Abstract
Sweet potato (Ipomoea batatas L. Lam.) is a major source of food in many parts of Ethiopia. In recent years, viral diseases have become the main threat to sweet potato production in Ethiopia. Previous virus survey studies carried out from 1986 to 2020 reported eight viruses infecting sweet potato in Ethiopia. Consequently, obtaining and multiplying virus-free planting materials have been difficult for farmers and commercial multipliers. This study was conducted to detect viruses infecting the five sweet potato varieties used as source plants and compare the virus elimination efficiency between meristem cultures from untreated and heat-treated mother plants and production of virus-free sweet-potato-planting materials. Seven common viruses were tested for, using grafting to Ipomoea setosa, enzyme-linked immunosorbent assay (ELISA) and reverse-transcription polymerase chain reaction (RT–PCR) before and after elimination procedures as screening and confirmatory methods. The sweet potato feathery mottle virus (SPFMV) elimination efficiencies of meristem cultures from untreated (grown at 25 ± 1 °C) and heat-treated (grown at 39 ± 1 °C) potted plants of sweet potato varieties were evaluated and compared. Sweet potato feathery mottle virus (SPFMV) was detected in 12 of the 15 source plants tested. Triple infections of SPFMV, sweet potato chlorotic stunt virus (SPCSV), and sweet potato virus C (SPVC) were detected in one of the fifteen plants. This study reports the detection of SPVC for the first time in sweet potato plants from Ethiopia. The cutting of meristems from heat-treated plants further increased the percentage of virus-free plantlets by ca 10% to ca 16%, depending on the plant variety. Elimination efficiency also seemed to vary among varieties: the greatest difference was observed for ‘Tola’, and the least difference was observed for ‘Guntute’. The present study provided protocols for detecting viruses and generating virus-free sweet-potato-planting materials in Ethiopia.
1. Introduction
Sweet potato (Ipomoea batatas L. Lam.) is expanding faster than any other major food crop in terms of area under production in sub-Saharan Africa [1]. The sweet potato is the second most important root crop grown by small-scale farmers in two regions of Ethiopia: the Southern Nation, Nationalities, and People’s Region (SNNPR) and the Oromia Region. Sweet potato is used as a source of food and feed in Ethiopia [2]. It is a key crop for combatting food insecurity and enhancing nutritional balance due to its high drought resistance and relatively high yield per unit area (14 tons/ha world average) [3]. Therefore, nongovernmental organizations (NGOs), including the International Potato Center (CIP) and the USAID Ethiopia project “Better Potato for a Better Life”, are promoting the distribution of orange-fleshed sweet potato (high in pro-vitamin A) to new areas in the northern part of the country [4]. Nevertheless, viral diseases are still the main factor limiting the production of this crop.
Viral diseases have become the main threat to sweet potato production in Ethiopia. To date, thirty sweet-potato-infecting viruses have been identified worldwide [5]. Among these viruses, only the following eight have been reported in Ethiopia during studies carried out from 1986 to 2020: sweet potato feathery mottle virus (SPFMV), sweet potato chlorotic stunt virus (SPCSV), sweet potato virus 2 (SPV-2), sweet potato virus G (SPVG), cucumber mosaic virus (CMV), sweet potato virus C (SPVC), sweet potato symptomless mastrevirus (SPSMV), and three different strains of badna viruses (A, B, C) [6,7,8,9,10]. SPFMV and SPCSV are the most frequently detected viruses, with high incidence levels in both farmers’ fields and research stations in the SNNPR of Ethiopia [7,8]. These viruses tend to cause single infections and mixed infections [7,8,10,11]. These viruses are still infecting and being distributed with planting materials because of the lack of a supply of virus-free materials in Ethiopia.
In Ethiopia, farmers often use their own planting materials that were saved from previous harvests and exchange them with their neighbors. There is no dependable certification scheme to verify that the planting material they use is free from viruses. In vegetatively propagated crops such as sweet potato, viruses accumulate, perpetuate, and disseminate over seasons using infected planting material. This is more concerning when farmers’ knowledge about viruses and insect-transmitting viruses is low [12]. Popular sweet potato varieties that are farmers-preferred and grown in southern Ethiopia are reportedly infected by viruses [7,11,12]. These factors could be the main reasons why viruses are distributed in the SNNPR of Ethiopia. The use of virus-infected planting materials not only reduces yield and quality [13] but also significantly contributes to the persistent and continued spread of viral diseases [14]. Furthermore, research institutions where virus infections were reported to be very common in their sweet potato collection [7] are the only sources of planting materials for commercial propagators. The propagators sell planting material to national and international NGOs and government organizations that distribute it to farmers in drought-prone areas. Therefore, there is an obvious need for a more rigorous certification scheme in Ethiopia, which should also encompass a scheme for detection and virus elimination to provide clean stock for propagation.
Virus detection and elimination are important practices for virus management. The detection of pathogens (including viruses) is key to their management, the prediction of disease occurrence, and the prevention of subsequent crop loss due to infection [15]. Therefore, the detection and elimination of viruses from farmers’ preferred high-yielding varieties is essential before it distributes to the grower. The application of efficient therapeutic methodologies to eliminate viruses and the rigorous screening of virus-resistant accessions are required for clonally propagated plants [16], and these methods determine the success of securing quality-propagating material. Therefore, the use of more than one technique, depending on the different properties of the virus, is essential for correct detection and characterization. Different therapeutic agents, in combination with meristem culture and/or shoot tip culture, have been used to eliminate viruses from many crops, including sweet potato [17,18,19]. Virus elimination by heat therapy is achieved by exposing plants to temperatures between 35 °C and 54 °C [19] for different time periods, depending on the plant genotype. Heat therapy is the oldest, most effective, and most commonly used virus elimination method [20]. However, combinations of one or more of these methods could further enhance the elimination efficiencies of the methods.
There are no institutions that generate disease-free sweet potato materials for distribution to growers in Ethiopia. Few researchers have attempted to clean viruses from a few cultivars of Ethiopian sweet potato using meristem tip culture and thermotherapy [9,10]. However, the cleaned stock was not maintained for further multiplication and distribution to farmers and multipliers. As Ethiopia is one of the sweet potato producers in Africa, this study was conducted to (i) detect the viruses infecting five sweet potato varieties used as sources plant in Ethiopia, (ii) compare the virus elimination efficiencies of meristem culture and heat therapy combined with meristem culture, and (iii) produce virus-free sweet-potato-planting materials.
2. Materials and Methods
2.1. Sampling, Plant Materials, and Greenhouse Conditions
This study was conducted during the years 2017 and 2018 at the Plant Cell Laboratory at the Norwegian University of Life Sciences (NMBU) and the Virus Laboratory at the Norwegian Institute of Bioeconomy Research (NIBIO in Ås, Norway).
Five high-yielding sweet potato varieties were used for this study (Table 1). Vines from three plants of each variety were collected from farmers’ fields, university research sites, and research institutions in Ethiopia. These vines were then established in an insect-proof screen net house at Hawassa University, Ethiopia.

Table 1.
Descriptions of the Ethiopian sweet potato varieties used in this study.
Vine cuttings from three plants of each variety (designated plants A, B, and C) were transported to Norway, established in quarantine rooms at NMBU, and grown at 25 °C, with a light intensity of 150 μmol.m−2.s−1 and an RH of 70%. These plants were used for the virus-testing and virus-elimination experiments.
2.2. Virus Testing
Three standard testing methods were used: enzyme-linked immunosorbent assay (ELISA), reverse-transcription polymerase chain reaction (RT–PCR), and graft inoculation using Ipomea setosa as the indicator plant.
2.2.1. ELISA
The ELISA was applied to leaf samples of initial sources/sweet potato plants (before graft indexing to I. setosa) and leaf samples of I. setosa grafts indexed to initial plants (after graft indexing to I. setosa) before virus elimination. In addition, leaf samples of I. setosa graft-inoculated with plantlets regenerated after virus elimination experiments were tested. This was conducted to determine the initial status of the plants prior to subjecting them to procedures for virus elimination and to evaluate whether the virus elimination procedures were effective.
The initial plants were inspected, and virus symptoms (if any) were recorded during the active growth stage of the plants for a period of up to three months. Five common sweet-potato-infecting viruses, SPFMV, SPMMV, SPCSV, SPV2 and CMV, were tested for using double antibody sandwich (DAS)–ELISA and triple antibody sandwich (TAS)–ELISA following the procedures in [21] with some modifications. The modifications were as follows: 0.5 g of fresh leaf sample (composite to make it representative of the test plant) and 4.5 mL of sample extraction buffer were added to extraction bags (BioReba, Switzerland). The sap was extracted using a homogenizer (semi-automated HOMEX 6, BioReba, Switzerland). Polyclonal antibodies (DAS–ELISA), SPFMV (RT-0898), SPMMV (RT-0900), SPV2 (RT-1036), and CMV (RT-0929) and TAS–ELISA, SPCSV (RT-1064–1064/1), as well as the respective conjugates and positive controls of the target viruses, were obtained from the Leibniz Institute DMSZ-German collection of microorganisms and cell culture (Braunschweig, Germany). The antibodies and conjugates were diluted as advised by the manufacturers. However, we used 100 μL of the antibody-coating solution in each microtiter plate instead of 200 μL. One hundred microliters of unpurified sample extracts were dispensed into microtiter plates. All buffers used were prepared based on the recommendations of [21]. All incubation and washing steps were carried out according to [21]. Positive and negative controls were added into two wells per plate to verify the results of the ELISA. The results were measured two hours after the addition and incubation of the colorless substrate solution made from P-nitrophenyl phosphate disodium (20 mg*.Ta; Sigma) commercial tablet dissolved in 40 mL of substrate buffer. The absorbance of the virus-infected samples was determined based on visual assessments of the yellow color that developed, and the absorbance (AV) was measured at 405 nm using a SpectraMax 190 absorbance microplate reader (Molecular Devices, San Jose, CA, USA). Samples that developed a yellow color and had a mean absorbance greater than twice the mean of the absorbance of the negative control sample were considered virus-infected samples.
2.2.2. RT–PCR
RT–PCR was used to verify the results obtained with ELISA and as a main testing method for SPVC and SPVG. Briefly, total RNA was extracted from 0.1 g fresh leaf samples. These samples were ground to a fine powder in liquid nitrogen using a mortar and pestle. RNA extraction was performed using TRIzol reagent (Invitrogen, Life Technologies) following the manufacturer’s procedures. Total RNA quantity and quality were determined using a Nanodrop spectrophotometer (Thermo Fisher Scientific, USA), Agilent 2100 Bioanalyzer (Agilent Technologies, USA), and 1% agarose gel electrophoresis.
First-strand cDNA was synthesized using 2.5 μg of total RNA template, random primer (Invitrogen), and Super Script™ II Reverse Transcriptase (Invitrogen), and all other reaction components and reaction conditions were selected according to the manufacturer’s recommendations (Invitrogen, California ©2010 Life Technologies).
PCR was carried out in a 25 μL volume, using first-strand cDNA as a template; virus-specific primers for SPVC, SPVG, and SPV2 [22]; SPFMV, SPCSV and SPMMV [23]; and TIF DNA polymerase (Invitrogen) were used following the procedures of the manufacturers and the appropriate controls. Amplification was performed under the following reaction conditions: 94 °C for 2 min, 35 cycles of 94 °C for 20 s, 52 °C (for SPVC, SPVG, SPMMV and SPCSV), or 53 °C (for SPFMV) for 20 s, and 72 °C for 30 s. A final elongation was carried out at 72 °C for 5 min.
The presence of SPVC from Ethiopia was confirmed using next-generation sequencing (NGS) as described in [11]. In brief, total RNA was extracted from 100 mg of leaf tissue using Trizol (Invitrogen, CA, USA), and the samples were purified using PureLink RNA Mini purification kits, and DNase was treated using the TURBO DNA-free kit (Invitrogen by thermofisher scientific) according to the manufacturer’s instructions. The quantity and purity of RNA samples were checked using a Nanodrop (NanoDropTechnologies, Wilmington, DE, USA). Furthermore, the RNA integrity was confirmed using Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The total RNA (3 µg) was sent to and sequenced at Fasteris genome-sequencing services center (Geneva, Switzerland), and the sequences’ data were analyzed using a Virus Detect, bioinformatic program [24].
2.2.3. Graft Indexing to I. setosa
Shoot tips of the initial sweet potato plants and in vitro plantlets regenerated after heat treatments were grafted onto the indicator plant I. setosa before and after virus elimination treatments, respectively. Previous studies [25] have shown that some viruses occur in sweet potato at very low titers, which might be below the detection threshold of ELISA. Therefore, all plants that initially tested negative by ELISA were grafted to I. setosa (a highly susceptible indictor host) to increase the virus concentration for a second round of testing. In all cases, graft indexing to I. setosa was carried out as follows: a three-week-old I. setosa was graft-inoculated with a 3 cm long shoot tip, with 3–4 internodes (side graft indexing to I. setosa) taken from test plants. Three I. setosa plants were grafted to the shoot tips of the test plants. Two shoot tips were grafted onto each I. setosa plant to maximize the amount of inoculum. The graft union was tightly wrapped with Para-film, covered with a plastic bag for one week, and cultivated under greenhouse conditions consisting of a day/night temperature of 21/19 °C, an RH of 70%, and a light intensity of 150 μmolm−2s−1. Grafted I. setosa plants were checked daily, and the presence of systemic symptoms was recorded for up to 45 days after graft indexing to I. setosa. However, I. setosa leaf samples were collected one month after graft inoculation and stored at −80 °C for virus testing via ELISA and RT–PCR, as described previously.
In vitro plantlets regenerated after virus elimination were also graft-inoculated into I. setosa as described above. Graft indexing to I. setosa was performed twice, at 5 months (after five subcultures) and 18 months (after nine subcultures) after meristem culture. In vitro plantlets were randomly selected, and shoot tips approximately 3 cm long were grafted at each screening time following a procedure described earlier.
2.3. Heat Therapy, Meristem Culture, and Plant Regeneration
Of the five sweet potato varieties tested in this study, four were infected with SPFMV only and were free of SPMMV, SPCSV, SPV2 and SPVC, SPVG, and CMV. Consequently, these four varieties were used in the virus elimination experiment. Heat-therapy treatment, followed by meristem tip culture, was applied according to the methods described for sweet potato [26] with some modifications. Six vine cuttings from each variety were potted and grown in a growth chamber at a daily temperature cycle of 25/18 °C under a 12/12 h photoperiod with a light intensity of 150 μmolm−2s−1 provided by cool, white, fluorescent tubes (Osram L 58 W/840 Lumilux). After four weeks, the potted plants were divided into two sets, each consisting of three potted plants of each variety. One set continued to grow under the same conditions and was used as a meristem donor for meristem culture alone (non-heat-treated control). The other set was transferred to a different growth room where it was subjected to heat therapy for 8 weeks at a daily temperature cycle of 39 ± 1 °C/25 ± 1 °C day/night, a 12/12 h photoperiod, and a light intensity of 150 μmol.m−2.s−1 provided by cool, white, fluorescent tubes (Osram L 58 W/840 Lumilux). Meristems were excised from both experimental conditions (non-heat-treated and heat-treated donor plants) and cultured in vitro on nutrient media optimized for sweet potato plants [26,27]. MS media [28] supplemented with vitamins, plant growth regulators (in mg/L; 0.3% benzylaminopurine, 0.03% 1-naphthaleneacetic acid, and 0.5% gibberellin A3), iron (48 mg/L), and 2% parts per million (ppm)) were used to initiate regeneration. The cultures were placed under light (18 h)/dark (8 h) conditions for one week at 25 ± 1 °C and then subjected to more intense light (28 μmol.m−2.s−1) provided by cool, white, fluorescent tubes (Osram L 58 W/840 Lumilux). After five weeks, the surviving meristems were counted and sub-cultured into test tubes containing fresh media of the same composition for further shoot initiation. After initiation, the plantlets were elongated and multiplied on plant-growth-regulator-free 1/2 strength basal MS media for subsequent virus indexing using combinations of the methods described in Section 2.2 of this paper. The virus-elimination efficiency of the meristem culture alone and combined heat therapy was compared based on the percentages of plantlets that were cured of infecting viruses by each method.
3. Results and Discussion
3.1. Graft Inoculation Drastically Improves the Detection of SPFMV by DAS–ELISA and RT–PCR
Among the 15 plant sources collected from the field and research stations and grown in the greenhouse, only one plant (variety ‘Guntute’, plant B) displayed typical virus-like symptoms. The plant presented general chlorosis, vein clearing, leaf distortion, mosaic, reduced leaf size, and stunting of the whole plant (Figure 1b). The remaining plants showed no visible symptoms (Figure 1c). The DAS–ELSA and TAS–ELISAs used to detect SPFMV, SPMMV, SPCSV, SPV2, and CMV revealed the presence of SPFMV and SPCSV in ‘Guntute’ plant B only, a plant that displayed typical virus-like symptoms. No other viruses we tested for were detected in the other 14 plants according to DAS–ELISA and TAS–ELISA (Table 2). The RT–PCR results confirmed the results obtained by ELISA (Table 1). However, there was a greater number of plants that were positive for SPFMV (6 out of 15), as detected by the RT–PCR (Table 2).

Figure 1.
Multiple infections of sweet potato plants. (a): PCR products of triple infection of virus in samples of the Guntute variety. Lane 1 is a 100 bp lader, lanes 2 and 3 are the positive control and PCR products of SPFMV (185 bp) detected in the sample, respectively; lanes 4 and 5 are the positive control and PCR products of SPCSV (200 bp), respectively; and lanes 6 and 7 are the positive control and PCR products of SPVC (836), respectively. (b) shows stunted plants infected with three viruses: SPFMV, SPCSV, and SPVC. (c): Symptomless plant infected with SPFMV.
Table 2.
Induced viral symptoms and viruses detected in orginal source sweet potato varieties from Ethiopia and I. setosa upon graft-inoculation with the source plant at 30 days post-inoculation.
Subsequently, scions from the 15 plants were used to graft-inoculate I. setosa. From each orginal source plant, six I. setosa plants were graft-inoculated. All I. setosa plants inoculated with scions of the Hawassa 83, Tola, Kulfo, and Guntute varieties showed virus-like symptoms 3 weeks after graft indexing to I. setosa (Table 2). Furthermore, all of these plants were positive for SPFMV according to the DAS–ELISA and RT–PCR (Table 2). Thus, the number of SPFMV-infected plants detected after graft inoculation increased from 6 to 12. In addition, none of the plants graft inoculated with scions belonging to Berkume plants developed any virus-like symptoms, nor did they test positive for any of the viruses tested by ELISA or RT–PCR.
As described above, ELISA was not able to detect SPFMV in 11 out of 12 plants prior to graft inoculation. On the other hand, RT–PCR assays detected SPFMV in 6 of the 12 plants prior to graft inoculation. Thus, RT–PCR is a more sensitive and reliable method for detecting SPFMV from sweet potato plants. Nevertheless, RT–PCR assays efficiently detected the virus in only 50% of the plants. Our findings are in line with previous studies reporting the challenges of detecting SPFMV by ELISA [25,29,30]. Consequently, we recommend (as previous authors have also recommended) that graft inoculation of I. setosa followed by ELISA and/or RT–PCR yields the most reliable results. We also suggest that ELISA is able to detect SPFMV in plants infected with SPCSV. Our results in Table 2 show that the DAS–ELISA and TAS–ELISA detected SPFMV and SPCSV, respectively, without the need for prior grafting from the severe symptomatic plant Guntute B (Figure 1b).
Previous studies have indicated that symptomless infection of sweet potato plants by SPFMV is not uncommon [31,32]. These findings are also exemplified in our study, in which 11 out of the 12 plants found to be infected with SPFMV were symptomless. In Ethiopia, there is a very weak plant virus certification scheme, since it relies only on visual evaluation in fields that do not consider symptomless infections. Using virus-infected planting materials not only reduces yield and quality [13] but also spreads the virus, since viruses accumulate and perpetuate over seasons in vegetatively propagated materials.
3.2. Report of the SPVC in Ethiopia
Although SPVC was detected first in this study, the presence of the virus was reported previously in our NGS experimental results generated without being claimed as a first report [11] after the results presented in this manuscript. In this study, SPVC was detected in one plant of the sweet potato variety Guntute, which was obtained from Hawassa Research Center, Ethiopia. This virus was detected via RT–PCR, in which a PCR product with a fragment size of approximately 1 kb was amplified and matched the expected size of 836 bp (Figure 1a). The identity of the virus was verified using next-generation sequencing in our other related experimental work, which has been published [11]. The infected plant appeared severely stunted and showed vein yellowing and chlorotic blotches (Figure 1b). These symptoms differed from those of all the other plants tested in this study, and it was subsequently revealed that this plant suffered from a mixed infection of SPVC, SPFMV, and SPCSV. None of the other plants or varieties tested by RT–PCR in this study were found to be infected by SPVC. However, NGS revealed the presence of SPCV in two more varieties of sweet potato from Ethiopia [11]. To the best of our knowledge, this is the first report of SPVC infecting sweet potato in Ethiopia. Previous surveys carried out in Ethiopia [6,7,8] did not include SPVC as a target virus. Therefore, it cannot be established whether SPVC has been present in the country for a long time or if it has recently been introduced by imported plant material.
3.3. Heat Therapy Followed by Meristem Tip Culture Increased the Efficiency of SPFMV Removal
The heat treatment of meristem donor plants before meristem excision resulted in a greater number of virus-free plants than the meristem excision without heat treatment (Figure 2). Samples from shoot tips/plants that did not induce virus symptoms in I. setosa after graft indexing to I. setosa were SPFMV-negative, according to the ELISA and RT–PCR. On the other hand, grafted plants that showed virus-like symptoms were positive for SPFMV, as confirmed by ELISA and RT–PCR.
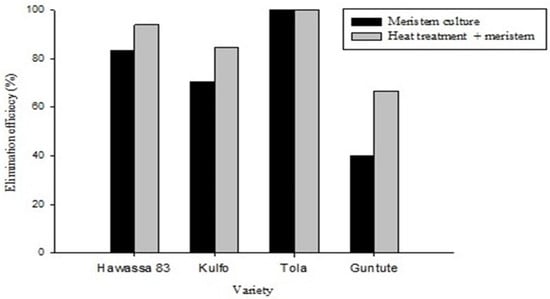
Figure 2.
Efficiency of virus elimination methods; the black-shaded graph represents the meristem culture alone, and the gray-shaded graph represents the meristem combined with heat treatment for four virus-infected genotypes of sweet potato.
Heat treatment of ‘Hawassa-83’ meristem donor plants followed by meristem culture eliminated SPFMV from 93.8% of the plantlets, whereas only 83.3% of ‘Hawassa-83’ plants obtained from the nontreated plants were SPFMV-free. In ‘Kulfo’, 84.7% and 70.6% of the SPFMV-free plantlets were obtained from heat-treated and -non-treated meristem donor plants, respectively. Only a small number of plants were regenerated from the variety ‘Tola’. We believe that this is due to poor regeneration media for this genotype; the optimum medium has not been found. Nevertheless, the plants derived from both sources (heat-treated and non-heat-treated) were 100% virus free. In the case of ‘Guntute’, only 66.7% and 40.0% of the plants derived from the heat-treated and -non-treated meristem donor plants, respectively, were found to be free of SPFMV.
Differences in response to virus elimination treatment were prominent among the varieties tested. The highest (100%) SPFMV elimination efficiency was obtained for ‘Tola’, followed by ‘Hawassa-83’ (93.8%), and ‘Kulfo’ (84.7%). The variety with the lowest efficiency (66.7%) was ‘Guntute’.
Differences in virus/viroid elimination efficiency between varieties of the same crop have previously been reported [33,34] and seem to be mediated by the specific interactions that occur between the virus and the host. For example, in the case of Argyranthemum plants infected by Chrysanthemum stunt viroid (CSVd), the accumulation of callose (β-1,3-glucan) around the plasmodesmata of shoot apical meristems differs between varieties and correlates with the feasibility of eliminating the viroid via meristem tip culture. Varieties that accumulate high amounts of callose are less likely to become viroid free after meristem tip culture. Studies have shown that callose accumulation in plasmodesmata is associated with a decrease in plasmodesmata opening [35], whereas virus/viroid movement from cell to cell is accomplished through plasmodesmata [36]. Thus, it has been suggested that callose deposition at plasmodesmata limits cell-to-cell movement of CSVd and is most likely responsible for the different frequencies of obtaining viroid-free materials among Argyranthemum varieties [37]. All of the sweet potato varieties in our study had different genetic backgrounds [38]. Thus, it is conceivable that the different frequencies observed in response to virus elimination treatment are due to specific virus–host interactions that vary among the sweet potato varieties tested.
Moreover, meristem tip culture alone and in combination with other virus elimination methods, heat therapy, chemotherapy, and cryotherapy have been successfully used as virus-elimination methods in sweet potato [39]. Previous studies have shown that heat treatment combined with meristem culture has better elimination efficiency in sweet potato and potato than meristem culture alone [34,40,41]. Thus, our study confirms that heat treatment of meristem donor plants increases the efficiency of virus elimination compared with the use of meristem culture alone.
4. Conclusions
In this study, we successfully eliminated SPFMV from four of the five most widely used sweet potato varieties in Ethiopia. Our data show that a combination of heat therapy and meristem tip culture is more efficient than meristem tip culture alone. We have also shown that efficient and reliable detection of SPFMV requires a combination of graft inoculation of I. setosa with either ELISA or RT–PCR. Moreover, our study is the first to report the SPVC in Ethiopia. Our study represents a step toward efficient virus-free production of sweet potato in Ethiopia. We envision that the experimental procedures presented in this paper will be used for the development of a virus-free sweet potato production scheme in Ethiopia.
Author Contributions
Conceptualization, D.H.B., C.S. and T.H.-E.; methodology, D.H.B. and T.H.-E.; data curation, D.H.B.; writing—original draft, D.H.B.; writing—review and editing, C.S. and T.H.-E.; project administration, T.H.-E. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the NORAD-funded project “Controlling disease in sweet potato and enset in South Sudan and Ethiopia to improve productivity and livelihoods under changing climatic conditions using modern technologies” under the NORHED program (Agreement no ETH-13/0017, 2013).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are available on request.
Acknowledgments
We received support from technicians Astrid Helene Sivertsen (meristem cuttings to clean viruses, media preparation) and Gry Skjeseth (heat treatment, screening by grafting, and taking care of plants in the greenhouse). They are duly acknowledged for their contributions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Low, J.; Lynam, J.; Lemaga, B.; Crissman, C.; Barker, I.; Thiele, G.; Namanda, S.; Wheatley, C.; Andrade, M. Sweet potato in Sub-Saharan Africa. In The Sweetpotato; Springer: Berlin/Heidelberg, Germany, 2009; pp. 359–390. [Google Scholar]
- Belehu, T. Agronomical and Physiological Factors Affecting Growth, Development and Yield of Sweet Potato in Ethiopia. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2003. [Google Scholar]
- Truong, V.D.; Avula, R.Y.; Pecota, K.V.; Yencho, G.C. Sweetpotato production, processing, and nutritional quality. In Handbook of Vegetables and Vegetable Processing; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 811–838. [Google Scholar]
- Aldow, A.M. Factors Affecting Sweet Potato Production in Crop-Livestock Farming Systems in Ethiopia. Master’s Thesis, Norwegian University of Life Sciences (NMBU), As, Norway, 2017. [Google Scholar]
- Clark, C.; Davis, J.A.; Abad, J.A.; Cuellar, W.J.; Fuentes, S.; Kreuze, J.F.; Gibson, R.W.; Mukasa, S.B.; Tugume, A.K.; Tairo, F.D. Sweet potato viruses: 15 years of progress on understanding and managing complex diseases. Plant Dis. 2012, 96, 168–185. [Google Scholar] [CrossRef] [PubMed]
- Alemu, T. Characterization of Viruses of Pepper (Capsicum spp.) and Sweet Potato (Ipomoea batatas) from Ethiopia; Cuvillier Verlag: Göttingen, Germany, 2004; 126p. [Google Scholar]
- Adane, A. Associated viruses threatening sweet potato improvement and production in Ethiopia. Afr. Crop Sci. J. 2010, 18, 207–213. [Google Scholar]
- Tesfaye, T.; Feyissa, T.; Abraham, A. Survey and serological detection of sweet potato (Ipomoea batatas (L.) Lam) viruses in Ethiopia. J. Appl. Biosci. 2011, 41, 2746–2756. [Google Scholar]
- Feyissa, T.; Dugassa, G. In vitro production of virus-free sweet potato [Ipomoea batatas (L.) Lam] by meristem culture and thermotherapy. Ethiop. J. Sci. 2011, 34, 17–28. [Google Scholar]
- Wondimu, T.; Feyissa, T.; Bedadav, G. Meristem culture of selected sweet potato (Ipomoea batatas L. Lam.) cultivars to produce virus-free planting material. J. Hortic. Sci. Biotechnol. 2012, 87, 255–260. [Google Scholar] [CrossRef]
- Buko, D.H.; Spetz, C.; Hvoslef-Eide, A.K. Next generation sequencing as a method to verify virus elimination using heat treatment and meristem tip culture in the five most widely used sweet potato varieties in Ethiopia. Afr. J. Biotechnol. 2020, 19, 458–463. [Google Scholar]
- Buko, D.H. Sweet Potato Virus in Ethiopia: Detection, Characterization, Elimination and Management. Ph.D. Thesis, Norwegian University of Life Sciences, Ås, Norway, 2019. Available online: https://nmbu.brage.unit.no/nmbu-xmlui/bitstream/handle/11250/2711545/105146_Dereje%20Haile%20Buko%20PhD%20thesis.pdf?isAllowed=y&sequence=1 (accessed on 1 June 2021).
- Clark, C.A.; Moyer, J.W. Compendium of Sweet Potato Diseases; American Phytopathological Society: St. Paul, MN, USA, 1988. [Google Scholar]
- Opiyo, S.; Ateka, E.; Owuor, P.; Manguro, L.; Miano, D. Development of a multiplex PCR technique for simultaneous detection of Sweet potato feathery mottle virus and Sweet potato chlorotic stunt virus. J. Plant Pathol. 2010, 92, 363–366. [Google Scholar]
- Van der Want, J.; Dijkstra, J. A history of plant virology. Arch. Virol. 2006, 151, 1467–1498. [Google Scholar] [CrossRef] [PubMed]
- Golino, D.; Fuchs, M.; Al Rwahnih, M.; Farrar, K.; Schmidt, A.; Martelli, G. Regulatory aspects of grape viruses and virus diseases: Certification, quarantine, and harmonization. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Berlin/Heidelberg, Germany, 2017; pp. 581–598. [Google Scholar]
- Spiegel, S.; Stein, A.; Tam, Y. In vitro thermotherapy of rosaceous fruit trees. In Proceedings of the XVIth International Symposium on Fruit Tree Virus Diseases, Rome, Italy, 27 June–2 July 1994; pp. 419–420. [Google Scholar]
- Wang, Q.C.; Valkonen, J.P. Elimination of two viruses which interact synergistically from sweet potato by shoot tip culture and cryotherapy. J. Virol. Methods 2008, 154, 135–145. [Google Scholar] [CrossRef]
- Panattoni, A.; Luvisi, A.; Triolo, E. Elimination of viruses in plants: Twenty years of progress. Span. J. Agric. Res. 2013, 11, 173–188. [Google Scholar] [CrossRef]
- Nyland, G.; Goheen, A.C. Heat therapy of virus diseases of perennial plants. Annu. Rev. Phytopathol. 1969, 7, 331. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zuo, R.; Abad, J.; Xu, D.; Bao, G.; Li, R. Simultaneous detection and differentiation of four closely related sweet potato potyviruses by a multiplex one-step RT–PCR. J. Virol. Methods 2012, 186, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Kathurima, T.; Bett, B.; Miano, D.; Kim, D. Diagnostics of viruses infecting local farmer preferred sweet potato cultivars in Kenya. Afr. J. Agric. Res. 2011, 6, 3718–3724. [Google Scholar]
- Zheng, Y.; Gao, S.; Padmanabhan, C.; Li, R.; Galvez, M.; Gutierrez, D.; Fuentes, S.; Ling, K.S.; Kreuze, J.; Fei, Z. VirusDetect: An automated pipeline for efficient virus discovery using deep sequencing of small RNAs. Virology 2017, 500, 130–138. [Google Scholar] [CrossRef]
- Aritua, V.; Bua, B.; Barg, E.; Vetten, H.; Adipala, E.; Gibson, R. Incidence of five viruses infecting sweet potato es in Uganda; the first evidence of Sweet potato caulimo-like virus in Africa. Plant Pathol. 2007, 56, 324–331. [Google Scholar] [CrossRef]
- Dennien, S.; Homare, D.; Hughes, M.; Lovatt, J.; Coleman, E.; Jackson, G. Growing Healthy Sweet Potato: Best Practices for Producing Planting Material; ACIAR: Canberra, Australian, 2013. [Google Scholar]
- Buko, D.H.; Hvoslef-Eide, T.A. Optimization of plant growth regulators for meristem initiation and subsequent multiplication of five virus tested elite sweet potato varieties from Ethiopia. Afr. J. Biotechnol. 2020, 19, 332–343. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Green, S.; Kuo, Y.; Lee, D. Uneven distribution of two potyviruses (Sweet potato feathery mottle virus and sweet potato latent virus) in sweet potato plants and its implication on virus indexing of meristem derived plants. Int. J. Pest Manag. 1988, 34, 298–302. [Google Scholar] [CrossRef]
- Moyer, J.W.; Salazar, L. Viruses and virus-like diseases of sweet potato. Plant Dis. 1989, 73, 451–455. [Google Scholar] [CrossRef]
- Gibson, R.; Mwanga, R.; Kasule, S.; Mpembe, I.; Carey, E. Apparent absence of viruses in most asymptomatic field-grown sweet potato in Uganda. Ann. Appl. Biol. 1997, 130, 481–490. [Google Scholar] [CrossRef]
- Tugume, A.; Mukasa, S.; Valkonen, J. Natural wild hosts of sweet potato feathery mottle virus show spatial differences in virus incidence and virus-like diseases in Uganda. Phytopathology 2008, 98, 640–652. [Google Scholar] [CrossRef]
- Paprstein, F.; Sedlak, J.; Polak, J.; Svobodova, L.; Hassan, M.; Bryxiova, M. Results of in vitro thermotherapy of apple cultivars. Plant Cell Tissue Organ Cult. 2008, 94, 347–352. [Google Scholar] [CrossRef]
- Waswa, M.; Kakuhenzire, R.; Ochwo-Ssemakula, M. Effect of thermotherapy duration, virus type and cultivar interactions on elimination of potato viruses X and S in infected seed stocks. Afr. J. Plant Sci. 2017, 11, 61–70. [Google Scholar]
- Bilska, A.; Sowiński, P. Closure of plasmodesmata in maize (Zea mays) at low temperature: A new mechanism for inhibition of photosynthesis. Ann. Bot. 2010, 106, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Plant Virology; Academic Press: Norwich, UK, 2013; 1104p. [Google Scholar]
- Zhang, Z.; Lee, Y.; Spetz, C.; Clarke, J.L.; Wang, Q.; Blystad, D.-R. Invasion of shoot apical meristems by Chrysanthemum stunt viroid differs among Argyranthemum cultivars. Front. Plant Sci. 2015, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Kosmowski, F.; Aragaw, A.; Kilian, A.; Ambel, A.; Ilukor, J.; Yigezu, B.; Stevenson, J. Varietal identification in household surveys: Results from three household-based methods against the benchmark of DNA fingerprinting in southern Ethiopia. Exp. Agric. 2018, 55, 1–15. [Google Scholar] [CrossRef]
- Wang, Q.; Panis, B.; Engelmann, F.; Lambardi, M.; Valkonen, J. Cryotherapy of shoot tips: A technique for pathogen eradication to produce healthy planting materials and prepare healthy plant genetic resources for cryopreservation. Ann. Appl. Biol. 2009, 154, 351–363. [Google Scholar] [CrossRef]
- Dodds, J.; Ng, S.Y.C. In vitro methods for pathogen elimination and international distribution of sweet potato germplasm exploration, maintenance and utilization of sweet potato genetic resources. In Proceedings of the Report 1st Planning Conference of Exploration, Maintenance and Utilization of Sweet Potato Resources, International Potato Centre, Lima, Perú, 23–27 February 1987. [Google Scholar]
- El Far, M.M.; Ashoub, A. Utility of thermotherapy and meristem tip for freeing sweet potato from viral infection. Aust. J. Basic Appl. Sci. 2009, 3, 153–159. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).