Abstract
Paphiopedilum micranthum, an IUCN Red List species, is discontinuously distributed in the karst limestone mountain of southwest China and exhibits ecological specialization, typically through lithophytic and terrestrial ecotypes. Whether the distribution of rhizosphere bacteria and fungi in these different habitats is random or reflects soil preferences requires further investigation. A total of 73 samples from the core distribution areas in China, representing all habitats in two sites, were analyzed for soil differences by comparing edaphic properties and microbial community structure based on high-throughput sequencing of bacterial 16S rRNA genes and fungal ITS region sequences, alongside soil physiochemical data. The results showed no significant differences in microbial community richness and diversity across the heterogeneous habitats. However, significant differences in taxa were observed across various habitats. Dominant bacterial phyla included Actinobacteriota, Proteobacteria and Acidobacteriota, with dominant genera such as Crossiella, Pseudonocardia, 67-14, Mycobacterium and RB41. The primary fungal phyla were Basidiomycota and Ascomycota, featuring prominent genera such as Phlegmacium, Archaeorhizomyces, Trechispora, and Lepiota. There were 16 bacterial genera and 13 fungal genera associated with nitrogen transformation and fixation. Alkali-hydrolyzed nitrogen (AN) was identified as a main driver of soil bacterial and fungal community variation. Based on an analysis of soil physicochemical properties, ammonium nitrogen content was consistently higher than nitrate nitrogen across different habitats. Furthermore, across all heterogeneous habitats, P. micranthum showed no significant differences in nitrate nitrogen, ammonium nitrogen, or their ratio. The nitrogen-use efficiency of P. micranthum ranged from 7.73% to 9.87%, with the highest efficiency observed in the terrestrial habitat of Shedu. These results suggest that P. micranthum prefers habitats rich in organic matter and nitrogen, showing a preference for ammonium nitrogen uptake in natural conditions. Heterogeneous habitats affect plant nitrogen-use efficiency as well as changes in microbial community composition.
1. Introduction
The genus Paphiopedilum, listed in appendix I of CITES, is known as a rare and endangered orchid with high ornamental value, making it one of the most threatened orchid genera [1,2]. Paphiopedilum micranthum, belonging to the most primitive paphiopedilum, exhibits a narrow niche breadth, contributing to its discontinuous distribution at altitudes of 333–1648 m across the karst limestone regions of southeast Yunnan, southwest Guizhou, Guangxi China and North Vietnam [3,4]. The karst limestone ecosystem, marked by severe water and soil erosion, is characterized by thin, calcareous soils with nitrogen deficiencies [5].
The Wenshan area in southeast Yunnan is considered a central distribution area and a key spot for the ecological diversity of this species in China [4]. P. micranthum typically prefers higher elevations, steeper slopes, and north-facing aspects within evergreen and deciduous broad-leaved forests or humid shrublands [6]. Based on its varied topographical settings, it can be categorized into two ecotypes: lithophyte and terrestrial. Lithophytes grow almost exclusively on steep, rocky cliffs or rock crevices with shallow soil deposition. However, terrestrial forms are commonly found in moist, structured soils at the base of trees or in low shrubs, often forming dense groups, clusters, or blocks [6]. Universally, the species’ discontinuous, block-like distribution pattern suggests its ecological specialization [7]. The obligate lithophyte typically denotes species with specific habitat preferences [8]. Nitrogen limitation and its seasonal variation in subtropical limestone areas play a significant role in shaping the patchy distribution in karst ecosystems [9,10]. However, it remains unclear whether P. micranthum exhibits a distinct preference for specific nitrogen forms or if its nitrogen-use efficiency varies across its habitats.
Climatic factors such as solar radiation, temperature, precipitation during the warmest quarter, and elevation have significant effects on the geographical distribution of taxa within Paphiopedilum Subgenus Brachypetalum (Orchidaceae) in southwest China [11]. It is widely recognized that the geographical patterns of plants at small scales, especially in compact habitats, are profoundly influenced by edaphic conditions such as soil nutrients, pH, organic content, symbiotic relationships, and topography [12,13,14,15]. Certain habitat types or associated vegetation types are important determinants of orchid distribution [16,17]. Furthermore, the specificity of symbiotic relationships between the orchid family and mycorrhizal fungi suggests that mycorrhizal fungal communities within their root systems and surrounding soils can significantly influence orchid distribution and population dynamics [18,19].
Generally, soil physicochemical properties, enzyme activities, and microbial communities accurately reflect nutrient characteristics [5]. Tian et al. [20] discovered significant differences between P. micranthum and other Paphiopedilum species in terms of soil nutrients, enzyme activities, and fungal communities. Habitat preference characteristics were highlighted by Russula purpureovirescens, which made the largest contribution to the fungal community [20]. However, there is limited research on the spatial nutrient preferences of P. micranthum, and several key questions remain unaddressed: (1) The nitrogen absorption characteristics and nitrogen-use efficiency of P. micranthum in heterogeneous habitats remain poorly understood. (2) Are there variations in soil properties and microbial communities across different heterogeneous habitats?
Previous studies have focused on symbiotic endomycorrhizal fungus [21], geographic distribution [1,17], and population genetics and propagation with a conservation perspective [22,23]. To compare the influences of soil, topography, and vegetation on Paphiopedilum settlement and elucidate its soil preferences, we investigated the soil physicochemical characteristics and microorganism communities across three habitats representing typical ecotypes within the core distribution area in China. Soil samples from all habitat types were collected at two sites and analyzed for organic matter, total nitrogen, alkali-hydrolysable nitrogen, soil nitrate nitrogen, ammonium-N, available phosphorus, and pH. Fungi associated with plant roots were identified through DNA analysis of key fungal barcoding genes from root samples. Microbial populations were quantified using high-throughput sequencing of the 16S gene (for Bacteria/Archaea) and ITS region (for Fungi). Microbial community diversity and composition were used to analyze and compare the similarity among the four habitats.
2. Materials and Methods
2.1. Soil Sample Collection
The Wenshan limestone region in southeastern Yunnan serves as the original and evolutionary center, as well as the largest trading hub, for P. micranthum worldwide. Two core distribution sites of P. micranthum were selected: Xiaobazi (XBZ) mountain in Wenshan County (104°20′726″ E, 23°22′776″ N) and Shedu (SD) mountain in Maguan County(23°07′18″ N, 104°16′34″ E) (Figure 1).

Figure 1.
Different habitats of P. micranthum in different locations: (A) under tree base in Shedu—SDT; (B) under tree base in Xiaobazi—XBZT; (C) under shrub base in Xiaobazi—XBZS; (D) lithophyte in Xiaobazi—XBZL. Red arrows in the pictures showed the individuals of P. micranthum.
In accordance with the actual plant distribution, undisturbed and well-established natural sites of P. micranthum were selected in the two core distribution areas. Due to the discontinuous nature of P. micranthum’s natural distribution and significant human disturbance in some areas, the number of sampling points for each ecological type could not be equal. Three habitats including one lithophyte, and two terrestrial types were collected from XBZ, respectively, at an altitude of 1490–1510 m. The lithophyte ecotype of P. micranthum is primarily distributed in the northern slopes near mountainsides, growing predominantly on stone walls and in stone crevices. Terrestrial ecotype growing under tree foot and shrub habitats was mainly distributed in northwest of evergreen and deciduous broad-leaved forest. In SD, P. micranthum was primarily distributed on the north slopes near the mountain peaks at altitudes of 1360–1370 m (Table 1).

Table 1.
The habitat conditions of P. micranthum in sample plots of southeast Yunnan in China.
Soil samples were collected at a distance of 10~20 cm from the P. micranthum roots and passed through a 2-mm sieve to remove plant roots, debris, and stones. The rhizosphere soil was the portion of soil attached to the P. micranthum roots [24]. In February 2023, 4~6 P. micranthum plants were selected from each of the 14 plots (3 × 5 + 3 × 6 + 4 × 6 + 4 × 4 = 73 total plants), and their rhizosphere soil was collected. All selected plants exhibited 3 to 4 leaves, demonstrating uniform growth. Whole P. micranthum plants were removed from the ground and then the rhizosphere soil samples were collected by shaking the roots vigorously to separate the soil from the roots, and then, the sifted soil particles (1 mm diameter) were collected using a 1 mm sieve [25]. The rhizosphere soil from the 5~6 P. micranthum plants in each plot was mixed to obtain 14 composite rhizosphere soil samples. Similar to the rhizosphere soil samples, 14 bulk soil samples were collected at four habitats. In total, 28 soil samples (bulk soil 14 + rhizosphere soil 14 = 28) were prepared for analysis. The first part was stored in a portable refrigerator and stored at −20 °C for soil chemical property analysis after being transported to the laboratory. The second part was stored in sterilized centrifuge tubes in dry ice during transportation, then kept in an ultra-low-temperature refrigerator (−80 °C) at the laboratory for subsequent microbial diversity sequencing [26].
2.2. Determination of Soil Physicochemical Properties and Nitrogen
The analysis of soil physical and chemical properties was briefly introduced [27]. Soil pH was determined using the potentiometric method (pH 700 Jingheng Company, Jinan, China). Water content was assessed by the 105 °C drying-weighing method. Soil organic matter was determined by the alkali N-proliferation method. Available phosphorus (AP) in soil was measured using the molybdenum antimony anti-colorimetric method (TU-1900). Available potassium (AK) was extracted with neutral ammonium acetate (NH4OAc) solution and analyzed by flame photometry (FP6400) [28,29]. Total nitrogen (TN) was determined by the Kjeldahl method (K9840). Ammonium nitrogen and nitrate nitrogen were extracted with 2 mol/L KCl and analyzed using a UV spectrophotometer (TU-1900) [30]. The plants were divided into different parts, defoliated at 105 °C for 30 min, dried at 70 °C to a constant weight, and then weighed. Subsequently, the plant organs were ground through 100 mesh sieves, and the total nitrogen content was determined using the Neisser-colorimetric method [31]. Nitrogen use efficiency was calculated by nitrogen balance method. Nitrogen balance method refers to the ratio of crop nitrogen uptake to total nitrogen input [32,33].
2.3. DNA Extraction and Amplification with Polymerase Chain Reaction (PCR)
The OMEGA Soil DNA Kit (D5625-01) (Omega Bio-Tek, Norcross, GA, USA) was used to extract genomic DNA from samples, which was subsequently analyzed by 1% agarose gel electrophoresis. According to the selection of sequencing region, the selected variable region was amplified by PCR using specific primers with Barcode and high-fidelity DNA polymerase. Primers 341F (5′-CCTAYGGGRBGCASCAG-3′) and 860R (5′-GGACTACNNGGGTATCTAAT-3′) targeted the V3–V4 regions of 16S rRNA genes, while Primers 1713F (5′-GGAAGTAAAAGTCGT AACAAGG-3′) and 2023R (5′-GCTGCGTTCTTCATCGATGC-3′) targeted the fungal ITS-1 regions. The PCR conditions included an initial denaturation at 98 °C for 1 min, followed by 30 cycles of denaturation at 98 °C for 10 s, annealing at 50 °C for 30 s, and extension at 72 °C for 30 s, with a final extension at 72 °C for 5 min. PCR products were analyzed using 2% agarose gel electrophoresis, and the target fragments were subsequently excised and purified using the Quant-iT PicoGreen dsDNA Assay Kit (Omega Bio-Tek, Norcross, GA, USA). Referring to the preliminary quantitative results of electrophoresis, the PCR amplification recovered products were detected and quantified using the Microplate reader (BioTek, FLx800) fluorescence quantitative system, and the corresponding proportions were mixed according to the sequencing requirements of each sample. The library was constructed using TruSeq Nano DNA LT Library Prep Kit from Illumina (San Diego, CA, USA). The constructed library was inspected with an Agilent (Agilent Technologies Inc., Santa Clara, CA, USA) Bioanalyzer 2100 and Promega (Promega Beijing Biotech, Beijing, China) QuantiFluor. Once qualified, the library proceeded to sequencing.
2.4. Statistical Analysis
Paired-end reads were initially preprocessed using cutadapt(2.6.0) software to detect and remove adapters. After trimming, the paired-end reads underwent filtering for low-quality sequences, denoising, merging, and removal of chimera reads using DADA2 (1.26.0) with the default parameters in QIIME2 (21.11.0). Subsequently, the software generated representative reads and an abundance table of ASVs. Using the QIIME2 software package, representative sequences for each ASV were selected, annotated, and compared against a database using blast. ITS sequences were aligned against the Unite (v6) database (https://unite.ut.ee/ accessed on 15 June 2023), while 16S sequences were aligned against the Silva database (version 138) (https://www.arb-silva.de/ accessed on 15 June 2023). α and β Diversity indices were calculated using Qiime2 software. α Diversity analysis involved assessing species richness and diversity of microbial communities within each sample [34]. This included generating species accumulation box plots and calculating indices such as Chao1, ACE, Shannon, and Simpson. To analyze β diversity of microbial communities, Principal Coordinate Analysis (PCoA) based on the Bray-Curtis distance algorithm was performed. similarity within and between groups was assessed using analysis of similarity (Anosim). Taxonomic analysis was conducted to assess the composition of microbial communities at the phylum and genus levels. Significant differences in species between samples were identified using the LEfSe tool with an LDA score threshold of 4.
FUNGuild was used for functional prediction of soil fungi from different habitats. PICRUSt (2.3.0) software was used to infer the functional gene composition in the samples by comparing the species composition information obtained from 16S sequencing data, and to analyze differences in function between different samples or subgroups. Redundancy Analysis (RDA) was performed to investigate the relationship between microbial communities and soil physicochemical properties. The correlation between microbial communities at the genus level and soil properties was assessed using Spearman’s correlation coefficient. Differences in soil physicochemical properties across different habitats were evaluated using one-way ANOVA with a significance level set at p < 0.05, using SPSS software (25.0).
3. Results
3.1. Soil Physiochemical Properties of P. micranthum in Different Habitats
The physicochemical properties of P. micranthum were analyzed across different locations and ecotypes (Table 2). Soil pH was significantly higher in SDT than in XBZL compared to different sites. Soil OM content was significantly higher in XBZS than in SDT. Additionally, soil AN was significantly lower in SD compared to XBZL. Soil water content in SD was significantly higher than in tree foot and shrub foot habitats of XBZ. Soil AP was also significantly lower in SDT compared to XBZS. AK was significantly higher in XBZS compared to XBZL. However, total nitrogen, ammonium nitrogen, nitrate nitrogen, and the ratio of ammonium nitrogen to nitrate nitrogen did not show significant differences between SD and XBZ. When comparing different habitats within the same site, soil pH at XBZ ranged from 6.67 to 6.85, indicating weak acidity, with no significant differences observed between habitats. OM (289.50 g/kg), AN (725.25 mg/kg), and NO3−-N (4.29 mg/kg) were the highest in XBZS. In contrast, XBZT exhibited the lowest values with OM at 277.33 g/kg, AN at 656.67 mg/kg, and NO3−-N at 2.67 mg/kg, respectively. Soil water content in XBZL was the highest, with no significant differences observed between habitats. AP was highest in XBZT (11.53 mg/kg) and lowest in XBZL (9.43 mg/kg), while AK was highest in XBZT (177.33 mg/kg) and lowest in XBZL (141.00 mg/kg).
Table 2.
Soil physical and chemical properties and one-way ANOVA (analyses of variance) results of P. micranthum in different habitats.
In summary, among the rhizosphere soil collected from the four types of sample plots, soil physicochemical properties showed better homogeneity within the same geographical area. However, significant differences were observed in soil physicochemical properties among different geographical areas for several indicators, except for total nitrogen, nitrate nitrogen, and ammonium nitrogen. It is noteworthy that ammonia nitrogen levels were consistently higher than nitrate nitrogen across all sites. This observation suggests that Paphiopedilum species are highly selective in their choice of specific soil types. Therefore, strict criteria exist for selecting suitable habitats for P. micranthum in various soil environments. P. micranthum preferred to the specific soils with high organic matter, rich total nitrogen and high ammonium nitrogen, which soil nitrogen pool could maintain a stable ratio of ammonium nitrogen to nitrate nitrogen to improve nitrogen use and reduce its leaching under thin soils by converting the form of inorganic nitrogen.
3.2. Nitrogen Content and Nitrogen-Use Efficiency of P. micranthum in Different Habitats
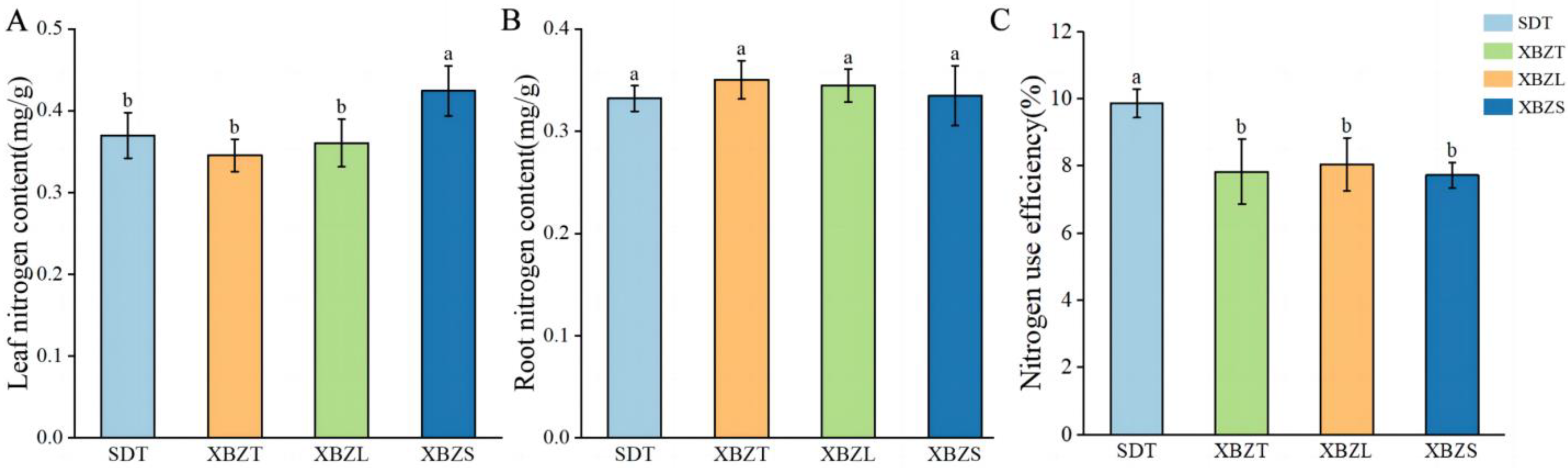
The nitrogen content across different habitats of P. micranthum was determined (Figure 2). Leaf nitrogen content ranged from 0.37 mg/g to 0.424 mg/g, with XBZS exhibiting significantly higher leaf nitrogen content (0.424 mg/g) compared to the other three habitats (p < 0.05). Root nitrogen content ranged from 0.33 mg/g to 0.35 mg/g, and there were no significant differences observed among the four habitats (p > 0.05). Comparison of leaf nitrogen and root nitrogen revealed varying trends. The nitrogen-use efficiency of P. micranthum ranged from 7.73% to 9.87%. Nitrogen-use efficiency among the four plot types was compared (Figure 2), showing that the SDT habitat had significantly higher nitrogen-use efficiency than the other three habitats (p < 0.05). However, there were no significant differences in nitrogen-use efficiency observed among same regions within the different habitats.
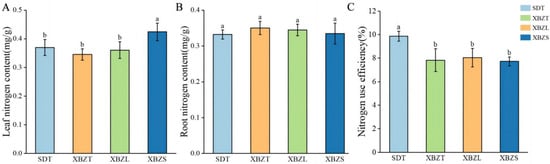
Figure 2.
Nitrogen content and nitrogen-use efficiency of P. micranthum in different habitats: (A) leaf nitrogen content; (B) root nitrogen content; (C) nitrogen-use efficiency. Different letters indicate a significant difference at p < 0.05.
3.3. Analysis of the Illumina Sequencing Data
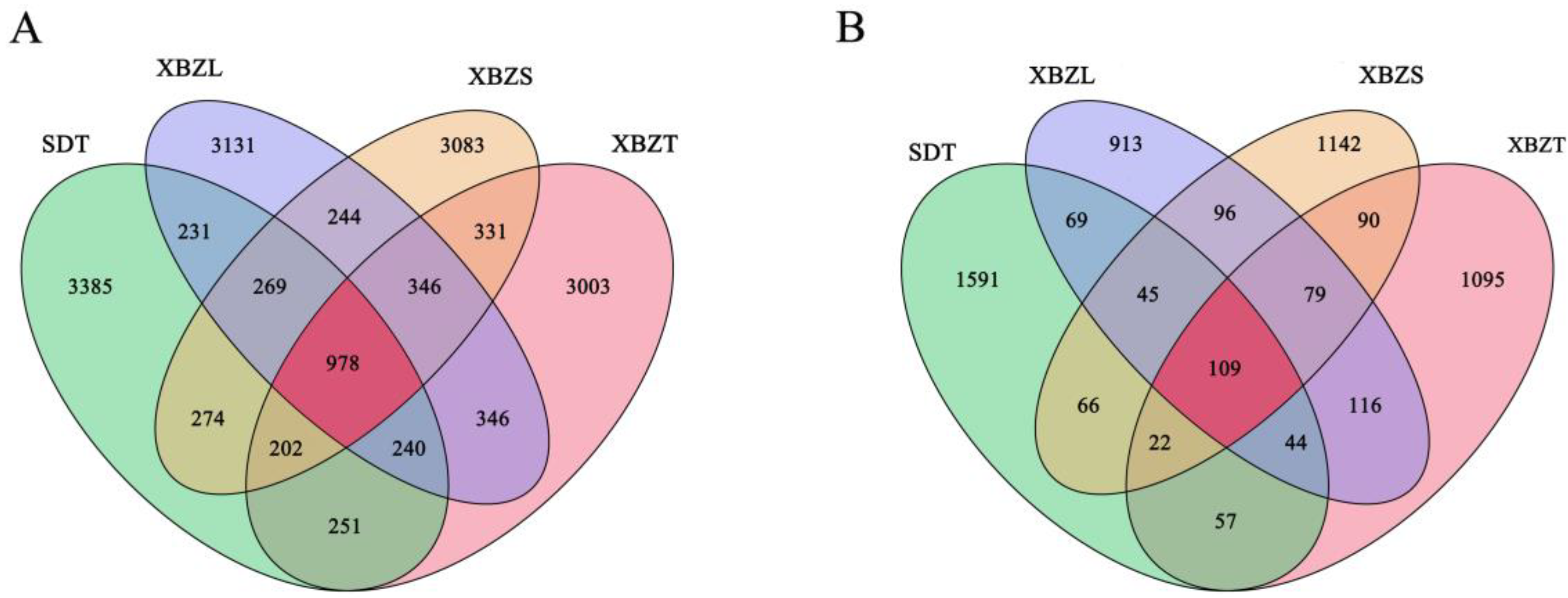
In this study, soil samples underwent Illumina NovaSeq high-throughput sequencing, resulting in a total of 2,005,967 bacterial and 1,828,241 fungal raw reads. Following the filtering of low-quality sequences, denoising, merging, and removal of chimera reads, 721,180 clean tags for bacteria and 151,669 for fungi were obtained. Valid sequences with 100% similarity in the soil samples were clustered into features. Venn diagrams were generated to visually illustrate the number of common and unique ASVs in soil samples from each habitat. These tags were classified into 16,314 bacterial and 5534 fungal ASVs. Among bacteria, the total number of bacterial features in the four groups was 3385, 3003, 3131, and 3083, respectively, encompassing a total of 978 ASVs, which accounted for 5.99% of the total number of classified individuals as ASVs (Figure 3A). For fungi, the effective fungal ASVs in the four groups were 2003, 1612, 1471, and 1649, totaling 109 ASVs, which accounted for 1.97% of the total. Meanwhile, the number of endemic ASVs for each habitat was 1591, 1095, 913 and 1142, respectively, making up 28.75%, 19.77%, 16.5%, and 20.64% of the total (Figure 3B). In summary, there was a higher proportion of shared ASVs among bacteria (5.99%) compared to fungi (1.97%) across all samples.
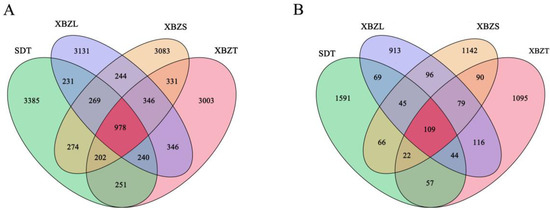
Figure 3.
Venn diagram of microbial communities in the rhizosphere soil of P. micranthum at different sites. Different colors represent distinctive groups. (A) Bacterial ASVs in soil from four habitats; (B) Fungal ASVs in soil from four habitats.
3.4. Diversity of Soil Bacterial and Fungal Communities in Different Habitats
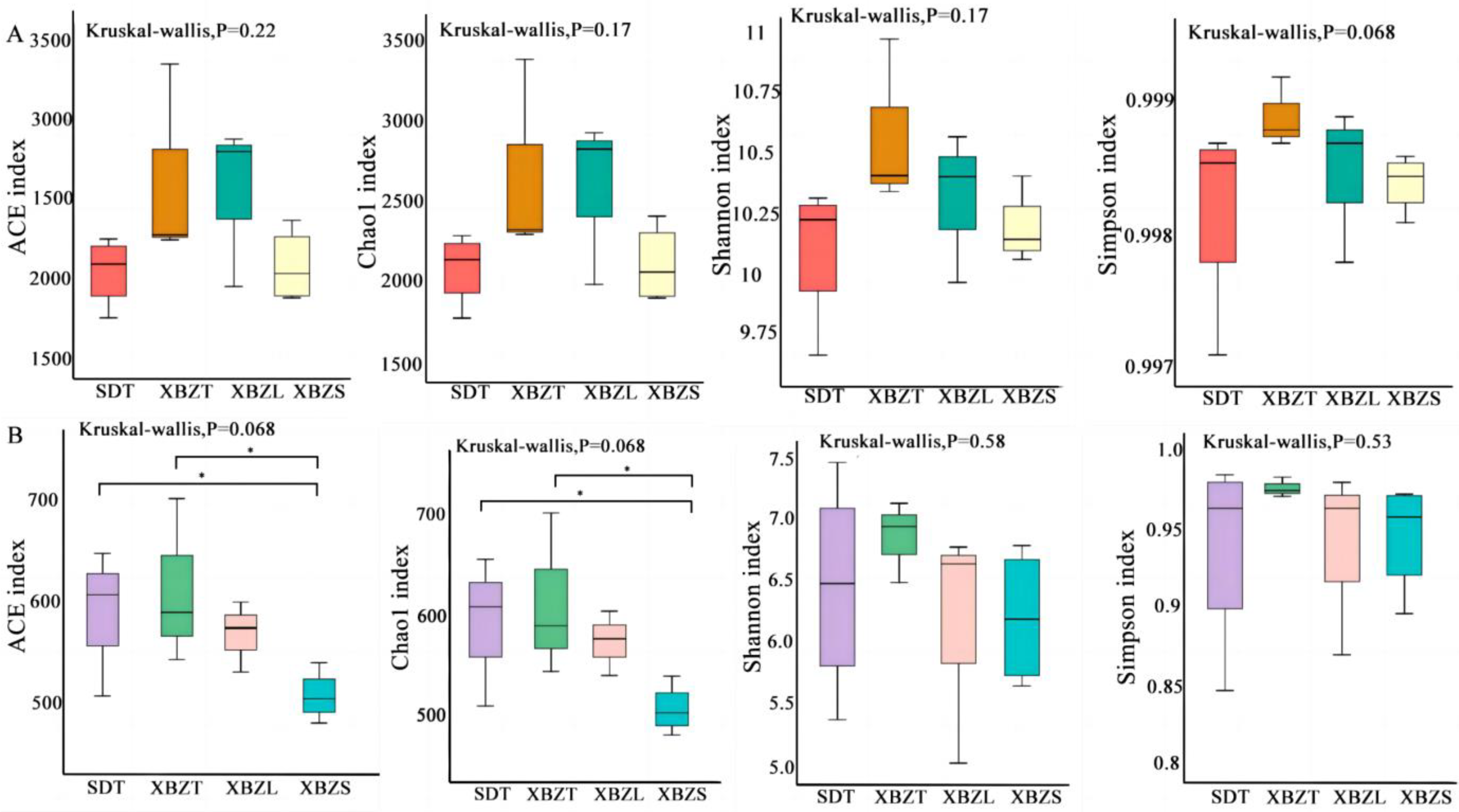
To understand the variation in microbial community composition among different habitats of P. micranthum, the rhizosphere soil microbial community was analyzed for α diversity indices, including ACE, Chao1, Shannon, and Simpson indices. According to KW tests of ACE and Chao1 indices between different regions (SD and XBZ), bacterial community richness was higher in XBZ compared to SD, although the difference was not statistically significant. Similarly, there were no significant differences observed in Shannon and Simpson indices for both bacterial and fungal community diversities between these regions (Figure 4). Adversely, the ACE index indicated significantly higher fungal community diversity in SDT compared to XBZS. Likewise, the Chao1 index showed a similar trend.
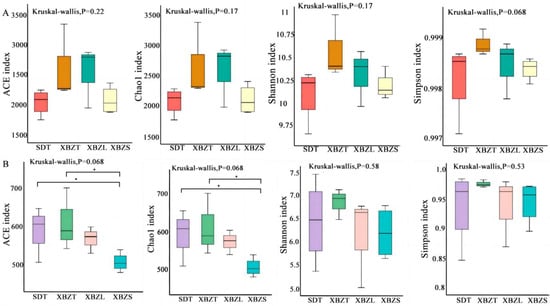
Figure 4.
Alpha diversity of soil fungi in different habitats of species in P. micranthum, including community richness (Chao, ACE indexes) and diversity (Shannon, Simpson indexes). KW was performed for factors (* p < 0.05) (A) the Alpha diversity indices of bacterial soil microbial communities associated with P. micranthum. (B) the Alpha diversity indices of fungal soil microbial communities associated with P. micranthum.
Comparing the community richness of different habitats within the XBZ region, results from ACE, Chao1, Shannon, and Simpson indices indicated no significant difference in bacterial community richness among them(Figure 4A). However, the ACE and Chao1 indices for the fungal community were significantly higher in XBZT than in XBZS (Figure 4B). In summary, there were no significant differences in the α diversity of bacterial communities among the different soil samples. Conversely, among the various microbial environments studied, the fungal community richness was highest in the tree foot habitat.
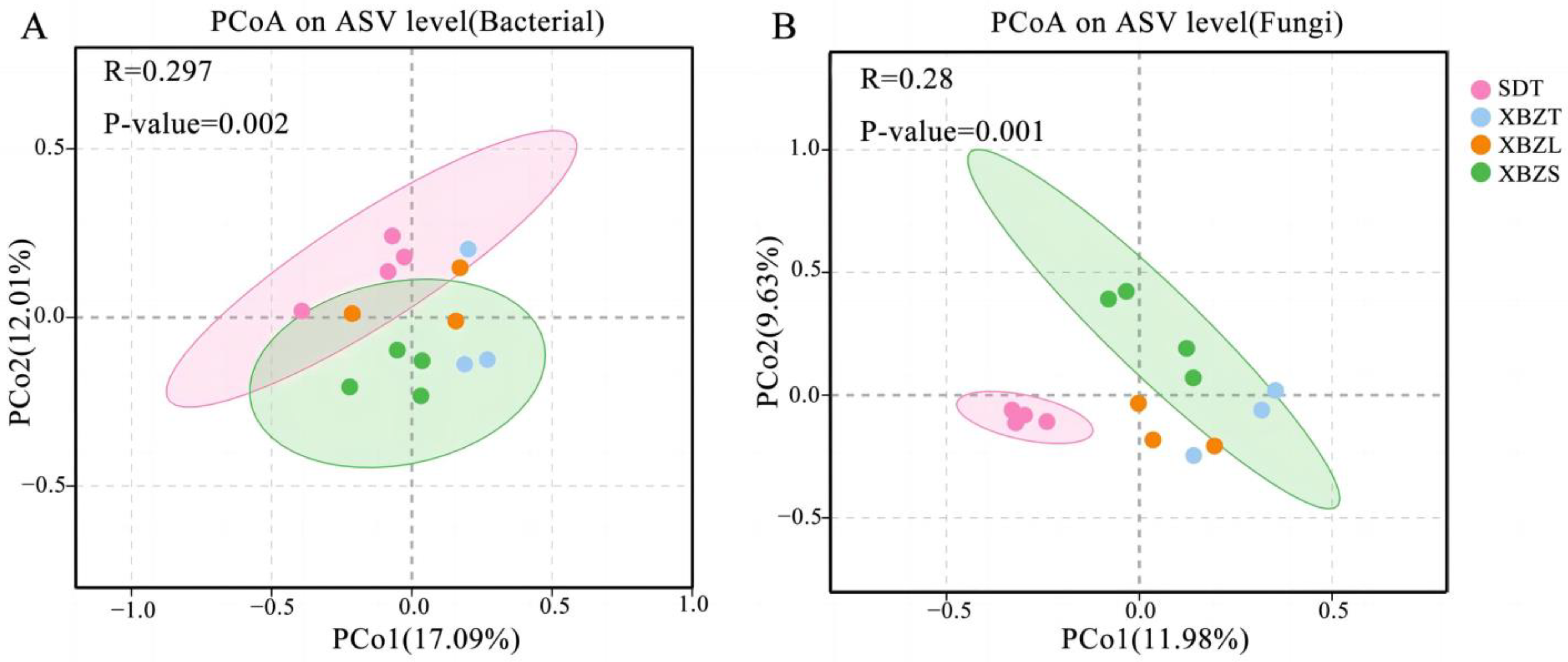
PCoA based on Bray Curtis distance revealed the dissimilarities in species composition among soil samples from different habitats of P. micranthum. For bacteria, PCoA of the 16S sequencing data showed that PCo1 and PCo2 contributed 17.09% and 12.01% to the total variance, respectively, explaining a combined 29.1% of the variation (Figure 5A). For fungi, PCoA of the ITS sequencing data indicated that PCo1 and PCo2 contributed 11.98% and 9.63% to the total variance, respectively, explaining a combined 21.61% of the variation (Figure 5B). The Anosim analysis indicated significant differences in microbial community composition between soil samples (Table S1). Bacterial and fungal community diversity showed significant differences (p < 0.05) between SDT and XBZT, XBZS.
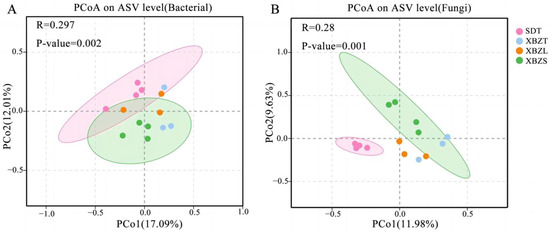
Figure 5.
Communities in the soil of P. micranthum at various habitats were analyzed using PCoA, based on the Bray–Curtis distance. (A) Principal coordinate analysis (PCoA) of bacterial; (B) Principal coordinate analysis (PCoA) of fungal.
3.5. Rhizosphere Soil Microbial Community Structure of Different Ecotypes of P. micranthum
3.5.1. Soil Microbial Community Composition
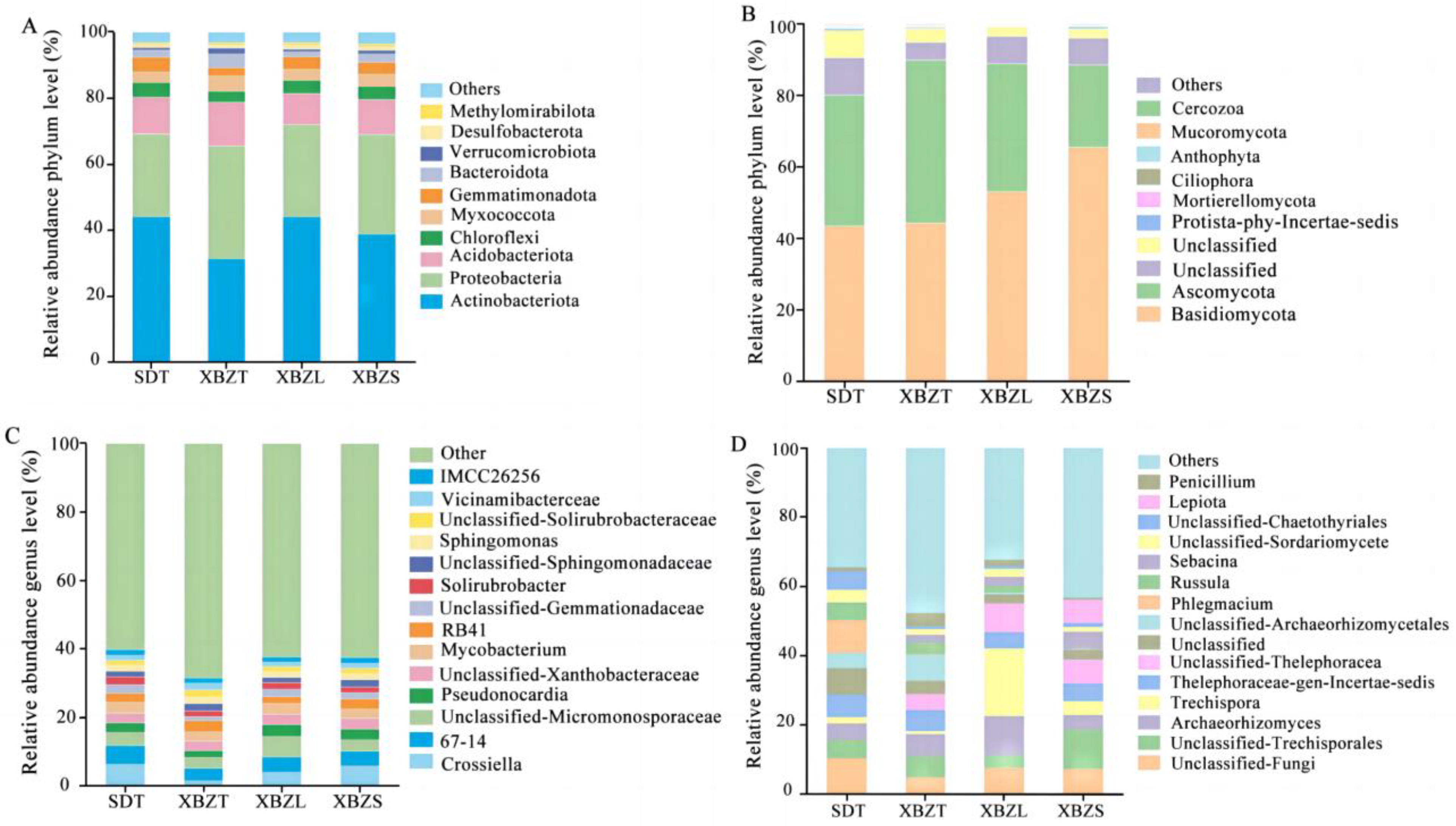
Based on 16S sequencing, the bacteria communities in P. micranthum belonged to 40 phyla, 112 classes, 257 orders, 418 families and 727 genera. The dominant bacterial phyla were Actinobacteriota (31.29–44.18%), Proteobacteria (24.91–34.27%), and Acidobacteriota (9.46–13.13%) (Figure 6A), collectively constituting more than 70% of the bacterial communities in each soil sample. Actinobacteriota exhibited the highest relative abundance in SDT, while Proteobacteria showed the lowest relative abundance in this habitat. Common dominant bacterial genera across different habitats included Crossiella (1.4–6.2%), 67-14 (3.62–5.52%), Mycobacterium (2.56–3.20%), RB41 (2.71–3.38%) and Pseudonocardia (2.20–3.49%) (Figure 6C). Simultaneously, the main dominant bacterial taxa (from phylum to genus) associated with P. micranthum were identified. SDT, XBZL, and XBZS shared the same dominant genera, while Vicinamibacteraceae (2.02%) ranked prominently in relative abundance in XBZT (Table S2).
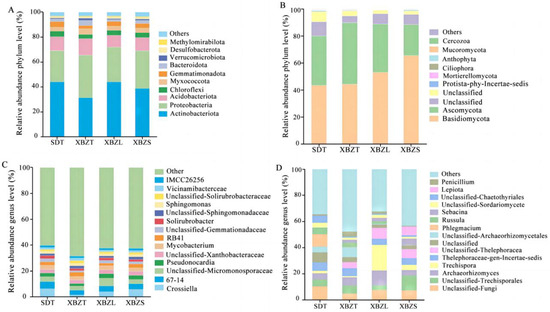
Figure 6.
Composition map of the main soil microbial groups at the phylum level and genus level in different habitats of P. micranthum. (A) Phylum-level composition of the bacterial community. (B) Phylum-level composition of the fungal community. (C) Genus-level composition of the bacterial community. (D) Genus-level composition of the fungal community.
Based on ITS sequencing, the fungi communities in P. micranthum belonged to 35 phyla, 82 classes, 181 orders, 367 families and 628 genera. Two taxonomic levels (phylum and genus) were selected for further analysis. At the phylum level, Basidiomycota and Ascomycota were the dominant fungal taxa in different habitats, collectively accounting for 86.87% of the relative abundance of fungal phyla across all soil samples (Figure 6B). However, the relative abundance varied among different habitats. XBZS in Basidiomycota had a significantly higher relative abundance than SDT. XBZT in Ascomycota had a significantly higher relative abundance than XBZS. At the genus level, the soils of P. micranthum habitats were rich in fungi. In summary, the dominant genera were Phlegmacium (9.67%), Archaeorhizomyces (6.40%), Trechispora (19.67%), and Lepiota (6.67%) in SDT, XBZT, XBZL, and XBZS, respectively(Figure 6D). However, there were also numerous undefined fungal groups and other fungi. Additionally, the primary dominant fungal communities within each taxonomic unit (from phylum to genus) of P. micranthum were identified (Table S3). Common dominant fungal genera included Thelephoraceae-gen-Incertae-sedis (4.63–7.63%) and Archaeorhizomics (4.12–11.35%).
3.5.2. Differential Analysis of Microbial Community Composition
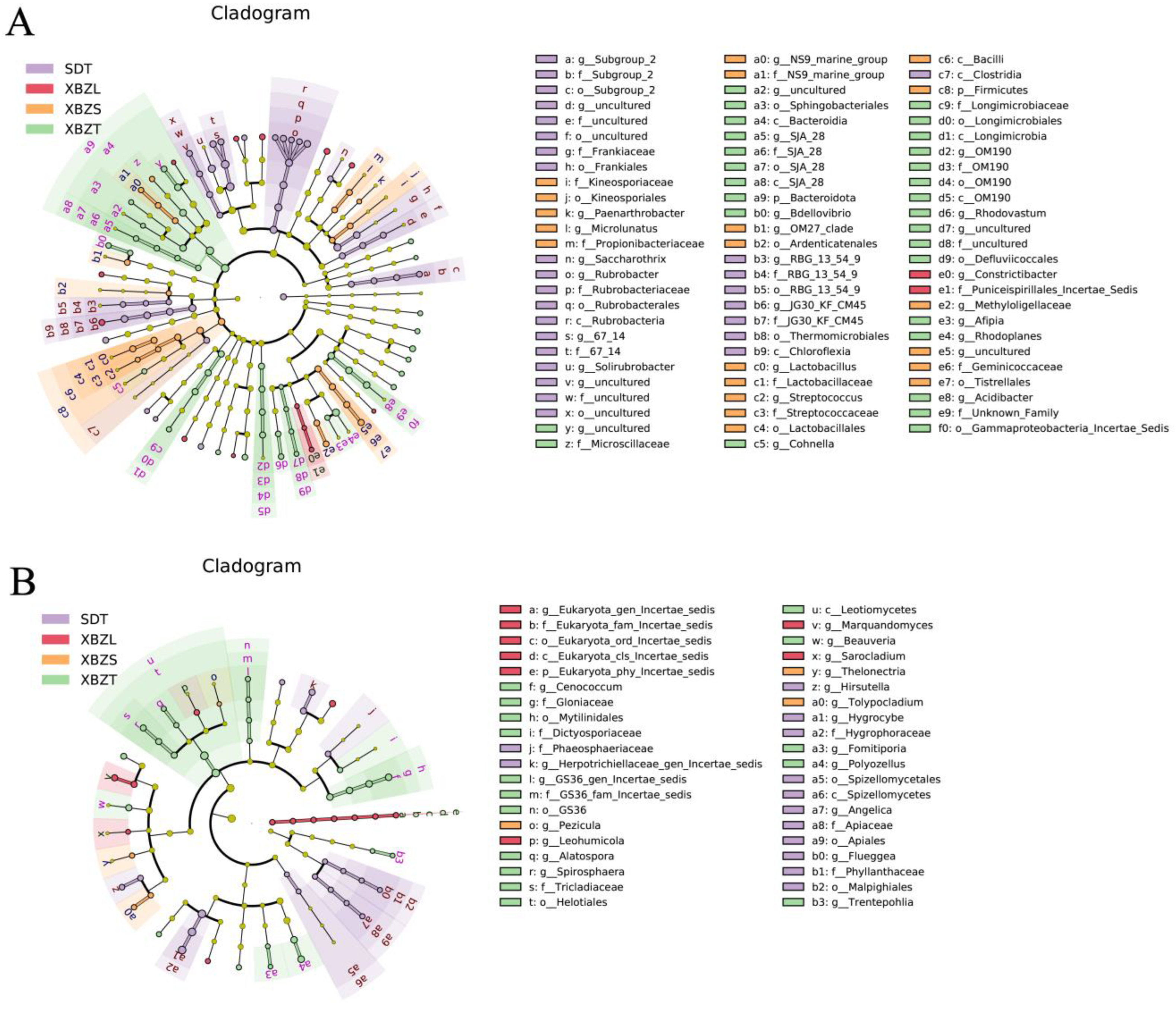
Setting the linear discriminant analysis (LDA) threshold to 2 revealed that the rhizosphere bacterial communities across different habitats exhibited variations in richness among significantly different taxa (Figure 7A). The lithophyte habitat showed the lowest diversity of differentially significant taxa, with Constrictibacter and others at the genus level. In SDT habitat, the differentially significant taxa included 67-14, Rubrobacter, Soilrubrobacter, Subgroup-2. In XBZT habitat, notable taxa were Rhodoplanes, Acidibacter, Rhodovastum, Bdellovibrio. Meanwhile, the XBZS habitat featured differentially significant taxa such as Microlunatus, Methyloligellaceae, OM27, and Streptococcus (Figure S2).
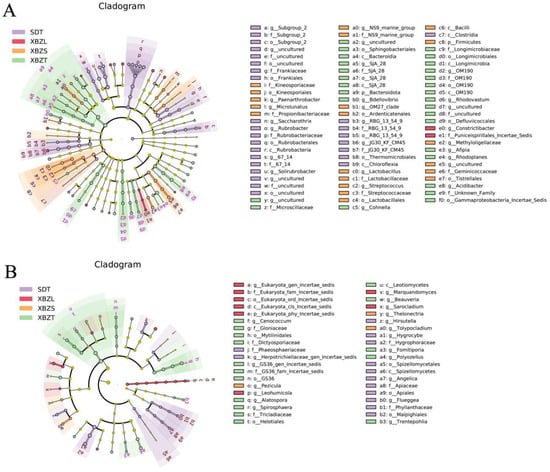
Figure 7.
LEfSe analysis of microbial abundance in four rhizosphere soil samples. Cladogram showing taxa with different abundance values of bacterial (A) and fungal (B) community. Taxa from various sites are depicted as colored dots.
Lefse analysis of the rhizosphere fungal community revealed distinct differentially significant fungal groups across various habitats (Figure 7B). The XBZT habitat exhibited the highest number of these groups at the genus level, including Cenococcum, Polyozellus, Beauveria, and Spirosphaera. In the SDT habitat, significant taxa included Hygrocybe, Herpotrichiella-gen-incertae-sedis, Hirsutella, Angelica. The XBZL habitat was characterized by significant taxa such as Marquandomyces, Leohumicola. In the XBZS habitat, the main taxa contributing to community differences included Tolypocladium, Thelonectria, Pezicula (Figure S3).
3.5.3. Soil Microbial Functional Prediction
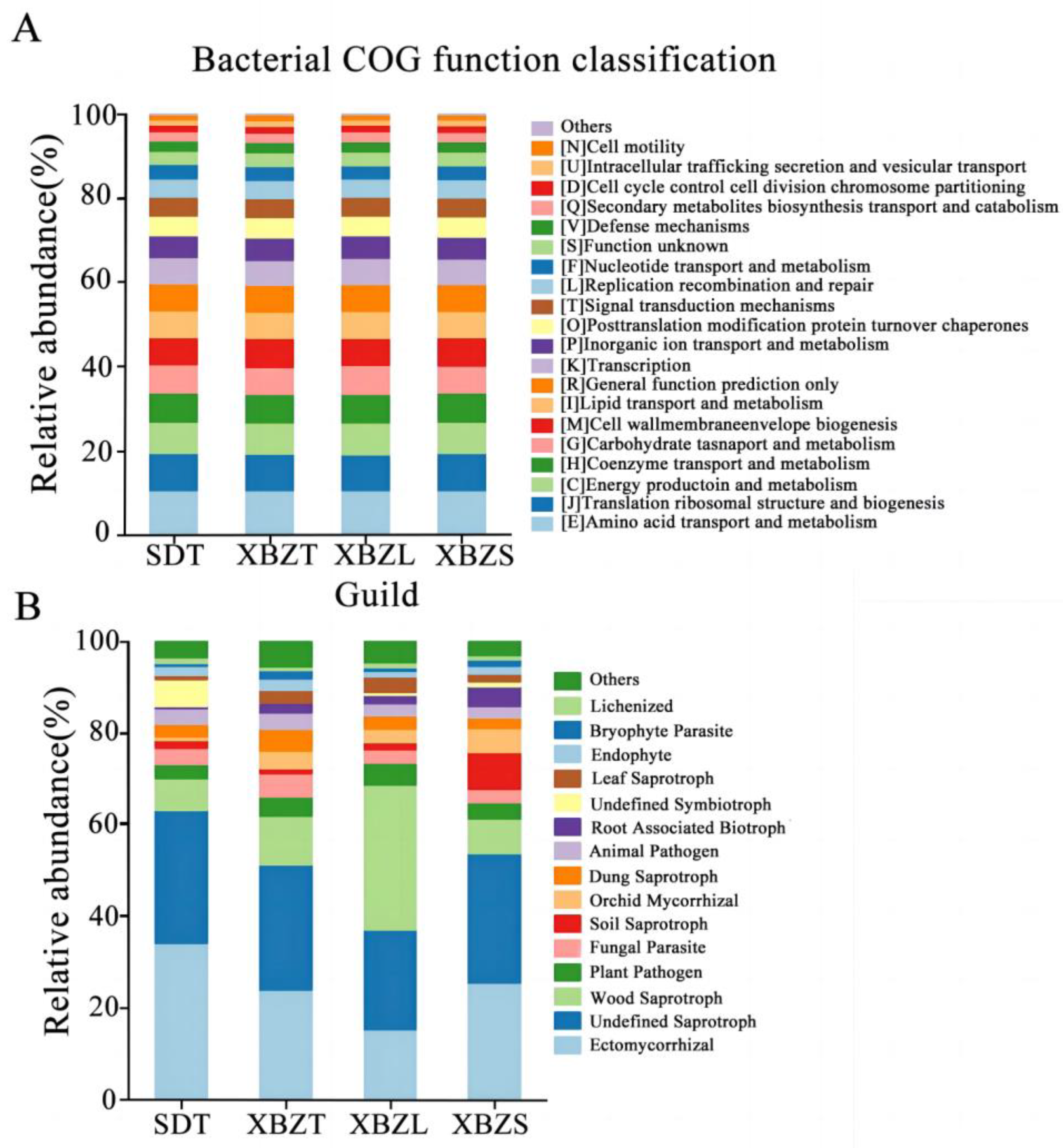
Among the identified taxa, functions related to bacterial metabolism were abundant, including amino acid transport and metabolism, energy production and conversion, and carbohydrate transport and metabolism (Figure 8A). This suggests that P. micranthum exerts selective action on the assembly of rhizosphere bacteria, favoring specific functional traits essential for its adaptation to heterogeneous environments.
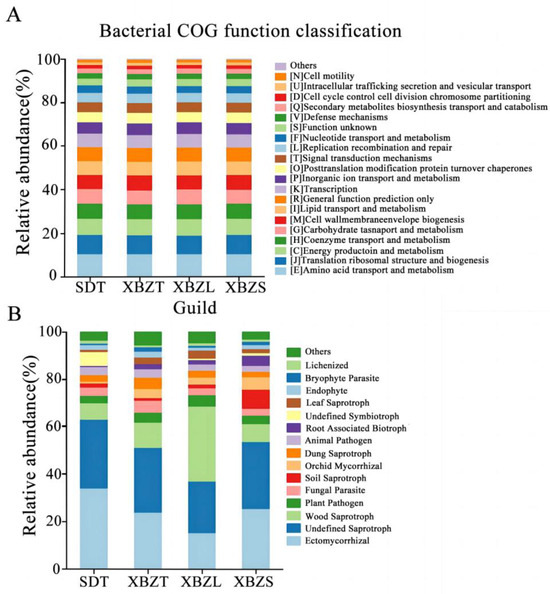
Figure 8.
Taxa composition of rhizosphere soil microbiomes with high relative abundance and their functional characteristics of P. micranthum at various habitats. (A) Functional features of the bacterial community and (B) functional features of the fungal community.
The functional groups of soil fungi in the different habitats of P. micranthum were similar and categorized by trophic type. The functional groups of soil fungi in among habitats of species were all symbiotroph, saprotroph and pathoproth fungi (Figure S1). Saprotrophic fungi were predominant in the rhizosphere soil samples of XBZ, comprising over 44.67% of the community. This group encompassed various types, including undefined saprotrophs, wood saprotrophs, and soil saprotrophs (Figure 8B). Symbiotic fungi dominated the rhizosphere soil samples in SD, comprising over 50.59% of the community. Ectomycorrhizal and Orchid Mycorrhizal fungi were relatively abundant among the symbiotrophs. Pathogens represented the smallest proportion, ranging from 8.83% to 14.96%. In summary, saprophytic and symbiotic fungi were the primary types observed in P. micranthum. This suggested that P. micranthum preferred saprotrophic nutrition and thrives in organic matter-rich habitats. Furthermore, it underscores the symbiotic relationships P. micranthum forms with other organisms.
3.6. Relationship Between Soil Microbial Community Structure and Soil Properties
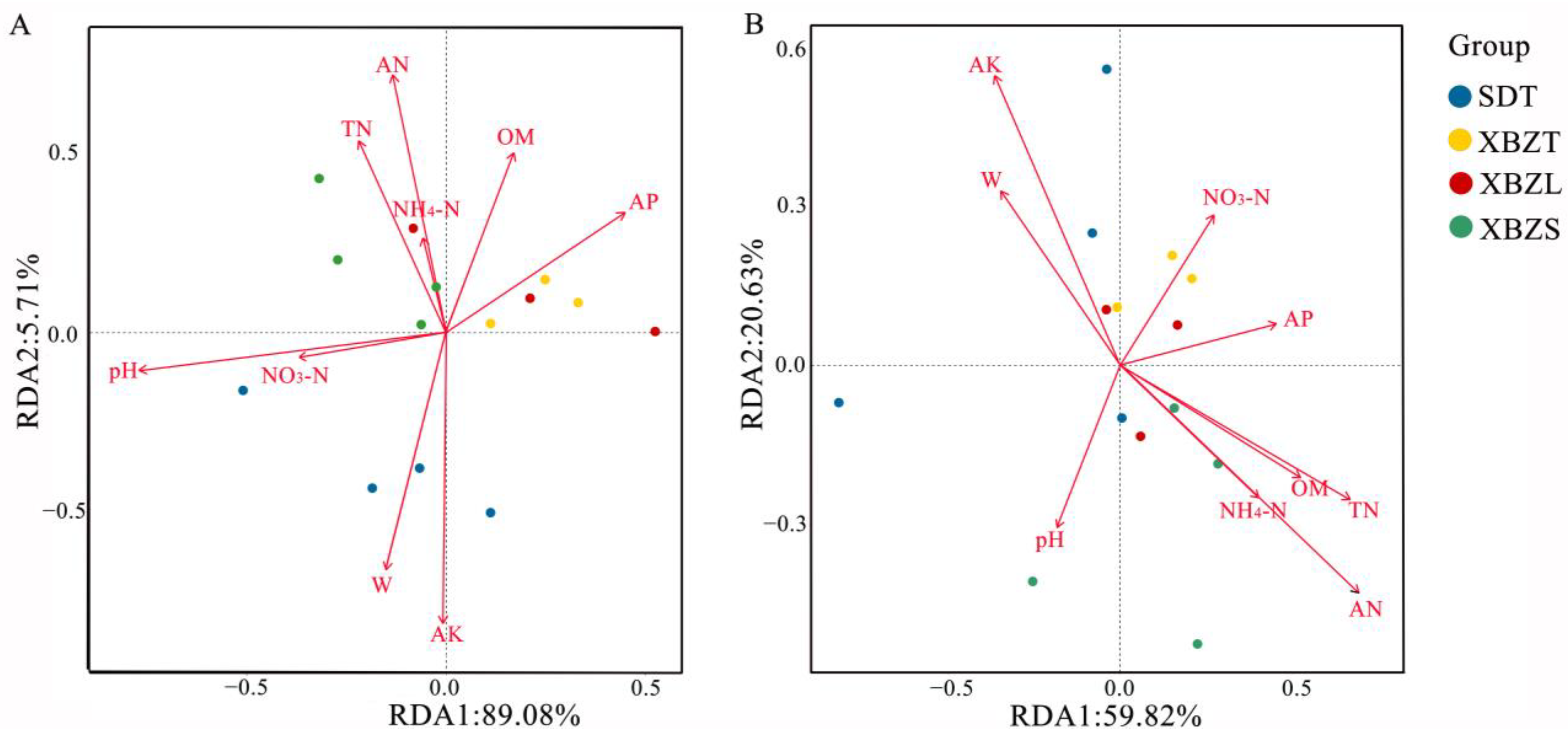
Soil physicochemical properties are important factors in the variation of microbial community structure. To delve deeper into the environmental factors influencing microbial diversity in P. micranthum soils, Redundancy Analysis (RDA) was employed to examine the relationship between soil physicochemical properties, samples, and flora. In the bacterial community, RDA analysis revealed that 94.78% of the total variation was explained, with RDA1 accounting for 89.07% and RDA2 for 5.71% (Figure 9A). Among them, pH, W, AN, and AK were identified as the primary soil properties influencing the structure of bacterial communities (r2 = 0.507, p = 0.018 for pH; r2 = 0.519, p = 0.022 for W; r2 = 0.604, p = 0.015 for AN; r2 = 0.752, p = 0.002 for AK). Similarly, in the fungal community, RDA analysis results explained 80.45% of the total variation, with RDA1 contributing 59.82% and RDA2 contributing 20.63% (Figure 9B). AN, AK, and TN were identified as the primary soil properties influencing fungal community structure (r2 = 0.603, p = 0.002 for AN; r2 = 0.376, p = 0.025 for AK; r2 = 0.461, p = 0.032 for TN). Notably, AN and AK exerted significant effects on both bacterial and fungal community structures.
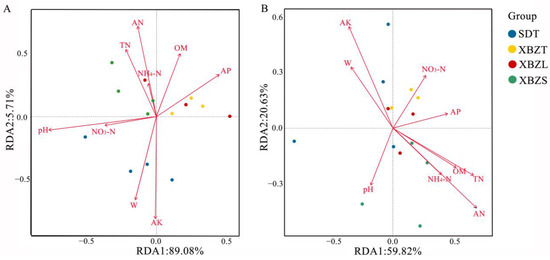
Figure 9.
Redundancy analysis (RDA) of soil properties and bacterial (A) or fungal (B) community in rhizosphere soil of P. micranthum.
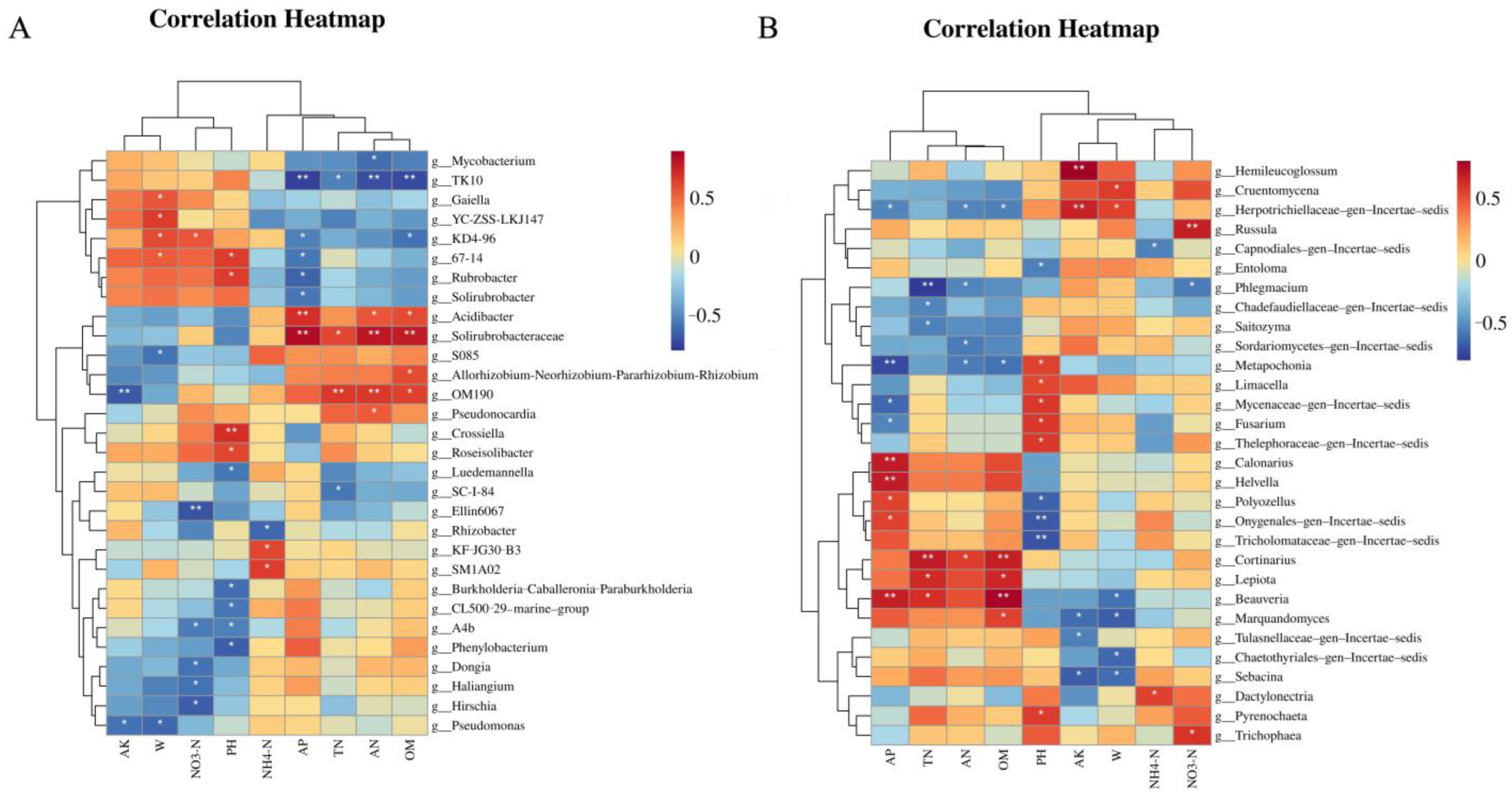
Spearman’s coefficient was used to analyze the relationships between soil physicochemical variables and the top 30 genera by abundance in the soil microbial community, represented in a heatmap plot. Within the bacterial community, five out of six highly relevant genera (Hirschia, Haliangium, Dongia, A4b, and Ellin6067) exhibited negative correlations with soil NO3−-N (p < 0.05). Additionally, M1A02 and KF-JG30-B3 showed positive correlations with soil NH4+-N (Figure 10A). A total of six genera were significantly associated with AN content, and four were linked to TN variation. Overall, these 16 genera may be related to nitrogen metabolism. Within the fungal community, six out of ten highly relevant genera exhibited positive correlations with soil pH (p < 0.05). Moreover, six genera were significantly related to TN content (Figure 10B), and five genera were associated with AN variation. Additionally, Russula and Trichophaea showed a significant positive correlation with NO3−-N, and Dactylonectria exhibited a significant positive correlation with NH4+-N. In conclusion, we hypothesize that 13 fungal genera in this study may be associated with nitrogen metabolism.
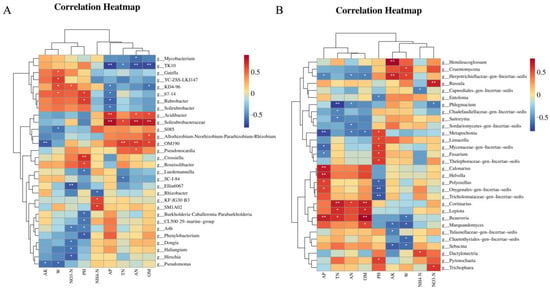
Figure 10.
Heatmaps showing correlations between soil factors and the first 30 genera of bacteria (A) and fungi (B). Color depth represents the magnitude of the correlation R value. * means significant correlation at the 0.05 level; ** means significant correlation at the 0.01 level.
4. Discussion
4.1. Nitrogen Preference of P. micranthum in Different Habitats
Plants can enhance nitrogen utilization efficiency through a complex and synergistic regulatory network under nitrogen-constrained conditions [35]. Majority tend to absorb inorganic nitrogen involving nitrate nitrogen and ammonium nitrogen [36]. However, plant nitrogen preference was closely associated with their physiological requirements and the dominant available nitrogen form in the soil [37,38]. Generally, it was hypothesized that the nitrogen form preferred by plants often aligns with the dominant nitrogen form present in the soil [38,39]. Many studies report that plants preferentially uptake ammonium nitrogen when growing in acidic soils where ammonium nitrogen was the dominant form, such as in high-altitude or high-humidity habitats [38]. Our findings indicate that ammonium nitrogen was the dominant form of inorganic nitrogen in P. micranthum soil. In the diverse habitats we studied, NH4+-N consistently exceeded NO3−-N in the rhizosphere soils of P. micranthum. However, there were no significant differences observed in the levels of either ammonium nitrogen or nitrate nitrogen across all habitats of P. micranthum. This preference was determined by the availability of energy, which influences the uptake patterns of NH4+-N and NO3−-N in plants. NH4+-N uptake relies on ion exchange, whereas NO3−-N uptake involves an active process [40]. Plants growing under high light intensity can acquire more photosynthetic energy to promote NO3−-N uptake.
Moreover, environmental factors influencing nitrogen fixation and transformation, such as pH, soil temperature, humidity, and light intensity, crucially govern the preference of trees for nitrogen forms [41,42,43]. P. micranthum consistently prefers higher elevations, steeper slopes, and north-facing aspects in secondary evergreen and deciduous broad-leaved forests or humid shrubs [6]. This implies that P. micranthum adaptively favors cold, semi-shaded, and moist environments, which help to maintain a relatively high ammonium nitrogen level. Cold temperatures promote the retention of ammonium nitrogen and reduce nitrate nitrogen denitrification in shallow soil layers due to reduced ammonia volatilization and denitrification [42]. PH was the main factor associated with the bacterial and archaeal community structure to interfere nitrogen transformation in soil [44]. PH range of 6.5 to 6.8 was found to be unfavorable for the denitrification process. Previous studies proved that seedlings typically prefer NO3−-N in strongly acidic soils, but NH4+-N in neutral to alkaline soils [45]. Isotopic tracing will be employed to further investigate this assertion.
4.2. Effects of Different Habitats on the Rhizosphere Microbial Communities of P. micranthum
P. micranthum thrives in mixed secondary evergreen and deciduous broad-leaved forests or humid shrubs in karst mountains, primarily in limestone and humus soils [6,20]. Karst habitats are characterized by rich microbial diversity, high pH levels, and nitrogen scarcity [45]. This may be an important factor affecting the growth of P. micranthum. Currently, studies on the rhizosphere microbial community structure of P. micranthum have not been reported. This study provides preliminary insights into the relative abundance and dominant flora of rhizosphere microorganisms associated with P. micranthum in different habitats. Actinobacteriota, Proteobacteria and Acidobacteriota were the dominant bacterial phyla across all four habitats, while Basidiomycota and Ascomycota were the dominant fungal phyla, aligning with findings from Zhang et al. on rhizosphere soil microorganisms of Paphiopedilum [46]. Among the common dominant bacteria genera, Mycobacterium and Pseudonocardia have been documented in other tropical orchids, which can promote the growth of orchids and produce IAA in vitro [47,48]. The genus 67-14 has also been reported in P. helenae, P. emersonii, P. spicerianum [46]. Among the dominant fungal genera, Archaeorhizomyces has been found in the rhizosphere of Holopogon pekinensis. However, the biological functions of these dominant genera for P. micranthum require further investigation. Additionally, the presence of other dominant bacterial genera, such as Crossiella and RB41, along with fungal genera like Thelephoraceae-gen-Incertae-sedis, and Lepiota in the rhizosphere of P. micranthum, has not been previously reported in orchids. We speculated that these genera may play key roles in the rhizosphere soil of P. micranthum. Furthermore, significant numbers of unclassified fungi were identified in rhizosphere soil, suggesting that these mycorrhizal resources warrant further exploration to uncover the ecological characteristics underlying the intermittent distribution of P. micranthum.
In soil, there were important interactions between plant rhizosphere and microbial communities. Plants supply essential nutrients for rhizosphere microorganisms through root exudates and shape rhizosphere microbial community [49]. Microorganisms also influence plant growth and development through various complex mechanisms [50]. In this study, no significant differences were observed in the number of ASVs of rhizosphere bacteria and fungi in four different habitats, nor in the α diversity of bacterial communities and the Shannon and Simpson indices for fungi. Additionally, the dominant bacterial and fungal genera in rhizosphere soil showed considerable overlap across different habitats, potentially indicating that P. micranthum influences its associated microbial communities. However, Lefse analysis revealed significant differences in the types of both bacterial and fungal communities across habitats. RDA analysis also showed that soil pH, W, AN, and AK significantly affected the bacterial community structure, while AN, AK, and TN had a significant impact on fungal community structure. The composition, abundance, and activity of soil microbial communities vary with the microenvironment for different vegetation types within the same environment. In this study, there were differences in vegetation type and soil type between different habitats of P. micranthum. It was also confirmed that habitat variations would significantly affect the structure of plant microbial communities [51].
4.3. Soil Adaptation of P. micranthum and the Function of Soil Microbial Communities
Analysis of physicochemical properties across different habitats of P. micranthum revealed a preference for environments with high organic matter content and abundant nitrogen. High organic matter levels could enhance nitrogen conversion and increase the activity of microorganisms involved in this process. Additionally, variations in microbial flora across different habitats contribute to differences in nitrogen-use efficiency among plants.
Nitrogen (N) is one of the major elements essential for plant growth [52,53] and is often the most limiting nutrient for plants [54]. Ammonification and nitrification of soil nitrogen were key processes in the nitrogen cycle and plant nutrient uptake, which were mainly mediated by soil microorganisms [55,56]. At the same time, soil microorganisms can mineralize organic nitrogen into plant-available inorganic nitrogen, thereby influencing the soil’s capacity to supply nitrogen to plants [57]. Additionally, they retain a portion of nitrogen to form a nitrogen pool through assimilation [58]. Actinobacteriota, Proteobacteria, and Acidobacteriota were the dominant phyla in the four habitats, exhibiting strong nitrogen fixation capabilities, nutrient decomposition and contribution to plant productivity through their roles in the nitrogen cycle [54,59,60]. The common dominant bacterial genera across these heterogeneous habitats included Crossiella, 67-14, Mycobacterium, RB41, and Pseudonocardia. Crossiella is an aerobic actinomycetes and a nitrate reducing bacterium [61]. The genus 67-14 is typically found in environments with limited total organic carbon (TOC) and is primarily associated with autotrophic carbon dioxide fixation [62,63]. RB41 belongs to a group of rhizosphere bacteria that generally enhance nitrogen uptake by host plants [64]. Also, in this study, five bacterial genera were found to be negatively correlated with nitrate nitrogen in relation to soil properties. This was similar to previous studies, Haliangium and Ellin6067 were involved in ammonia oxidation [65] and Haliangium was capable of nitrification-denitrification [66]. Basidiomycota and Ascomycota were the dominant fungal phyla, consistent with the dominant fungal phyla reported by Tian et al. in the habitats of P. armeniacum, P. wenshanense, and P. concolor, where they play essential ecological roles in material decomposition and nutrient cycling [20]. The dominant fungi genera Phlegmacium, Archaeorhizomyces and Trechispora existed widely in the rhizosphere of orchidaceae, and have also been observed in the root surface of Arundina graminifolia and Cymbidium ensifolium [67,68]. Meanwhile, Russula and Trichophaea showed a significant positive correlation with NO3−-N, while Dactylonectria exhibited a significant positive correlation with NH4+-N. Therefore, it was speculated that P. micranthum may overcome the limitations of low nitrogen and nutrient poor environment through these dominant genera, while promoting the uptake and utilization of nitrogen and other nutrient elements by the plant.
Orchids are adapted to ecological constraints, utilizing unique mechanisms to access limited resources and promote nutrient-use efficiency [35]. Nitrogen uptake and utilization efficiency in most plants are influenced by soil nitrogen availability and growth stages. Additionally, the growth environment significantly impacts nitrogen-use efficiency, which varies greatly under different soil types, nitrogen levels, and climatic conditions [69]. Nitrogen uptake and utilization can be affected by different soil nutrients as well as rhizosphere microbial species in the same plant in different habitats [70]. The results of the present study showed that heterogeneous habitats could have an effect on nitrogen-use efficiency. The nitrogen-use efficiency was observed to be lowest in the shrub habitat during this study. This may be linked to the composition of microorganisms in the shrub habitat. Previous studies have shown that due to the particularity of orchids, mycorrhizal fungi could provide considerable nitrogen nutrition to the plant at different stages of development [71,72]. Mycorrhiza makes the nitrogen concentration in orchid tissues higher than surrounding autotrophic plants [73]. In this paper, nitrogen content in the root leaves of P. micranthum was analyzed, revealing inconsistent trends in nitrogen levels across different samples. However, further research is needed to understand how microorganisms affect the nitrogen uptake and utilization efficiency of P. micranthum. The relationship between nitrogen-use efficiency and microbial community is also unclear.
5. Conclusions
In this paper, changes in the structure of the rhizosphere microbial community of P. micranthum were investigated across various habitats. There were no significant differences observed in microbial community richness and diversity across the heterogeneous habitats. The dominant bacterial phyla included Actinobacteriota, Proteobacteria and Acidobacteriota and the dominant genera were Crossiella, Pseudonocardia, 67-14, Mycobacterium and RB41. Among fungi, the predominant phyla were Basidiomycota and Ascomycota, with prominent genera such as Phlegmacium, Archaeorhizomyces, Trechispora, and Lepiota, which play essential roles in the rhizosphere of P. micranthum, particularly in regulating nitrogen transformation, uptake, and utilization. However, there were significant differences in taxa across habitats, such as Rubrobacter, Rhodoplanes, Microlunatus, Cenococcum, Hygrocybe, Marquandomyces, Tolypocladium. AN was identified as a primary driver of variation in soil bacterial and fungal communities. P. micranthum demonstrated a preference for habitats rich in ammonium nitrogen, which enhances nitrogen uptake. The highest nitrogen-use efficiency (9.78%) was observed in the terrestrial habitat of the Shedu population. Further research is recommended to explore the mechanisms underlying ammonium nitrogen uptake and metabolism in P. micranthum.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14111909/s1, Figure S1: Function prediction of fungi in different habitats according to nutrient classification; Figure S2: Histogram of LDA values distribution graph of significantly different rhizosphere bacteria biomarker for four habitats of P. micranthum. SDT-under tree foot in Shedu; XBZT-under tree foot in Xiaobazi; XBZL-lithophyte in Xiobazi; XBZS-under Shrub foot in Xiobazi; Figure S3. Histogram of LDA values distribution graph of significantly different rhizosphere fungal biomarker for four habitats of P. micranthum. SDT-under tree foot in Shedu; XBZT-under tree foot in Xiaobazi; XBZL-lithophyte in Xiobazi; XBZS-under Shrub foot in Xiobazi; Table S1: Anosim data analysis; Table S2: Bacterial dominant community in different habitats; Table S3: Fungi dominant community in different habitats.
Author Contributions
Conceptualization, Y.L. and Z.L.; methodology and investigation, Z.L.; data curation, Y.L., J.H. and Q.W.; writing—original draft preparation, Y.L., J.H. and Z.L.; writing—review and editing, Z.L., Y.L., Y.R. and Y.Y.; funding acquisition, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 32260414.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are presented in Tables and Figures within the main text or Supplemental Files.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, Z.J.; Chen, S.C.; Chen, L.J.; Lei, S.P. The Genus Paphiopedilum in China; Science Press: Beijing, China, 2009; pp. 4–12. [Google Scholar]
- CITES—Convention on International Trade in Endangered Species of Wild Fauna and Flora. Available online: https://cites.org/eng (accessed on 10 July 2023).
- Chen, S.C. Paphiopedilum. Flora Reipublicae Popularis Sinicae Tomus. Biodivers. Sci. 1999, 17, 52–72. [Google Scholar]
- Zhang, Y.; An, M.T.; Wu, J.Y.; Liu, F.; Wang, W. Geographical distribution pattern and dominant climatic factors of the Paphiopedilum Subgen. Brachypetalum in China. Chin. J. Plant Ecol. 2022, 46, 40. [Google Scholar]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Influencing factors on soil nutrients at different scales in a karst area. Caten 2019, 175, 411–420. [Google Scholar] [CrossRef]
- Li, Z.Y.; Li, J.; Li, M.Y. Effect of human disturbance on genetic structure of rare and endangered Paphiopedilum micranthum implied the habitat status. Trop. Conserv. Sci. 2020, 13, 1940082920942012. [Google Scholar] [CrossRef]
- Zotz, G.; Armenia, L.; Einzmann, H.J. A new approach to an old problem: How to categorize the habit of ferns and lycophytes. Ann. Bot. 2023, 132, 513–522. [Google Scholar] [CrossRef]
- Averyanov, L.V.; Hiep, N.T.; Lộc, P.K.; Averyanova, A.L. Preliminary orchid checklist of Cao Bang province (Vietnam). Lindleyana 2000, 15, 130–164. [Google Scholar]
- Toure, D.; Ge, J.W.; Zhou, J.W. Interactions between soil characteristics, environmental factors, and plant species abundance: A case study in the Karst Mountains of Longhushan Nature Reserve, southwest China. J. Mt. Sci. 2015, 12, 943–960. [Google Scholar] [CrossRef]
- Geekiyanage, N.; Goodale, U.M.; Cao, K.; Kitajima, K. Plant ecology of tropical and subtropical karst ecosystems. Biotropica 2019, 51, 626–640. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, H.C.; Cheng, Z.; Wang, M.N.; LI, L.Q.; Long, C.L. Current Status of Wild Orchid Resources in China, Focusing on Their Conservation and Utilization. China Biotechnol. 2022, 42, 59–72. [Google Scholar]
- Gaskett, A.C.; Gallagher, R.V. Orchid diversity: Spatial and climatic patterns from herbarium records. Ecol. Evol. 2018, 8, 11235–11245. [Google Scholar] [CrossRef]
- Mikryukov, V.; Dulya, O.; Zizka, A.; Bahram, M.; Hagh-Doust, N.; Anslan, S.; Prylutskyi, O.; Delgado-Baquerizo, M.; Maestre, F.T.; Nilsson, H.; et al. Connecting the multiple dimensions of global soil fungal diversity. Sci. Adv. 2023, 9, eadj8016. [Google Scholar] [CrossRef] [PubMed]
- Eiserhardt, W.L.; Svenning, J.C.; Kissling, W.D.; Balslev, H. Geographical ecology of the palms (Arecaceae): Determinants of diversity and distributions across spatial scales. Ann. Bot. 2011, 108, 1391–1416. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Peng, W.X.; Song, T.Q.; Zeng, F.P.; Wang, K.L.; Song, M.; Zhang, H. Spatial pattern of woody plants and their environmental interpretation in the karst forest of southwest China. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2015, 149, 121–130. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Jakovljević, K.; Stevanović, V. Patterns of distribution, abundance and composition of forest terrestrial orchids. Biodivers Conserv. 2020, 29, 4111–4134. [Google Scholar] [CrossRef]
- Ye, P.; Wu, J.; An, M.; Chen, H.; Zhao, X.; Jin, X.; Si, Q. Geographical distribution and relationship with environmental factors of Paphiopedilum subgenus Brachypetalum Hallier (Orchidaceae) taxa in southwest China. Diversity 2021, 13, 634. [Google Scholar] [CrossRef]
- Kaur, J.; Phillips, C.; Sharma, J. Host population size is linked to orchid mycorrhizal fungal communities in roots and soil, which are shaped by microenvironment. Mycorrhiza 2021, 31, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wu, S.; Yang, W.; Selosse, M.A.; Gao, J. How mycorrhizal associations influence orchid distribution and population dynamics. Front. Plant Sci. 2021, 12, 647114. [Google Scholar] [CrossRef]
- Tian, L.; An, M.; Wu, M.; Liu, F.; Zhang, Y. Habitat ecological characteristics and soil fungal community structure of Paphiopedilum subgenus Brachypetalum Hallier (Orchidaceae) plants in Southwest China. Plant Signal. Behav. 2023, 18, 2227365. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, Z.L.; Li, S.Y.; Hu, H.; Huang, J.L. Mycorrhizal specificity, preference, and plasticity of six slipper orchids from South Western China. Mycorrhiza 2010, 20, 559–568. [Google Scholar] [CrossRef]
- Karbarz, M.; Szlachcikowska, D.; Zapał, A.; Leśko, A. Unlocking the Genetic Identity of Endangered Paphiopedilum Orchids: A DNA Barcoding Approach. Genes 2024, 15, 689. [Google Scholar] [CrossRef]
- Deb, C.R.; Jakha, H.Y. Factors affecting asymbiotic immature seed culture and in vitro propagation of Paphiopedilum insigne (Wall. Ex. Lindl.) Pfitzer, a horticultural important vulnerable orchid. Plant Cell Biotechnol. Mol. Biol. 2020, 21, 129–141. [Google Scholar]
- McPherson, M.R.; Wang, P.; Marsh, E.L.; Mitchell, R.B.; Schachtman, D.P. Isolation and analysis of microbial communities in soil, rhizosphere, and roots in perennial grass experiments. JoVE 2018, 137, e57932. [Google Scholar]
- Wang, B.; Chen, H.; Qu, P.; Lin, R.; He, S.; Li, W.; Zhang, C.; Shi, X.; Liu, Y.; Du, H.; et al. Effect of Different Cultivation Patterns on Amomum villosum Yield and Quality Parameters, Rhizosphere Soil Properties, and Rhizosphere Soil Microbes. Horticulturae 2023, 9, 306. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, G.; Ren, S.; Li, L.; Li, C.; Li, Y.; Yu, X.; Yin, Y.; Liu, T.; Liu, X. Responses of soil microbial community structure, potential ecological functions, and soil physicochemical properties to different cultivation patterns in cucumber. Geoderma 2023, 429, 116237. [Google Scholar] [CrossRef]
- Bao, S.D. Agrochemical Analysis of Soil; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer Science and Business Media: New York, NY, USA, 2012. [Google Scholar]
- Zhang, M.; Wang, N.; Zhang, J.; Hu, Y.; Cai, D.; Guo, J.; Wu, D.; Sun, G. Soil physicochemical properties and the rhizosphere soil fungal community in a mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Forests 2019, 10, 167. [Google Scholar] [CrossRef]
- Huang, M.; Wang, Z.; Luo, L.; Wang, S.; Hui, X.; He, G.; Cao, H.; Ma, X.; Huang, T.; Zhao, Y.; et al. Soil testing at harvest to enhance productivity and reduce nitrate residues in dryland wheat production. Field Crops Res. 2017, 212, 153–164. [Google Scholar] [CrossRef]
- Lü, W.X.; Ge, Y.; Wu, J.Z.; Chang, J. Study on the method for the determination of nitric nitrogen, ammoniacal nitrogen and total nitrogen in plant. Guang Pu Xue Yu Guang Pu Fen Xi 2004, 24, 204–206. [Google Scholar]
- Lee, Y.J.; Sung, J.K.; Lee, S.B.; Lim, J.E.; Song, Y.S.; Lee, D.B.; Hong, S.Y. Plant analysis methods for evaluating mineral nutrient. Korean J. Soil Sci. Fertil. 2017, 50, 93–99. [Google Scholar] [CrossRef]
- Quan, Z.; Zhang, X.; Fang, Y.; Davidson, E.A. Different quantification approaches for nitrogen use efficiency lead to divergent estimates with varying advantages. Nat. Food 2021, 2, 241–245. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- De, L.C.; Biswas, S.S. Adaptational Mechanisms of Epiphytic Orchids: A Review. Int. J. Bio-Resour. Stress Manag. 2022, 13, 1312–1322. [Google Scholar] [CrossRef]
- Ashton, I.W.; Miller, A.E.; Bowman, W.D.; Suding, K.N. Niche complementarity due to plasticity in resource use: Plant partitioning of chemical N forms. Ecology 2010, 91, 3252–3260. [Google Scholar] [CrossRef]
- Warren, C.R. Does nitrogen concentration affect relative uptake rates of nitrate, ammonium, and glycine? J. Plant. Nutr. Soil. Sci. 2009, 172, 224–229. [Google Scholar] [CrossRef]
- Sun, S.; Chen, J.; Feng, W.; Zhang, C.; Huang, K.; Guan, M.; Feng, Y. Plant strategies for nitrogen acquisition and their effects on exotic plant invasions. Biodivers Sci. 2021, 29, 72. [Google Scholar]
- Liu, Q.; Wu, X.; Xing, H.; Chi, K.; Wang, W.; Song, L.; Xing, X. Orchid diversity and distribution pattern in karst forests in eastern Yunnan Province, China. For. Ecosyst. 2023, 10, 100117. [Google Scholar] [CrossRef]
- Li, S.X.; Wang, Z.H.; Stewart, B.A. Responses of crop plants to ammonium and nitrate N. Adv. Agron. 2013, 118, 205–397. [Google Scholar]
- Bartelheimer, M.; Poschlod, P. The response of grassland species to nitrate versus ammonium coincides with their pH optima. J. Veg. Sci. 2014, 25, 760–770. [Google Scholar] [CrossRef]
- Boczulak, S.A.; Hawkins, B.J.; Roy, R. Temperature effects on nitrogen form uptake by seedling roots of three contrasting conifers. Tree Physiol. 2014, 34, 513–523. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, H.; Xu, X. Root nitrogen acquisition strategy of trees and understory species in a subtropical pine plantation in southern China. Eur. J. Forest. Res. 2020, 139, 791–804. [Google Scholar] [CrossRef]
- Santillán, J.; López-Martínez, R.; Aguilar-Rangel, E.J.; Hernández-García, K.; Vásquez-Murrieta, M.S.; Cram, S.; Alcántara-Hernández, R.J. Microbial diversity and physicochemical characteristics of tropical karst soils in the northeastern Yucatan peninsula, Mexico. Appl. Soil Ecol. 2021, 165, 103969. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Shen, R.F.; Zhao, X.Q. Nitrogen source preference in maize at seedling stage is mainly dependent on growth medium pH. Agronomy 2022, 12, 2149. [Google Scholar] [CrossRef]
- Zhang, Z.L. Study on the Endangered Mechanism of Key Microorganisms in the Root-Endophytic and Rhizosphere of Four Extremely Small Populations of Paphiopedilum Plant Populations; Henan University of Science and Technology: Luoyang, China, 2023. [Google Scholar]
- Saikia, J.; Mazumdar, R.; Thakur, D. Phylogenetic afliation of endophytic actinobacteria associated with selected orchid species and their role in growth promotion and suppression of phytopathogens. Front. Plant Sci. 2022, 13, 1058867. [Google Scholar] [CrossRef] [PubMed]
- Tedsree, N.; Likhitwitayawuid, K.; Sritularak, B.; Tanasupawat, S. Diversity and Antimicrobial activity of plant growth promoting endophytic Actinomycetes isolated from Thai orchids. Environ. Nat. Resour. J. 2022, 20, 379–392. [Google Scholar] [CrossRef]
- Li, Y.Y.; Xu, T.T.; Ai, Z.; Wei, L.L.; Ma, F. Diversity and predictive functional of Caragana jubata bacterial community in rhizosphere and non-rhizosphere Soil at different altitudes. Environ. Sci. 2023, 44, 2304–2314. [Google Scholar]
- Yuan, R.W.; Liu, L.; Zhang, R.; Fan, S.Y. The interaction mechanism between plant rhizosphere secretion and soil microbe: A review. Chin. Agric. Sci. Bull. 2020, 36, 26–35. [Google Scholar]
- Liu, L.; Zhu, K.; Wurzburger, N.; Zhang, J. Relationships between plant diversity and soil microbial diversity vary across taxonomic groups and spatial scales. Ecosphere 2020, 11, e02999. [Google Scholar] [CrossRef]
- Aanderud, Z.T.; Bledsoe, C.S. Preferences for 15N-ammonium, 15N-nitrate, and 15N-glycine differ among dominant exotic and subordinate native grasses from a California Oak woodland. Environ. Exp Bot. 2009, 65, 205–209. [Google Scholar] [CrossRef]
- Wekesa, C.; Kiprotich, K.; Okoth, P.; Asudi, G.O.; Muoma, J.O.; Furch, A.C.U.; Oelmüller, R. Molecular characterization of indigenous rhizobia from Kenyan soils nodulating with common beans. Int. J. Mol. Sci. 2023, 24, 9509. [Google Scholar] [CrossRef]
- Xu, P.; Wang, E. Diversity and regulation of symbiotic nitrogen fixation in plants. Curr. Biol. 2023, 33, 543–559. [Google Scholar] [CrossRef]
- Yi, X.M.; Yuan, J.; Zhu, Y.H.; Yi, X.J.; Zhao, Q.; Fang, K.K.; Cao, L.K. Comparison of the abundance and community structure of N-cycling bacteria in paddy rhizosphere soil under different rice cultivation patterns. Int. J. Mol. Sci. 2018, 19, 3772. [Google Scholar] [CrossRef]
- Bai, B.; Liu, W.; Qiu, X.; Zhang, J.; Zhang, J.; Bai, Y. The root microbiome: Community assembly and its contributions to plant fitness. J. Integr. Plant Biol. 2022, 64, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.R.; Shi, R.H. The effect of soil pretreatment on the mineralization of nitrogen derived from different forms. J. Nanjing Agric. Univ. 1991, 14, 54–58. [Google Scholar]
- Fan, Q.S. Effect of soil microbial biomass on nitrogen conservation. Soils 1987, 19, 46–49. [Google Scholar]
- Finn, D.R.; Ziv-El, M.; Van Haren, J.; Park, J.G.; del Aguila-Pasquel, J.; Urquiza-Muñoz, J.D.; Cadillo-Quiroz, H. Methanogens and methanotrophs show nutrient-dependent community assemblage patterns across tropical peatlands of the Pastaza-Maranon Basin, Peruvian Amazonia. Front. Microbiol. 2020, 11, 746. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent understanding of soil acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Portillo, M.C.; Alloza, R.; Gonzalez, J.M. Three different phototrophic microbial communities colonizing a single natural shelter containing prehistoric paintings. Sci. Total Environ. 2009, 407, 4876–4881. [Google Scholar] [CrossRef]
- Rodriguez, V.; Moskwa, L.M.; Oses, R.; Kühn, P.; Riveras-Muñoz, N.; Seguel, O.; Scholten, T.; Wagner, D. Impact of climate and slope aspects on the composition of soil bacterial communities involved in pedogenetic processes along the Chilean coastal cordillera. Microorganisms 2022, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Filippini, G.; Bugnot, A.B.; Johnston, E.L.; Ruszczyk, J.; Potts, J.; Scanes, P.; Ferguson, A.; Ostrowski, M.; Varkey, D.; Dafforn, K.A. Sediment bacterial communities associated with environmental factors in intermittently closed and Open Lakes and lagoons (ICOLLs). Sci. Total Environ. 2019, 693, 133462. [Google Scholar] [CrossRef]
- Fu, Y.; Kumar, A.; Chen, L.; Jiang, Y.; Ling, N.; Wang, R.; Pan, Q.; Singh, B.P.; Redmile-Gordon, M.; Luan, L.; et al. Rhizosphere microbiome modulated effects of biochar on ryegrass 15N uptake and rhizodeposited 13C allocation in soil. Plant Soil 2021, 463, 359–377. [Google Scholar] [CrossRef]
- Wang, L.; Qiu, S.; Guo, J.; Ge, S. Light irradiation enables rapid start-up of nitritation through suppressing nxrB gene expression and stimulating ammonia-oxidizing bacteria. Environ. Sci. Technol. 2021, 55, 13297–13305. [Google Scholar]
- Li, L.; Dong, Y.; Qian, G.; Hu, X.; Ye, L. Performance and microbial community analysis of bio- electrocoagulation on simultaneous nitrification and denitrification in submerged membrane bioreactor at limited dissolved oxygen. Bioresour. Technol. 2018, 258, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Xie, T.X.; Liu, J.F.; Lan, S.R.; Peng, D.H.; Zhang, Q.H. Community structure and biological function of the root symbiotic fungi of Arundina graminifolia. Mycosystema 2019, 38, 1631–1642. [Google Scholar]
- Zhou, J.; Xie, T.X.; Liu, J.F.; Lan, L.Y.; Xu, Y.F.; Liu, Z.Y.; Ai, Y.; Zhang, Q.H. Community structure and biological function of the root symbiotic fungi of wild Cymbidium ensifolium. Acta Microbiol. Sin. 2021, 07, 2136–2153. [Google Scholar]
- Ying, Y.; Jiang, W.; He, X.; Fiaz, S.; Ahmad, S.; Lei, X.; Wang, X. A review of nitrogen translocation and nitrogen-use efficiency. J. Plant Nutr. 2019, 42, 2624–2641. [Google Scholar]
- Mou, Z.-m.; Yan, N.; Li, S.-y.; Hu, H. Nitrogen require ments for vegetative growth, flowering, seed production, and ramet growth of Paphiopedilum armeniacum (Orchid). HortScience 2012, 47, 585–588. [Google Scholar] [CrossRef]
- Shan, T.T.; Zhou, L.S.; Li, B.; Chen, X.M.; Guo, S.X.; Wang, A.R.; Tian, L.X.; Liu, J.T. The plant growth-promoting fungus MF23 (Mycena sp.) increases production of Dendrobium officinale (Orchidaceae) by affecting nitrogen uptake and NH4 + assimilation. Front. Plant Sci. 2021, 12, 693561. [Google Scholar] [CrossRef]
- Zahn, F.E.; Lee, Y.I.; Gebauer, G. Fungal association and root morphology shift stepwise during ontogenesis of orchid Cremastra appendiculata towards autotrophic nutrition. AoB Plants 2022, 14, plac021. [Google Scholar] [CrossRef]
- Hynson, N.A.; Madsen, T.P.; Selosse, M.A.; Adam, I.K.; Ogura-Tsujita, Y.; Roy, M.; Gebauer, G. The physiological ecology of mycoheterotrophy. In Mycoheterotrophy: The Biology of Plants Living on Fungi; Springer Science & Business Media: New York, NY, USA, 2013; pp. 297–342. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).