April-May to June-July. Freshly emerged females of L. rubi seem to not have conidia [36], as in the case of L. arundinis [37,38,39]. Freshly emerged females probably collect mycelium from the plants, like in the case of L. arundinis [37,38,39], but it is not yet exactly known how this occurs in the case of L. rubi. Females lay eggs in May-June, in groups from a few to a few dozen, on the surface of canes, close to developing flower buds, sometimes on the lateral shoots or on the petioles. The first instar larva appears after 8–10 days [22]. Most probably, the mycelium from the mycangia covers the corium of the eggs in the moment of the oviposition. L. rubi has three larval stages which develop as endophagous inside the stems of Rubus species, R. idaeus in this case, producing galls visible from the exterior of the host plant. As in L. arundinis, the newly hatched larvae likely puncture the epidermal cells with their mandibles, facilitating the entry of the fungus into the plant and lysing the middle lamellae of the cells. The hyphae induce cell dissociation, creating spaces where the moving larvae inoculate the mycelium using the long spines on their bodies, forming tunnels in the process. [37]. The larvae from the first instar are white and begin to make tunnels in the plant host tissues (Figure 2a) in May–June, with the first exterior effects being visible on the stems of R. idaeus, appearing as very small black dots with a shallow depression and small deformations on the surface of the stem. Larvae are somewhat gregarious, feeding in a group, coming from the same mass of eggs having the same larval stage inside the gall. The body is cylindrical, elongated, and typical for a stem-boring insect larva. During the first larval instar, the mycelium is not macroscopically visible in the gall (Figure 2a). The cuticle is transparent, and some conspicuous orange-brown masses can be visible inside the first larval instar which is the remains of the yolk proteins from the eggs and some tissue consumed from the plant host. To the anterior part of the larva, Bolwig’s organ can be visible (Figure 2a), which can detect light, having a role in the larva’s behavior [25].

Figure 2.
Lasioptera rubi larvae and pupa: (a)—gall with first instar larvae with tunnels (black arrow), larva (lv), Bolwig’s organ (red arrow); (b)—gall with second instar larvae and mycelium with hole (h), cavity (c), larva (lv), mycelium (m); (c)—third instar larva ventral with sternal spatula (white arrow); (d)—third instar larva dorsal; (e)—third instar larva lateral with sternal spatula (white arrow); (f)—pupa in gall, (g)—pupa lateral and ventral.
The tegument has dentate and spinulate microverrucae and long sensilla trichodea (Figure 3c–f). All these structures help in disseminating the fungus inside the tunnels in the tissue of the gall. The first instar larva does not have a sternal spatula. Some capitate sensilla trichodea and middle fusiform sensilla trichodea can also be observed on the cuticle (Figure 3e,f). The development of the first instar larva can be correlated with the visibility of the galls in June-July, even in August (Figure 4a–i and Figure 5a–d), generally 3–6 weeks after oviposition of the eggs [22], but the timing can be different and can correlate with early or late spring every year. The second instar larva (Figure 2b and Figure 3g–i) also does not have a sternal spatula and the cuticle is transparent, but the color is more yellowish, and the antennae are more elongated (Figure 3i). During the second larval instar, the mycelium is macroscopically visible in the galls, which can have big holes, irregular cavities and tunnels covered with the mycelium, with the larva staying in the mycelium and feeding from it (Figure 2b and Figure 3g).
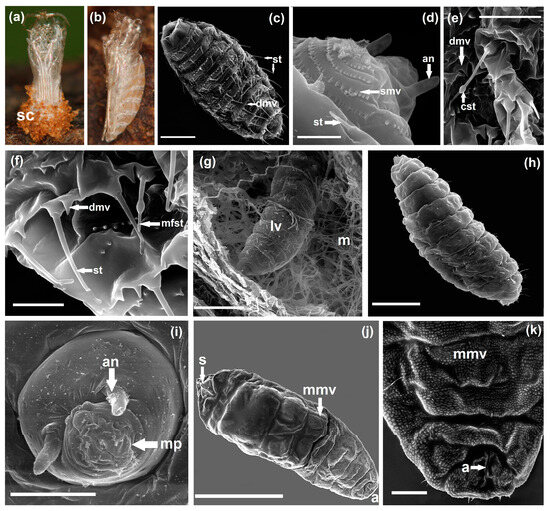
Figure 3.
Lasioptera rubi pupal exuviae and SEM micrographs of larvae (a)—pupal exuviae ventral on gall with silken cocoon (sc), (b)—pupal exuviae lateral, (c)—first instar larva with sensilla trichodea (st), dentate microverrucae (dmv) (scale bar 50 µm), (d)—first segments of first instar larva with antenna (an), spinulate microverrucae (smv), sensilla trichodea (st) (scale bar 10 µm), (e)—tegument detail of first instar larva with capitate sensilla trichodea (cst), dentate microverrucae (dmv) (scale bar 20 µm), (f)—tegument detail of first instar larva with sensilla trichodea (st), middle fusiform sensilla trichodea (mfst), dentate microverrucae (dmv) (scale bar 10 µm), (g)—second instar larva (lv) in gall with mycelium (m) (scale bar 100 µm), (h)—second instar larva (scale bar 100 µm), (i)—second instar larva head with antenna (an), mouthparts (mp) (scale bar 20 µm), (j)—third instar larva with sternal spatula (s), minutely microverrucae (mmv), anus (a) (scale bar 1 mm), (k)—terminal part of third instar larva with minutely microverrucae (mmv), anus (a) (scale bar 200 µm).

Figure 4.
Lasioptera rubi galls: (a)—first visible signs of future galls, (b)—small gall in progress, (c)—typical gall, (d)—multiple galls in stem and petiole, (e)—round gall in stem, (f)—gall in node at the base of lateral canes, (g)—gall ending a lateral cane, (h)—galls in petioles, (i)—gall at the base of a petiole and a puncture mark of another future gall (pm).

Figure 5.
Lasioptera rubi galls: (a)—cracking gall at the base of a stem, (b–d)—cracking galls, (e)—contorted gall, (f)—atypical galls.
The body of the third instar larva is hemicephalic, asymmetrically fusiform, and compressed dorsoventrally, with most of the segments swelling laterally; it is more of a yellow to red-orange color (Figure 2c–e and Figure 3j,k). The epiderma of the third instar larva is without the macropapillae, projections and microdentition observed in the first and second instar. The cuticle is minutely verrucose (cuticular verrucae) (Figure 3j,k), with these structures making transversal lines, probably with the role of the traction ridges, on the ventral side [25]. On the ventral side of the prothorax, there is a sternal spatula, a protuberant sturdily sclerotized armature protruding from the body, and a bilobed with two strong forwardly directed teeth (Figure 2c,e). Internal muscles are attached to the round base (manubrium) of the sternal spatula. The sternal spatula can be used for perforating and enlarging the plant tissues and for making the escaping tunnel for the adults. It can also be used as an extraoral masticating organ for scraping and scratching the mycelium mixed with excreted digestive fluids, with the nutrients obtained like this being sucked in a process of extraoral digestion [25,38].
L. rubi has a third larval stage inside the galls over winter in a silk cocoon, made with the silk produced by the tubular salivary glands [25]. The overwintering larva is not damaged by the fungus mycelium and generally looks to not be negatively affected by the humid conditions in winter. In other situations, the generally humid conditions in winter can in fact help the emergence of the adult insect by moisturizing the galls, but in this case the third instar larva makes an escaping tunnel with the sternal spatula close to the surface of the gall, with just a very thin epidermal tissue surface as a barrier to the outside, which is easy to break by the adult to emerge from the gall in spring. It is also possible that the fungus protects the insect larva against some parasites [38]. The pupa is exarate, without evident apical spines, enclosed in a silken cocoon, with an orange tegument before the formation of the imago (Figure 2f). The full mature pupa is darker (Figure 2g), with the wing pads overlapping the metafemurs. When the adult emerges from the gall, it leaves the pupal exuviae (Figure 3b) protruding from the exit hole with the rest of the silken cocoon at the base, impregnated with wooden debris as a result of the process of drilling the exit tunnel (Figure 3a). The emergence of the fresh adult (Figure 1f) takes around 5–15 min [40].
3.2. Stem Galls’ Morphology and Morphometry
The galls induced by L. rubi at the level of the stems show great diversity, both from the morphological and dimensional point of view.
The initial zone where the larvae enter in the stems has the appearance of small black dots with a thin depression and a small deformation on the surface of the canes (Figure 4a). After that, a light brown area appears with a more visible deformation of the cane (Figure 4b). The typical young gall produced by L. rubi looks like a lateral light brown deformation of the cane on the one side of the stem, with the visible green epidermis cracking on the surface of the gall (Figure 4c). A typical mature gall is brown to dark brown with numerous longitudinal fissures and cracks, sometimes holes, on the surface (Figure 1b).
The galls can be localized on the principal stem or on the lateral canes from the base of the plant a few cm above the soil (Figure 5a) to 1–2 m above the soil. The galls can be dispersed on the canes or can be localized side by side, very close each other (Figure 4d), depending on where the adults lay the eggs. It is common situation to find two galls almost agglutinated each other, as a result of eggs being laid very close together. Sometimes, the eggs can be laid very close to a formed gall (Figure 4i). As a general conclusion, it does not seem that L. rubi disperse the eggs too much or avoid canes with formed galls. Sometimes, the galls can be rounded, encapsulating the stem (Figure 4e), or can be in a node, at the base of lateral canes and petioles (Figure 4f) or even at the end of a lateral cane (Figure 4g).
Galls can also be located inside the petioles, at the base of a petiole, in the middle or at the end, usually as a rhombic gall with green stripes of the epidermis. They sometimes appear as a rounded gall or even as a prolonged gall (Figure 4h,i). Cracked galls can generally be found on the main stems from the base of the plants up to a 1–1.5 m distance from the soil, but also on the lateral canes (Figure 5a–d). Sometime, the galls can distort the stem by totally bending it (Figure 5e), or the galls can have an atypical aspect (Figure 5f).
The gall length (GL) has an average of 15.9 ± 5.41 mm, with a minimum of 6.72 mm and a maximum of 44.71 mm. The gall width (GW), measured perpendicular to the stem, has an average of 9.70 ± 2.74 mm, with a minimum of 3.07 mm and a maximum of 24.42 mm. The gall height (GH), measured from the stem to its edge, has an average of 4.56 ± 2.01 mm, with a minimum of 0.75 mm and a maximum of 17.07 mm. The gall height plus stem (GSH) has an average of 9.38± 2.69 mm, with a minimum of 3.57 mm and a maximum of 19.19 mm.
The gall volume has an average of 422.49 ± 407.33 mm3, with a minimum of 53.81 mm3 and a maximum of 3964.63 mm3. The distribution of the values of the investigated parameters is presented in Figure 6.
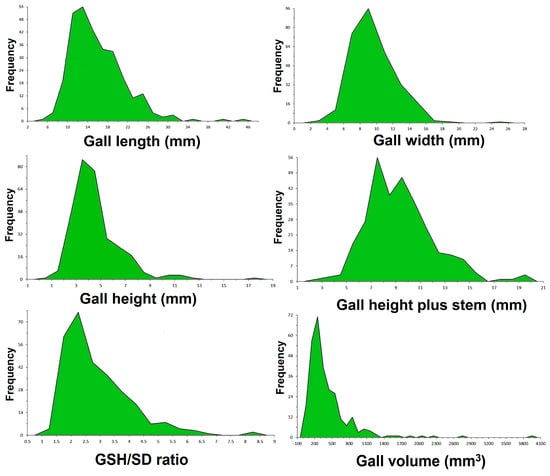
Figure 6.
Graphical representation of the distribution of the values recorded in the measurements of the biometric parameters of the galls on the stem.
The galls of L. rubi can be superficially confused with the galls produced by Diastrophus rubi (Bouché) (Hymenoptera: Cynipidae) in the stems of Rubus idaeus, but the galls of D. rubi are more elongated with a smooth surface, without the longitudinal fissures, cracks, or even holes that are featured on the surface of the galls of L. rubi (Figure 1d). In the interior, each larva has its own chamber, and the irregular cavities, tunnels and holes that can be seen on the inside of L. rubi galls are not present. It is normal to find galls of D. rubi destroyed by birds who consume the larvae (Figure 1d). We have never found a gall of L. rubi with the larvae consumed by birds. Sometimes, the larva of Agrilus cuprescens (Ménétries) (Coleoptera: Buprestidae) can produce fusiform galls at the base of R. idaeus plants (Figure 1c), with just one larva inside a single chamber. Also, from Buprestidae, we frequently find adults of Coraebus rubi (Linnaeus) on the plants of R. idaeus (Figure 1c), but this species does not produce visible galls.
3.3. Galls’ Anatomy
3.3.1. Non-Galled Stems Structure
The structure of the current year’s stems is secondary, formed mainly through the activity of the cambium. The contour of the cross-section is circular and slightly ribbed. The phellogen appears from the last layers of the cortical parenchyma, at the edge of the central cylinder. The conducting tissues have a typical secondary structure, arranged in the form of rings (of xylem and secondary phloem). On the periphery of the phloem, bundles of sclerenchyma are visible. The secondary phloem is thick, the cambium is thin, and the secondary xylem is composed of libriform fibers, dispersed vessels, parenchyma cells and sclerified medullary rays. The pith is compact, with parenchyma cells.
3.3.2. Non-Galled Petiole Structure
The non-galled petiole structure has a triangular outline, with a deep groove on the adaxial face. Under the epidermis, columns of tangential collenchyma can be observed in the three ribs. The vascular system consists of 3–5 fascicles—a larger central one and 2–4 smaller lateral ones with the primary structure.
3.3.3. Galled Stems’ Structure
Galls present an anatomical structure with increased variability, and the final appearance is determined by the place where the larval chambers develop.
In the case of the stems, we can distinguish three morphotypes: GL 1—when the gall is asymmetrical, affecting only one part of the stem and the larval chambers do not touch the pith; GL 2—when the gall is asymmetric but the pith is affected and invaded by neo-formed tissue; GL 3—when the gall develops around the stem, completely affecting the conducting tissue and the pith.
- Anatomical structure of GL 1 galls
The most frequently encountered galls are located laterally on the stem, which affect only part of its structure (Figure 7a).
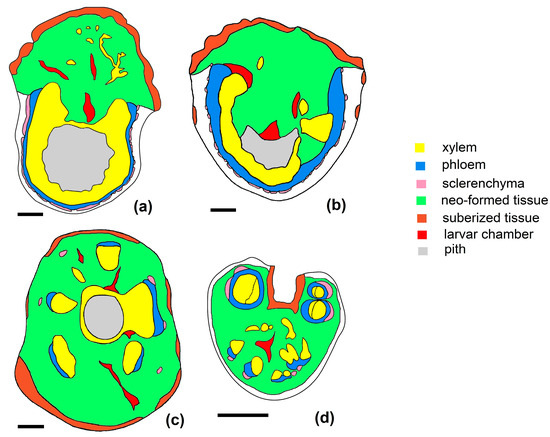
Figure 7.
Diagrams with the structures of galls produced by L. rubi on the stems and petiole: (a–c) galls from the stem: (a)—morphotype GL1, (b)—morphotype GL2, (c)—morphotype GL3, (d)—galls in the petiole (scale bar 1 mm).
The activity of cambium and phellogen is stimulated. The cambium produces modified vascular elements even in the distal areas, in the vicinity of the gall. Also, in the vicinity of the gall, the amount of secondary xylem and secondary phloem generated is increased compared to the unaffected area of the stem. The structure of the galls is relatively compact, consisting of neo-formed tissue in which numerous larval chambers can be observed, irregular in number and distribution (Figure 8a–c). The neo-formed tissue is compact, made up of cells with small air spaces between them, many of them with visible dividing walls. Isolated xylem elements, with a modified structure, can sometimes be observed in the mass of neo-formed tissue. Meristematic nodules made of small cells are also visible (Figure 8b).
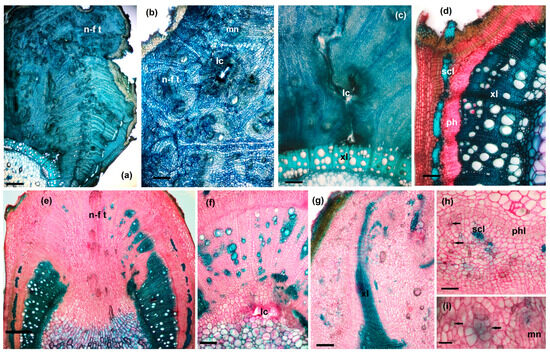
Figure 8.
Cross-sections through galled stems: (a–d) morphotype GL1: (a)—general view (scale bar 500 µm), (b)—detail from neo-formed tissue (scale bar 200 µm), (c)—detail from the internal area, with secondary xylem, and larval chamber (scale bar 100 µm), (d)—detail from the lateral area; (e–h) morphotype GL 2: (e)—general view (scale bar 500 µm), (f)—detail from the internal area and larval chamber (scale bar 200 µm), (g)—detail from the lateral area (scale bar 200 µm), (h)—detail from the external cortical area (scale bar 100 µm); (i)—detail from neo-formed tissue (scale bar 100 µm): lc—larval chamber, mn—meristematic nodules, n-f t—neo-formed tissue, ph—phloem, phl—phelogen, scl – sclerenchyma, xl – xylem, black arrows—calcium oxalate crystals.
The induction of the new formation of meristematic-type tissues is also observed around the perivascular sclerenchyma. Between this and the secondary phloem, the development of a tissue similar to phellogen is visible. After the gall grows, radically changing the structure of the stem, this phellogen-like tissue is found grouped around sclerenchyma fibers now isolated at the edge of the gall (Figure 8d).
With the initiation of gall formation, xylem development is deeply disrupted, with lignification being inhibited in the area where gall develops. The effect of inhibiting lignin production is observed to decrease towards the edges of the gall.
The larval chambers are numerous (multilocular gall) and arranged randomly (in neo-formed tissue, in modified secondary xylem or in pith). They are lined in the first layers of the neo-formed tissue consisting of rounded, thin-walled cells, which later become necrotic and serve as nutrient materials for the mycelium of the fungus that grows in the mature larval chamber. The mycelium of the fungus penetrates very little in the areas adjacent to the larval chamber, in the living cells (Figure 9a,c,d). Fungal hyphae are observed in the first 2–4 layers of the parenchyma cells (Figure 9e,f) in the vicinity of the larval chamber and sometimes in the nearby wooden vessels (Figure 9b). But, we have never identified mycelium in the deep areas of the gall, in neo-formed tissues.

Figure 9.
Microscopic aspects of ambrosia galls fungi induced by L. rubi in R. idaeus stems: (a–c) light microscopy images: (a)—larval chamber developed in modified secondary xylem (scale bar 100 µm), (b)—fungal hyphae located in a secondary xylem vessel (arrows) (scale bar 50 µm), (c)—fungal hyphae in the larval chamber (stained with Astra Blue) (scale bar 50 µm); (d–f) scanning electron microscopy (SEM) images: (d)—chamber larva near the pith (scale bar 100 µm), (e)—detail of the parenchyma tissue of the pith, adjacent to the larval chamber (scale bar 100 µm), (f)—pith cell with fungal hyphae (arrows) (scale bar 20 µm): lc—larval chamber.
In the second year of life, the stems that present lateral galls of the GL 1 type usually maintain their viability; this is due to the intact maintenance of the vascular tissue structure in half of the stem structure. Also, their hypertrophy in the vicinity of the gall replaces the sap transport that is suppressed in the gall area.
- Anatomical structure of GL 2 galls
The structure of GL 2 galls differs from that of GL 1 galls, primarily by complete interruption of the continuity of the conducting tissue rings (Figure 7b). The xylem and secondary phloem elements, as well as sclerenchyma bundles, are pushed to the periphery of the gall by the newly formed tissue. A large part of the pith is also replaced by the neo-formed tissue, and the larval chambers can also be found inside it (Figure 8e,f). As a consequence, only a part of the stem retains a normal structure, which leads to a weakening of its viability. The secondary xylem also extends into the neo-formed tissue area, ensuring the vascularization of the gall and the transport of nutrients to it (Figure 8g).
From the activity of the hypertrophied phellogen, neo-formed tissue is also generated in the cortical area (Figure 8g,h). And in these new tissues, as in the unaffected stems, numerous calcium oxalate druses can be observed (Figure 8h,i).
- Anatomical structure of GL 3 galls
In the case of the galls that completely surround the entire stem at maturity, the normal structure is disturbed by the tissues formed following larval induction (Figure 7c). The xylem appears in separate areas and is small in size (Figure 10a); after larval induction, its development is inhibited, unlike the other two gall morphotypes where, in their vicinity, xylem formation is stimulated.
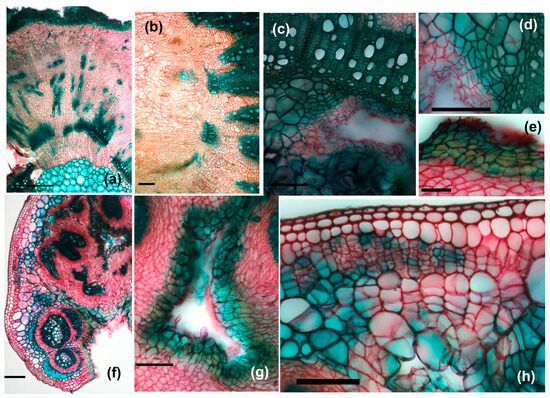
Figure 10.
Cross-sections through galled stems: (a–e) morphotype GL3: (a)—general view (scale bar 500 µm), (b)—detail of the central area with neo-formed tissue (scale bar 100 µm); (c,d)—details of internal area with larval chamber (scale bar 100 µm), (e)—detail of the external area (scale bar 50 µm); (f–h)—cross-sections through petiole galls, (f)—overview (scale bar 200 µm), (g)—detail of larval chamber (scale bar 100 µm), (h)—detail from the external area (scale bar 100 µm).
Hypertrophy and hyperplasia of neo-formed tissue lead to the disintegration of the stem structure, through the fragmentation of the xylem masses already formed at the time of larval induction. Meristematic nodules are also observed in this case (Figure 10b). In isolated cases, we reported the presence of an organized meristematic tissue in the vicinity of the larval chambers (Figure 10c,d). The induction of the pre-existing parenchymal cells’ de-differentiation is intense; the division walls can be observed in the already slightly sclerified pith cells (Figure 10d).
At the periphery of the gall, with the growth of its dimensions, the epidermis of the stem appears interrupted, with the neo-formed tissue coming into direct contact with the external environment. In order to protect the gall, scar tissue is built up over time, consisting of dead cells at maturity with slightly suberized walls (Figure 10e).
3.3.4. Petiole Galls’ Structure
Inducing gall development in the petiole leads to the disorganization of its normal structure in large part or even completely (Figure 7d and Figure 10f). With small dimensions, the proliferation of the unformed tissue changes the position of the conducting bundles, which can become fragmented. Cell divisions are initiated in the fundamental parenchyma of the petiole both in the external region (between the epidermis or collenchyma and vascular bundles) (Figure 10h), around the vascular bundles, and in the central part (Figure 10g). These lead to the appearance of hyperplastic tissue, in which meristematic nodules are organized. Some of them give birth to neo-formed conductive elements. The larval chambers are 2–5 in number, and due to insufficient space—despite the proliferation of parenchymal cells—some of them are open, in direct contact with the external environment.
3.3.5. Changes in Vascular Tissue Morphogenesis Induced by Gall Formation
With the growth of gall, the conducting tissues in the area adjacent to it are deeply affected. The changes aim at the appearance of some discontinuities in its structure (mainly caused by the appearance of neo-formed tissue) and structural changes (in the morphogenesis of the secondary xylem in particular), all of which affect its functionality (Figure 11).
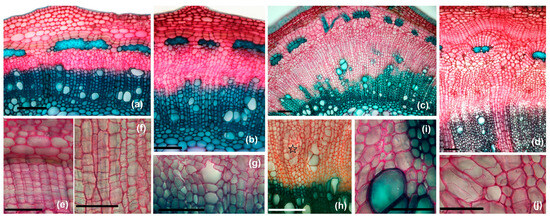
Figure 11.
Changes induced by the induction of galls at the level of the vascular and meristematic systems in the stem: (a)—cross-section through normal stem (scale bar 100 µm), (b)—cross-section through gall margin—beginning of stimulation of cambium activity (scale bar 100 µm), (c)—cross-section through the gall; modified secondary xylem begins to result from the activity of the cambium (scale bar 100 µm), (d)—cross-section through the gall; modified secondary conductive tissues and the appearance of a phellogen-like tissue around the perivascular sclerenchyma can be observed (scale bar 100 µm), (e)—hypertrophied phellogen (scale bar 50 µm), (f)—hypertrophied cambium (scale bar 50 µm), (g)—non-lignified secondary wood vessel (scale bar 50 µm), (h)—emergence of an area with intense divisions in the modified secondary xylem (star) (scale bar 100 µm), (i)—non-lignified xylem fibers (scale bar 50 µm), (j)—hypertrophied wood parenchyma cells, with multiple division walls (arrows) (scale bar 50 µm).
The structure of the xylem within the gall is significantly altered due to the appearance of abnormal tissue associated with gall formation. The xylem vessels undergo hypertrophy (enlargement) and hyperplasia (increased cell number, especially concerning parenchyma and xylem fibers) (Figure 11a–d) and the formation of irregular, swollen or deformed xylem vessels is observed (Figure 11g,i); at the same time, the xylem elements (especially the fibers) no longer lignify their cell walls after larval induction (Figure 11i). These changes cause the xylem to lose its typical structure, which is crucial for efficient sap transport. The altered structure may also result in increased resistance to water flow, reducing the overall efficiency of the xylem.
The results of the histometric study are presented in Figure 12.
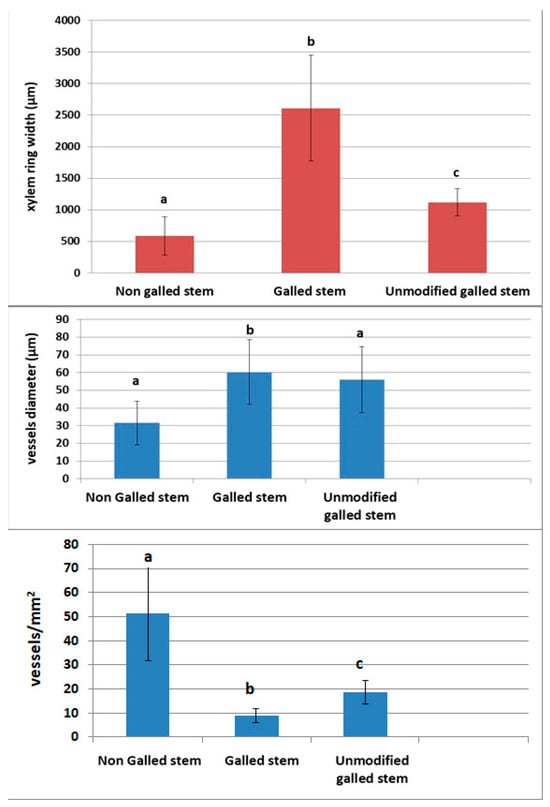
Figure 12.
Histometric analyses of the anatomical parameters of secondary xylem; secondary xylem ring width, xylem vessels diameter, xylem vessels density (means ± SD); bars with different letters are significantly different (p < 0.05, one-way ANOVA).
The width of the secondary xylem ring increases by 443.45% in the area in the vicinity of the gall (p < 0.001) compared to the non-galled stem and by 232.92% compared to the unmodified xylem area in the gall (p < 0.001). Also, the diameter of the secondary xylem vessels in the gall increases by 191.2% compared to those in the normal stem (p < 0.001), but there are no significant differences between them and the vessels in the unmodified area (p = 0.2511). The variation in the density of the xylem vessels is opposite to the size, with those in the normal stem being 5.72 times (p < 0.001) denser than those in the vicinity of the gall and 2.75 times (p < 0.001) denser than those in the unmodified area of the gall.
3.4. Histochemical Peculiarities of the Gall Tissues
The histochemical tests carried out on the investigated galls were positive in the majority of cases (with the exception of the Sudan Red 3 test, which identified only very small amounts lipids in the neo- formed tissues of the gall structure (Figure 13c); it was positive for the cuticle (Figure 13d) and suberized cells.
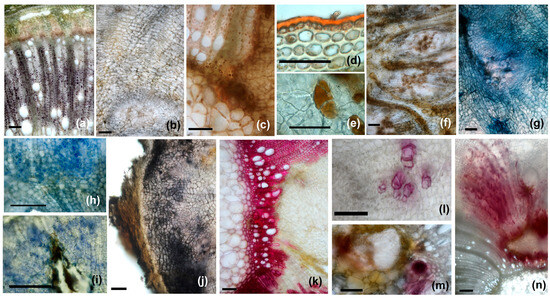
Figure 13.
Histochemical profiles of the galls: (a,b)—starch stained in purple with Lugol’s reagent ((a)—scale bar 200 µm, (b)—scale bar 100 µm), (c,d)—lipids stained in red with Sudan Red (weak positive reaction) ((c)—scale bar 100 µm, (d)—scale bar 50 µm), (e,f)—reducing sugars stained in reddish brown with Fehling’s reagent ((e)—scale bar 5 µm, (f)—scale bar 100 µm), (g)—terpenes stained in dark blue with NADI reagent (scale bar 100 µm), (h,i)—proteins stained in blue with Coomassie brilliant blue (scale bar 50 µm), (j)—phenolic compounds stained in dark violet with Ferric chloride (scale bar 100 µm), (k,l)—lignin stained in purple red with phloroglucinol (scale bar 100 µm), (m,n)—IAA (indoleacetic acid) stained in burgundy with Ehrlich reagent ((m)—scale bar 100 µm, (n)—scale bar 200 µm).
Starch was identified with Lugol’s solution in the wood parenchyma (Figure 13a) cells and very rarely and in small amounts in the neo-formed parenchyma (Figure 13b). In non-galled stems, starch is present in large quantities in the pith near the primary xylem. At the gall level, it is no longer found in these areas. On the other hand, reducing sugars, detected by Fehling’s reagent (Figure 13e,f), were identified in large quantities in all morphotypes of galls, both those in the stem and those in the petiole. If non-galled stems are moderately positive for reduced carbohydrates, with these being found in the cortex and parenchyma of the secondary phloem, increased amounts are observed both in the peripheral area and in the nutritious tissue at the level of the galls. In GL 3-type galls, they are found in large quantities and in neo-formed tissue in the central area, which takes the place of the pith, while in GL1- and GL2-type galls, the reduced sugars in the pith are located especially in small cells (as in non-galled stems).
The NADI reagent used for the identification of terpenoids led to positive reactions in the neo-formed parenchyma of the galls, especially near the larval chambers, and also in vascular tissues (Figure 13g). The proteins, identified with Coommasie brilliant blue, are mainly located in the meristematic nodules and are found in neo-formed tissue from all the morphotypes of the range analyzed (Figure 13h,i).
Intensely positive reactions were observed in all the galls tested with ferric chloride for phenolic compounds (Figure 13j). They were identified in neo-formed tissues, especially in the outer and middle part, as well as around the larval chambers. Lignin was intensely stained purple-red with phloroglucinol at the level of primary and secondary xylem elements formed before larval induction and was weaker in those not formed after this moment (Figure 13k,l).
IAA (indoleacetic acid) was detected only in the galls, both in the petiole and in the stem, especially in the neo-formed tissue, around the larval chamber (Figure 13m,n). However, its distribution is not uniform, and it is most often linked to the disposition of the larval chambers; sometimes, IAA was observed on the periphery of the galls, in the areas with phellogen-like meristematic tissue.
The results are summarized in Table 1.

Table 1.
Histochemical analysis of the gall tissues.
4. Discussion
From 80 worldwide pests in raspberry plantations, 25 of these species are insects [22]. For Europe, 22 arthropod pests are listed as being associated with cultivated Rubus, including Lasioptera rubi [41], which is considered to be a specific pest for the raspberry [6]. Damage to the raspberry by L. rubi has been mentioned dating back to 1898 (“damage in 1898 very apparent, few leaves and no fruit”), 1912 (“not only the young shoots die, but the canes become liable to be attacked by a fungus, large numbers in past few years”) and 1925 (“causes the stem to droop”) [42], and in 1948, it was noted in a series of eight volumes dedicated to “Gall Midges of Economic Importance” [43]. In 1978, Viggiani and Mazzone [44] found an infestation in southern Italy, in the Campania region, with 70–80% damaged canes resulting in small leaves and fruits, likely in addition to infection from the damaging fungi Didymella applanate (raspberry spur blight). But still, some authors consider L. rubi as a “minor pest” [41,45,46], stating that “it should not be regarded as a serious pest” as it only causes “cosmetic damage” [41].
The reality is that in recent years, L. rubi has become a serious pest for raspberry crops. Serbia is the third country in the world in terms of raspberry production, with “red gold” being the name of the fresh raspberry here; most of the harvest goes to export. In Serbia, the increased incidence of these pests is very well documented, where they were considered to be “little known” in the 1960s and 1970s but, since 2000, have been viewed as an “economically important pest” and even “extremely serious economic pests” [47,48,49,50,51,52,53,54]. The average infestation of canes in Serbia is between 12.5% and 51.5% with 4.5–24.5 larvae (51 maxim) per gall. The growth of the infested canes and fruit formation can be inhibited, and the canes can break beneath the galls [22,52,53]. In Russia, the leading country in the world raspberry production, it has been mentioned that L. rubi can inhibit the growth of plants and cause the stems to break due to wind because of the partial desiccation of the stems, reducing the production of the raspberry [55]. The European and Mediterranean Plant Protection Organization recognize the economic importance of L. rubi as a raspberry pest and requested monitoring by visual inspection and elimination from the fields in a 1993 Certification scheme for Rubus (PM 4/10 (1)) [56].
In Romania, L. rubi was recognized as a pest just after 1989 [57], with few mentions in the following years [58,59,60]; in 2020, it was presented across two pages in the “Guide for identification of the disease and pests of fruit shrubs”, elaborated by the National Phytosanitary Agency of Romania [61]. In this study, we found an average of 20% infestation of the canes and L. rubi was present in all the investigated sites, so we can consider it to be a widespread pest of the raspberry in Romania. In compact plantations, with more natural bushes growing, L. rubi prefer plants from the margins of the crops, which are areas with a small density of plants and so are more exposed to the attack of L. rubi. This is generally the situation in small farms, around private houses, in small gardens. But generally, in big raspberry plantations, plants are cultivated in rows, sometimes with vertical supports, especially horizontal wires, and here all the plants can be infested with L. rubi, without any special preferences.
The main observed damages produced by L. rubi are linked to the development of the larvae inside the stems of Rubus idaeus (Figure 2a,b). This results in apparition of visible galls (Figure 1c, Figure 4a–g and Figure 5a–f) inside the canes, sometimes in petioles (Figure 4h,i). The growth of the infested canes is inhibited and the fruit formation is affected, leading to yield loss [22,53]. It is also possible that the canes may break just below the galls (Figure 5a–d). Cracked galls can be located just a few cm above the soil (Figure 5a), affecting the entire plant, or even at 1.5–2 m high. In some cases, the gall can be located at the end of a lateral cane, stopping the growth of it (Figure 4g). Sometimes, the gall can result in total contortion of the cane (Figure 5e).
For Diptera galls associated with fungus, Krishnamurthy [62] considered that “only one fungus is associated with each insect gall”, but recent investigations on L. arundinis, L. carophila Löw and L. eryngii (Vallot) show that each species of galling Diptera, if we consider the larva, gall interior and gall surface, have a higher fungal richness and taxonomic variety, with complex mycobiomes. Just for the interior of the galls, the Cercospora beticola species complex for L. arundinis and Botryosphaeria dothidea for L. eryngii were identified, but for the larvae, Alternaria alternata species complex, Apenidiella spp., Aureobasidium pullulans, Mucor fragilis, Pseudopithomyces rosae, Botrytis cinerea, Filobasidium oeirense, Fusarium sporotrichioides, Papiliotrema frias, Cercospora zebrina, Tilletiopsis washingtonensis, Myrmecridium sp., Cladosporium sp., Periconia sp., etc., were identified. From the three gall compartments analyzed, the larval mycobiome composition looks to be more species-specific and can have an important functional role in the relation with the host [63]. Unfortunately, similar data are not yet available for L. rubi but may be very similar. Kaiser cultivates on Agar-Agar hyphae from L. rubi galls, and indeterminate species of Dothiorella have been obtained in 75% of cases [64].
Around 22 viruses are known to infect the raspberry [65], but none are known to be transmitted by L. rubi, so it is not clear whether it may be a potential raspberry virus vector for transmitting viruses or diseases to raspberry plants [65], but further research studies must investigate this.
L. rubi’s gall structure is not as well known as the gall structure of other Lasioptera galls such as L. arundinis Schiner, and generally not much is known on the fungus/gall midge association in Lasiopterini-induced galls [13,37,38,39]. Meyer [66] was the first who highlighted the role of the mycelium in the nourishment of the L. rubi larva, but other researchers were not so convinced of this. Kaiser [64] tried to cultivate the larvae of L. rubi on Agar-Agar but with no success, with the larvae dying in the absence of the mycelium. So, the statement of Krishnamurthy [62] that the “insect larva never survives without the fungus or vice versa” [62] can be true in this case. Kaiser [64] found glycogen inside the larvae and the only explanation offered for the presence of this polysaccharide is the fungal origin [64]. The larvae of the galling species from Lasiopterini have inherit ancestral mycetophagous capabilities, with the skills to feed in a saprophytic medium with fungal mycelium, and also consume plant tissues, in fact being a case of a phytomycetophagy. It is a situation of symbiosis where the fungus and the insect are mutually dependent for survival, with the insect larva being helped by the fungus to penetrate the stem, to lyse the middle lamellae of the cell, and to obtain adequate nutrition with a supplement from the mycelium. The fungus is helped by the insect in its transportation, inoculation, development and expansion inside the plant tissues [37,38,39,63,64,67]. Ambrosia galls are a particular case of structures resulting from the interaction between three categories of organisms: the plant (host), the insect (inducer) and the fungi (food source) [37]. The galls produced by L. rubi are classified as “cecidii caulinare laterale”, with larva situated inside the formative zone of the secondary xylem–phloem tissue [67], but the specific situations are more complicated, as demonstrated by the present research.
In all the gall morphotypes analyzed in this paper, the fundamental tissue in which larval chambers develop is predominantly composed of neo-formed tissue. The appearance of new meristematic centers was also reported in the case of galls induced by Cecidomyiidae in the superhost plant species Copaifera langsdorffii Desf. [68]. The presence of the larva induces dedifferentiation in the host’s tissues by reprogramming the pre-existing anatomical structures for the generation of new formations with meristematic characteristics. The formation of meristematic centers in galls is similar to that in callus generated in vitro cultures [69]; this can be explained by the existence of similar mechanisms for inducing cellular dedifferentiation mediated by phytohormones (such as IAA, identified in the case of the analyzed galls).
The structure and development of the secondary xylem is affected by the induction of gall development. The morphometric investigations carried out show a significant increase in the thickness of the secondary wood ring at the gall level (both near it and in the unaffected area), as well as in the diameter of the xylem vessels. But these increases are accompanied by a significant decrease in the density of xylem vessels, which reduces the efficiency of raw sap conduction in the area of the stem affected by the gall.
A decrease in the density of the xylem vessels in the gall compared to the normal stem was also observed in the case of galls induced by Dasineura sp. (Cecidomyiidae) on Peumus boldus Molina [18], but the differences were not significant. Instead, an increase in the size of xylem vessels is a constant in most organs affected by galls [17,70].
As xylem vessels are responsible for transporting water and dissolved nutrients from the roots to the rest of the plant, any disruption can prevent the flow, potentially leading to water stress or nutrient deficiencies in parts of the plant above the gall.
This reduced functionality of the xylem can lead to localized issues such as water stress in the tissues above the gall, especially during periods of high water demand (during hot weather in summer, when the fruits reach maturity and require an increased amount of water). In extreme cases, if the gall severely restricts xylem function (particularly in the case of GL3 galls, which encircle the stem), it could lead to wilting or even dieback of the plant tissue above the gall [71]. But in many cases, the flow of water through the modified xylem of the gall is ensured and thus the stem can continue to develop [69].
Histochemical investigations of galls help us to better understand the way in which the inducer interacts with the host plant, diverting its normal metabolism and meristematic activity, complete with the appearance of new structures, which favor its development [72].
Ambrosia galls induced by species of the tribes Asphondyliini and Lasiopterini, in general, lack typical nutritive tissue [37,73]; this consists of specialized parenchymatic cells, which are located on the edges of the larval chamber. However, there are exceptions; in galls from the leaves of Machilus spp. (Lauraceae), some authors described the presence of nutritive tissue lining the larval chamber [73]. Also, in galls induced by Cecidomyiidae at the leaflet level of the species Inga ingoides (Rich.) Willd. (Fabaceae: Caesalpinioideae), the presence of typical nutritive tissue around the larval chamber was reported [74].
In galls induced by L. rubi, typical nutritive tissue is missing, with the larval chamber being full of fungal mycelium, which comes into contact with the partially disintegrated cells of the host plant. The neo-formed tissue is actually a common storage tissue, as described by Bronner [75] and Costa et al. [76]. In it, both primary products of the metabolism (which provide the nutrients necessary for the inducer and, to a greater extent, the associated fungi) and secondary products are produced as a defense reaction to its aggression, but also with a possible protective role of the insects against natural predators. In the absence of a typical nutritive tissue, L. rubi larvae have a mixed diet (phytomycetophagy), using both the mycelium of the fungus and the nutrients provided by the host plant [63]. Primary metabolites (reducing sugars and proteins) as well as secondary metabolites (phenolic compounds, terpenoids) were identified in neo-formed tissue. The absence of lipids from all the morphotypes of galls investigated, as well as the reduced amounts of starch observed in them, can be noted. The nutrients stored in this tissue can be directed towards the gall inducer larva to contribute to its nutrition [18]. This occurs because larvae in fungal-associated galls consume plant and fungal cells, lacking classical nutritive tissue [37]. Significant fungal development, expected if larvae are fed only on mycelia, was not observed [77].
Changes in the normal structure of the stem, along with the induction of the formation of galls, appear first at the level of the cambium. Its normal activity is modified and the intensity of divisions increases, which leads to the formation of a much larger number of cells, especially from the secondary xylem (hyperplasia). However, simultaneously with these changes, change in the normal course of secondary xylem morphogenesis is also induced: lignification is inhibited and the number of wood vessels decreases at the expense of fibers (which are maintained with thin, cellulosic walls). This is how the unformed tissue appears, in which the divisions continue, along with an increase in the size of the component cells (hypertrophy). Later, meristematic nodules appear in the newly formed tissue, consisting of small cells with a dense cytoplasm. Conductive elements (xylem and phloem) are sometimes differentiated from these nodules, but they appear isolated in the neo-formed tissue, without apparent connections with the vascular system of the normal stem. Sap transport at the level of the gall is both apoplastic and symplastic [17], ensuring a moist environment which is conducive to the development of the larva and also of the fungus that serves as its main food. We thus distinguish three distinct types of divisions that participate in the formation and consolidation of neo-formed tissue: divisions that occur in parenchymal cells (typical for the phenomenon of dedifferentiation), divisions in meristematic nodules (also neo-formed), and divisions of secondary, pre-existing meristematic tissues—cambium and phelogen—whose activity is stimulated by larval induction.
The formation of larval chambers is accompanied by different changes in the adjacent tissues, depending on the area where they are located. The formation of the larval chambers in the neo-formed tissue is accompanied by minimal changes to it, as it is already intended to host the inducer larva. When the larval chamber comes into contact with the secondary xylem, breakages occur to create the space necessary for the larva’s development. This phenomenon is also observed in the case of galls induced by Dasineura sp. (Cecidomyiidae) in the stems of Peumus boldus [18]. Fungal hyphae contribute to the degradation of cells located near the larval chambers, thus obtaining a favorable substrate for their development [74]. The fungus degrades the cells in the vicinity of the larval chamber, helping the larva to feed on the host’s tissues directly or indirectly (since it also consumes hyphae from the young mycelium) [38].
IAA (indoleacetic acid) is an auxin that is normally found in plant organisms, where it is involved in growth and development processes [78]. At the cambium level, the activity of its cells is supported by increased amounts of IAA, while the differentiation of xylem elements, the thickening of cell walls, and lignification are induced by the presence of reduced amounts of IAA [79]. IAA is considered to have an important role in the phenomenon of gall induction [80], and insect larvae from Hymenoptera and Diptera have the ability to synthesize this compound [81]. Studies have shown that there is a gradient decrease in IAA concentration as we move away from the cambium, towards the secondary xylem elements in the process of development [82]. In the case of ambrosia galls induced by L. rubi, the increased amounts of IAA at the level of neo-formed tissue inhibit the maturation of xylem vessels and fibers. The same phenomenon is also observed around the larval chambers, where the large amount of IAA prevents lignification of adjacent tissues and maintains cell division for the production of new cells in the newly formed tissue.
Phenolic compounds are known to protect plants against herbivores and parasites, appearing as a response to both biotic and abiotic stress [83]. Research on their role in gall morphogenesis suggests that they are involved in hormonal regulation by inhibiting indole-3-acetic acid (IAA) oxidases, which indirectly leads to an accumulation of auxins [69,84]; the interaction of phenolics and phytohormones leads to increased and localized cell hypertrophy and hyperplasia.
Using pesticides against L. rubi is not the best solution, both for humans and environmental health. In raspberry fields in Poland, pesticide remains were detected in more than 50% of the investigated samples, and residues of 26 pesticides were found from 140 samples evaluated [85]. Generally, systemic pesticides from neonicotinoids are used against L. rubi, but in Russia the synthetic pyrethroid insecticide tau-fluvalinate is also used [71]. Neonicotinoid insecticides can have negative effects for non-target organisms, both for humans and other animals, including pollinating insects [86,87]. Using big quantities of pesticides leads to a weakening of the canes and a reduced value of the fruits [6]. Some environmentally friendly alternatives like essential oils used as green insecticides have not yet been investigated for controlling L. rubi [88,89].
The best environmentally friendly way to fight against L. rubi is visual control of the canes, cutting out the ones with galls and eliminating and destroying the damaged canes. Of course, this solution is applicable just for small farms and is not economically viable for big plantations. The best period for this preventive monitoring measure, by visual controls, is from late autumn until spring, before blooming when the leaves make this operation very difficult. When planting, the canes must also be visually investigated and stems without galls of L. rubi should be selected. Experience shows that the best time for planting the canes is in autumn [6]. If we plant canes with galls of L. rubi, it is possible that they may not produce any viable plants. From our experience of planting twenty canes with galls of L. rubi, none produced viable plants. In terms of agricultural methods, pruning must be carried out, taking care to removing all galled canes before the emergence of L. rubi adults [44]. Mechanical injuries to the canes can increase the attachment rate of L. rubi [22]. A solution to avoid reinfestation of the canes with L. rubi can be crop rotations, but the raspberry is a perennial plant with a life span of up to 15–20 years [22], so this could be difficult in practice. It is also important to have knowledge of the abundance of L. rubi in the surrounding environment in order to implement an effective pest management strategy. L. rubi attacks many species of wild Rubus, so the surrounding environment can be a natural reservoir for this pest [42].
5. Conclusions
Galls induced by L. rubi in the stems and leaf petiole of R. idaeus show great diversity both morphologically and structurally. The sizes of the galls vary widely, and the anatomical structure is different depending on the positioning of the larval chambers. The larval induction generates significant changes in the structure of the stem, affecting the activity of the secondary meristems whose cellular descendants are diverted from the normal morphogenetic program: a neo-formed tissue appears as a result of cambium hyperactivity, in which meristematic nodules are organized “de novo” from their activity and vascular elements. The structure of the secondary xylem is majorly affected, especially in the vicinity of the gall, through the generalized hyperplasia of the tissue, the inhibition of lignification and the stimulation of the formation of wide vessels, but with a low density. IAA plays an important role in gall morphogenesis, identified by histochemistry tests in the neo-formed tissue area and near the larval chambers. Its presence inhibits the normal development of wood, ensuring formation in the proximity of the larval inducer of an environment favorable to its development. Although in many situations vascular continuity is maintained in the case of galls appearing in the stem, sometimes (especially in GL3-type galls) ruptures occur at the level of the stem, leading to the death of its terminal part. From this perspective, L. rubi can be considered a pest that can reduce fruit production in R. idaeus by affecting the integrity of the attacked stems.
Author Contributions
Conceptualization, I.E.P. and I.N.G.; methodology, I.E.P. and I.N.G.; investigation, I.E.P. and I.N.G.; writing—original draft preparation, I.E.P. and I.N.G.; writing—review and editing, I.E.P. and I.N.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study as phytophagous Diptera are not considered to be laboratory animals according to EU Directive 2010/63.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Weber, H.E.; Rubus, L. Illustrierte Flora von Mitteleuropa. Band IV. Teil 2A. Spermatophyta: Angiospermae: Dicotyledones; Conert, H.J., Jäger, E.J., Kadereit, J.W., Schultze-Motel, W., Wagenitz, G., Weber, H.E., Eds.; Blackwell: Berlin, Germany, 1995. [Google Scholar]
- POWO—Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: https://powo.science.kew.org/ (accessed on 26 July 2024).
- WFO. World Flora Online. Version 2024.06. 2024. Available online: http://www.worldfloraonline.org (accessed on 29 July 2024).
- Lingdi, L.; Boufford, D.E.R.; Editorial Committee of Flora of China. Flora of China; Wu, Z., Raven, P.H., Eds.; Chinese Academy of Sciences, Science Press: Beijing, China, 2003. [Google Scholar]
- Beldie, A.; Buia, A.; Guşuleac, M.; Nyárády, E.I.; Prodan, I.; Răvăruţ, M. Flora Republicii Populare Române; Editura Academiei Republicii Populare Române: Bucharest, Romania, 1956; Volume 6. [Google Scholar]
- Totic, I. Raspberry Breeding and Protection against Disease and Pests. Bulg. J. Agric. Sci. 2014, 20, 391–404. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2024-1. 2024. Available online: www.iucnredlist.org (accessed on 20 July 2024).
- Khela, S. Rubus idaeus (Europe Assessment). The IUCN Red List of Threatened Species 2012: e.T203456A2765767. Rubus idaeus (Raspberry). Available online: https://www.iucnredlist.org/ (accessed on 28 July 2024).
- Aronsson, M.; Bioret, F.; Bita-Nicolae, C.; Capelo, J.; Čarni, A.; Dimopoulos, P.; Janssen, J.; Loidi, J. F3.1b Temperate Rubus Scrub; European Red List of Habitats—Heathland Habitat Group: Gland, Switzerland, 2016. [Google Scholar]
- Tobyn, G.; Denham, A.; Whitelegg, M. Rubus idaeus, Raspberry. In Medical Herbs; Elsevier: Amsterdam, The Netherlands, 2011; pp. 271–282. ISBN 978-0-443-10344-5. [Google Scholar]
- Rao, A.V.; Snyder, D.M. Raspberries and Human Health: A Review. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef]
- Bojkovska, K.; Jankulovski, N.; Mihajlovski, G.; Momirceski, J. Analysis of Market Opportunities for Raspberry Production in The Republic of North Macedonia. Int. J. Res. GRANTHAALAYAH 2020, 8, 149–154. [Google Scholar] [CrossRef]
- Redfern, M. Plant Galls; Harper Collins Publishers: New York City, NY, USA, 2011. [Google Scholar]
- Gagné, R.J.; Jaschhof, M. A Catalog of the Cecidomyiidae (Diptera) of the World, 5th ed.; Systematic Entomology Laboratory, Agricultural Research Service, U.S. Department of Agriculture, U.S. National Museum: Washington, DC, USA, 2021. [Google Scholar]
- Skuhravá, M.; Skuhravý, V. Species Richness of Gall Midges (Diptera: Cecidomyiidae) in Europe (West Palaearctic): Biogeography and Coevolution with Host Plants. Acta Soc. Zool. Bohem. 2009, 73, 87–156. [Google Scholar]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; deWaard, J.R. Counting Animal Species with DNA Barcodes: Canadian Insects. Philos. Trans. R. Soc. B 2016, 371, 20150333. [Google Scholar] [CrossRef] [PubMed]
- Bragança, G.P.P.; Freitas, M.D.S.C.; Isaias, R.M.D.S. The Influence of Gall Position over Xylem Features in Leaflets of Inga ingoides (Rich.) Willd. (Fabaceae: Caesalpinioideae). Trees 2021, 35, 199–209. [Google Scholar] [CrossRef]
- Guedes, L.M.; Aguilera, N.; Gavilán, E.; Péndola, J.A.; Villagrán, N.E. Vascular Implications of Dasineura sp. Galls’ Establishment on Peumus boldus Stems. Plant Biol. J. 2023, 25, 965–972. [Google Scholar] [CrossRef]
- Carneiro, R.G.; Burckhardt, D.; Isaias, R.M. Biology and systematics of gall-inducing triozids (Hemiptera: Psylloidea) associated with Psidium spp.(Myrtaceae). Zootaxa 2013, 3620, 129–146. [Google Scholar] [CrossRef]
- Arduin, M.; Kraus, J.E. Anatomia de Galhas de Ambrosia Em Folhas de Baccharis concinna e Baccharis dracunculifolia (Asteraceae). Braz. J. Bot. 2001, 24, 63–72. [Google Scholar] [CrossRef]
- Neger, F.W. Ambrosiapilze. Berichte Dtsch. Bot. Ges. 1908, 26A, 735–754. [Google Scholar]
- Tanasković, S.; Milenković, S.; Sretenović, D. Intensity of Attack of Raspberry Gall Midge Lasioptera rubi Schrank (Diptera, Cecidomyiidae) on Some Raspberry Genotypes. Acta Entomol. Serbica 2008, 13, 43–50. [Google Scholar]
- Möhn, E. Beiträge Zur Systematik Der Larven Itonididae (Cecidomyiidae, Diptera). 1. Teil: Porricondylinae Und Itonidinae Mitteleuropas. Lieferung 1. In Zoologica Original-Abhandlungen aus dem Gesamtgebiete der Zoologie; Forgotten Books: London, UK, 1955; Volume 105, pp. 1–247. [Google Scholar]
- Yukawa, J. A Revision of the Japanese Gall Midges (Diptera: Cecidomyiidae); Memoirs of the Faculty of Agriculture Kagoshima University: Kagoshima, Japan, 1971; Volume 8, pp. 1–203. [Google Scholar]
- Thomas, D.B.; Goolsby, J.A. Morphology of the Preimaginal Stages of Lasioptera donacis Coutin (Diptera: Cecidomyiidae), a Candidate Biocontrol Agent for Giant Arundo Cane. Psyche J. Entomol. 2015, 2015, 262678. [Google Scholar] [CrossRef]
- Jensen, W.A. Botanical Histochemistry: Principles and Practice; W. H. Freeman and Co: San Francisco, CA, USA, 1962. [Google Scholar]
- Şerbănescu-Jitariu, G.; Andrei, M.; Mitroiu-Rădulescu, N.; Petria, E. Practicum de Biologie Vegetală; CERES: Bucureşti, Romania, 1983. [Google Scholar]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Srebotnik, E.; Messner, K. A Simple Method That Uses Differential Staining and Light Microscopy to Assess the Selectivity of Wood Delignification by White Rot Fungi. Appl. Environ. Microbiol. 1994, 60, 1383–1386. [Google Scholar] [CrossRef] [PubMed]
- Johansen, D.A. Plant Microtechnique; McGrawHill: New York, NY, USA, 1940. [Google Scholar]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan red 7B or fluoral yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Sass, J.E. Botanical Microtechnique, 2nd ed.; Iowa State College Press: Ames, IA, USA, 1951. [Google Scholar]
- David, R.; Carde, J.P. Histochimie-Coloration Differentielle Des Inclusions Lipidiques et Terpeniques des Pseudophylles du Pin Maritime au Moyen du Reactif NADI. Comptes Rendus Hebd. Seances Acad. Sci. 1964, 258, 1338. [Google Scholar]
- Gahan, P.B. Plant Histochemistry and Cytochemistry: An Introduction; Academic Press: Orlando, FL, USA, 1984. [Google Scholar]
- Bedetti, C.S.; Modolo, L.V.; Isaias, R.M.D.S. The Role of Phenolics in the Control of Auxin in Galls of Piptadenia gonoacantha (Mart.) MacBr (Fabaceae: Mimosoideae). Biochem. Syst. Ecol. 2014, 55, 53–59. [Google Scholar] [CrossRef]
- Tastás-Duque, R.; Sylvén, E. Sensilla and Cuticular Appendages on the Female Abdomen of Lasioptera rubi (Schrank) (Diptera, Cecidomyiidae). Acta Zool. 1989, 70, 163–174. [Google Scholar] [CrossRef]
- Rohfritsch, O. Plants, Gall Midges, and Fungi: A Three-component System. Entomol. Exp. Appl. 2008, 128, 208–216. [Google Scholar] [CrossRef]
- Rohfritsch, O. A fungus associated gall midge, Lasioptera arundinis (Schiner), on Phragmites australis (Cav.) Trin. Bull. Soc. Bot. Fr. Lett. Bot. 1992, 139, 45–59. [Google Scholar] [CrossRef]
- Rohfritsch, O. Morphological and Behavioural Adaptations of the Gall Midge Lasioptera arundinis (Schiner) (Diptera, Cecidomyiidae) to Collect and Transport Conidia of Its Fungal Symbiont. Tijdschr. Voor Entomol. 1997, 140, 59–66. [Google Scholar]
- Barnes, H.F. On Some Factors Governing the Emergence of Gall Midges (Cecidomyidæ: Diptera). Proc. Zool. Soc. Lond. 1930, 100, 381–393. [Google Scholar] [CrossRef]
- Gordon, S.C.; Woodford, J.A.T.; Birch, A.N.E. Arthropod Pests of Rubus in Europe: Pest Status, Current and Future Control Strategies. J. Hortic. Sci. 1997, 72, 831–862. [Google Scholar] [CrossRef]
- Barnes, H.F. The Gall Midges of Blackberries and Raspberries. J. Pomol. Hortic. Sci. 1926, 5, 137–140. [Google Scholar] [CrossRef]
- Barnes, H.F. Gall Midges of Economic Importance, Volume 3, Fruit; Crosby Lockwood & Son Ltd.: London, UK, 1948. [Google Scholar]
- Viggiani, G.; Mazzone, P. Infestazioni di Lasioptera rubi (Schrank) (Diptera, Cecidomyiidae) in Lamponeti Dell’ltalia Meridionale. Inf. Fitopatol. 1978, 2, 3–4. [Google Scholar]
- Darvas, B.; Skuhravá, M.; Andersen, A. Agricultural Dipteran Pests of the Palaearctic Region. In Contributions to a Manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; General and Applied Dipterology; Science Herald: Budapest, Romania, 2000; Volume 1, pp. 565–650. [Google Scholar]
- Skuhravá, M.; Skuhravý, V.; Meyer, H. Gall Midges (Diptera: Cecidomyiidae: Cecidomyiinae) of Germany; Druckzentrum, Neumünster: Kiel, Germany, 2014. [Google Scholar]
- Simova, D.; Dobrivojević, K. Raspberry Gall Midge (Lasioptera rubi Heeg.) A Little Known Pests of Raspberry. Plant Prot. 1966, 91, 323–330. [Google Scholar]
- Dobrivojević, K. Ekonomski Značajne Štetočine Maline u Proizvodnim Područjima Valjeva i Čačka. Zaštita Bilja 1968, 100–101, 253–271. [Google Scholar]
- Simova-Tošić, D A Little Known Pests of Raspberry from Family Cecidomyiidae (Diptera). J. Yugosl. Pomol. 1970, 11–12, 193–197.
- Milenković, S.; Tanasković, S. Harmfulness of Raspberry Gall Midge, Lasioptera rubi Schrank (Diptera, Cecidomyiidae), to Some Raspberry Cultivars. IOBC/WPRS Bull. 2008, 39, 71–75. [Google Scholar]
- Gordon, S.C.; Milenković, S.; Cross, J.V.; Stanisavljevic, M. Raspberry Pests in Serbia. Bull. OILB/SROP 2003, 26, 23–27. [Google Scholar]
- Tanasković, S.; Milenković, S. Occurrence of Raspberry Gall Midge Lasioptera rubi Schrank (Diptera, Cecidomyiidae) in Some Raspberry Cultivars. Acta Agric. Serbica 2009, 14, 79–85. [Google Scholar]
- Tanasković, S.T.; Milenković, S.N. Open Field Surveys to Evaluate the Susceptibility of Red Raspberry Genotypes to Raspberry Gall Midge, Lasioptera rubi Schrank (Diptera, Cecidomyiidae)—4 Year Results. Acta Hortic. 2012, 946, 247–251. [Google Scholar] [CrossRef]
- Milenković, S.; Ranković, M. Integral Raspberry Protection. Proc. Winter Sch. Agron. Fac. Agron. 2001, 5, 59–65. [Google Scholar]
- Yegorenkova, E.; Yefremova, Z. Notes on Lasioptera rubi (Schrank) (Diptera: Cecidomyiidae) and Its Larval Parasitoids (Hymenoptera) on Raspberries in Russia. Entomol. Fenn. 2019, 27, 15–22. [Google Scholar] [CrossRef][Green Version]
- OEPP/EPPO Recommendations made by EPPO Council in 1992: Scheme for the production of classified vegetatively propagated ornamental plants to satisfy health standards. Bull. OEPP/EPPO Bull. 1993, 23, 735–736.
- Teodorescu, G. Contribuții La Studiul Biologiei și Ecologiei Musculiței Galicole a Lăstarilor de Zmeur și Mur, Lasioptera rubi Heeger. Bul. Prot. Plantelor 1989, 2, 55–57. [Google Scholar]
- Popescu, I.E. Torymid Wasps (Hymenoptera, Chalcidoidea, Torymidae) New for Romanian Fauna. Analele Ştiinţifice Univ. “Al. I. Cuza” Iaşi Ser. Biol. Anim. 2003, 49, 83–86. [Google Scholar]
- Neacşu, P. Galls from Romania; Victor Bortaş Publishing House: Bucureşti, Romania, 2006. [Google Scholar]
- Chiriceanu, C.; Chiriloaie, A.; Teodoru, A.; Sivu, C. Contribution to Knowledge of the Gall Insects and Mites Associated with Plants in Southern Romania. Sci. Pap. Ser. B Hortic. 2015, 59, 27–36. [Google Scholar]
- National Phytosanitary Agency Guide for Identification of the Disease and Pests of Fruit Shrubs. Bucharest 2020. Available online: https://www.anfdf.ro/sanatate/ghid/ghid_arbusti.pdf (accessed on 10 August 2024).
- Krishnamurthy, K.V. Plant-Insect-Fungus Association in Some Plant Galls. Proc. Anim. Sci. 1984, 93, 265–273. [Google Scholar] [CrossRef]
- Pyszko, P.; Šigutová, H.; Kolařík, M.; Kostovčík, M.; Ševčík, J.; Šigut, M.; Višňovská, D.; Drozd, P. Mycobiomes of Two Distinct Clades of Ambrosia Gall Midges (Diptera: Cecidomyiidae) Are Species-Specific in Larvae but Similar in Nutritive Mycelia. Microbiol. Spectr. 2024, 12, e02830-23. [Google Scholar] [CrossRef]
- Kaiser, P. Importance of Fungal Hyphae in Mycotic Galls. Studies on Lasioptera rubi (Schrank, 1803) (Diptera, Cecidomyiidae). Entomol. Mitteilungen Zool. Mus. Hambg. 1978, 6, 41–48. [Google Scholar]
- Tan, J.L.; Trandem, N.; Fránová, J.; Hamborg, Z.; Blystad, D.-R.; Zemek, R. Known and Potential Invertebrate Vectors of Raspberry Viruses. Viruses 2022, 14, 571. [Google Scholar] [CrossRef]
- Meyer, J. Cécidogenèse de La Galla de Lasioptera rubi Heeger et Rôle Nourricier d’un Mycélium Symbiotique. Comptes Rendus Hebd. Seances Acad. Des Sci. 1952, 234, 2256–2558. [Google Scholar]
- Mani, M.S. Ecology of Plant Galls; Springer-Science+Business Media, B.V.: Dordrecht, The Netherlands, 1964. [Google Scholar]
- Carneiro, R.G.S.; Isaias, R.M.S.; Moreira, A.S.F.P.; Oliveira, D.C. Reacquisition of New Meristematic Sites Determines the Development of a New Organ, the Cecidomyiidae Gall on Copaifera langsdorffii Desf. (Fabaceae). Front. Plant Sci. 2017, 8, 1622. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Isaias, R.M.; De Oliveira, D.C.; Da Silva Carneiro, R.G.; Kraus, J.E. Developmental Anatomy of Galls in the Neotropics: Arthropods Stimuli Versus Host Plant Constraints. In Neotropical Insect Galls; Fernandes, G.W., Santos, J.C., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 15–34. ISBN 978-94-017-8782-6. [Google Scholar]
- Jorge, N.D.C.; Freitas, M.D.S.C.; Caffaro, R.M.; Vale, F.H.A.; Lemos-Filho, J.P.; Isaias, R.M.D.S. Vascular Traits of Stem Galls: Cell Increment versus Morphogenetic Constraints in Wood Anatomy. Plant Biol. J. 2022, 24, 450–457. [Google Scholar] [CrossRef]
- Marini-Filho, O.J.; Fernandes, G.W. Stem Galls Drain Nutrients and Decrease Shoot Performance in Diplusodon orbicularis (Lythraceae). Arthropod-Plant Interact. 2012, 6, 121–128. [Google Scholar] [CrossRef]
- Coelho Kuster, V.; Costa Rezende, U.; Fernandes Cardoso, J.C.; Dos Santos Isaias, R.M.; Coelho De Oliveira, D. How Galling Organisms Manipulate the Secondary Metabolites in the Host Plant Tissues? A Histochemical Overview in Neotropical Gall Systems. In Co-Evolution of Secondary Metabolites; Mérillon, J.-M., Ramawat, K.G., Eds.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Switzerland, 2020; pp. 823–842. ISBN 978-3-319-96396-9. [Google Scholar]
- Chao, J.-F.; Liao, G.-I. Histocytological Aspects of Four Types of Ambrosia Galls on Machilus zuihoensis Hayata (Lauraceae). Flora—Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 157–164. [Google Scholar] [CrossRef]
- Bragança, G.P.P.; Ferreira, B.G.; Isaias, R.M.D.S. Distinct Cytological Mechanisms for Food Availability in Three Inga ingoides (Fabaceae)—Cecidomyiidae Gall Systems. Protoplasma 2022, 259, 155–162. [Google Scholar] [CrossRef]
- Bronner, R. The role of nutritive cells in the nutrition of cynipids and cecidomyiids. In Biology of Insect-Induced Galls; Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 118–140. [Google Scholar]
- Costa, E.C.; Oliveira, D.C.; Ferreira, D.K.L.; Isaias, R.M.S. Structural and Nutritional Peculiarities Related to Lifespan Differences on Four Lopesia Induced Bivalve-Shaped Galls on the Single Super-Host Mimosa gemmulata. Front. Plant Sci. 2021, 12, 660557. [Google Scholar] [CrossRef]
- Sá, C.E.M.D.; Silveira, F.A.O.; Santos, J.C.; Isaias, R.M.D.S.; Fernandes, G.W. Anatomical and Developmental Aspects of Leaf Galls Induced by Schizomyia macrocapillata Maia (Diptera: Cecidomyiidae) on Bauhinia brevipes Vogel (Fabaceae). Braz. J. Bot. 2009, 32, 319–327. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin Biosynthesis and Its Role in Plant Development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Sorce, C.; Giovannelli, A.; Sebastiani, L.; Anfodillo, T. Hormonal Signals Involved in the Regulation of Cambial Activity, Xylogenesis and Vessel Patterning in Trees. Plant Cell Rep. 2013, 32, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Gatjens-Boniche, O. The Mechanism of Plant Gall Induction by Insects: Revealing Clues, Facts, and Consequences in a Cross-Kingdom Complex Interaction. Rev. Biol. Trop. 2019, 67, 1359–1382. [Google Scholar] [CrossRef]
- Bailey, S.; Percy, D.M.; Hefer, C.A.; Cronk, Q.C.B. The Transcriptional Landscape of Insect Galls: Psyllid (Hemiptera) Gall Formation in Hawaiian Metrosideros polymorpha (Myrtaceae). BMC Genom. 2015, 16, 943. [Google Scholar] [CrossRef] [PubMed]
- Bhalerao, R.P.; Bennett, M.J. The Case for Morphogens in Plants. Nat. Cell Biol. 2003, 5, 939–943. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The Roles of Plant Phenolics in Defence and Communication during Agrobacterium and Rhizobium Infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef]
- Hori, K. Insect secretions and their effect on plant growth, with special references to Hemipteran. In Biology of Insect-Induced Galls; Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 157–170. [Google Scholar]
- Łozowicka, B.; Kaczyński, P.; Jankowska, M.; Rutkowska, E.; Hrynko, I. Pesticide Residues in Raspberries (Rubus idaeus L.) and Dietary Risk Assessment. Food Addit. Contam. Part B 2012, 5, 165–171. [Google Scholar] [CrossRef]
- Thompson, D.A.; Lehmler, H.-J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; LeFevre, G.H.; Peeples, T.L.; Poch, M.C.; LaDuca, L.E.; et al. A Critical Review on the Potential Impacts of Neonicotinoid Insecticide Use: Current Knowledge of Environmental Fate, Toxicity, and Implications for Human Health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef] [PubMed]
- Alsafran, M.; Rizwan, M.; Usman, K.; Saleem, M.H.; Jabri, H.A. Neonicotinoid Insecticides in the Environment: A Critical Review of Their Distribution, Transport, Fate, and Toxic Effects. J. Environ. Chem. Eng. 2022, 10, 108485. [Google Scholar] [CrossRef]
- Gostin, I.N.; Popescu, I.E. Evaluation of the Essential Oils Used in the Production of Biopesticides: Assessing Their Toxicity toward Both Arthropod Target Species and Beneficial Pollinators. Agriculture 2023, 14, 81. [Google Scholar] [CrossRef]
- Popescu, I.E.; Gostin, I.N.; Blidar, C.F. An Overview of the Mechanisms of Action and Administration Technologies of the Essential Oils Used as Green Insecticides. AgriEngineering 2024, 6, 1195–1217. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).