Abstract
The optimization of artificial pollination is crucial in breeding programs for allogamous plants. In maize, achieving a balance between the labor-intensive nature of controlled pollinations and the need for large-scale production of hybrid seeds, along with considerations of germinability and pollen production, determines the success of genetic improvement programs. Breeding programs in maize have resulted in a reduction in the number of tassel branches to increase light interception and plant density in production fields. However, despite this genetic improvement, the decreased pollen production per plant has raised critical concerns regarding pollination and subsequent ear filling, especially under adverse environmental conditions. The aim of this work was the analysis of factors that can contribute to increasing the efficiency of controlled pollination in maize. The data obtained showed that pollen diameter, flavonoid, and phenolic acid content do not influence the percentage of germination and therefore the efficiency of pollination. The quantity of pollen is a central factor in ensuring the efficiency of controlled pollinations, and the data obtained by comparing traditional varieties with modern hybrids of maize showed that an increase in pollen production is determined by the increase in branching of the male inflorescence. Furthermore, we propose the use of a “smart” pollination method to make this step in maize breeding programs easier and faster.
1. Introduction
Maize is a cross-pollinated species with distinct female and male flowers present on the same plant [1,2]. In maize populations, pollination generally occurs with pollen coming from different plants, belonging to the same or different populations, carried by gravity, wind, and insects [3,4].
Starting from the pioneering studies of Shull and East regarding the constitution of inbred lines and hybrids [5,6,7,8], pollen management has been investigated as a method for researchers to limit cross-contaminations in breeding programs and seed multiplication [9,10,11]. The artificial pollination method, based on the use of paper bags to isolate tassel and ear inflorescences and avoid pollen contamination, is commonly used in breeding programs [12]. Two important traits to consider for a good seed set are the number of lateral branches in the tassel, a key factor in pollen production, and the pollen germination rate, defined as the number of viable grains out of the total number of pollen grains. Furthermore, in addition to genetic factors dependent on the cultivated variety, several environmental factors can affect pollen viability.
Flowering time is a critical phase influencing yield. In maize, its duration varies from 5 to 14 days [13,14], starting with pollen emission from the tassel. After an interval of a few days, depending on the genetic background, silks protrude from the husks. Each plant can produce about 25 million pollen grains; considering that one ear has from 700 to 1000 silks, up to 25,000 pollen grains are available per silk [15].
Pollen production and germinability can be affected by biotic and abiotic stresses. The critical phase lasts from stamen initiation to anthesis, and even short periods of stress can have severe effects on fertility [16]. Water stress can affect pollen production and germinability, while UV-B rays and the time elapsed between pollen dispersion from anthers and its arrival on silks decrease its germinability [17,18,19,20].
Heat stress (35 °C) during the bicellular stage of pollen development has been shown to misregulate cell cycle regulatory genes, disrupting the metaphase-to-anaphase transition. This likely impairs the development and transport of sperm cells within the pollen tube, leading to sterility [21]. Impaired early pollen tube growth has been identified as a key factor in heat stress-induced (38 °C) maize sterility, with reduced sugar levels, increased ROS content, and hormonal imbalances in the silks, leading to misdirected pollen tube growth [22].
Flavonoids play an important role in pollen protection from UV rays, and mutations in genes involved in the flavonoid biosynthetic pathway can affect pollen production [21,22,23,24].
In breeding programs aimed at increasing maize yield, the focus has been the selection of smaller tassels to reduce the shading on upper leaves, enhance photosynthetic capacity, and increase the planting density. However, this resulted in reduced pollen production [25]. For this reason, studies on factors affecting pollen production and germinability are crucial for both breeding programs and hybrid production.
Tassel size (i.e., the number and length of tassel branches, number of spikelets, and tassel area index), has been reported to be negatively correlated with yield but positively correlated with pollen production [26,27,28,29,30]. In maize, there are three Ramosa genes (Ra1, Ra2, and Ra3) whose function is related to the regulation of tassel branch number. Literature data have shown that mutations in these three genes increase branching by changing the identity of the spikelet-pair meristems to branch meristems [31,32].
The Ra1 and Ra2 genes encode putative transcription factors while Ra3 encodes a trehalose-6-phosphate phosphatase [31]. Characterization of the loss of function mutant Ra1-154, with a deletion in the putative zinc-finger motif, suggested a role as a DNA-binding protein for this gene [32].
In breeding programs, in order to obtain the desired progeny and maintain the purity of the breeding stock, it is fundamental to avoid unwanted pollination. Cross-contamination has been reported in open-pollinated varieties, resulting in out-of-type individuals in the progeny. Examples of cross-contamination have been reported involving both seed (shape, volume, weight, color, oil and protein content, and fatty acid composition) and plant traits (plant height, tassel shape, and the presence of secondary metabolites as anthocyanins) [33,34,35,36,37].
The problem of cross-contamination can be bypassed by controlled pollination or isolating plants via spatial (at least 200–300 mt between different maize accessions) or temporal distance (e.g., staggered sowing). In breeding programs, pollen contamination is generally avoided using controlled pollinations, as isolating the different accessions would be more time-consuming and less efficient.
Various artificial pollination systems are used as a replacement or in addition to natural systems to increase the efficiency of natural pollination, i.e., due to the action of the wind or insects. These systems (reviewed by Brossard et al., 2023) [38] involve the first phase of collection and the second phase of the distribution of the pollen and are based on the use of relatively simply handled tools such as vibratory wands and blowers, more complex tools mounted on tractors such as pollen blowers or high-pressure/fogger/electrostatic sprayers, or tools based on more advanced technology such as drones and robotic pollinators [38]. In the case of maize, for the production of hybrid seeds, harvesting machinery has been developed to collect the plumes and extract the pollen, while for pollination, robotic sprayers carried by tractors are available, particularly suitable for commercial-scale crops [38].
The aim of this study was to assess the in vitro and in vivo viability of maize pollen grains from different genetic materials and to study possible correlations of this parameter with traits such as the pollen diameter and the content of different antioxidant compounds inside the pollen grain. Furthermore, we presented a simple and fast method for controlled pollination, of which the accuracy will be compared with the classic method by monitoring the percentage of out-of-type individuals in the progeny using phenotypic markers for seed and plant traits.
2. Materials and Methods
2.1. Plant Materials
The maize genotypes analyzed in this study were cultivated in the experimental field located at Landriano (PV), Italy, 45°55′50.66″ N, 10°13′48.98″ E, during the agronomic season in 2021. For each genotype studied, the seeds were sown in 80 m rows with a plot seeder, with 70 cm inter-row spacing and 20 cm intra-row spacing.
To measure the pollen traits such as the percentage of germinability, diameter, and flavonoid and phenolic acid content, we chose 11 commercial inbred lines, commonly cultivated in Italy, indicated by the codes from R1 to R11.
The following genotypes were used to analyze the tassel morphology: two inbred lines and their F1/F2 progenies, respectively, R6708 (B73), R6706 (Mo17), R6603 (F1 B73/Mo17), and R6732 (F2 B73/Mo17); three commercial hybrids, R6600 (Hybrid PI817 Pioneer FAO 700), R6601 (Hybrid LG 32.85 Limagrain FAO 200), and R6602 (Hybrid PR33A46 Pioneer FAO 500); and five landraces, R6571 (Nero Spinoso), R6573 (Rostrato Rosso di Rovetta), R6637 (Spinato di Gandino), R6639 (Mais delle Fiorine), and R6640 (Millo Corvo) [39,40,41].
Regarding the cross-contamination study, we used the recessive sugary1 mutation, the dominant Purple plant1 allele [42], and the recessive brachytic2 mutation [43]. The Pl1 plants, used as the pollen donor, were sown in a single row with 5 rows on the left and 5 rows on the right of pl1 plants, used as the female parent. The su1 plants were sown in a single row with 5 rows on the left and 5 rows on the right of Su1 plants. The br2 plants were sown in a single row with 5 rows on the left and 5 rows on the right of the Br2 plants. The out-of-type plants were analyzed in the following agronomic season (2022).
To study pollen longevity, we used the F1 hybrid B73 × Mo17 (ID R6116) and the inbred line B73 (ID R6123).
2.2. In Vitro Pollen Germination and Measurement of Pollen Diameter
Tassels were harvested at 50% blooming at about 10 AM in the morning (Supplementary Figure S1).
After harvesting, tassels were placed in a beaker with water and left for one hour at room temperature. The pollen was collected by shaking the tassels on a sheet of paper, and then it was distributed uniformly on Petri dishes containing 6 mL of a semi-solid culture medium for pollen germination (60 g of sucrose, 0.15 g of CaCl2, 0.05 g of H3BO3, 5 g of agar, 375 mL of distilled water, and a pH of 5.8).
The Petri dishes were incubated for one hour at room temperature to allow pollen germination. Toluidine blue 0.05% (v/w) was added to arrest the development of the pollen tube and visualize pollen grains. Pictures were taken using a Leica stereoscope equipped with an Olympus Camedia C-7070 wide-zoom digital camera.
For each genotype, at least 200 pollen grains were analyzed.
Pollen germinability was calculated as the percentage of germinated pollen grains out of the total number of pollen grains analyzed.
Pollen diameter was measured using the ImageJ program, version 1.51, on the pictures previously used to analyze pollen germinability.
2.3. Determination of Flavonoids and Phenolic Acids Content
To quantify flavonoid and phenolic acid content, 0.002 g of pollen was collected in a 1.5 mL tube and 250 µL of extraction buffer (EtOH 95%-HCl 1%) was added. After 3 min of centrifugation at 13,000 rpm, the pollen was ground for 1 min and centrifuged again for 2 min at 13,000 rpm. Then, 750 µL of extraction buffer was added to the tube, and the samples were centrifuged for 10 min at 13,000 rpm. Next, 500 µL of the supernatant was transferred to a new tube with 500 µL of extraction buffer. The absorbance was measured using a quartz cuvette at three different wavelengths: 530 nm for anthocyanins, 350 nm for flavonoids, and 280 nm for phenolic acids. Then, 1 mL of the extraction buffer was used as a blank. The extinction coefficient (ε) was 21,877 L m−1 mol−1 for flavonoids and 14,700 L m−1 mol−1 for phenolic acids.
2.4. Analysis of Tassel Morphology
For each of the varieties used to analyze the tassel morphology, listed in Section 2.1, we measured the weight and total length of 5 tassels and counted the total number of branches and spikelets for each tassel.
2.5. “Smart” Pollination
In canonical self-pollination, the ears were isolated with a paper bag before the silk emergence to prevent pollen contamination. The day before pollination, the tassels at anthesis were isolated with a paper bag, and the tip of ears that reached maturity were cut back to facilitate the development of a short, thick brush of silks ensuring a good seed set. The ears were immediately re-covered with the bag.
The next day, the tassel bag was gently shaken to collect the pollen, which was poured onto the silks. The pollinated ear was then covered with the tassel bag, firmly staked on the stalk of the plant. The following information was written on the tassel bag: the date of pollination, the code number of the genotype(s), and a symbol identifying the type of pollination (self-pollination or cross).
In “smart” pollination, the main difference was that the tassel was not isolated with a paper bag the day before the pollination, making the preparation of plants for controlled pollination faster and less laborious. Immediately before pollination, the pollen was collected in a paper bag and poured onto the silks (Figure 1D,E).

Figure 1.
Canonical (A–C) and “smart” (D,E) pollinations. (A) The day before pollination, the tassel was isolated using a paper bag; (B) collection of pollen; (C) pollination; (D) collection of pollen from a tassel not previously isolated by a bag; (E) pollination.
To test the accuracy of “smart” pollination, we compared the percentage of off-type progeny produced in “smart” and canonical controlled pollination. To easily recognize the out-of-type progeny, we performed pollinations using plants carrying the mutations sugary1 (su1), resulting in wrinkled and translucent seeds when dry (ID R3475, R5795, R5796, and R 5797), and brachytic2 (br2), resulting in reduced-height and dark green, erect leaves (ID R3535). Both homozygous mutants, su1 and br2, were self-pollinated, and the progeny were scored for out-of-type phenotypes (wild-type seeds and wild-type seedling/plant, respectively). The total number of sugary1 and wild-type seeds were counted with the help of a seed counter (CGOLDENWALL Semi-Automatic Counter Machine Counter SLY-C Hangzhou, China).
The inbred line Pl1-Pl1 (Pl: Purple Plant) (code number R 3371), with red anthers, was used as the pollen donor in crosses with the B73 inbred line, homozygous pl1/pl1, with yellow anthers. In this cross, the out-of-type progeny was easily identified by the presence of yellow anthers.
2.6. Evaluation of Pollen Longevity in the Field
To test pollen longevity in the field, we performed canonical pollination using the F1 hybrid B73 × Mo17 as the female parent and the B73 inbred line as the pollen donor. The pollen was collected in paper bags from tassels in anthesis, at 10:00 a.m., stored in the field in the shade (for T0 to T3 pollinations), and used for pollinations at successive times. Pollen used 24 h or more after collection was stored in a dark room at room temperature (RT). The first pollinations (T0) were made immediately after pollen collection at 10:00 AM, followed by pollinations after 1 h (T1), 2 h (T2), 3 h (T3), 24 h (T4), 2 days (T5), 3 days (T6), 5 days (T7), 6 days (T8), and 7 days (T9) of storage. After the harvest, the ears were analyzed and classified according to the seed set into four different categories ranging from 0 to 100% of seeds. The range of 0–25% corresponds to category 1; 25–50% to category 2; 50–75% to category 3; and 75–100% to category 4 (Supplementary Figure S2).
2.7. Informatic Tools
Microsoft Excel®, Microsoft Corporation, Redmond, WA, USA, was used to collect data. The PAST program (Paleontological Statistics, version 4.12) was used to perform statistical analysis. All statistical analyses performed are described in the table and figure captions. Generally, the analyses were conducted using ANOVA and post-hoc tests.
Results are presented as mean ± standard deviation. Statically significant differences are considered for p ≤ 0.05.
3. Results
3.1. Analysis of Pollen Germinability and Diameter, Flavonoid and Phenolic Acid Content
Different parameters on pollen (pollen germinability, pollen diameter, and flavonoid and phenolic acid content) were analyzed in 11 commercial inbred lines commonly cultivated in Italy (Table 1).

Table 1.
Parameters collected on pollen of 11 different inbred lines. For each parameter measured, mean and SD are reported. Different letters indicate statistically significant differences (Tukey’s test, p < 0.05). n.d.: not determined. N = 200.
The values of pollen germinability showed a gradient from the lowest value 36.54% (R10) to the highest value 79.82% (R6) (Table 1).
Regarding the pollen diameter, the lowest value was observed in line R5 (61 μm) while the highest was shown in line R7 (89.56 μm).
Analyzing flavonoid and phenolic acid content, the lowest values for both parameters were detected in line R4 (219 mg/100 g and 1.64 mg/100 g, respectively), while the highest were found in line R9 (1208 mg/100 g and 9.13 mg/100 g, respectively) (Table 1).
A positive correlation between phenolic acid content and flavonoid content was observed with an R value of 0.9293 (p < 0.05).
3.2. Analysis of Tassel Morphology
We measured the length of tassel branches and tassel weight and counted the number of tassel branches and total number of spikelets in inbred lines B73 and Mo17, their F1 and F2 generations, three commercial hybrids, and five landraces (Figure 2 and Table 2). The analysis of tassel morphology showed that the number of lateral branches is higher in local landraces, ranging from 20 to 33, in comparison with modern hybrids, ranging from 6 to 8. Inbred lines Mo17 and B73 showed values of 11 and 12, respectively, with tassels of the F1 and F2 progenies showing 8 branches (Table 2).

Figure 2.
Tassels analyzed to measure number and length of branches, tassel weight, and total spikelet number. From (A–D) tassel of inbred line and their F1 and F2 progeny. (A) R6708 B73; (B) R6706 Mo17; (C) R6603 F1 B73/Mo17; (D) R6732 F2 B73/Mo17. From (E–G) Hybrid tassels (E) R 6600 Hybrid PI 817 Pioneer FAO 700; (F) R 6601 Hybrid LG 32.85 Limagrain FAO 200; (G) R 6602 Hybrid PR33A46 Pioneer FAO 500. From (H–L) landrace tassels (H) R6571 Spinoso nero; (I) R6573 Rostrato rosso di Rovetta; (J) R6637 Spinato di Gandino; (K) R6639 Mais delle Fiorine; (L) R6640 Millo Corvo. The same ruler is present in each individual image as a size standard.
Table 2.
Different parameters collected on tassels of the different maize genotypes tested (n = 5). Mean and SD are reported. For each parameter analyzed, different letters indicate statistically significant differences (Tukey’s test, p < 0.05).
To find the easiest and most accurate way to estimate the total number of spikelets, correlations were made using the total length of tassel branches and tassel weight to elaborate two regression equations: one between the tassel weight and spikelet number and another between the total branch length and spikelet number (Figure 3A,B). The values of the correlation coefficient R indicated that the regression equation based on the length of tassel branches (R: 0.9655) will give more accurate estimates of the total number of spikelets than the regression equation based on tassel weight (R: 0.9281) (Figure 3A,B).
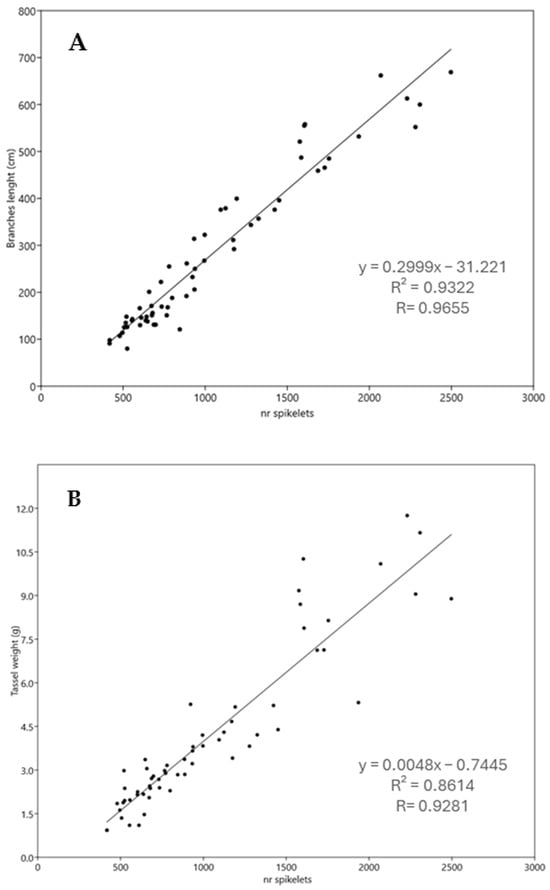
Figure 3.
Regression lines between tassel branch length (A) or tassel weight (B) and the total number of spikelets. In (A), the regression line illustrates a significant positive correlation between tassel branch length and the total number of spikelets (r = 0.9655). (B) shows a significant positive correlation between tassel weight and the total number of spikelets, with correlation (r = 0.9281).
3.3. “Smart” Pollination Accuracy
To verify the accuracy of “smart” pollinations, we compared the frequency of pollinations with unwanted pollen in canonical and “smart” pollination (Table 3 and Figure 4). This frequency was measured as the percentage of out-of-type seeds/plants present in the progeny of controlled self-pollinations of su1/su1 and br2/br2 plants, and in the F1 progeny of the cross pl1/pl1 × Pl1/Pl1, with plants Pl1/Pl1used as male parents.
Table 3.
Cross-contamination comparison between canonical and “smart” pollination. Inbred lines carrying sugary1 (su1, wrinkled seed) and brachytic2 (br2, short plant) recessive mutations were self-pollinated, and the progeny obtained were scored for the off-type phenotypes. We also crossed the inbred line carrying Purple plant1 allele (Pl1, red anther) as the pollen donor with the B73 inbred line (pl1/pl1, yellow anther). Cross-contamination was detected as wt seeds on su1/su1 ears, while in the progeny of br2/br2 plants, the out-of-type plants were tall, and in the progeny of Pl1/Pl1 plants, the out-of-type plants had yellow anthers. Cross-contamination frequency is shown as percentage ± SD. Different letters indicate statistically significant differences (Tukey’s test, p < 0.05).

Figure 4.
The off-type phenotypes observed in canonical and “smart” pollinations. (A) Wild-type seeds (off-type) on an ear homozygous for the sugary1 mutation. (B) Wild-type plant (off-type) in the progeny of the self-pollination of a plant homozygous for brachytic2 mutation. (C) Plant with yellow anthers (off-type) in the Pl/pl progeny showing the phenotype “red anthers”. The off-type phenotypes are indicated by white arrows.
In the progeny of sugary1 plants, the out-of-type phenotype was represented by the wild-type seeds. In the progeny of brachytic2 plants, the out-of-type phenotype was represented by the wild-type plants, identified and counted in the next field season. In the F1 progeny Pl1/pl1, the out-of-type phenotype was represented by the plants with yellow anthers, identified and counted in the next field season. As reported in Table 3, the two different methods of pollination gave almost the same results, suggesting that “smart” pollination is a rapid and simple method to produce hybrids and maintain inbred lines. Figure 4 shows the phenotype of the off-type progeny for each cross.
3.4. Pollen Longevity
To test how long pollen maintains its ability to fertilize, we crossed the hybrid B73 x Mo17, used as the female parent, with the B73 inbred line used as the pollen donor. We carried out pollinations starting immediately after pollen emission from anthers (at 10 AM) and then at 8 subsequent times, up to 7 days after pollen collection. After harvesting, we classified the ears, as reported in the materials and methods section, according to seed set into five different categories, ranging from 0 to 100% of seeds (Table 4). Good seed production was observed with pollen stored for up to 48 h (showing between 84 and 99% of seeds produced). After three days of pollen storage, seed production dropped around 40%, and after five days, the pollen had totally lost its ability to fertilize (Table 4).
Table 4.
Seed set (mean ± SD) obtained using pollen stored for different times (from 1 h to 7 days). The four categories refer to different seed sets ranging from 0% (category 1) to 100% (category 4). Different letters indicate statistically significant differences (Tukey’s test, p < 0.05).
4. Discussion
Seed production and yield are crucial aspects in the cultivation of maize, influenced by both environmental and genetic factors that affect pollen quantity and viability. Currently, most maize cultivated worldwide consists of hybrids from different FAO classes. Over the last 50 years, breeding programs have led to a decrease in the number of tassel lateral branches to enhance plant light perception and reduce tassel competition with the ear for nutrients [25,44]. Literature data have shown a negative correlation between the tassel weight and maize yield [28], with tassel weight decreasing by 36% from 1967 to 1991 [25,28]. However, to ensure the production of high amounts of hybrid seeds, abundant and prolonged pollen availability is required [44], which is dependent on tassel architecture, i.e., the total branch length and total number of spikelets [29].
This study aimed to examine different parameters related to pollen production and functionality in maize (Table 1 and Table 2). Both genetic and environmental factors strongly influence pollen production and activity. Environmental factors, such as drought stress, UV rays, and high temperatures, play a pivotal role [16,18,20,45]. Drought stress during pollen formation is related to sterile pollen production. Two peaks of sensitivity to drought have been described: the first, from meiosis to tetrad break-up in anthers, induces pollen sterility, while the second, during the initial stages of grain development, causes abnormalities in floral organs and the abortion of newly formed grains [16]. Pollen viability after anthesis is influenced by its water content, which depends on the temperature and relative humidity conditions in the atmosphere [45]. UV rays also impact pollen viability. Early in the day, when pollen dispersal begins, UV rays are at their lowest level but are sufficient to reduce pollen viability by 10%, increasing to 20% at midday [46]. Therefore, together with genetic factors, these environmental factors significantly influence pollen longevity and thus the period during which controlled pollinations can be carried out. Knowledge of this period is essential in genetic improvement programs, especially for crosses between lines with different flowering times. A common strategy involves staggered sowing to synchronize flowering, but this can fail under environmental stresses such as high temperatures, drought, and high UV levels.
In the 2021 season, we measured how long pollen collected from a tassel and stored in a paper bag at an ambient temperature could be preserved while maintaining its fertility. Our data showed that up to 48 h after collection, pollen maintained good fertility. After 72 h, fertility was significantly reduced, and after five days, pollen nearly lost its fertility (Table 4). These results align with previously reported data indicating pollen longevity at ambient temperature ranging from 1–2 h up to 2 days [19,47,48]. This range can be explained by factors such as ambient temperature and relative humidity, which have been reported to influence pollen longevity [48], in addition to the fact that different accessions were chosen in the different papers. Unfortunately, in the 2022 season, continuous heat waves and very low rainfall prevented us from repeating the experiment to confirm the previous year’s data on pollen viability (Supplementary Figure S3).
Among the genetic factors affecting pollen germination, an important role is played by genes involved in flavonoid biosynthesis. Flavonoids are a class of compounds that have been reported to protect DNA and cellular membranes from damage caused by UV-B rays [24]. UV-B radiation increases ROS and lipid peroxidation and decreases antioxidant activities, reducing pollen germination, tube growth, and fertilization capacity [20]. We analyzed flavonoid and phenolic acid content in 11 commercial inbred lines, looking for correlations with pollen traits such as germination percentage and grain diameter. Our data did not show any correlation between these parameters, except for a positive correlation between flavonoid and phenolic acid content, as expected, since both metabolites are products of the phenylpropanoid pathway [49]. The literature data have reported a positive correlation between pollen fertility and flavonoid content. Maize mutants lacking chalcone synthase are flavonoid-deficient and male-sterile due to failure to produce functional pollen tubes [50]. Our results suggested that the flavonoid levels in the 11 commercial inbred lines analyzed reach the threshold level necessary to ensure pollen germination and fertility (Table 1).
Breeding programs aimed at increasing grain yield over the last 50 years have resulted in modern hybrids with significantly fewer tassel branches compared to landraces [25] (Table 2 and Figure 2). Spikelet number is positively correlated with tassel length and weight [27,29]. Therefore, modern hybrids, due to reduced tassel branches, produce fewer pollen grains. This could result in pollen production insufficient to ensure a good seed set, especially under environmental stresses that are common during summer such as high temperatures and drought [17,18,51].
Tassel branch number (TBN) is a key trait determining tassel morphology and is a primary determinant of tassel size. TBN is a quantitative trait controlled by multiple genes. The literature data have reported the identification of 21 QTLs, with 7 being major QTLs, suggesting TBN is controlled by major QTLs, with modification by minor QTLs [44]. A more recent paper reported the identification of 13 QTLs, with 2 being major QTLs, and 22 quantitative trait nucleotides (QTNs) [52]. Eight of these QTLs are localized on chromosomes 3 and 7, hotspots for QTLs containing genes associated with tassel development, i.e., Lg2 (Liguleless2), Na1 (Nana plant 1), Ba1 (Barren stalk1), Sid1 (Sister of indeterminate spikelet1), Tsh4 (Tasselsheath4), and Ra3 (Ramosa 3) [52].
The developmental processes of male and female inflorescences are very similar, with several mutations influencing both tassel and ear traits. Maize mutants in the three Ramosa (Ra) genes, Ra1, Ra2, and Ra3, are characterized by higher TBNs and branched ears, suggesting a role in meristem determinacy of these genes, with Ra1 pivotal in the determination of plant inflorescence morphology [31,53,54]. In particular, the mutant ra1-154, bearing a deletion in the putative zing-finger domain, indicated a role for RA1 as a DNA-binding protein [32]. The overexpression of this gene in A. thaliana increased reproductive and vegetative organ size, suggesting a further possible role of the Ra1 gene in cell expansion regulation [55]. RAMOSA ENHANCER LOCUS2 (REL2), encoding a transcriptional repressor and physically interacting with RA1, has been reported to enhance the mutant phenotype of ra1 and ra2 [31].
A potential strategy to increase tassel branching and therefore pollen production in a line of interest is the use of new breeding technologies based on CRSPR-Cas technology. The site-specific gene knockout mediated by CRISPR-Cas allows for improved phenotypes in the desired background. CRISPR-Cas gene editing has been used in maize to validate the role played by genic male-sterility (GMS) genes in pollen and anther development [56,57,58] to study genes involved in plant and inflorescence architecture in wheat [59], rice and maize [60], tomato [61,62], Solanaceae [63], and Rapeseed [64] and to generate male sterile mutant in rice, soybean, maize, and tomato [65]. Based on the phenotype of Ramosa mutants described in the literature, knocking out the Ramosa1 gene using CRISPR-Cas would increase the number of tassel branches (Figure 5A). In particular, considering the highly branched tassel driven by the ra-154 mutation [32], genome editing should be directed to the zinc-finger domain of RA1 (Figure 5B,C). The advantage of using CRISPR-Cas technology is that increased pollen production can be achieved without altering the genetic structure of the line under study, unlike traditional breeding methods. In fact, this precise breeding approach allows the knock-out of the gene of interest without compromising the genetic background that has already been enhanced.
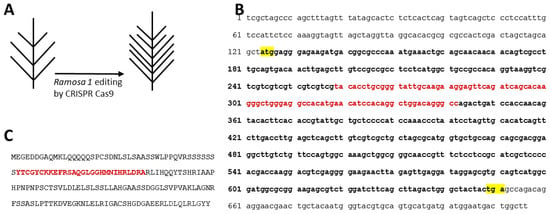
Figure 5.
Genome editing on putative target, Ramosa 1 (ra1) transcription factor, to improve the number of tassel branches (A). Ra1, mRNA sequence (GeneID:100276104, chr. 7, 716 bp). The start and stop codons are shown in yellow and the Zinc finger C2H2 type domain, putative genome editing target, is shown in red (B). Ramosa1 C2H2 zinc-finger transcription factor protein (175 aa), where the Zinc finger C2H2 type domain is shown in red (C).
Furthermore, we proposed a fast and easy method for controlled pollination (“smart” pollination), and we presented data about its accuracy through quantification of the frequency of out-of-type progeny in “smart” and conventional pollinations (Table 3 and Figure 4). These data suggested that “smart” pollinations provide nearly the same accuracy as the more laborious and time-consuming canonical pollination. Literature data have reported the use of kernel phenotypic traits to control pollen contamination [66,67], but data regarding the percentage of off-type seeds are missing. Regarding the progeny of Pl1/pl1 plants, the significantly higher presence of off-type (1.78%) compared to crosses with other phenotypic markers (ranging from 0.38% to 0.6%) could be due to two phenomena: higher levels of pollen contamination or the combined effect of pollen contamination with the paramutation phenomenon, determining the inactivation of the Pl allele [68,69,70].
Participatory Plant Breeding (PPB) is a form of plant breeding where breeding programs are carried out by scientists/breeders in collaboration with local farmers so that user preferences and the socio-economic environment can also be taken into account [71]. In particular, PPB approaches enhance the acceptability of new crop varieties among disadvantaged farmers by incorporating their preferences into the criteria for developing, testing, and releasing these varieties [71]. The “smart” pollination method we proposed is simpler, less time-consuming, and less labor-intensive, yet as efficient as the classic method. This approach could be easily used by farmers in PPB to propagate and select open-pollinated varieties (OPV) in purity and, less frequently, hybrid varieties, even without a breeder’s experience. These activities, in fact, are often performed in the best fields, where different genotypes are cultivated in adjacent areas, with the possibility of cross-contamination. Once the best materials are selected, their multiplication will be performed using spatial isolation, without the need for artificial pollination.
Another easy procedure that can be used in PPB programs is the conservation of pollen for several days at room temperature, allowing crosses between lines, which, due to the genetic characteristics or the effect of the environment, flower asynchronously.
5. Conclusions
Pollen production and vitality are important issues in breeding programs for grain and seed production. In this paper, we proposed the use of a simple method, based on measuring the tassel weight, to estimate the number of spikelets as an indicator of the amount of pollen produced. Additionally, we propose that “smart” pollination and storing maize pollen for up to 2–3 days at an ambient temperature are two methods that can be used to implement breeding programs, also in areas with little access to modern technologies, and in particular in PPB projects. Furthermore, to increase pollen production, we suggested the use of the CRISPR-Cas genome editing technique to create loss-of-function mutants in genes regulating the branch number as Ra1, resulting in increased tassel branches and increased pollen production, in the selected genetic background.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14101791/s1, Figure S1: Tassel at 50% of anthesis (left) and pollen grains germinated on a semi-solid medium (right); Figure S2: Examples of ears belonging to the different categories used to classify the ears according to the seed set. From left to right: category 1: 0–25% of seed set; category 2: 25–50% of seed set; category 3: 50–75% of seed set; category 4: 75–100% of seed set); Figure S3: Climatic data collected in the years 2021–2022. A: temperatures, B: rainfalls, C: % of relative humidity. The year 2022 was characterized by exceptionally high temperatures, very scarce rainfall, and low RH. Pollen collection and hand pollinations were performed between the 15 and 25 of July.
Author Contributions
Conceptualization, R.P.; methodology, R.P., S.S., E.C. and M.L.; software, R.P.; validation, S.S. and R.P.; formal analysis, S.S., M.G., E.C. and M.L.; investigation, S.S. and R.P.; resources, R.P.; data curation, M.G., E.C. and M.L.; writing—original draft preparation, S.S.; writing—review and editing, R.P. and M.L.; visualization, S.S.; supervision, R.P.; project administration, R.P.; funding acquisition, R.P. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This publication is part of the projects NODES, which received funding from the MUR—M4C2 1.5 of PNRR funded by the European Union—NextGenerationEU (Grant agreement no. ECS00000036) (to M.L.) and Agritech National Research Centre, who received funding from the European Union NextGenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)–MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4–D.D. 1032 17/06/2022, CN00000022) (to R.P.).
Institutional Review Board Statement
This manuscript reflects only the authors’ views and opinions; neither the European Union nor the European Commission can be considered responsible for them.
Data Availability Statement
This article includes all data generated or analyzed during this study; further inquiries can be directed to the corresponding authors.
Acknowledgments
We wish to thank Davide Reginelli for his work in the field.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ertdman, G. Pollen Morphology and Plant Taxonomy, Angiosperms; Almqvist and Wiksell: Stockholm, Sweden, 1952. [Google Scholar] [CrossRef]
- Smith, E.G. Sampling and Identifying Allergenic Pollens and Molds; Blewstone Press: San Antonio, TX, USA, 1990; ISBN 0930961021/9780930961022. [Google Scholar]
- Percival, M.S. Pollen presentation and pollen collection. New Physiol. 1950, 49, 40–63. [Google Scholar] [CrossRef]
- Purseglov, J.W. Tropical Crops. Monocotyledons; Longiman Group: London, UK, 1972. [Google Scholar]
- Shull, G.H. The composition of a field of maize. J. Hered. 1908, 4, 296–301. [Google Scholar] [CrossRef]
- Shull, G.H. What is Heterosis? Genetics 1948, 33, 439–446. [Google Scholar] [CrossRef]
- Shull, G.H. Beginnings of the heterosis concept. In Heterosis; Gowen, J.W., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 14–48. [Google Scholar]
- East, E.M. Heterosis. Genetics 1936, 21, 375–397. [Google Scholar] [CrossRef]
- Rodgers, H.J.; Parkes, H.C. Transgenic plants and the environment. J. Exp. Bot. 1995, 46, 467–488. [Google Scholar]
- Louette, D.; Charrier, A.; Berthaud, J. In situ conservation of maize in Mexico: Genetic diversity and maize seed management in a traditional community. Econ. Bot. 1997, 51, 20–38. [Google Scholar] [CrossRef]
- Garcia, C.; Figueroa, M.J.; Gomez, L.; Towsend, R.; Schoper, J. Pollen control during transgenic hybrid maize development in Mexico. Crop Sci. 1998, 38, 1597–1602. [Google Scholar] [CrossRef]
- Oz, A.; Tugay, M.E. Variation on some agronomic characters in selfing generations in corn (Zea mays indentata Sturt). JAFAG 2003, 20, 123–132. [Google Scholar]
- Ogden, E.C.; Hayes, J.V.; Raynor, G.S. Diurnal patterns of pollen emission in Ambrosia, Phleum, Zea and Ricinus. Am. J. Bot. 1969, 56, 16–21. [Google Scholar] [CrossRef]
- Jarosz, N.; Loubet, B.; Durand, B.; Mc Cartney, H.A.; Foueillassar, X.; Huber, L. Field measurement of airborne concentration and deposition rate of maize pollen (Zea mays L.) downwind of an experimental field plot. Agric. For. Meteorol. 2003, 119, 37–51. [Google Scholar] [CrossRef]
- Kiesselbach, T.A. The Structure and Reproduction of Corn; Research Bulletin; University of Nebraska Press: Lincoln, NE, USA, 1949; p. 161. [Google Scholar]
- Saini, H. Effects of water stress on male gametophyte development in plants. Sex. Plant Reprod. 1997, 10, 67–73. [Google Scholar] [CrossRef]
- Barnabás, B. Effect of water loss on germination ability of maize (Zea mays L.) pollen. Ann. Bot. 1985, 55, 201–204. [Google Scholar] [CrossRef]
- Buitink, J.; Walters-Vertucci, C.; Hoekstra, F.A.; Leprince, O. Calorimetric properties of dehydrating pollen (analysis of a desiccation-tolerant and an intolerant species). Plant Physiol. 1996, 111, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Luna, S.; Figueroa, J.M.; Baltzar, M.; Gomez, L.; Towsend, R.; Schoper, J.B. Maize pollen longevity and distance isolation requirements for effective pollen control. Crop Sci. 2001, 41, 1551–1557. [Google Scholar] [CrossRef]
- Wang, S.; Xie, B.; Yin, L.; Duan, L.; Li, Z.; Eneji, A.E.; Tsuji, W.; Tsunekawa, A. Increased UV-B radiation affects the viability, reactive oxygen species accumulation and antioxidant enzyme activities in maize (Zea mays L.) pollen. Photochem. Photobiol. 2010, 86, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bruckmann, A.; Dresselhaus, T.; Begcy, K. Heat stress at the bicellular stage inhibits sperm cell development and transport into pollen tubes. Plant Physiol. 2024, 195, 2111–2128. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, X.; Sheng, D.; Hou, X.; Mandal, S.; Liu, X.; Zhang, P.; Shen, S.; Wang, P.; Krishna Jagadish, S.V.; et al. Heat-dependent postpollination limitations on maize pollen tube growth and kernel sterility. Plant Cell Environ. 2023, 46, 3822–3838. [Google Scholar] [CrossRef]
- Mo, Y.; Nagel, C.; Taylor, L.P. Biochemical complementation of chalcone synthase mutants defines a role for flavonols in functional pollen. Proc. Natl. Acad. Sci. USA 1992, 89, 7213–7217. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shirley, B.V. Flavonoids biosynthesis: “new” functions for an “old” pathway. Trends Plant Sci. 1996, 1, 11. [Google Scholar]
- Duvick, D.N.; Cassmann, K.G. Post-green revolution trends in yield potential of temperate maize in the North-Central United States. Crop Sci. 1999, 39, 16221630. [Google Scholar] [CrossRef]
- Postlethwait, S.N.; Nelson, O.E. Characterization of development in maize through the use of mutants. The polytypic (Pt) and ramosa-1 (Ra1) mutants. Am. J. Bot. 1964, 51, 238–243. [Google Scholar] [CrossRef]
- Fonseca, A.E.; Westgate, M.E.; Grass, L.; Dornbos, D.L. Tassel Morphology as an Indicator of Potential Pollen Production in Maize. Crop Manag. 2003, 2, 1–15. [Google Scholar] [CrossRef]
- Bódi, Z.; Pepó, P.; Kovács, A. Morphology of tassel components and their relationship to some quantitative features in maize. Cereal Res. Commun. 2008, 36, 353–360. [Google Scholar] [CrossRef]
- Ricci, B.; Monod, H.; Guérin, D.; Messéan, A.; Maton, C.; Balique, B.; Angevin, F. Predicting maize pollen production using tassel morphological characteristics. Field Crop Res. 2012, 136, 107–115. [Google Scholar] [CrossRef]
- Dari, S.; MacRobert, J.; Labuschagne, M. Effect of the few-branched-1 (Fbr1) tassel mutation on performance of maize inbred lines and hybrids evaluated under stress and optimum environments. Maydica 2016, 62. [Google Scholar]
- Gallavotti, A.; Long, J.A.; Stanfield, S.; Yang, X.; Jackson, D.; Vollbrecht, E.; Schmidt, R.J. The control of axillary meristem fate in the maize ramosa pathway. Development 2010, 137, 2849–2856. [Google Scholar] [CrossRef]
- Cassani, E.; Landoni, M.; Pilu, R. Characterization of the Ra1 mazie gene involved in inflorescence architecture. Sex. Plant Reprod. 2006, 19, 145–150. [Google Scholar] [CrossRef]
- Krieger, K.M.; Pollak, L.M.; Brumm, T.J.; White, P.J. Effects of pollination method and growing location on starch thermal properties of corn hybrids. Cereal Chem. 1998, 75, 656–659. [Google Scholar] [CrossRef]
- Letchworth, M.B.; Lambert, R.J. Pollen parent effects on oil, protein, and starch concentration in maize kernels. Crop Sci. 1998, 38, 363–367. [Google Scholar] [CrossRef]
- Sulewska, H.; Adamczzyk, H.; Gygert, J.; Rogacki, G.; Szymanska, K.; Smiatacz, K.; Panasiewicz, K.; Tomaszyk, K. A comparison of controlled self-pollination and open pollination results based on maize grain quality. Span. J. Agric. Res. 2014, 12, 492–500. [Google Scholar] [CrossRef]
- Kahriman, F.; Egesel, C.O.; Aydin, T.; Subasi, S. The role of artificial pollination and pollen effect on ear development and kernel structure of different maize genotypes. J. Pollinat. Ecol. 2015, 15, 6–14. [Google Scholar] [CrossRef]
- Kahriman, F.; Egesel, C.O.; Zorlu, E. Effects of open and self-pollination treatments on genetic estimations in maize diallel experiment. Span. J. Agric. Res. 2015, 13, e0704. [Google Scholar] [CrossRef]
- Broussard, M.A.; Coates, M.; Martinsen, P. Artificial Pollination Technologies: A Review. Agronomy 2023, 13, 1351. [Google Scholar] [CrossRef]
- Lago, C.; Landoni, M.; Cassani, E.; Cantaluppi, E.; Doria, E.; Nielsen, E.; Giorgi, A.; Pilu, R. Study and characterization of an ancient European flint white maize rich in anthocyanins: Millo Corvo from Galicia. PLoS ONE 2015, 10, e0126521. [Google Scholar]
- Giupponi, L.; Leoni, V.; Colombo, F.; Cassani, E.; Hejna, M.; Rossi, L.; Pilu, R. Characterization of “Mais delle Fiorine” (Zea mays L.) and nutritional, morphometric and genetic comparison with other maize landraces of Lombardy region (Northern Italy). Genet. Resour. Crop Evol. 2021, 68, 2075–2091. [Google Scholar] [CrossRef]
- Sangiorgio, S.; Colombo, F.; Ghidoli, M.; Giupponi, L.; Ferro, G.; Ferro, C.G.; Cassani, E.; Landoni, M.; Pilu, R. The ancient varieties of mountain maize: The inheritance of the pointed character and its effect on the natural drying process. Agronomy 2021, 11, 2295. [Google Scholar] [CrossRef]
- Lago, C.; Landoni, M.; Cassani, E.; Atanassiu, S.; Cantaluppi, E.; Pilu, R. Development and characterization of a coloured sweet corn line as a new functional food. Maydica 2014, 59, 191–200. [Google Scholar]
- Landoni, M.; Cassani, E.; Ghidoli, M.; Colombo, F.; Sangiorgio, S.; Papa, G.; Adani, F.; Pilu, R. Brachytic2 mutation is able to counteract the main pleiotropic effects of brown midrib3 mutant in maize. Sci. Rep. 2022, 12, 2446. [Google Scholar] [CrossRef]
- Chen, Z.J.; Cong, Y.; Tang, D.G.; Zhang, L.; Zhang, L.; Qu, J.T.; Jian, L. Dissection of the genetic architecture for tassel branch number by QTL analysis in two related populations in maize. J. Integr. Agric. 2017, 16, 1432–1442. [Google Scholar] [CrossRef]
- Aylor, D.E. Rate of dehydration of corn (Zea mays L.) pollen in the air. J. Exp. Bot. 2003, 54, 2307–2312. [Google Scholar] [CrossRef]
- Torabinejad, J.; Caldwell, M.M.; Flint, S.D.; Durham, S. Susceptibility of pollen to UV-B radiation: An assay of 34 taxa. Am. J. Bot. 1998, 85, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Youmbi, E.; The, C.; Tedjacno, A. Conservation of the germination capacity of pollen grains in three varieties of maize (Zea mays L.). Grana 2005, 44, 152–159. [Google Scholar] [CrossRef]
- Kaefer, K.A.C.; Chiapetti, R.; FogaÃ, L.; Muller, A.L.; Calixto, G.B.; Dallâ, E.I.; Chaves, Ã. Viability of maize pollen grains in vitro collected at different times of the day. Afr. J. Agric. Res. 2016, 11, 1040–1047. [Google Scholar]
- Lephatsi, M.; Nephali, L.; Meyer, V.; Piater, L.A.; Buthelezi, N.; Dubery, I.A.; Opperman, H.; Brand, M.; Huyser, J.; Tugizimana, F. Molecular mechanisms associated with microbial biostimulant-mediated growth enhancement, priming and drought stress tolerance in maize plants. Sci. Rep. 2022, 21, 12–10450. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions and biotechnological applocations. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef]
- Westgate, M.E.; Lizaso, J.; Batchelor, W. Quantitative relationships between pollen shed density and grain yield in maize. Crop Sci. 2003, 43, 934–942. [Google Scholar] [CrossRef]
- Ruidong, S.; Shijin, H.; Yuwei, Q.; Yimeng, L.; Xiaohang, Z.; Ying, L.; Xihang, L.; Mingyang, D.; Xiangling, L.; Fenghai, L. Identification of QTLs and their candidate genes for the number of maize tassel branches in F2 from two higher generation sister lines using QTL mapping and RNA-seq analysis. Front. Plant Sci. 2023, 14, 1202755. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bortiri, E.; Chuck, G.; Vollbrecht, E.; Rocheford, T.; Martienssen, R.; Hake, S. ramosa2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell 2006, 18, 574–585. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Satoh-Nagasawa, N.; Nagasawa, N.; Malcomber, S.; Sakai, H.; Jackson, D. A trehalose metabolic enzyme controls inflorescence architecture in maize. Nature 2006, 441, 227–230. [Google Scholar] [CrossRef]
- Landoni, M.; Cassani, E.; Pilu, R. Arabidopsis thaliana plants overexpressing Ramosa 1 maize gene show an increase in organ size due to cell expansion’. Sex. Plant Reprod. 2007, 20, 191–198. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, J.; Wei, X.; Wu, S.; Fang, C.; Li, Z.; Qi, Y.; Gao, Y.; Dong, Z.; Wan, X. Genetic Structure and Molecular Mechanisms Underlying the Formation of Tassel, Anther, and Pollen in the Male Inflorescence of Maize (Zea mays L.). Cells 2022, 11, 1753. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S.; Jiang, Y.; Yan, T.; Fang, C.; Hou, Q.; Wu, S.; Xie, K.; An, X.; Wan, X. Use of CRISPR/Cas9-Based Gene Editing to Simultaneously Mutate Multiple Homologous Genes Required for Pollen Development and Male Fertility in Maize. Cells 2022, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Zhang, S.; Jiang, Y.; Liu, X.; Fang, C.; Wang, J.; Zhao, L.; Hou, Q.; Zhang, J.; Xiangyuan Wan, X.; et al. CRISPR/Cas9-based genome editing of 14 lipid metabolic genes reveals a sporopollenin metabolon ZmPKSB-ZmTKPR1-1/-2 required for pollen exine formation in maize. Plant Biotechnol. J. 2024, 22, 216–232. [Google Scholar] [CrossRef]
- Errum, A.; Rehman, N.; Uzair, M.; Inam, S.; Ali, G.M.; Khan, M.R. CRISPR/Cas9 editing of wheat Ppd-1 gene homoeologs alters spike architecture and grain morphometric traits. Funct. Integr. Genom. 2023, 23, 66. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lindsay, P.L.; Jackson, D. Next generation cereal crop yield enhancement: From knowledge of inflorescence development to practical engineering by genome editing. Int. J. Mol. Sci. 2021, 22, 5167. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J. Genome editing of tomatoes and other Solanaceae. In Genome Editing for Precision Crop Breeding; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 421–440. [Google Scholar]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering quantitative trait variation for crop improvement by genome editing. Cell 2017, 171, 470–480. [Google Scholar] [CrossRef]
- Kwon, C.T.; Heo, J.; Lemmon, Z.H.; Capua, Y.; Hutton, S.F.; Van Eck, J.; Park, S.J.; Lippman, Z.B. Rapid customization of Solanaceae fruit crops for urban agriculture. Nat. Biotechnol. 2020, 38, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Sriboon, S.; Li, H.; Guo, C.; Senkhamwong, T.; Dai, C.; Liu, K. Knock-out of TERMINAL FLOWER 1 genes altered flowering time and plant architecture in Brassica napus. BMC Genet. 2020, 21, 52. [Google Scholar] [CrossRef]
- Farinati, S.; Draga, S.; Betto, A.; Palumbo, F.; Vannozzi, A.; Lucchin, M.; Barcaccia, G. Current insights and advances into plant male sterility: New precision breeding technology based on genome editing applications. Front. Plant Sci. 2023, 14, 1223861. [Google Scholar] [CrossRef]
- Della Porta, G.; Ederle, D.; Bucchini, L.; Prandi, M.; Verderio, A.; Pozzi, C. Maize pollen mediated gene flow in the Po valley (Italy): Source–recipient distance and effect of flowering time. Eur. J. Agron. 2008, 28, 255–265. [Google Scholar] [CrossRef]
- Devos, Y.; Reheul, D.; de Schrijver, A. ‘The co-existence between transgenic and non-transgenic maize in the European Union: A focus on pollen flow and cross-fertilization’. Environ. Biosafety Res. 2005, 4, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Hollick, J.B.; Patterson, G.I.; Asmundsson, I.M.; Chandler, V.L. ‘Paramutation alters regulatory control of the maize pl locus’. Genetics 2000, 154, 1827–1838. [Google Scholar] [CrossRef] [PubMed]
- Pilu, R. Paramutation: Just a Curiosity or Fine Tuning of Gene Expression in the next generation? Curr. Genom. 2011, 12, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Pilu, R. Paramutation phenomena in plants. Semin. Cell. Biol. 2015, 44, 2–10. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Decentralized-participatory plant breeding: An example of demand driven research. Euphytica 2007, 155, 349–360. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).