Abstract
One of the most serious diseases affecting dairy cattle, causing significant losses both in breeding and economy, is mastitis, an inflammation of the mammary gland. Due to the economic importance of this issue, many research teams are striving to develop an easy-to-apply and, most importantly, effective method to prevent mastitis. The use of traditional methods for mastitis detecting and treating, as well as improvement in hygienic conditions, have not yielded the expected results in combating this disease combating. Currently, the main task is to find the tools that would allow for the rapid detection of mastitis and the improvement of udder health in cows while maintaining high milk production, which is essential for the profitability of dairy cattle farming. Accurate and rapid diagnostic tools, with the simultaneous capability of identifying pathogens, may help to reduce losses. Sufficient sensitivity and specificity for tests are required to minimize the number of false-positive and false-negative cases. Efforts are also being made to determine the optimal threshold value for detecting the disease at its earliest possible stage. The estimation of somatic cell count (SCC) as a phenotypic indicator of mastitis is widely used. A more precise parameter for accurately describing udder health is the differential somatic cell count (DSCC). The well-known California Mastitis Test (CMT) is an inexpensive, easy, and rapid method for mastitis detection useful on farms. The latest diagnostic methods for mastitis utilize tests based on the activity of N-acetyl-β-d-glucosaminidase (NAGase) or lactate dehydrogenase (LDH) as well as the determination of acute phase proteins (APPs) in blood serum and milk (such as haptoglobin, serum amyloid A, fibrinogen, and ceruloplasmin). Research also focuses on the genomic improvement of mastitis resistance in successive generations, and for this purpose, many quantitative trait loci (QTLs) and single nucleotide polymorphisms (SNPs) have been identified. In recent years, immunotherapy has become an increasingly common area of research, including vaccinations, T/B cell immunotherapy, RNA immunotherapy, epigenetic immunotherapy, stem cell therapy, and native secretory factors. An important aspect of the control of mastitis is the implementation of strategies that focus primarily on preventing the disease through appropriate breeding and farm management practices. In the forthcoming years, a significant challenge will be the development of universal diagnostic and therapeutic strategies that can be effectively implemented as alternatives to antibiotic therapy. Future research should prioritize the advancement of preventive and therapeutic techniques, such as immunotherapies, bacteriocins, herbal therapy, and nanoparticle technology.
1. Introduction
Mastitis is a multi-etiological disease associated with the cattle production system and rearing environment. It is manifested by physical, chemical, and bacteriological changes in the mammary gland tissue [1,2,3]. Mastitis is a result of inflammation of the udder gland tissue and the defense reaction of the organism to some factors, mainly the microorganisms present inside the mammary gland [4]. The infection develops when the organism’s defense mechanisms are weakened. The teat canal forms the first line of defense against invading pathogens by providing a physical barrier and a source of antimicrobial substances (Figure 1). Teat canal keratin plays particular importance in non-specific defense of the gland, serving as a physical barrier [5,6]. Keratin consists of fatty acids and fibrous proteins. Upon pathogen entry into the teat canal, these fibrous proteins bind electrostatically to the pathogens, resulting in alterations to their cell walls. This modification increases the pathogens’ susceptibility to osmotic pressure. The inability to regulate osmotic pressure leads to lysis of the cell membranes and subsequent death of the invading pathogens [7]. Additional factors limiting the penetration of glandular tissue by pathogens include such substances as lysozyme, lactoferrin, and lactoperoxidase. With their nonspecific effect on the infectious agent, they are classified in the group of non-specific humoral substances. Important components of the innate defense mechanisms of the mammary gland are endothelial cells (which control blood flow to affected tissues and regulate leukocyte migration and activation) and oxylipids (responsible mainly for regulation of microvasculature). Neutrophils are the most active cell line with direct involvement in the defense of the udder against microbes. Like neutrophils, macrophages and dendritic cells are active mammary gland defense cells that are capable of phagocytosis and cytokine production. Dendritic cells, as well as macrophages, are also capable of antigen presentation [5,8,9]. Another form of defense is the specific cellular and humoral immunity associated with the release of antibodies and various fractions of T and B cells into the bloodstream. They produce cytokines, e.g., interleukins (ILs), tumor necrosis factor-alpha (TNF-α), and eicosanoids [10,11,12]. The differences in their production and kinetics may cause large variations in the course of the inflammatory process. The accompanying symptoms also vary, ranging from a lack of visible abnormalities in the udder and milk through reduced milk production and changes in its composition and/or appearance as well as redness, hardness, sensitivity, pain, and itching of the udder to systemic symptoms [13].
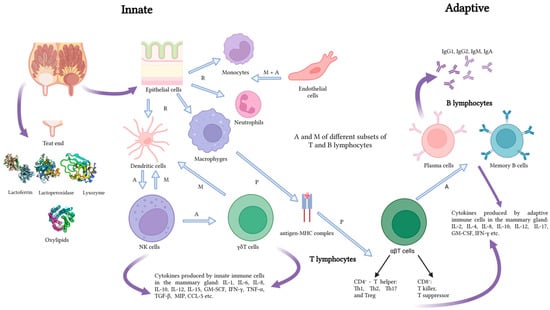
Figure 1.
Predominant innate and adaptive defense mechanisms of the mammary gland. Explanations: A—activation, M—migration, P—presentation, R—recognition. Source: own study.
Since high-yielding breeds are more prone to inflammation, higher losses in milk production are recorded, as shown by Bronzo et al. [8]. The reduced synthetic activity of the infected mammary gland is also manifested by changes in the content of milk constituents and its physicochemical properties (pH, density, conductivity), which in turn reduce its nutritional value and suitability for processing [13]. Mastitis leads to changes in protein distribution (the proportion of whey proteins increases at the expense of casein), which leads to reduced fat and lactose content and changes in the concentration of minerals, mainly a reduction in the calcium content [2]. Unfavorable changes in the milk composition caused by mastitis exert a negative effect on its quality and suitability for processing. As reported by Pegolo et al. [13], mastitis leads to the reduction of curd firmness, recovery of nutrients (protein and fat) in the curd, milk coagulation properties, and cheese traits. The magnitude of these changes depends on the etiological factor.
Prevention plays a critical role in mastitis management in dairy cattle [14]. Effective mastitis prevention encompasses a multifaceted approach, integrating appropriate milking procedures, environmental management, and herd health strategies [15,16]. Regular monitoring and highly sensitive methods serve as fundamental tools for early detection and intervention [17]. There is an urgent need for the development of new and implementation of existing portable mastitis detection systems [18]. Immunotherapy targeted at common mastitis pathogens can also enhance herd immunity. Ultimately, a comprehensive prevention program not only reduces the incidence of mastitis but also improves the overall health and productivity of the dairy cattle herd. The aim of our study was to review and systematize the current state of knowledge and the latest discoveries concerning the diagnosis of mastitis and the study of biomarkers of udder inflammation. We also included some management practices in mastitis prevention and mastitis immunotherapy as well as nanoparticle-based treatments as an alternative to the increasing antibiotic resistance of the etiological agents of mastitis.
2. Methods
The review and analysis of information available in the world scientific literature were conducted in April and May 2024 using the Science Direct, Scopus, and Agricola databases. The databases were searched according to the keyword combinations included in Table 1. The date range of the search was from 2010 to 2024. The search field was selected, where the search was directed through titles, abstracts, and keywords, among others; this was specific to each database.

Table 1.
Search strategy keyword combinations.
The results of this search were downloaded to RIS library files and then imported into Rayyan Software as a Service (web application) for systematic reviews [19]. The Rayyan database of records was supplemented with articles from other sources, such as citations, as well as with publications important for our review but published before 2010. In the next step, duplicate selection was performed and then records were screened for inclusion or exclusion from the review. Finally, 244 articles that meet the established criteria were gathered and analyzed. Figure 2 illustrates the process in a flow chart using the PRISMA methodology.
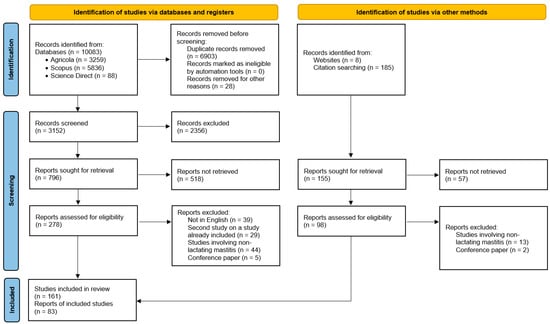
Figure 2.
Study selection flow diagram showing the stages from initial search to the finalization of publications included in the review.
3. Pathogens Involved and Severity of Clinical Signs in Mastitis
Bovine mastitis can be divided into clinical and subclinical types. Clinical mastitis (CM) is a severe condition with local symptoms visible in the mammary gland and systemic symptoms, e.g., redness and inflammation of affected areas, pain, loss of appetite, elevated body temperature, reduced milk yield, and changes in the milk composition [20]. In severe cases, abnormal teat discharge can be noticed, e.g., floccules and watery milk or blood in the milk [21]. According to Cheng and Han [22], the severe clinical mastitis caused by E. coli may cause irreversible tissue damage in the mammary gland, complete milk production loss, and sometimes even lead to the death of the dairy cow. The intensity and course of infection are influenced by a variety of factors, e.g., the type of pathogens, the age, immunity, and health status of the animal, and the course of lactation [23]. The clinical form can be further classified as hyperacute, acute, and subacute depending on the severity of symptoms [15]. Hyperacute mastitis is characterized by severe inflammation and significant milk yield reduction and changes in its composition. It is accompanied by systemic symptoms such as fever, depression, chills, and loss of appetite and body weight. Acute mastitis is similar to hyperacute mastitis but with fewer systemic symptoms such as fever and mild depression. In subacute mastitis, the inflammatory symptoms are minimal and there are no visible systemic symptoms [24].
In subclinical mastitis (SCM), the appearance of milk does not change. One of the indicators of the possible development of inflammation is the increased somatic cell count. Other indicators of SCM include an increase in the abundance of bacterial populations in milk, reduction of milk yields, and changes in milk composition and quality [25]. As reported by Griffioen et al. [26], detection of SCM is indispensable for the choice of strategies targeted at control, prevention, and treatment of the disease. The prophylaxis of SCM requires laboratory diagnostics targeted at the isolation and identification of the pathogens responsible for the disease [27,28,29].
The etiological agents of mastitis are mainly bacteria, especially staphylococci and streptococci, as well as mycoplasmas, fungi, algae, protozoa, and viruses. Inflammation may also develop upon physical trauma to the udder, contact with irritating chemicals triggering immune response, or any physiological damage [16,30]. Bacterial mastitis is highly important due to its physiological complications and economic consequences [14,30]. As reported by the National Mastitis Council [31], approximately 200 microorganisms causing mastitis in cattle have been identified, but only a few species are isolated on a regular basis. Kuang et al. [32] have shown that over 150 species of both Gram-positive and Gram-negative bacteria are responsible for development of mastitis. Gram-positive bacteria include various species of staphylococci and streptococci, while E. coli and Klebsiella pneumoniae are the most common Gram-negative bacteria causing the disease [33].
Depending on the etiological factor (mode of infection), two categories of mastitis have been distinguished, that caused by infectious agents and that caused by environmental agents, although Klaas and Zadoks [34] suggest that this division may be misleading in some cases. The infectious agents (e.g., Staphylococcus aureus, Streptococcus agalactiae, Streptococcus dysgalactiae) are mainly responsible for subclinical and chronic mammary gland inflammation [35,36]. They colonize the udder skin, adapt to survive and multiply in the mammary gland, and so are transferred between animals during milking. In turn, environmental pathogens, i.e., bacteria from the coli group (Escherichia coli, Klebsiella pneumoniae), Streptococcus uberis, or Enterococci spp., present in the animals’ environment (soil, water, litter) are quickly fought by the host’s immune system [30,35,36,37].
Yeasts are a less common cause of mastitis [38]. However, in herds characterized by poor milking hygiene, the frequent occurrence of udder irritation and injuries, and excessive use of antibiotics or multiple intramammary treatments, mastitis was caused by, e.g., several species of the genera Candida, Cryptococcus, Rhodotorula, and Trichosporum [39,40,41]. Noteworthily, mastitis can also be caused by two of the eight known species of Prototheca algae, i.e., P. zopfii and P. blaschkeae. It should also be noted that only genotype 2 of the two P. zopfii genotypes has been isolated from diseased animals [42].
In addition to pathogenic agents, udder inflammation can be caused by factors that are directly associated with the organism of the cow. These include the presence or absence of natural immune mechanisms, milk yield, age, lactation phase, stress factors, and morphological features of the udder and teats. Mastitis can also be induced by environmental factors, e.g., the methods and conditions of milking, housing type, microclimatic conditions, production season, and nutrition [7,30,43].
4. Losses Associated with Dairy Cow Mastitis
Mastitis is one of the costliest diseases in dairy cattle. The economic losses associated with udder inflammation stem mainly from decreased milk production and quality, increased labor and treatment costs, and the shortened productive life of affected cows. Following infertility and limb diseases (lameness), mastitis is one of the most frequent reasons for cow culling [30,44,45]. As suggested by Van Soest et al. [46], the economic costs of mastitis should include not only the actual losses but also expenditure on preventive measures.
As demonstrated by Awale et al. [30] and Bezman et al. [44], it is difficult to estimate the magnitude of mastitis-related economic losses. The differences in the volume of losses depend mainly on the actual price of milk, milk yield reduction, applied drugs, herd treatment, renovation, nutrition costs, and the duration, advancement, and course of the disease. Huijps et al. [47] emphasize that milk producers tend to underestimate the importance of these costs as they are difficult to assess and evaluate.
While considering the costs, it is worth realizing the extent of the prevalence of this disease. As demonstrated by Krishnamoorthy [48], its clinical form affects 29% of the European cow population and about 22% of the North American population. However, this form is slightly less frequent in Asia (18%) and Africa (12%). At the same time, it should be emphasized that mastitis accounts for approximately 70% of economic losses on dairy farms [47]. In the case of the clinical form, the losses are generated mainly by the cost of culling and replacing (48%), reduction of milk yields (34%), unsuitability of milk for consumption and processing (11%), increased labor costs (3%), and diagnosis and animal treatment costs (2%). In contrast, the chronic form includes costs related to reduced milk yield (72%), the likelihood of culling (25%), and, to a lesser extent, discarded milk and diagnosis and veterinary services [49].
As reported by Stevens et al. [50], mastitis prophylaxis and treatment measures account for approximately 60–70% of all antimicrobials used on dairy farms. Cha et al. [51] showed that the cost of CM varied according to the etiologic factor and was the highest for inflammation caused by Gram-negative bacteria (211.03 USD), lower for Gram-positive bacteria (133.73 USD), and the lowest for other pathogens (95.31 USD). In contrast, much higher values were estimated for CM by Liang et al. [52], i.e., 326 USD and even 444 USD for CM during the first 30 days of lactation [53].
The amount of production losses estimated by Hagnestam et al. [1] varied from 0 to 705 kg for the first and from 0 to 902 kg for the second and subsequent lactations. Heikkilä et al. [36], in turn, estimated daily milk loss for E. coli mastitis of 3.5 kg/d. Bezman et al. [44] found an average 25% decline in the milk yield in mastitis-affected cows. The authors observed differences in the yield reduction depending on the mastitis-causing pathogen. A 20% or 50% decrease in milk yield was reported for mastitis caused by S. dysgalactiae or E. coli, respectively. Cobirka et al. [7] suggest that the milk produced by cows with CM is unsuitable for processing. Therefore, the economic losses associated with the necessity to discard milk (e.g., costs of feeding) are substantially higher than the losses caused by reduced milk yields.
The losses generated by SCM are even higher. This is mainly related to the long-term reduction in milk yield, as many cows produce amounts of milk much below their potential. It is worth mentioning that mastitis reduces milk production not only during the inflammation but also throughout the lactation period [1,36]. However, some researchers did not observe a significant effect of SCM caused by various pathogens on the amount of milk loss [54,55,56]. Mastitis alters not only the milk yield but also its chemical composition. A reduction in non-fat solids and lactose in milk from infected quarters was observed by Malek dos Reis et al. [57]. The level of these changes was influenced by the causative agent. Batavani et al. [58] reported that milk from quarters with subclinical mastitis showed lowered contents of calcium (90.45 mg/dL), potassium (151.56 mg/dL), inorganic phosphorous (24.40 mg/dl), α-lactalbumin (22.25%), and β-lactoglobulin (34.21%). They also found elevated levels of sodium (91.97 mg/dL), chloride (>0.14 g/dL), pH (6.69), albumin (5.62 g/dL), lactate dehydrogenase (LDH) activity (1524.04 IU/L), and immunoglobulins (26.86%). Gonçalves et al. [55] found no significant changes in protein and fat content between healthy cows and those with SCM. As shown by the authors, cows with SCM had a moderately higher somatic cell count in milk, which had no significant effect on the milk yield.
5. Mastitis Detection
As suggested by Martin et al. [17], mastitis induced by the milking process itself is extremely difficult to eliminate. Therefore, accurate and rapid diagnostic tools for continuous surveillance and early detection of mastitis with a concurrent possibility to identify pathogens can help to reduce the mastitis-related rearing and production losses.
A diagnostic test with high sensitivity and specificity is the key to solving accurate mastitis diagnosis. Sensitivity describes the ability to designate an animal with a disease as positive; if the diagnostic test is highly sensitive, it has few false-negative results, so fewer positive cases are missed. Specificity refers to the ability to identify an animal that does not have a disease as negative; if a test has high specificity, there are fewer false-positive results. It is desirable to use a test that exhibits both high sensitivity and high specificity (with similar values c.a. 0.7–0.8), with the combined value of these indicators not falling below 1.5 [59].
5.1. Classic on-Farm and Laboratory Methods
The somatic cell count (SCC) in milk has been widely used as a phenotypic indicator of mastitis since the late 1970s. It shows the total number of immune cells in milk, mainly neutrophils, lymphocytes, and macrophages [60,61]. Addis et al. [62] emphasize the high importance of monitoring the SCC level in milk as an indicator of mastitis, especially in its subclinical form. As shown by Viguier et al. [63] and Adkins and Middleton [64], the SCC value can be determined in the laboratory; however, this time-consuming method requires high-quality equipment and qualified personnel. Alternatively, somatic cell counters based on flow cytometry combined with imaging techniques are available [65]. However, their results may be distorted, as milk is a water–fat emulsion [66]. Flow cytometry can also be used for the detection and quantifying of mastitis-related bacteria (Gram-negative bacteria stained less intensely compared with Gram-positive bacteria) [67].
Many authors highlight the diagnostic sensitivity and specificity of various methods and tests used for the estimation of the SCC and the types of immune cells for the detection of mastitis [3,68,69,70,71,72,73,74]. The SCC value may be influenced by many factors, e.g., stress, nutrition, lactation phase, parturition, and the quality of the sampled milk fraction. Due to the variability in SCC, this method has varying sensitivity and specificity (Table 2). Much depends also on the adopted cut-off value (threshold). Lowering the threshold increases sensitivity and, consequently, provides minimal false-negative results, whereas raising the threshold increases specificity, providing minimal false-positive results [3,69,71]. To reduce the diagnostic error, a threshold of 200,000 cells/mL was generally accepted as the level of SCM, although a threshold of ≥100,000 cells/mL [75] or >150,000 cells/mL [76] is increasingly suggested, particularly for primiparous. For the threshold of 200,000 cells/mL, Pantoja et al. [69] estimated 0.64, 0.69, and 0.65 sensitivity for milk samples obtained in the dry-off, post-calving, and first dairy herd improvement (DHI) test of the subsequent lactation, respectively. The specificity was estimated at 0.66, 0.84, and 0.93, respectively. In contrast, Ferronatto et al. [70] suggested varying cut-off values for optimizing sensitivity versus specificity at different postpartum periods, with particular emphasis on 1–3 d postpartum (colostrum and transitional milk). In these studies, a significant correlation (r = 0.61) was also found for the automated and microscopic cell counting. These two methods also had differences in sensitivity and specificity during 1–30 d after calving, and interestingly, the highest prediction area under a receiver operator characteristic curve (AUC) value (0.74–0.75) was found on the day of calving (with relatively high cut-off values). The AUC quantifies the overall ability of a test to discriminate between healthy and infected quarters.
Table 2.
The diagnostic performance of tests based on the classical methods for detecting dairy cows with intramammary infections.
Noteworthily is the use of the somatic cell count (SCC) in milk at the end of lactation as an indicator of udder infection during the dry period [3]. As reported by Scherpenzeel et al. [88] and Vanhoudt et al. [89], the SCC results registered in the pre-dry period have become the main selection criterion for selective dry cow therapy (SDCT). In this strategy, long-acting antimicrobials are administered only to the cows that are most likely to be infected with the major pathogens during the dry period. It is therefore possible to make an informed choice of the SCC threshold depending on the farm strategy in terms of the level of antibiotics used and possible intention to reduce the use of the drugs. For example, in the case of strategies aimed at the reduction of the risk of infected quarters being omitted from antibiotic therapy (i.e., high sensitivity), a relatively low SCC threshold should be employed. The side effect for this relatively sensitive approach will be a decrease in specificity and, consequently, a higher proportion of “healthy” quarters being treated with antibiotics at dry-off. In contrast, when the assumption is the reduction of antibiotic use, a higher SCC threshold should be adopted, as this will increase specificity and decrease the apparent prevalence (decrease sensitivity) [3,68].
The California Mastitis Test (CMT) is a well-known inexpensive, easy, and quick (although not very accurate) method of SCC assessment that can be used on the farm. It is carried out by the addition of a sodium lauryl sulfate reagent to destroy the membranes of the cells present in milk and release their DNA, in contact with which the reagent forms a visible gel. Changes are visually scored using a five-point scale (negative, trace, weak positive, distinct positive, strong positive) [64,90,91]. As it is based on somatic cells, this method has similar limitations (e.g., false positives in cows in the postpartum period or immediately before drying off). Additionally, it is subject to human error in the form of subjective reading and interpretation of changes after the addition of the detergent. Therefore, this method is characterized by the variable sensitivity and specificity of the prediction of SCM. The high sensitivity (0.95) and specificity (0.86) of this method for the threshold at the CMT score of trace or higher was confirmed by Kandeel et al. [77] immediately prior to drying off. In cows after calving, the sensitivity decreased (0.79) while the specificity increased (0.95), and both resulted in a high AUC value (0.95). The above studies also showed higher sensitivity values for CMT than commercial tests based on esterase activity: PortaSCC color test, PortaSCC reader, and PortaSCC quick. Similar sensitivity values were confirmed by Bhutto et al. [78] for a threshold of trace or higher (0.86). The specificity was significantly lower in this case (0.26). Increasing the threshold to weak positive or higher resulted in decreased sensitivity (0.75) and increased specificity (0.50). Classical analysis at the weak positive cut-off points for levels >200,000 cells/mL showed slightly lower sensitivity (0.67) and specificity (0.87). However, these values were increased using Bayesian latent class analysis and resulted in a sensitivity of 0.95 and a specificity of 0.77, while increasing the AUC from 0.79 to 0.93 [79]. It should also be noted that the first week of lactation is characterized by high variability in the quality of inflammation prediction, as confirmed by Dingwell et al. [80], and the highest values of sensitivity (0.82) and specificity (0.81) are observed for trace or higher CMT scores. A relatively high positive predicted value (PPV) (0.42) and negative predicted value (NPV) (0.96) were found on the fourth day of lactation. This is confirmed by the results obtained by Sargeant et al. [92], who also differentiated the quality of this test according to the etiological factor.
A relatively new and more accurate parameter for the precise description of actual udder health in dairy cows is the differential somatic cell count (DSCC). The DSCC is the combined percentage of polymorphonuclear leukocytes (PMNs) and lymphocytes compared to the total number of PMNs, macrophages, and lymphocytes. In milk samples obtained from healthy udders, macrophages and lymphocytes dominate the total cells, while PMNs dominate the cell count in milk obtained from infected mammary glands or milk with a high SCC; this is related to the different roles of individual cells in the immune response [93]. Monitoring this indicator provides more information about udder health than the SCC, since an increase in PMNs can be observed even at low SCC levels [60,93,94]. Therefore, elevated DSCC scores indicate the presence of mastitis much earlier [60,95]. Certainly, intramammary inflammation is not the only factor influencing the proportion of individual cell populations; others include days in milk (DIM) and lactation number [96,97,98]. Zecconi et al. [98] showed that lymphocytes decreased over DIM, while macrophages and PMNs increased. On the other hand, Pilla et al. [95] found that only macrophages were significantly influenced by DIM, without a significant effect of lactation number or quarter position. Since this is a relatively new method, there is no well-defined cut-off point to predict SCM. The use of only the DSCC score in the 60–80% range in linear mixed models resulted in 0.34–0.92 sensitivity and 0.58–0.88 specificity, with a low PPV of 0.19–0.48 and a high NPV of 0.89–0.98. In contrast, the higher values of sensitivity while maintaining specificity allowed both the DSCC (60–80%) and SCC (100,000–200,000/mL) to be included in the model [72]. As a result, several optimal cut-off values were proposed, i.e., 63–68% DSCC depending on the milking time and sample collection and, similarly, for the combination of DSCC and SCC 67% and 163,000 cells/mL for the DHI a.m. samples, 65% and 131,000 cells/mL for the DHI p.m. samples, and 65% and 184,000 cells/mL for the 24h DHI samples. Similar relationships were found in the study conducted by Schwarz et al. [73] on the identification of mastitis at the end of lactation, where the combination of the DSCC and SCC showed higher sensitivity parameters, unfortunately at the expense of specificity. This was not confirmed based on statistical differences in AUC. Again, an optimal cut-off was proposed, i.e., DSCC 59%, 135,000 cells/mL, and 55% for the combination of DSCC and SCC and 131,000 cells/mL for SCC. In turn, with the combined DSCC and SCC method, Pilla et al. [86] obtained 0.97 sensitivity and 0.92 specificity using a threshold of 49.5%. As suggested by Wall et al. [99] and Schwarz et al. [85], the combination of the DSCC and SCC methods in mastitis diagnostics may increase the sensitivity in monitoring mastitis as part of DHI programs.
An important element in the strategy for the reduction or treatment of mastitis-affected cows is to identify the etiological factors of the disease. Hence, it is necessary to perform microbiological assays. Microbiological tests are also carried out to determine the sensitivity of pathogens to the active substances of antibiotics. An issue that is widely discussed in the literature is the methodology of collecting and handling milk samples intended for microbiological cultures. The problems considered include, e.g., the timing of sampling (pre-milking vs. post-milking or both), the sample size (e.g., 2–3 samples in a given pre-milking term), and the procedure (fresh or frozen milk cultures) [81,82,83,84]. As shown by Sears et al. [81], the sensitivity of pre-milking samples was 0.91 for Staphylococcus aureus, 0.91 for coagulase-negative staphylococci (CNS), and 0.97 for streptococci other than agalactiae. Significantly lower sensitivity values were determined in post-milking samples, i.e., 0.81, 0.45, and 0.58 for the same pathogens, respectively. In turn, the specificity was in the range of 0.86–0.92 depending on the pathogen in the case of pre-milking samples and 0.96–0.99 in the post-milking samples. These results were confirmed by Godden et al. [82]. The authors demonstrated that the sensitivity of S. aureus cultures was improved by the sample freezing process. The effectiveness of the multiplication of sampling was highlighted by Souza et al. [83]. They found that duplicate samples had a higher AUC value and higher sensitivity and specificity. Reyher and Dohoo [84] showed significant differences in sensitivity depending on the etiological factor of mastitis; the sensitivity values were 0.25 for Enterococcus spp. and Corynebacterium spp., 0.73 for S. dysgalactiae, and 0.77 for S. aureus. The specificity in these studies ranged from 0.83 for CNS to 1.00 for S. uberis and S. dysgalactiae. These values also differed depending on the number of infected quarters and the lactation phase. This may impede the identification of the etiological factor.
5.2. Physics-Based Methods
Conductivity (mS/cm) and electrical resistance (µΩ) are two opposing but interrelated concepts that describe the properties of materials (include fluids) in relation to the flow of electric current (1 mS = 1.000 µΩ) [100]. Automatic milking systems (AMS) are equipped with sensors measuring milk quality, somatic cell count, and electrical conductivity (EC) [101,102,103]. EC depends on the concentration of anions and cations in milk. Cows with mastitis have an elevated concentration of Na+ and Cl- in their milk, which causes an increase in EC [104]. Norberg et al. [105] noted an EC value of 5.3 mS/cm for healthy, 5.75 for subclinically infected, and 6.73 for clinically infected udders (Table 3). Another classification is as follows: healthy udder (>30 mΩ/cm), suspected mastitis (25–30 mΩ/cm), and mastitis positive (<25 mΩ/cm) [79]. Milner et al. [106] showed that 55% of the CM cases caused by S. aureus and S. uberis can be detected by measuring EC as early as two milking events prior to the onset of overt mastitis symptoms. The increase in EC (defined as a 10% increase compared to the moving average of four previous milking events) was reported to be only associated with infection, indicating its high sensitivity. In a later study, Norberg et al. [105] found that the EC level assisted in the detection of 80.6% of CM cases, 45.0% of SCM cases, and 74.8% of healthy cows. Lukas et al. [107] reported a significant increase in EC three days before the detection of mild mastitis. Only EC measured using a hand-held meter (cow or quarter average) to predict intramammary infection was characterized by relatively low AUC (0.47–0.58), with a simultaneously wide range of values for sensitivity (0.40–0.58) and specificity (0.47–0.98). Explaining these values, it should be noted that factors affecting the electrical conductivity of milk include breed, lactation number and stage, variations in milk composition during milking, and individual animal predisposition [108]. The indicators were corrected using other parameters in the combined model (i.e., last or the maximum SCC before drying off, CMT, inter-quarter ratio of EC (IQR EC) within a cow). The AUC for the combined tests were 0.85 for the last SCC and IQR EC and 0.82 for the highest SCC and IQR EC. As reported by of Fosgate et al. [79], the sensitivity ranged from 0.20 to 0.89, with a high specificity value of 0.81–0.99. The rates were significantly increased by the use of Bayesian latent class analysis, which was further confirmed by the high AUC value (0.91), compared to 0.71 for the classical method. On the other hand, Naqvi et al. [109] used recurrent neural networks, including a range of information extracted from AMS in a validation test, and showed 0.68–0.90 sensitivity and 0.84 specificity. As expected, the test set performance was worse than both the validation and training sets for all metrics. As suggested by Juozaitienė et al. [110], the use of an AMS may facilitate the early diagnosis of mastitis, which allows for the early detection of infected cows at the initial stage of inflammation, enabling the prompt initiation of therapy. The possibility of the on-line recording of data on the EC of milk and the high heritability of this parameter can be used in dairy cattle breeding programs targeted at effective improvement of the productivity and health of cows. As highlighted by these authors, the positive genetic correlation between milk EC and SCC found in their research indicates that it is advisable to monitor changes in the EC of milk in order to diagnose early-stage mastitis in cows.
Table 3.
The diagnostic performance of a tests based on physics methods for detecting dairy cows with intramammary infections.
Infrared thermography (IRT) imaging can be used to diagnose subclinical mastitis. The sensitivity of modern cameras allows the detection of abnormal thermal patterns even in a small area on one of the mammary gland quarters [114]. The thermographic imaging technique and appropriate calculations (predictive modeling) facilitate the effective detection of mastitis at an early stage of the disease [111,114,115,116]. Zaninelli et al. [111] indicated the relatively high sensitivity (0.71–0.79) and specificity (0.72–0.78) of this method, with a relatively high AUC value (0.81). In turn, Velasco-Bolaños et al. [112] considered the problem of the selection of an optimal cut-off value for thermographic measurement and defined it at the level of 32.6 °C for manual milking and 33.7–34.0 °C for automated milking. The value of AUC was shown by these authors to range from 0.70 to 0.90. As suggested by Machado et al. [116], there is a need to standardize the udder area to be measured and analyzed using the software. The authors proposed the anterior udder quarters as the most suitable for recording images and diagnosis of early inflammation, as the correlation coefficients between the temperature of the left and right quarters and the SCC had the highest values, i.e., 0.87 and 0.88, respectively, in comparison with the coefficient of 0.53 for the correlation between the SCC and the temperature of the hindquarters. The effectiveness of thermography in diagnosing mastitis was also indicated by Hovinen et al. [117] and Pezeshki et al. [115]. However, faster diagnosis in these studies was provided by the observations of physical changes in the milk samples and physical examination of the mammary gland [117], as well as measurements of rectal temperature [115]. As suggested by Bortolami et al. [4], IRT may be a useful screening tool for assessment of the severity of mastitis. However, the absence of a significant relationship between the etiological factor and udder temperature indicates a low diagnostic value for thermography in the determination of the etiology of SCM. Another method currently under testing involves the use of data from 3D accelerometer sensors. These sensors are placed on the animal’s ears and serve as tools for measuring resting time, feed intake, rumination, and activity. Grodkowski et al. [113], by constructing artificial neural network and logistic regression models, showed that among cow behaviors, time spent on feed taking and resting appeared to be significant factors for mastitis detection. The effect of mastitis on the cows’ feeding behavior was previously studied by Sepúlveda-Varas [118], who found that cows reduced their feed intake by 1.2 kg DMI/day five days before a CM diagnosis.
Ultrasonography is another helpful non-invasive technique used in the diagnosis of mastitis [119]. However, it should be remembered that it does not replace the traditional methods of identification of pathogens or SCC but provides additional information about the appearance of the parenchyma and teats and helps to determine a likely prognosis for the animal. However, this technique requires extensive experience. The image of a normal bovine mammary gland is moderately echogenic (especially connective tissue and parenchyma). Anechoic areas correspond to the blood vessels and milk ducts [120]. Similarly, the gland cistern is anechoic when distended with milk, while it is surrounded by a hyperechoic mucosal membrane. In contrast, the homogeneous hypoechogenic layer corresponds to muscles. In cases of mastitis, ultrasonographic abnormalities exhibit some variations with the chronicity of the disease and the genus of etiological agent [119,121]. The differences are mainly evident in the appearance of the parenchyma. During infection with Gram-negative bacteria, gas is formed and appears as small shaded hyperechogenic areas. In the case of Arcanobacterium pyogenes, there are multiple spherical regions (approximately 1 cm in diameter) that are more hypoechogenic than the parenchyma and have hyperechogenic centers [121]. Differences can also be seen in the highly echogenic images of milk in cisterns and teats related to increased SCCs [122]. Interestingly, in infections caused by some staphylococcal species, there are no changes in the ultrasonographic images of the parenchyma [119].
5.3. Acute-Phase Proteins
In recent years, the determination of acute-phase proteins (APPs) in blood serum or milk as markers of mammary gland inflammation in cows has been validated. APPs are produced in the liver during an acute inflammatory reaction [123]. The most important APPs in ruminants are haptoglobin (Hp), serum amyloid A (SAA), fibrinogen, and ceruloplasmin [63,124,125]. SAA and Hp were shown to correlate with the SCC and bacteriological test results [126,127]; therefore, SAA and Hp are considered markers with potential diagnostic value [128,129,130,131,132,133,134] (Table 3). They showed high sensitivity both for the measurements in milk from 0.65–0.77 for SAA to 0.82–0.96 for Hp and in serum from 0.90 to 0.74, respectively (Table 4). The specificity was also high and ranged between 0.72 and 0.99 depending on the protein and research material [125,134,135,136,137]. In turn, Addis et al. [62] suggest that cathelicidin is another reliable inflammatory marker. As demonstrated by the authors, its presence is strongly correlated with the SCC. Its potential diagnostic relevance was also highlighted by Smolenski et al. [138] and Cubeddu et al. [139]. Moreover, it is characterized by high values for sensitivity (0.76–0.98) and specificity (0.86–0.99) with relatively high AUC (0.78) and low threshold [62,134]. The high sensitivity (0.98) and specificity (1.00) levels were also obtained using C-reactive protein (CRP) measurement in milk [140]. O’Reilly et al. [141] found that the levels of Hp, MAA, CRP, lactoferrin (LF), α-lactalbumin (LA), and cathelicidin (CATHL) were significantly higher in the milk of diseased cows (p ≤ 0.001). Adopting an assay cut-off for the concentration of the following biomarkers, CRP < 9.5 μg/mL, LF ≥ 325 μg/mL, and MAA < 16 μg/mL, together with the classification tree method, resulted in a sensitivity level of 0.64, a specificity of 0.91, an overall misclassification rate of 0.18, and an AUC of 0.84. As a result, this allowed the differentiation of inflammation caused by Gram-positive from Gram-negative bacteria.
Table 4.
The diagnostic performance of a tests based on acute-phase proteins for detecting dairy cows with intramammary infections.
5.4. Genetic Methods
Currently, methods for the quick identification of pathogenic microorganisms are more frequently based on polymerase chain reaction (PCR) tests. This technique can be applied to test bacteria that have already been isolated from milk or to detect their presence directly in milk. The results of such an analysis can be obtained within ca. 4 h [142].
The latest molecular techniques, such as MALDI-TOF and commercially available quantitative real-time PCR (qPCR) tests, play a crucial role in microbial identification. MALDI-TOF is used to identify the genus or species of isolated organisms, reducing identification errors compared to traditional methods. Commercially available qPCR assays facilitate storage and transport at room temperature and also allow identification of organisms that are difficult to grow, e.g., anerobic bacteria, Mycoplasma spp. [143].
As demonstrated by Wiggans et al. [144], the limited progress in the improvement of udder health with conventional methods and selective procedures based on indirect traits has resulted in an increased need for molecular markers of resistance to mastitis, which should be included in marker-assisted breeding and selection programs. In recent years, many Quantitative Trait Loci (QTLs) associated with bovine resistance to mastitis have been identified. The progress in the investigations of quantitative traits in cattle is documented in the Cattle Quantitative Trait Locus Database [145]. According to current data (June 2024), a total of 2618 QTLs have been associated with the mastitis phenotypes of the SCC, somatic cell score, and CM. These QTLs span all 29 autosomes and the X chromosome and were generated via a combination of traditional linkage analysis, QTL mapping, and genome-wide association studies [145,146,147,148,149,150].
Fang et al. [151] and Kurz et al. [152] conducted research to identify such genomic markers as single nucleotide polymorphisms (SNPs) for genomic traits in order to improve the genomic prognosis of mastitis and immunity traits in cows. There have also been attempts to identify the genes responsible for the ability of the organism to regenerate after mastitis and EPS15L1 (essential for lymphocyte development), PDGFD (involved in macrophage recruitment and wound healing), and PTX3 (involved in regulating inflammation) have been proposed as candidate genes [153]. Abdel-Schafey et al. [154] and Mustafa et al. [155] suggest that the identification of specific single nucleotide polymorphisms (SNPs) associated with mastitis in combination with genomic selection may increase resistance to mastitis in subsequent generations.
The application of these methods represents a significant advancement in the improvement of population resistance to mastitis, particularly in comparison to the pre-genomic era. This approach also accelerates the process relative to traditional genetic methods. However, the issue of heritability remains a concern, especially for mastitis, which is considered to have low heritability. For clinical mastitis in the first lactation, the heritability is estimated to be between 0.02 and 0.03 [156,157] and it is 0.05 for subsequent lactations [156]. The Canadian Dairy Network [158] estimates the heritability of mastitis resistance at a slightly higher level of 0.12. Nevertheless, this trait is largely dependent on management practices and a wide range of environmental factors.
5.5. Metabolomics, Proteomics, and Other Biomarkers
In the near future, the methods with the broadest application for identifying inflammation, but also for identifying etiological factors, will most likely be metabolomics and proteomics. Lisuzzo et al. [159] point to the possibility of using the serum metabolome, finding in animals with subclinical inflammation a lower function of phenylalanine and tyrosine biosynthesis, protein synthesis containing the amino acids phenylalanine, alanine, isoleucine, threonine and tyrosine, and increased synthesis of glycine, serine, histidine, and proline. At the same time, the cited authors showed a different synthesis of glycine, methionine, and leucine-containing proteins when comparing S. agalactiae to Prototheca spp. infections. On the other hand, Dervishi et al. [160] considered valine, serine, tyrosine, and phenylalanine determined 4–8 weeks before parturition and valine, isoleucine, serine, and proline determined 4–8 weeks after parturition as reliable markers of SCM.
Hovinen et al. [161] proposed the detection of udder inflammation with the use of fluorometric tests based on the catalytic activity of N-acetyl-β-d-glucosaminidase (NAGase). This substance releases 4-MU (4-Methylumbelliferone) from the substrate 4-MUAG in an acidic environment. The 4-MU exhibits a blue fluorescence at pH 7.5. The fluorescence intensity of 4-MU is pH dependent and increases to a maximum at pH 10 (plateau above pH 10). The formation of 4-MU can be followed by difference absorption spectrometry at 347 nm or by fluorometric measurements at pH 10, which can be measured to indicate milk NAGase activity. A study conducted by Hiss et al. [136] showed that the SCC was positively correlated (r = 0.76) with the activity of lactate dehydrogenase (LDH) determined directly in the milking parlor with the use of a portable spectrophotometer (Table 5). The use of LDH as an indicator of mastitis resulted in a moderate or relatively high sensitivity (0.61–0.81) and specificity (0.56–0.87) at an AUC value of 0.61–0.88. Similar values were recorded for NAGase [74,136,162]. In the case of LDH, the rates were significantly improved by the inclusion of SCC in the combined model.
Table 5.
The diagnostic performance of tests based on lactate dehydrogenase and N-acetyl-β-d-glucosaminidase and cathelicidin for detecting dairy cows with intramammary infections.
At the same time, simple, fast, and low-cost assays are being tested, i.e., a chemiluminescent bioassay based on magnetite nanoparticles, bioluminescence-based high throughput techniques, cross-linked gold nanoparticles for sensitive detection of haptoglobin, or silver-coated PSi (Ag-PSi) Fabry–Pérot interferometers for quantifying NAGase activity using surface-enhanced Raman spectroscopy (SERS) [18,163,164,165,166]. Serum proteins are also regarded as potential markers of mammary gland inflammation. It has been shown [18,167,168] that an increase in the SCC in milk was accompanied by an increase in the concentration of total protein and globulin in blood serum. Concurrently, high SCC values were accompanied by a decline in albumin concentration and a reduced albumin-to-globulin ratio.
6. Some Management Practices in Mastitis Prevention
For many years, the research on mastitis has comprehensively analyzed the role of the broad-sense farm management. The investigations focused not only on animals but also on their living environments, nutrition, and milking procedures. Importantly, farmers, as well as their knowledge and professional and reliable approach to the problem, were the subject of studies [14].
The variety of mastitis-causing pathogens has increased over the decades in parallel with the changing herd management systems and practices [169]. When control programs were introduced in the 1960s, S. agalactiae and S. aureus were the most common infectious pathogens. Control activities, known as the five-point plan, have proven to be largely successful in reducing the incidence of infectious pathogens, especially S. agalactiae, through the promotion of a widespread use of dry cow therapies and the disinfection of teats.
Prophylaxis, early diagnosis, and management can lead to better treatment of mastitis [26,50]. De Vliegher et al. [14] suggest that improvement of udder health on the farm should be based on shortening the duration of mastitis diagnosis and the reduction of new case frequency. As reported by Ganda et al. [170], the success of mastitis treatment is largely dependent on the type and correct identification of the causative pathogen, which prompts the application of the appropriate therapy.
Milking is one of the most important risk factors for the development of mammary gland inflammation. Appropriate milking does not lead to damage to udder tissues and does not make the mammary gland more susceptible to mastitis. However, any disturbances in the milking process (e.g., irregular vacuum fluctuations, cyclic vacuum fluctuations and very high teat-end vacuum levels, over-milking) are associated with tissue damage that may promote mastitis development and increase the SCC [171,172].
Udders with unevenly developed quarters are most prone to infections. Additionally, inflammations develop more frequently in long teats, which are more susceptible to damage [30]. In relation to health, the most important trait is the udder depth, and cows with pendulous udders are at greater risk of SCM, with an odds ratio (OR) of 1.66 for cows with an udder floor in line with the hock and 2.42 for cows with an udder floor below the hock [173]. Morphometric comparisons demonstrated that the diameter of the teat apex was observed to be a risk factor of CM increase. The OR of this trait was 1.2, which represents a 20% increase in the odds of developing CM for each 1 mm increase in the pre-milking diameter of the apex of the teat. This increased risk of developing mastitis may be related to the wider teat canals and larger teat orifices in wider teats [174]. The teat shape also seems to be important, and cows with flat or round teat ends were 7.6 or 3.2 times more likely to have mastitis than cows with pointed teat ends. This may be explained by the wider streak canals in teats with rounded or flat ends, which have been shown to allow more extensive pathogen penetration [175]. In terms of the risk factors at the level of quarters, a correlation was confirmed between the prevalence of infection and the anatomical position of the quarter, damage to udder skin, condition of the teat canal, and distance from the apex to the base of the teat [176]. A greater frequency of mastitis was observed in hindquarters and in cows with damaged teat skin or with teat orifice hyperkeratosis [177,178].
Poor hygiene during milking and unsatisfactory cleanliness of the hands of milking staff may lead to the development of mastitis. The causative agents are usually transferred from one cow to another during milking [179,180]. Shittu et al. [181] found a lower risk of the occurrence of SCM in herds where hand washing before milking was a common practice in comparison with herds where milking staff did not wash hands (OR = 0.173, p = 0.003). Abrahmsèn et al. [182] confirmed a statistical (p < 0.003) relationship between udder hygiene and the occurrence of SCM. They showed a higher susceptibility for a dirty udder to SCM. The mastitis rate in Holstein-Friesian cows characterized by good udder hygiene was 1.63 quarters/cow, while cows with poor udder hygiene exhibited infection in 2.72 quarters/cow.
The level of hygiene is one of the critical determinants of the occurrence of bacterial infections in the udder [183] leading to SCM and an SCC increase [184]. As demonstrated by Köster et al. [185], the degree of contamination of teats and udders depends on many factors, e.g., type of stall, available space per cow, bedding material and management, and time spent by cows in stalls. Zucali et al. [186] highlight the considerable impact of seasonal and environmental conditions on the cleanliness of cows, e.g., the season of the year and the occurrence of rainfall or snowfall, which make it difficult to keep the beds and traffic alleys clean.
Teat hygiene and the application of pre-milking and post-milking dipping is essential for harvesting high-quality milk with a low level of microbial contamination and a low SCC [187]. As emphasized Huijps et al. [188], proper disinfection of teats before milking can reduce the bacterial count on their surfaces by up to 75% and can be an effective method to control mastitis caused by environmental pathogens. In turn, Sandrucci et al. [187] report that assessment of the condition and hygiene of the teat apex is an easy and quick method providing information about teat health and the level of hyperkeratosis as well as cleanliness of cows in the herd. The information can be used as an indicator of the quality of herd management and welfare.
As demonstrated by Vissio et al. [189], post-milking disinfection of teats, cluster attachment on dry teats, and controlling the pulsation system were associated with a lower SCC in bulk milk, which indicates the prophylactic importance of these measures against new cases of mastitis. Concurrently, the authors reported that the SCC in milk was related to the udder health management rather than to the milking equipment factors. The application of post-milking dipping was strongly associated with a lower SCC in bulk milk, as confirmed by Kelly et al. [190] and Cortinhas et al. [191]. Studies conducted by Kelly et al. [190], Halasa et al. [192], Schewe et al. [193], and Tiwari et al. [194] showed strong relationships between the application of dry cow therapy and a lower SCC in milk, thus proving the suitability of the approach. Mineral supplementation and separate calving pens are also important during this period. It may also be beneficial to use fly control (especially in the pasture) and ruthlessly dispose of waste/discarded milk (Figure 3) [195]. The improvement of cleanliness and thus udder health will also be facilitated by ensuring bedding and animal hygiene by the daily cleaning of the alleyways (at least twice a day), tail and udder trimming (at least twice a year), and disinfection of the stalls [195,196]. The milking procedures and management practices are also extremely important. Of several practices, Huijps et al. [188] identified keeping cows standing after milking, rinsing clusters after milking in each clinical case, using a separate towel for each cow, and wearing milkers’ gloves as the most cost-effective procedures. From this point of view, the management of subclinical cases (milking last and hot water rinsing of clusters) and cleanliness issues (clean stalls and alleyways) were the least effective. These processes were simply expensive to carry out.
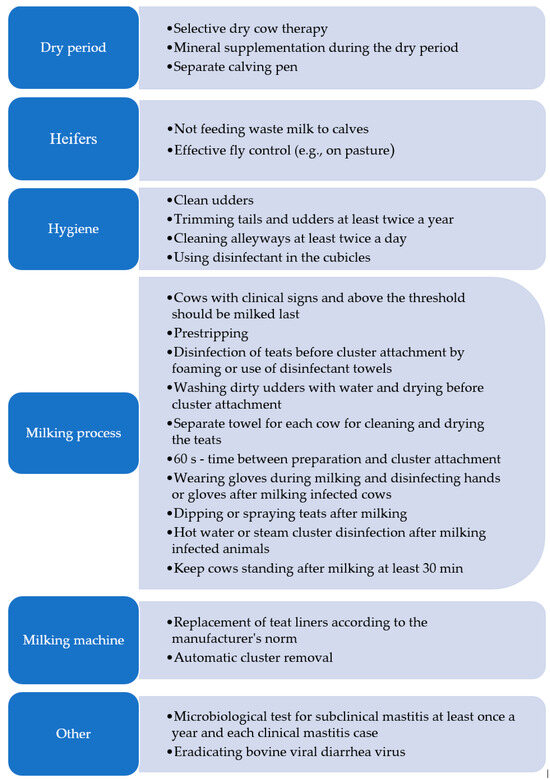
Figure 3.
Prevention and control practices used to reduce mastitis transmission in dairy herd. (own work based on: [50,188,195]).
As suggested Nickerson and Oliver [197], the tangible benefits of proper milking management may be ensured by monitoring the degree of milking in all udder quarters after a decrease in the milk flow and automatic cluster removal. This is intended to prevent over-milking and maintain the appropriate condition of the teats and their apices, which lowers their susceptibility to bacterial colonization and development of mastitis. It was showed [197] that the SCC in milk was significantly lower in herds with automatic cluster removal than in those where no such devices were used. The observance of the teat liner replacement times is also important. Microbiological tests should be performed at least once a year for SCM and for each clinical case. Control and eradication of bovine diarrhea virus is also recommended [50,195].
Replacement of the traditional milking system by an automatic milking system is associated with changes in the preparation of cows for milking. A question arises regarding how the automation of all procedures affects udder health. Salovuo et al. [198] reported that the introduction of such a milking system on farms in Finland was associated with an increase in the total SCC and total bacteria count in milk, which may have been related to technical aspects or improper milking management. This was confirmed by Kolenda et al. [199], who demonstrated a negative impact of AMS on udder health, evidenced by the SCC. The study showed an increase in the SCC in milk in the first year after the introduction of automatic milking on Danish farms compared to the previous year. The increase in the SCC was noted to occur suddenly and was synchronized with the introduction of automatic milking. Similarly, Ivemeyer et al. [200] reported that, compared to conventional milking, automatic milking had a negative effect on udder health and more cases of mastitis were diagnosed. Different results were obtained by Winnicki et al. [201] and Toušová et al. [202], who reported lower SCCs in milk from cows that were milked automatically.
7. Mastitis Immunotherapy and Nanoparticles-Based Treatments
Immunotherapy has gained significant importance in veterinary medicine over the past few decades. In essence, the immune system is manipulated or “fine-tuned” to enhance its natural defense mechanisms. This is achieved by stimulating the immune system to produce specific factors, such as antibodies, lymphocytes, or other immune cells, that are specially designed to target and eliminate harmful substances in the body [203]. Various therapeutic methods are being tested to enhance resistance to mastitis, including vaccinations, T/B cell immunotherapy, RNA immunotherapy, epigenetic immunotherapy, native secretory factors, cytokine therapy, and stem cells therapy [204]. It is important to note that the functioning of the immune system is still being explored and that the unique functions of some of its components are continually being discovered [205].
Since the 1960s, increasingly advanced generations of vaccines have been developed, tested, and commercialized, with the potential to reduce the incidence of mastitis. The focus has been on the three groups of microorganisms causing mastitis that are not effectively controlled by management methods: Gram-negative bacteria (“coliforms”), streptococci, and S. aureus [206]. Currently, in addition to inactivated and modified live vaccines (first generation), the commercially available vaccines also include those based on subunit components and recombinant antigens (second generation). For these vaccines (except live vaccines), a humoral immune response occurs via the MHC-II pathway, which may be insufficient [205,207]. Simultaneous induction of the humoral and cellular responses is possible through live or inactivated chimeric vaccines as well as DNA/RNA-based vaccines. However, these either have not demonstrated sufficient efficacy against mastitis [208,209] or are currently in the testing phase [210,211]. At present, the efficacy of vaccines against the bacteria causing mastitis does not meet expectations. Due to the complexity of the issue and the limited scope of this chapter, we refer to the critical evaluation of mastitis vaccinations and their “immunomechanics” by Rainard et al. [206].
Another tool based on the activation of the immune response is non-specific active cytokine immunotherapy, whose large-scale production has been enabled by recombinant DNA technology. Currently used in adjuvant therapies to support chemotherapy in human medicine, they have also been tested as initiators of immune response in cattle. For this purpose, recombinant granulocyte colony-stimulating factor [6,212,213,214], interleukin-1β and interleukin-2 [215,216,217,218], interleukin-8 [219,220], tumor necrosis factor-α [221], and interferon-gamma [9,222] have been used. Previous studies have shown that the administration of cytokines improves both the humoral and cellular immune responses and may even maintain the increased resistance to infections caused by S. aureus and E. coli. However, it should be emphasized that the administration of external cytokines merely mimics the host’s normal response to an infectious agent [216]. In combination with other therapies, they can enhance their efficacy [215], particularly in the cases of mastitis in individuals with weakened immune systems [204].
Although there are currently no reports on the typical application of T/B lymphocyte immunotherapy for mastitis, the relationships in this area are known. T lymphocytes are an integral part of the mechanism for most immunotherapies clinically approved for human treatment. Dynamic changes are observed in the ratio of T CD4:CD8 lymphocytes and effector T cells (Teff) to regulatory T cells (Treg) [11,223]. Conversely, B cells play a crucial role in activating the host’s innate immunity, enabling the body to combat infections independently [224]. Since the relationships between CD4/CD8 and B lymphocytes appear to be pathogen dependent, we refer to the review by Rainard et al. [12] due to the complexity of the topic. In conclusion, following these authors, we suggest that resuming research on immune cells inhabiting healthy, infected, or immunized udders seems to be the most promising approach for designing effective immunotherapies against mastitis.
It has been found that egg yolk immunoglobulins (IgY) from hens previously vaccinated with pathogenic strains of bovine mastitis, S. aureus, or E. coli killed with formaldehyde demonstrated phagocytic activity against these pathogens in in vitro studies [225,226]. For S. aureus, these results were confirmed also in vivo [227]. Another example is the study by Almeida et al. [228], which utilized anti-recombinant antibodies against the adhesive molecules of S. uberis. These antibodies prevented the adhesion and internalization of mammary gland cells, as confirmed by lower SCCs, bacterial colony numbers, and infected quarters.
A highly promising area in mastitis immunotherapy is the regeneration of mammary gland tissues. Contemporary regenerative medicine leverages the potential of stem cells to improve a wide spectrum of disease conditions in both humans and animals. Particular emphasis is placed on mesenchymal stem cells (MSCs), multipotent progenitor cells found in various tissues, including adipose [229]. Research indicates that MSCs are similar to pericytes, which are perivascular cells capable of detaching from blood vessels in response to tissue damage or inflammation [230]. Studies suggest that bovine adipose-tissue-derived MSCs may play a crucial role in controlling various infectious conditions, including bovine mastitis [231].
Extracellular vesicles also hold immunotherapeutic potential. In the context of mastitis therapy, scientific efforts have focused on the exosomes derived from milk, mammary epithelial cells, blood, and urine. It has been determined that the exosomal miRNA expression profile changes during inflammation and that high expression of certain miRNAs may serve as potential markers for the subclinical mastitis caused by S. aureus [232]. On the other hand, milk exosomes, being safe in terms of immunogenicity and cytotoxicity [233,234,235] and capable of being targeted [236], can be used as natural nanoparticles for the delivery of short interfering RNA (siRNA), miRNA, and drugs [237,238].
Nanoparticles can be valuable allies fighting against drug-resistant microorganisms due to their enhanced antibacterial efficacy, which facilitates the active uptake of substances by phagocytes [239,240]. Studies using nanoparticles of silver and copper [241], propolis [242], chitosan [243], nisin [240], and tilmicosin [244] have demonstrated inhibitory effects on certain etiological agents of mastitis under in vitro conditions, which is a promising indicator for further in vivo trials.
8. Conclusions and Future Perspectives
Mastitis is globally recognized as the most costly disease affecting dairy cattle and it poses significant challenges to management through zootechnical and veterinary methods. This is due to a wide range of causative factors, such as the susceptibility of cows to infections, maintenance conditions, risk factors for infection development, and antibiotic resistance. The reviewed literature indicates that while there are many diagnostic methods for mastitis, there is still a need to develop the new tools. Further research is necessary to establish objective detection parameters and prognostic indicators. It is desirable to implement tests that demonstrate both high sensitivity and high specificity. A highly sensitive diagnostic test gives few false-negative results, thus missing fewer positive cases. Conversely, a test with high specificity gives fewer false-positive results. Statistical tools that consider various variables, such as artificial neural networks and Bayesian latent class analysis may be used to improve test accuracy.
The challenge in the coming years will be to develop universal diagnostic and therapeutic strategies/techniques that can be considered and implemented as effective alternatives to antibiotic therapy. Future research should focus on developing preventive and therapeutic techniques (e.g., immunotherapies, bacteriocins, herbal therapy, nanoparticle technology).
Author Contributions
Conceptualization, P.S., P.Ż. and E.J.; project administration, P.Ż. and E.J.; visualization, P.Ż. and E.J.; writing—original draft preparation, P.S.; writing—review and editing, P.Ż. and E.J. All authors have read and agreed to the published version of the manuscript.
Funding
Publication financed by a statutory activity subsidy from the Polish Ministry of Science and Higher Education for Faculty of Animal Sciences and Bioeconomy University of Life Sciences in Lublin: SUBB.WZB.19.065.ZiR.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hagnestam, C.; Emanuelson, U.; Berglund, B. Yield Losses Associated with Clinical Mastitis Occurring in Different Weeks of Lactation. J. Dairy Sci. 2007, 90, 2260–2270. [Google Scholar] [CrossRef]
- Hussain, R.; Javed, M.T.; Khan, A. Changes in Some Biochemical Parameters and Somatic Cell Counts in the Milk of Buffalo and Cattle Suffering from Mastitis. Pak. Vet. J. 2012, 32, 418–421. [Google Scholar]
- Lipkens, Z.; Piepers, S.; De Visscher, A.; De Vliegher, S. Evaluation of Test-Day Milk Somatic Cell Count Information to Predict Intramammary Infection with Major Pathogens in Dairy Cattle at Drying Off. J. Dairy Sci. 2019, 102, 4309–4321. [Google Scholar] [CrossRef]
- Bortolami, A.; Fiore, E.; Gianesella, M.; Corrò, M.; Catania, S.; Morgante, M. Evaluation of the Udder Health Status in Subclinical Mastitis Affected Dairy Cows through Bacteriological Culture, Somatic Cell Count and Thermographic Imaging. Pol. J. Vet. Sci. 2015, 18, 104. [Google Scholar] [CrossRef]
- Senft, B.; Neudecker, J. Defense Mechanisms of the Bovine Mammary Gland. Tierärztliche Prax. 1991, 19, 357–363. [Google Scholar]
- Nickerson, S.C. Immunological Aspects of Mammary Involution. J. Dairy Sci. 1989, 72, 1665–1678. [Google Scholar] [CrossRef]
- Cobirka, M.; Tancin, V.; Slama, P. Epidemiology and Classification of Mastitis. Animals 2020, 10, 2212. [Google Scholar] [CrossRef]
- Bronzo, V.; Lopreiato, V.; Riva, F.; Amadori, M.; Curone, G.; Addis, M.F.; Cremonesi, P.; Moroni, P.; Trevisi, E.; Castiglioni, B. The Role of Innate Immune Response and Microbiome in Resilience of Dairy Cattle to Disease: The Mastitis Model. Animals 2020, 10, 1397. [Google Scholar] [CrossRef]
- Pighetti, G.M.; Sordillo, L.M. Specific Immune Responses of Dairy Cattle after Primary Inoculation with Recombinant Bovine Interferon-γ as an Adjuvant When Vaccinating against Mastitis. Am. J. Vet. Res. 1996, 57, 819–824. [Google Scholar] [CrossRef]
- Piotrowska-Tomala, K.K.; Siemieniuch, M.J.; Szóstek, A.Z.; Korzekwa, A.J.; Woclawek-Potocka, I.; Galváo, A.M.; Okuda, K.; Skarzynski, D.J. Lipopolysaccharides, Cytokines, and Nitric Oxide Affect Secretion of Prostaglandins and Leukotrienes by Bovine Mammary Gland Epithelial Cells. Domest. Anim. Endocrinol. 2012, 43, 278–288. [Google Scholar] [CrossRef]
- Mishra, S. CD8+ Regulatory T Cell—A Mystery to Be Revealed. Front. Immunol. 2021, 12, 8874. [Google Scholar] [CrossRef]
- Rainard, P.; Foucras, G.; Martins, R.P. Adaptive Cell-Mediated Immunity in the Mammary Gland of Dairy Ruminants. Front. Vet. Sci. 2022, 9, 4890. [Google Scholar] [CrossRef]
- Pegolo, S.; Tessari, R.; Bisutti, V.; Vanzin, A.; Giannuzzi, D.; Gianesella, M.; Lisuzzo, A.; Fiore, E.; Barberio, A.; Schiavon, E.; et al. Quarter-Level Analyses of the Associations among Subclinical Intramammary Infection and Milk Quality, Udder Health, and Cheesemaking Traits in Holstein Cows. J. Dairy Sci. 2022, 105, 3490–3507. [Google Scholar] [CrossRef]
- De Vliegher, S.; Ohnstad, I.; Piepers, S. Management and Prevention of Mastitis: A Multifactorial Approach with a Focus on Milking, Bedding and Data-Management. J. Integr. Agric. 2018, 17, 1214–1233. [Google Scholar] [CrossRef]
- Ashraf, A.; Imran, M. Causes, Types, Etiological Agents, Prevalence, Diagnosis, Treatment, Prevention, Effects on Human Health and Future Aspects of Bovine Mastitis. Anim. Health Res. Rev. 2020, 21, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, M.; Tantary, H. A Treatise on Bovine Mastitis: Disease and Disease Economics, Etiological Basis, Risk Factors, Impact on Human Health, Therapeutic Management, Prevention and Control Strategy. Adv. Dairy Res. 2015, 4, 150. [Google Scholar] [CrossRef]
- Martins, S.A.M.; Martins, V.C.; Cardoso, F.A.; Germano, J.; Rodrigues, M.; Duarte, C.; Bexiga, R.; Cardoso, S.; Freitas, P.P. Biosensors for On-Farm Diagnosis of Mastitis. Front. Bioeng. Biotechnol. 2019, 7, 186. [Google Scholar] [CrossRef] [PubMed]
- Pyörälä, S. Indicators of Inflammation in the Diagnosis of Mastitis. Vet. Res. 2003, 34, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan — a Web and Mobile App for Systematic Reviews. Available online: https://rayyan.ai/ (accessed on 1 August 2024).
- Peters, M.D.P.; Silveira, I.D.B.; Fischer, V. Impact of Subclinical and Clinical Mastitis on Sensitivity to Pain of Dairy Cows. Animal 2015, 9, 2024–2028. [Google Scholar] [CrossRef]
- Ferreira, G.M.; Petzer, I.M. Injectable Organic and Inorganic Selenium in Dairy Cows—Effects on Milk, Blood and Somatic Cell Count Levels. Onderstepoort J. Vet. Res. 2019, 86, 1664. [Google Scholar] [CrossRef]
- Cheng, W.N.; Han, S.G. Bovine Mastitis: Risk Factors, Therapeutic Strategies, and Alternative Treatments—A Review. Asian-Australas J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.L.; Theil, P.K. Perspectives on Immunoglobulins in Colostrum and Milk. Nutrients 2011, 3, 442–474. [Google Scholar] [CrossRef] [PubMed]
- Kibebew, K. Bovine Mastitis: A Review of Causes and Epidemiological Point of View. J. Biol. Agric. Healthc. 2017, 7, 1–14. [Google Scholar]
- Bian, Y.; Lv, Y.; Li, Q. Identification of Diagnostic Protein Markers of Subclinical Mastitis in Bovine Whey Using Comparative Proteomics. Bull. Vet. Inst. Pulawy 2014, 58, 385–392. [Google Scholar] [CrossRef]
- Griffioen, K.; Velthuis, A.G.J.; Koop, G.; Lam, T.J.G.M. Effects of a Mastitis Treatment Strategy with or without On-Farm Testing. J. Dairy Sci. 2021, 104, 4665–4681. [Google Scholar] [CrossRef] [PubMed]
- Juronen, D.; Kuusk, A.; Kivirand, K.; Rinken, A.; Rinken, T. Immunosensing System for Rapid Multiplex Detection of Mastitis-Causing Pathogens in Milk. Talanta 2018, 178, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, J.R.; Gonçalves, J.L.; Grenfell, R.; Leite, R.F.; Juliano, L.; Santos, M.V. Direct Identification of Bovine Mastitis Pathogens by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry in Pre-Incubated Milk. Braz. J. Microbiol. 2018, 49, 801–807. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.; Awad, W.; Abdou, N.E.; Castañeda Vázquez, H. Molecular Biological Tools Applied for Identification of Mastitis Causing Pathogens. Int. J. Vet. Sci. Med. 2017, 5, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Awale, M.; Dudhatra, G.B.; Avinash, K.; Chauhan, B.N.; Kamani, D.R.; Modi, C.M.; Patel, H.B.; Mody, S.K. Bovine Mastitis: A Threat to Economy. Open Access Sci. Rep. 2012, 1, 1–10. Available online: https://www.omicsonline.org/scientific-reports/srep295.php (accessed on 28 May 2024).
- National Mastitis Council Inc. Laboratory Handbook on Bovine Mastitis, 3rd ed.; National Mastitis Council Inc.: New Prague, MN, USA, 2017. [Google Scholar]
- Kuang, Y.; Tani, K.; Synnott, A.J.; Ohshima, K.; Higuchi, H.; Nagahata, H.; Tanji, Y. Characterization of Bacterial Population of Raw Milk from Bovine Mastitis by Culture-Independent PCR-DGGE Method. Biochem. Eng. J. 2009, 45, 76–81. [Google Scholar] [CrossRef]
- Contreras, G.A.; Rodríguez, J.M. Mastitis: Comparative Etiology and Epidemiology. J. Mammary Gland Biol. Neoplasia 2011, 16, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Klaas, I.C.; Zadoks, R.N. An Update on Environmental Mastitis: Challenging Perceptions. Transbound. Emerg. Dis. 2018, 65, 166–185. [Google Scholar] [CrossRef] [PubMed]
- Saidani, M.; Messadi, L.; Soudani, A.; Daaloul-Jedidi, M.; Châtre, P.; Ben Chehida, F.; Mamlouk, A.; Mahjoub, W.; Madec, J.Y.; Haenni, M. Epidemiology, Antimicrobial Resistance, and Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Clinical Bovine Mastitis in Tunisia. Microb. Drug Resist. 2018, 24, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Heikkilä, A.M.; Liski, E.; Pyörälä, S.; Taponen, S. Pathogen-Specific Production Losses in Bovine Mastitis. J. Dairy Sci. 2018, 101, 9493–9504. [Google Scholar] [CrossRef]
- Zi, C.; Zeng, D.; Ling, N.; Dai, J.; Xue, F.; Jiang, Y.; Li, B. An Improved Assay for Rapid Detection of Viable Staphylococcus Aureus Cells by Incorporating Surfactant and PMA Treatments in QPCR. BMC Microbiol. 2018, 18, 1–8. [Google Scholar] [CrossRef]
- Milanov, D.; Prunić, B.; Velhner, M.; Bojkovski, J. Diagnosis of Yeast Mastitis in Dairy Cows. Lucr. Stiint.—Univ. Stiint. Agric. A Banat. Timis. Med. Vet. 2014, 47, 56–64. [Google Scholar]
- Abou-Elmagd, S.; Kotb, H.; Sabry, K.; Refai, M. Prevalence of Candida Albicans and Cryptococcus Neoformans in Animals and Chickens in Quena Governorate, with Special Reference to RAPD-PCR Patterns of the Isolates. J. Am. Sci. 2011, 7, 20–31. [Google Scholar]
- Da Costa, G.M.; de Pereira, U.P.; Gomes Souza-Dias, M.A.; da Silva, N. Yeast Mastitis Outbreak in a Brazilian Dairy Herd. Braz. J. Vet. Res. Anim. Sci. 2012, 49, 239–243. [Google Scholar] [CrossRef][Green Version]
- Tarazona-Manrique, L.E.; Villate-Hernández, J.R.; Andrade-Becerra, R.J. Bacterial and Fungal Infectious Etiology Causing Mastitis in Dairy Cows in the Highlands of Boyacá (Colombia). Rev. Fac. Med. Vet. Zootec. 2019, 66, 208–218. [Google Scholar] [CrossRef]
- Jagielski, T.; Krukowski, H.; Bochniarz, M.; Piech, T.; Roeske, K.; Bakuła, Z.; Wlazło, Ł.; Woch, P. Prevalence of Prototheca Spp. on Dairy Farms in Poland—A Cross-Country Study. Microb. Biotechnol. 2019, 12, 556–566. [Google Scholar] [CrossRef]
- Shkromada, O.; Palii, A.; Palii, A.; Skliar, O.; Dudchenko, Y.; Necherya, T. Improvement of Milk Quality for Micro-Climate Formation on Cattle Farms. Bull. Sumy Natl. Agrar. Univ. Ser. Vet. Med. 2019, 4, 43–49. [Google Scholar] [CrossRef]
- Bezman, D.; Lemberskiy-Kuzin, L.; Katz, G.; Merin, U.; Leitner, G. Influence of Intramammary Infection of a Single Gland in Dairy Cows on the Cow’s Milk Quality. J. Dairy Res. 2015, 82, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Leitner, G.; Lavon, Y. The Disruptive Effects of Mastitis on Reproduction and Fertility in Dairy Cows. Ital. J. Anim. Sci. 2015, 14, 650–654. [Google Scholar] [CrossRef]
- Van Soest, F.J.S.; Santman-Berends, I.M.G.A.; Lam, T.J.G.M.; Hogeveen, H. Failure and Preventive Costs of Mastitis on Dutch Dairy Farms. J. Dairy Sci. 2016, 99, 8365–8374. [Google Scholar] [CrossRef] [PubMed]
- Huijps, K.; Lam, T.J.G.M.; Hogeveen, H. Costs of Mastitis: Facts and Perception. J. Dairy Res. 2008, 75, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, P.; Goudar, A.L.; Suresh, K.P.; Roy, P. Global and Countrywide Prevalence of Subclinical and Clinical Mastitis in Dairy Cattle and Buffaloes by Systematic Review and Meta-Analysis. Res. Vet. Sci. 2021, 136, 561–586. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadi, M.; Haine, D.; Kelton, D.F.; Barkema, H.W.; Hogeveen, H.; Keefe, G.P.; Dufour, S. Herd-Level Mastitis-Associated Costs on Canadian Dairy Farms. Front. Vet. Sci. 2018, 5, 100. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; Piepers, S.; De Vliegher, S. Mastitis Prevention and Control Practices and Mastitis Treatment Strategies Associated with the Consumption of (Critically Important) Antimicrobials on Dairy Herds in Flanders, Belgium. J. Dairy Sci. 2016, 99, 2896–2903. [Google Scholar] [CrossRef]
- Cha, E.; Bar, D.; Hertl, J.A.; Tauer, L.W.; Bennett, G.; González, R.N.; Schukken, Y.H.; Welcome, F.L.; Gröhn, Y.T. The Cost and Management of Different Types of Clinical Mastitis in Dairy Cows Estimated by Dynamic Programming. J. Dairy Sci. 2011, 94, 4476–4487. [Google Scholar] [CrossRef]
- Liang, D.; Arnold, L.M.; Stowe, C.J.; Harmon, R.J.; Bewley, J.M. Estimating US Dairy Clinical Disease Costs with a Stochastic Simulation Model. J. Dairy Sci. 2017, 100, 1472–1486. [Google Scholar] [CrossRef]
- Rollin, E.; Dhuyvetter, K.C.; Overton, M.W. The Cost of Clinical Mastitis in the First 30 Days of Lactation: An Economic Modeling Tool. Prev. Vet. Med. 2015, 122, 257–264. [Google Scholar] [CrossRef]
- Gonçalves, J.L.; Tomazi, T.; Barreiro, J.R.; Beuron, D.C.; Arcari, M.A.; Lee, S.H.I.; de Martins, C.M.M.R.; Araújo Junior, J.P.; Santos, M.V. dos Effects of Bovine Subclinical Mastitis Caused by Corynebacterium Spp. on Somatic Cell Count, Milk Yield and Composition by Comparing Contralateral Quarters. Vet. J. 2016, 209, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.L.; Kamphuis, C.; Martins, C.M.M.R.; Barreiro, J.R.; Tomazi, T.; Gameiro, A.H.; Hogeveen, H.; dos Santos, M.V. Bovine Subclinical Mastitis Reduces Milk Yield and Economic Return. Livest. Sci. 2018, 210, 25–32. [Google Scholar] [CrossRef]
- Bobbo, T.; Ruegg, P.L.; Stocco, G.; Fiore, E.; Gianesella, M.; Morgante, M.; Pasotto, D.; Bittante, G.; Cecchinato, A. Associations between Pathogen-Specific Cases of Subclinical Mastitis and Milk Yield, Quality, Protein Composition, and Cheese-Making Traits in Dairy Cows. J. Dairy Sci. 2017, 100, 4868–4883. [Google Scholar] [CrossRef] [PubMed]
- Malek dos Reis, C.B.; Barreiro, J.R.; Mestieri, L.; de Porcionato, M.A.F.; dos Santos, M.V. Effect of Somatic Cell Count and Mastitis Pathogens on Milk Composition in Gyr Cows. BMC Vet. Res. 2013, 9, 67. [Google Scholar] [CrossRef]
- Batavani, R.A.; Asri, S.; Naebzadeh, H. The Effect of Subclinical Mastitis on Milk Composition in Dairy Cows. Iran. J. Vet. Res. 2007, 8, 205–211. [Google Scholar]
- Power, M.; Fell, G.; Wright, M. Principles for High-Quality, High-Value Testing. Evid. Based. Med. 2013, 18, 5–10. [Google Scholar] [CrossRef]
- Damm, M.; Holm, C.; Blaabjerg, M.; Bro, M.N.; Schwarz, D. Differential Somatic Cell Count—A Novel Method for Routine Mastitis Screening in the Frame of Dairy Herd Improvement Testing Programs. J. Dairy Sci. 2017, 100, 4926–4940. [Google Scholar] [CrossRef]
- Martin, P.; Barkema, H.W.; Brito, L.F.; Narayana, S.G.; Miglior, F. Symposium Review: Novel Strategies to Genetically Improve Mastitis Resistance in Dairy Cattle. J. Dairy Sci. 2018, 101, 2724–2736. [Google Scholar] [CrossRef] [PubMed]
- Addis, M.F.; Tedde, V.; Puggioni, G.M.G.; Pisanu, S.; Casula, A.; Locatelli, C.; Rota, N.; Bronzo, V.; Moroni, P.; Uzzau, S. Evaluation of Milk Cathelicidin for Detection of Bovine Mastitis. J. Dairy Sci. 2016, 99, 8250–8258. [Google Scholar] [CrossRef]
- Viguier, C.; Arora, S.; Gilmartin, N.; Welbeck, K.; O’Kennedy, R. Mastitis Detection: Current Trends and Future Perspectives. Trends Biotechnol. 2009, 27, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Adkins, P.R.F.; Middleton, J.R. Methods for Diagnosing Mastitis. Vet. Clin. North Am.—Food Anim. Pract. 2018, 34, 479–491. [Google Scholar] [CrossRef]
- Erdbrügger, U.; Rudy, C.K.; Etter, E.M.; Dryden, K.A.; Yeager, M.; Klibanov, A.L.; Lannigan, J. Imaging Flow Cytometry Elucidates Limitations of Microparticle Analysis by Conventional Flow Cytometry. Cytom. Part A 2014, 85, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.F.; Uniacke-Lowe, T.; McSweeney, P.L.H.; O’Mahony, J.A. Dairy Chemistry and Biochemistry, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2015; ISBN 9783319148922. [Google Scholar]
- Langerhuus, S.N.; Ingvartsen, K.L.; Bennedsgaard, T.W.; Røntved, C.M. Gram-Typing of Mastitis Bacteria in Milk Samples Using Flow Cytometry. J. Dairy Sci. 2013, 96, 267–277. [Google Scholar] [CrossRef]
- Rowe, S.; Godden, S.; Nydam, D.V.; Gorden, P.; Lago, A.; Vasquez, A.; Royster, E.; Timmerman, J.; Thomas, M. Evaluation of Rapid Culture, a Predictive Algorithm, Esterase Somatic Cell Count and Lactate Dehydrogenase to Detect Intramammary Infection in Quarters of Dairy Cows at Dry-Off. Prev. Vet. Med. 2020, 179, 104982. [Google Scholar] [CrossRef]
- Pantoja, J.C.F.; Hulland, C.; Ruegg, P.L. Dynamics of Somatic Cell Counts and Intramammary Infections across the Dry Period. Prev. Vet. Med. 2009, 90, 43–54. [Google Scholar] [CrossRef]
- Ferronatto, J.A.; Ferronatto, T.C.; Schneider, M.; Pessoa, L.F.; Blagitz, M.G.; Heinemann, M.B.; Della Libera, A.M.M.P.; Souza, F.N. Diagnosing Mastitis in Early Lactation: Use of Somaticell®, California Mastitis Test and Somatic Cell Count. Ital. J. Anim. Sci. 2018, 17, 723–729. [Google Scholar] [CrossRef]
- Gohary, K.; McDougall, S. Predicting Intramammary Infection Status at Drying off Using Indirect Testing of Milk Samples. N. Z. Vet. J. 2018, 66, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Santschi, D.E.; Durocher, J.; Lefebvre, D.M. Evaluation of the New Differential Somatic Cell Count Parameter as a Rapid and Inexpensive Supplementary Tool for Udder Health Management through Regular Milk Recording. Prev. Vet. Med. 2020, 181, 105079. [Google Scholar] [CrossRef]
- Schwarz, D.; Lipkens, Z.; Piepers, S.; De Vliegher, S. Investigation of Differential Somatic Cell Count as a Potential New Supplementary Indicator to Somatic Cell Count for Identification of Intramammary Infection in Dairy Cows at the End of the Lactation Period. Prev. Vet. Med. 2019, 172, 104803. [Google Scholar] [CrossRef]
- Khatun, M.; Thomson, P.C.; García, S.C.; Bruckmaier, R.M. Suitability of Milk Lactate Dehydrogenase and Serum Albumin for Pathogen-Specific Mastitis Detection in Automatic Milking Systems. J. Dairy Sci. 2022, 105, 2558–2571. [Google Scholar] [CrossRef]
- Bludau, M.J.; Maeschli, A.; Leiber, F.; Steiner, A.; Klocke, P. Mastitis in Dairy Heifers: Prevalence and Risk Factors. Vet. J. 2014, 202, 566–572. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.G.A.; Olde Riekerink, R.G.M.; Sampimon, O.C.; van Schaik, G.; Lam, T.J.G.M. Incidence of Subclinical Mastitis in Dutch Dairy Heifers in the First 100 Days in Lactation and Associated Risk Factors. J. Dairy Sci. 2012, 95, 2476–2484. [Google Scholar] [CrossRef]
- Kandeel, S.A.; Megahed, A.A.; Ebeid, M.H.; Constable, P.D. Evaluation of 3 Esterase Tests for the Diagnosis of Subclinical Mastitis at Dry-off and Freshening in Dairy Cattle. J. Dairy Sci. 2019, 102, 1402–1416. [Google Scholar] [CrossRef] [PubMed]
- Bhutto, A.L.; Murray, R.D.; Woldehiwet, Z. California Mastitis Test Scores as Indicators of Subclinical Intra-Mammary Infections at the End of Lactation in Dairy Cows. Res. Vet. Sci. 2012, 92, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Fosgate, G.T.; Petzer, I.M.; Karzis, J. Sensitivity and Specificity of a Hand-Held Milk Electrical Conductivity Meter Compared to the California Mastitis Test for Mastitis in Dairy Cattle. Vet. J. 2013, 196, 98–102. [Google Scholar] [CrossRef]
- Dingwell, R.T.; Leslie, K.E.; Schukken, Y.H.; Sargeant, J.M.; Timms, L.L. Evaluation of the California Mastitis Test to Detect an Intramammary Infection with a Major Pathogen in Early Lactation Dairy Cows. Can. Vet. J. Rev. Vet. Can. 2003, 44, 413–415. [Google Scholar]
- Sears, P.M.; Wilson, D.J.; Gonzalez, R.N.; Hancock, D.D. Microbiological Results from Milk Samples Obtained Premilking and Postmilking for the Diagnosis of Bovine Lntramammary Infections. J. Dairy Sci. 1991, 74, 4183–4188. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M.; Jansen, J.T.; Leslie, K.E.; Smart, N.L.; Kelton, D.F. The Effect of Sampling Time and Sample Handling on the Detection of Staphylococcus Aureus in Milk from Quarters with Subclinical Mastitis. Can. Vet. J. 2002, 43, 38. [Google Scholar]
- Souza, F.N.; Cunha, A.F.; Rosa, D.L.S.O.; Brito, M.A.V.; Guimarães, A.S.; Mendonça, L.C.; Souza, G.N.; Lage, A.P.; Blagitz, M.G.; Della Libera, A.M.M.P.; et al. Somatic Cell Count and Mastitis Pathogen Detection in Composite and Single or Duplicate Quarter Milk Samples. Pesqui. Vet. Bras. 2016, 36, 811–818. [Google Scholar] [CrossRef]
- Reyher, K.K.; Dohoo, I.R. Diagnosing Intramammary Infections: Evaluation of Composite Milk Samples to Detect Intramammary Infections. J. Dairy Sci. 2011, 94, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Kleinhans, S.; Reimann, G.; Stückler, P.; Reith, F.; Ilves, K.; Pedastsaar, K.; Yan, L.; Zhang, Z.; Valdivieso, M.; et al. Investigation of Dairy Cow Performance in Different Udder Health Groups Defined Based on a Combination of Somatic Cell Count and Differential Somatic Cell Count. Prev. Vet. Med. 2020, 183, 105123. [Google Scholar] [CrossRef]
- Pilla, R.; Malvisi, M.; Snel, G.G.M.; Schwarz, D.; König, S.; Czerny, C.P.; Piccinini, R. Differential Cell Count as an Alternative Method to Diagnose Dairy Cow Mastitis. J. Dairy Sci. 2013, 96, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Robles, I.; Nolan, D.T.; Fendley, C.A.; Stokley, H.L.; France, T.L.; Ferrell, J.L.; Costa, J.H.C. Technical Note: Evaluation of a Commercial on-Farm Milk Leukocyte Differential Tester to Identify Subclinical Mastitis Cases in Dairy Cows. J. Dairy Sci. 2021, 104, 4942–4949. [Google Scholar] [CrossRef] [PubMed]
- Scherpenzeel, C.G.M.; den Uijl, I.E.M.; van Schaik, G.; Riekerink, R.G.M.O.; Hogeveen, H.; Lam, T.J.G.M. Effect of Different Scenarios for Selective Dry-Cow Therapy on Udder Health, Antimicrobial Usage, and Economics. J. Dairy Sci. 2016, 99, 3753–3764. [Google Scholar] [CrossRef] [PubMed]
- Vanhoudt, A.; van Hees-Huijps, K.; van Knegsel, A.T.M.; Sampimon, O.C.; Vernooij, J.C.M.; Nielen, M.; van Werven, T. Effects of Reduced Intramammary Antimicrobial Use during the Dry Period on Udder Health in Dutch Dairy Herds. J. Dairy Sci. 2018, 101, 3248–3260. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.J.G.M.; Olde Riekerink, R.G.M.; Sampimon, O.C.; Smith, H. Mastitis Diagnostics and Performance Monitoring: A Practical Approach. Ir. Vet. J. 2009, 62, 34–39. [Google Scholar] [CrossRef]
- Maćešić, N.; Bačić, G.; Božičević, K.; Benić, M.; Karadjole, T.; Babić, N.P.; Lojkić, M.; Efendić, M.; Bačić, I.; Pavlak, M. Assessment of the Zagreb Mastitis Test in Diagnosis of Subclinical Mastitis in Dairy Cattle. Vet. Arh. 2016, 86, 475–485. [Google Scholar]
- Sargeant, J.M.; Leslie, K.E.; Shirley, J.E.; Pulkrabek, B.J.; Lim, G.H. Sensitivity and Specificity of Somatic Cell Count and California Mastitis Test for Identifying Intramammary Infection in Early Lactation. J. Dairy Sci. 2001, 84, 2018–2024. [Google Scholar] [CrossRef]
- Halasa, T.; Kirkeby, C. Differential Somatic Cell Count: Value for Udder Health Management. Front. Vet. Sci. 2020, 7, 9055. [Google Scholar] [CrossRef]
- Zecconi, A.; Vairani, D.; Cipolla, M.; Rizzi, N.; Zanini, L. Assessment of Subclinical Mastitis Diagnostic Accuracy by Differential Cell Count in Individual Cow Milk. Ital. J. Anim. Sci. 2019, 18, 460–465. [Google Scholar] [CrossRef]
- Pilla, R.; Schwarz, D.; König, S.; Piccinini, R. Microscopic Differential Cell Counting to Identify Inflammatory Reactions in Dairy Cow Quarter Milk Samples. J. Dairy Sci. 2012, 95, 4410–4420. [Google Scholar] [CrossRef]
- Græsbøll, K.; Kirkeby, C.; Nielsen, S.S.; Halasa, T.; Toft, N.; Christiansen, L.E. Models to Estimate Lactation Curves of Milk Yield and Somatic Cell Count in Dairy Cows at the Herd Level for the Use in Simulations and Predictive Models. Front. Vet. Sci. 2016, 3, 115. [Google Scholar] [CrossRef] [PubMed]
- Kirkeby, C.; Toft, N.; Schwarz, D.; Farre, M.; Nielsen, S.S.; Zervens, L.; Hechinger, S.; Halasa, T. Differential Somatic Cell Count as an Additional Indicator for Intramammary Infections in Dairy Cows. J. Dairy Sci. 2020, 103, 1759–1775. [Google Scholar] [CrossRef] [PubMed]
- Zecconi, A.; Zanini, L.; Cipolla, M.; Stefanon, B. Factors Affecting the Patterns of Total Amount and Proportions of Leukocytes in Bovine Milk. Animals 2020, 10, 992. [Google Scholar] [CrossRef]
- Wall, S.K.; Wellnitz, O.; Bruckmaier, R.M.; Schwarz, D. Differential Somatic Cell Count in Milk before, during, and after Lipopolysaccharide- and Lipoteichoic-Acid-Induced Mastitis in Dairy Cows. J. Dairy Sci. 2018, 101, 5362–5373. [Google Scholar] [CrossRef]
- Thyarla, A.; Agrawal, A.K.; Sandey, K.K.; Suresh, V.; Prasanth, B.; Umapathy, K.S.; Sinha, G. Study of Physical and Electrical Characteristics of Skim Milk. Asian J. Dairy Food Res. 2018, 37, 18–21. [Google Scholar] [CrossRef]
- Hovinen, M.; Pyörälä, S. Invited Review: Udder Health of Dairy Cows in Automatic Milking. J. Dairy Sci. 2011, 94, 547–562. [Google Scholar] [CrossRef]
- Khatun, M.; Thomson, P.C.; Kerrisk, K.L.; Lyons, N.A.; Clark, C.E.F.; Molfino, J.; García, S.C. Development of a New Clinical Mastitis Detection Method for Automatic Milking Systems. J. Dairy Sci. 2018, 101, 9385–9395. [Google Scholar] [CrossRef]
- Wethal, K.B.; Svendsen, M.; Heringstad, B. A Genetic Study of New Udder Health Indicator Traits with Data from Automatic Milking Systems. J. Dairy Sci. 2020, 103, 7188–7198. [Google Scholar] [CrossRef]
- Youssif, N.H.; Hafiz, N.M.; Halawa, M.A.; Saad, M.F. Association of Selected Risk Factors with Bovine Subclinical Mastitis. Acta Vet. Bras. 2021, 15, 153–160. [Google Scholar] [CrossRef]
- Norberg, E.; Hogeveen, H.; Korsgaard, I.R.; Friggens, N.C.; Sloth, K.H.M.N.; Løvendahl, P. Electrical Conductivity of Milk: Ability to Predict Mastitis Status. J. Dairy Sci. 2004, 87, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Milner, P.; Page, K.L.; Walton, A.W.; Hillerton, J.E. Detection of Clinical Mastitis by Changes in Electrical Conductivity of Foremilk before Visible Changes in Milk. J. Dairy Sci. 1996, 79, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Lukas, J.M.; Reneau, J.K.; Wallace, R.; Hawkins, D.; Munoz-Zanzi, C. A Novel Method of Analyzing Daily Milk Production and Electrical Conductivity to Predict Disease Onset. J. Dairy Sci. 2009, 92, 5964–5976. [Google Scholar] [CrossRef] [PubMed]
- Kunes, R.; Bartos, P.; Iwasaka, G.K.; Lang, A.; Hankovec, T.; Smutny, L.; Cerny, P.; Poborska, A.; Smetana, P.; Kriz, P.; et al. In-Line Technologies for the Analysis of Important Milk Parameters during the Milking Process: A Review. Agric. 2021, 11, 239. [Google Scholar] [CrossRef]
- Naqvi, S.A.; King, M.T.M.; Matson, R.D.; DeVries, T.J.; Deardon, R.; Barkema, H.W. Mastitis Detection with Recurrent Neural Networks in Farms Using Automated Milking Systems. Comput. Electron. Agric. 2022, 192, 106618. [Google Scholar] [CrossRef]
- Juozaitienė, V.; Juozaitis, A.; Brazauskas, A.; Žymantienė, J.; Žilaitis, V.; Antanaitis, R.; Stankevičius, R.; Bobinienė, R. Investigation of Electrical Conductivity of Milk in Robotic Milking System and Its Relationship with Milk Somatic Cell Count and Other Quality Traits. J. Meas. Eng. 2015, 3, 63–70. [Google Scholar]
- Zaninelli, M.; Redaelli, V.; Luzi, F.; Bronzo, V.; Mitchell, M.; Dell’Orto, V.; Bontempo, V.; Cattaneo, D.; Savoini, G. First Evaluation of Infrared Thermography as a Tool for the Monitoring of Udder Health Status in Farms of Dairy Cows. Sensors 2018, 18, 862. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Bolaños, J.; Ceballes-Serrano, C.C.; Velásquez-Mejía, D.; Riaño-Rojas, J.C.; Giraldo, C.E.; Carmona, J.U.; Ceballos-Márquez, A. Application of Udder Surface Temperature by Infrared Thermography for Diagnosis of Subclinical Mastitis in Holstein Cows Located in Tropical Highlands. J. Dairy Sci. 2021, 104, 10310–10323. [Google Scholar] [CrossRef]
- Grodkowski, G.; Szwaczkowski, T.; Koszela, K.; Mueller, W.; Tomaszyk, K.; Baars, T.; Sakowski, T. Early Detection of Mastitis in Cows Using the System Based on 3D Motions Detectors. Sci. Rep. 2022, 12, 21215. [Google Scholar] [CrossRef]
- Sathiyabarathi, M.; Jeyakumar, S.; Manimaran, A.; Pushpadass, H.A.; Sivaram, M.; Ramesha, K.P.; Das, D.N.; Kataktalware, M.A.; Jayaprakash, G.; Patbandha, T.K. Investigation of Body and Udder Skin Surface Temperature Differentials as an Early Indicator of Mastitis in Holstein Friesian Crossbred Cows Using Digital Infrared Thermography Technique. Vet. World 2016, 9, 1386–1391. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, A.; Stordeur, P.; Wallemacq, H.; Schynts, F.; Stevens, M.; Boutet, P.; Peelman, L.J.; De Spiegeleer, B.; Duchateau, L.; Bureau, F.; et al. Variation of Inflammatory Dynamics and Mediators in Primiparous Cows after Intramammary Challenge with Escherichia Coli. Vet. Res. 2011, 42, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Machado, N.A.F.; Da Costa, L.B.S.; Barbosa-Filho, J.A.D.; De Oliveira, K.P.L.; De Sampaio, L.C.; Peixoto, M.S.M.; Damasceno, F.A. Using Infrared Thermography to Detect Subclinical Mastitis in Dairy Cows in Compost Barn Systems. J. Therm. Biol. 2021, 97, 102881. [Google Scholar] [CrossRef] [PubMed]
- Hovinen, M.; Siivonen, J.; Taponen, S.; Hänninen, L.; Pastell, M.; Aisla, A.M.; Pyörälä, S. Detection of Clinical Mastitis with the Help of a Thermal Camera. J. Dairy Sci. 2008, 91, 4592–4598. [Google Scholar] [CrossRef]
- Sepúlveda-Varas, P.; Proudfoot, K.L.; Weary, D.M.; von Keyserlingk, M.A.G. Changes in Behaviour of Dairy Cows with Clinical Mastitis. Appl. Anim. Behav. Sci. 2016, 175, 8–13. [Google Scholar] [CrossRef]
- Hallowell, G.D.; Palgrave, K. Ultrasound of the Bovine Mammary Gland and Its Role in Mastitis Control. Int. Dairy Top. 2012, 11, 43. [Google Scholar]
- Suzuki, N.; Kurose, T.; Kaneko, S.; Haraguchi, A.; Isobe, N. Outcome Prediction from the First Examination in Clinical Mastitis Using Ultrasonography in Dairy Cows. Anim. Sci. J. 2020, 91, 13452. [Google Scholar] [CrossRef]
- Flöck, M.; Winter, P. Diagnostic Ultrasonography in Cattle with Diseases of the Mammary Gland. Vet. J. 2006, 171, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.; Patil, D.; Kelawala, D.; Parikh, P.; Mer, D.; Gameti, K.; Gohil, K. Ultrasonography of Udder and Teat in Dairy Animals. Rumin. Sci. 2017, 6, 171–175. [Google Scholar]
- Jain, S.; Gautam, V.; Naseem, S. Acute-Phase Proteins: As Diagnostic Tool. J. Pharm. Bioallied Sci. 2011, 3, 118–127. [Google Scholar] [CrossRef]
- Turk, R.; Piras, C.; Kovačić, M.; Samardžija, M.; Ahmed, H.; De Canio, M.; Urbani, A.; Meštrić, Z.F.; Soggiu, A.; Bonizzi, L.; et al. Proteomics of Inflammatory and Oxidative Stress Response in Cows with Subclinical and Clinical Mastitis. J. Proteom. 2012, 75, 4412–4428. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.C.; Waterston, M.; Hastie, P.; Parkin, T.; Haining, H.; Eckersall, P.D. The Major Acute Phase Proteins of Bovine Milk in a Commercial Dairy Herd. BMC Vet. Res. 2015, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic-Filipovic, M.; Ilić, V.; Vujčić, Z.; Dojnov, B.; Stevanov-Pavlović, M.; Mijačević, Z.; Božić, T. Serum Amyloid A Isoforms in Serum and Milk from Cows with Staphylococcus Aureus Subclinical Mastitis. Vet. Immunol. Immunopathol. 2012, 145, 120–128. [Google Scholar] [CrossRef]
- Kleczkowski, M.; Kluciński, W.; Czerski, M.; Kudyba, E. Association between Acute Phase Response, Oxidative Status and Mastitis in Cows. Vet. Stanica 2017, 48, 177–186. [Google Scholar]
- Eckersall, P.D.; Young, F.J.; Nolan, A.M.; Knight, C.H.; McComb, C.; Waterston, M.M.; Hogarth, C.J.; Scott, E.M.; Fitzpatrick, J.L. Acute Phase Proteins in Bovine Milk in an Experimental Model of Staphylococcus Aureus Subclinical Mastitis. J. Dairy Sci. 2006, 89, 1488–1501. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Sugiyama, M.; Ito, T.; Tsukano, K.; Oikawa, S.; Suzuki, K. Diagnostic Utility of Measuring Serum Amyloid a with a Latex Agglutination Turbidimetric Immunoassay in Bovine Mastitis: Comparison with Haptoglobin and Alpha 1 Acid Glycoprotein. J. Vet. Med. Sci. 2021, 83, 329–332. [Google Scholar] [CrossRef]
- El-Deeb, W.M. Clinicobiochemical Investigations of Gangrenous Mastitis in Does: Immunological Responses and Oxidative Stress Biomarkers. J. Zhejiang Univ. Sci. B 2013, 14, 33–39. [Google Scholar] [CrossRef]
- Tothova, C.; Nagy, O.; Kovac, G. Acute Phase Proteins and Their Use in the Diagnosis of Diseases in Ruminants: A Review. Vet. Med. 2014, 59, 163–180. [Google Scholar] [CrossRef]
- Sadek, K.; Saleh, E.; Ayoub, M. Selective, Reliable Blood and Milk Bio-Markers for Diagnosing Clinical and Subclinical Bovine Mastitis. Trop. Anim. Health Prod. 2017, 49, 431–437. [Google Scholar] [CrossRef]
- Dalanezi, F.M.; Schmidt, E.M.S.; Joaquim, S.F.; Guimarães, F.F.; Guerra, S.T.; Lopes, B.C.; Cerri, R.L.A.; Chadwick, C.; Langoni, H. Concentrations of Acute-Phase Proteins in Milk from Cows with Clinical Mastitis Caused by Different Pathogens. Pathogens 2020, 9, 706. [Google Scholar] [CrossRef]
- Wollowski, L.; Heuwieser, W.; Kossatz, A.; Addis, M.F.; Puggioni, G.M.G.; Meriaux, L.; Bertulat, S. The Value of the Biomarkers Cathelicidin, Milk Amyloid A, and Haptoglobin to Diagnose and Classify Clinical and Subclinical Mastitis. J. Dairy Sci. 2021, 104, 2106–2122. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, M.C.; Healy, A.M.; Harte, D.; Walshe, K.G.; Torgerson, P.R.; Doherty, M.L. Milk Amyloid A: Correlation with Cellular Indices of Mammary Inflammation in Cows with Normal and Raised Serum Amyloid A. Res. Vet. Sci. 2006, 80, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Hiss, S.; Mueller, U.; Neu-Zahren, A.; Sauerwein, H. Haptoglobin and Lactate Dehydrogenase Measurements in Milk for the Identification of Subclinically Diseased Udder Quarters. Vet. Med. 2007, 52, 245–252. [Google Scholar] [CrossRef]
- Safi, S.; Khoshvaghti, A.; Jafarzadeh, S.R.; Bolourchi, M.; Nowrouzian, I. Acute Phase Proteins in the Diagnosis of Bovine Subclinical Mastitis. Vet. Clin. Pathol. 2009, 38, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Smolenski, G.A.; Wieliczko, R.J.; Pryor, S.M.; Broadhurst, M.K.; Wheeler, T.T.; Haigh, B.J. The Abundance of Milk Cathelicidin Proteins during Bovine Mastitis. Vet. Immunol. Immunopathol. 2011, 143, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Cubeddu, T.; Cacciotto, C.; Pisanu, S.; Tedde, V.; Alberti, A.; Pittau, M.; Dore, S.; Cannas, A.; Uzzau, S.; Rocca, S.; et al. Cathelicidin Production and Release by Mammary Epithelial Cells during Infectious Mastitis. Vet. Immunol. Immunopathol. 2017, 189, 66–70. [Google Scholar] [CrossRef]
- Ali, A.; Rehman, M.U.; Mushtaq, S.; Ahmad, S.B.; Khan, A.; Karan, A.; Bashir Wani, A.; Ganie, S.A.; Mir, M.U.R. Biochemical and Computational Assessment of Acute Phase Proteins in Dairy Cows Affected with Subclinical Mastitis. Curr. Issues Mol. Biol. 2023, 45, 338. [Google Scholar] [CrossRef]
- O’Reilly, E.L.; Viora, L.; Malcata, F.; Pepler, P.T.; Zadoks, R.; Brady, N.; Hanh, H.Q.; McLaughlin, M.; Horvatic, A.; Gelemanovic, A.; et al. Biomarker and Proteome Analysis of Milk from Dairy Cows with Clinical Mastitis: Determining the Effect of Different Bacterial Pathogens on the Response to Infection. Res. Vet. Sci. 2024, 172, 105240. [Google Scholar] [CrossRef] [PubMed]
- Spittel, S.; Hoedemaker, M. Mastitis Diagnosis in Dairy Cows Using PathoProof Real-Time Polymerase Chain Reaction Assay in Comparison with Conventional Bacterial Culture in a Northern German Field Study. Berl. Munch. Tierarztl. Wochenschr. 2012, 125, 494–502. [Google Scholar]
- Loy, J.D.; Clawson, M.L.; Adkins, P.R.F.; Middleton, J.R. Current and Emerging Diagnostic Approaches to Bacterial Diseases of Ruminants. Vet. Clin. North Am. Food Anim. Pract. 2023, 39, 93–114. [Google Scholar] [CrossRef]
- Wiggans, G.R.; Cole, J.B.; Hubbard, S.M.; Sonstegard, T.S. Genomic Selection in Dairy Cattle: The USDA Experience. Annu. Rev. Anim. Biosci. 2017, 5, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Cattle Quantitative Trait Locus Database. Available online: https://www.animalgenome.org/cgi-bin/QTLdb/BT/nscape?isID=1439 (accessed on 20 June 2024).
- Mai, M.D.; Rychtářová, J.; Zink, V.; Lassen, J.; Guldbrandtsen, B. Quantitative Trait Loci for Milk Production and Functional Traits in Two Danish Cattle Breeds. J. Anim. Breed. Genet. 2010, 127, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Minozzi, G.; Nicolazzi, E.L.; Strozzi, F.; Stella, A.; Negrini, R.; Ajmone-Marsan, P.; Williams, J.L. Genome Wide Scan for Somatic Cell Counts in Holstein Bulls. In Proceedings of the International Symposium on Animal Genomics for Animal Health (AGAH 2010), Paris, France, 31 May–2 June 2010; Volume 5. [Google Scholar]
- Sodeland, M.; Kent, M.P.; Olsen, H.G.; Opsal, M.A.; Svendsen, M.; Sehested, E.; Hayes, B.J.; Lien, S. Quantitative Trait Loci for Clinical Mastitis on Chromosomes 2, 6, 14 and 20 in Norwegian Red Cattle. Anim. Genet. 2011, 42, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Meredith, B.K.; Kearney, F.J.; Finlay, E.K.; Bradley, D.G.; Fahey, A.G.; Berry, D.P.; Lynn, D.J. Genome-Wide Associations for Milk Production and Somatic Cell Score in Holstein-Friesian Cattle in Ireland. BMC Genet. 2012, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wijga, S.; Bastiaansen, J.W.M.; Wall, E.; Strandberg, E.; de Haas, Y.; Giblin, L.; Bovenhuis, H. Genomic Associations with Somatic Cell Score in First-Lactation Holstein Cows. J. Dairy Sci. 2012, 95, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Sahana, G.; Ma, P.; Su, G.; Yu, Y.; Zhang, S.; Lund, M.S.; Sørensen, P. Exploring the Genetic Architecture and Improving Genomic Prediction Accuracy for Mastitis and Milk Production Traits in Dairy Cattle by Mapping Variants to Hepatic Transcriptomic Regions Responsive to Intra-Mammary Infection. Genet. Sel. Evol. 2017, 49, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kurz, J.P.; Yang, Z.; Weiss, R.B.; Wilson, D.J.; Rood, K.A.; Liu, G.E.; Wang, Z. A Genome-Wide Association Study for Mastitis Resistance in Phenotypically Well-Characterized Holstein Dairy Cattle Using a Selective Genotyping Approach. Immunogenetics 2019, 71, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Welderufael, B.G.; Løvendahl, P.; de Koning, D.J.; Janss, L.L.G.; Fikse, W.F. Genome-Wide Association Study for Susceptibility to and Recoverability from Mastitis in Danish Holstein Cows. Front. Genet. 2018, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafy, H.; Bortfeldt, R.H.; Reißmann, M.; Brockmann, G.A. P5000 Confirmation of Genome-Wide Associations for Clinical Mastitis in German Holstein Cattle. J. Anim. Sci. 2016, 94, 115–116. [Google Scholar] [CrossRef]
- Mustafa, H. Performance of Bovine High Density SNPs Genotyping Array in Indigenous Pakistani Cattle Breeds. Pure Appl. Biol. 2018, 7, 221–226. [Google Scholar] [CrossRef]
- Jamrozik, J.; Koeck, A.; Miglior, F.; Kistemaker, G.J.; Schenkel, F.S.; Kelton, D.F.; Doormaal, B.J. Van Genetic and Genomic Evaluation of Mastitis Resistance in Canada. Interbull Bull. 2013, 47, 43–51. [Google Scholar]
- Koeck, A.; Miglior, F.; Kelton, D.F.; Schenkel, F.S. Alternative Somatic Cell Count Traits to Improve Mastitis Resistance in Canadian Holsteins. J. Dairy Sci. 2012, 95, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Canadian Dairy Network Heritability Estimates Used for Genetic Evaluation in Canada. Available online: https://lactanet.ca/en/heritability-estimates-used-for-genetic-evaluation-in-canada/ (accessed on 20 July 2024).
- Lisuzzo, A.; Laghi, L.; Fiore, E.; Cecchinato, A.; Bisutti, V.; Pegolo, S.; Giannuzzi, D.; Tessari, R.; Barberio, A.; Schiavon, E.; et al. Serum Metabolome Differences Associated with Subclinical Intramammary Infection Caused by Streptococcus Agalactiae and Prototheca Spp. in Multiparous Dairy Cows. J. Dairy Sci. 2024, 107, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC-MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Hovinen, M.; Simojoki, H.; Pösö, R.; Suolaniemi, J.; Kalmus, P.; Suojala, L.; Pyörälä, S. N-Acetyl-β-D-Glucosaminidase Activity in Cow Milk as an Indicator of Mastitis. J. Dairy Res. 2016, 83, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Nyman, A.K.; Emanuelson, U.; Waller, K.P. Diagnostic Test Performance of Somatic Cell Count, Lactate Dehydrogenase, and N-Acetyl-β-d-Glucosaminidase for Detecting Dairy Cows with Intramammary Infection. J. Dairy Sci. 2016, 99, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Nirala, N.R.; Shtenberg, G. N-Acetyl-β-D-Glucosaminidase Biomarker Quantification in Milk Using Ag-Porous Si SERS Platform for Mastitis Severity Evaluation. Appl. Surf. Sci. 2021, 566, 150700. [Google Scholar] [CrossRef]
- Nirala, N.R.; Shtenberg, G. Gold Nanoparticle Size-Dependent Enhanced Chemiluminescence for Ultra-Sensitive Haptoglobin Biomarker Detection. Biomolecules 2019, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Nirala, N.R.; Harel, Y.; Lellouche, J.P.; Shtenberg, G. Ultrasensitive Haptoglobin Biomarker Detection Based on Amplified Chemiluminescence of Magnetite Nanoparticles. J. Nanobiotechnol. 2020, 18, 1–10. [Google Scholar] [CrossRef]
- Frundzhyan, V.G.; Parkhomenko, I.M.; Brovko, L.Y.; Ugarova, N.N. Improved Bioluminescent Assay of Somatic Cell Counts in Raw Milk. J. Dairy Res. 2008, 75, 3282. [Google Scholar] [CrossRef]
- Bobbo, T.; Ruegg, P.L.; Fiore, E.; Gianesella, M.; Morgante, M.; Pasotto, D.; Gallo, L.; Bittante, G.; Cecchinato, A. Short Communication: Association between Udder Health Status and Blood Serum Proteins in Dairy Cows. J. Dairy Sci. 2017, 100, 9775–9780. [Google Scholar] [CrossRef] [PubMed]
- Bobbo, T.; Fiore, E.; Gianesella, M.; Morgante, M.; Gallo, L.; Ruegg, P.L.; Bittante, G.; Cecchinato, A. Variation in Blood Serum Proteins and Association with Somatic Cell Count in Dairy Cattle from Multi-Breed Herds. Animal 2017, 11, 2309–2319. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L. A 100-Year Review: Mastitis Detection, Management, and Prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Ganda, E.K.; Bisinotto, R.S.; Decter, D.H.; Bicalho, R.C. Evaluation of an On-Farm Culture System (Accumast) for Fast Identification of Milk Pathogens Associated with Clinical Mastitis in Dairy Cows. PLoS ONE 2016, 11, e0155314. [Google Scholar] [CrossRef] [PubMed]
- Besier, J.; Lind, O.; Bruckmaier, R.M. Dynamics of Teat-End Vacuum during Machine Milking: Types, Causes and Impacts on Teat Condition and Udder Health—A Literature Review. J. Appl. Anim. Res. 2016, 44, 263–272. [Google Scholar] [CrossRef]
- Cerqueira, J.L.; Araújo, J.P.; Cantalapiedra, J.; Blanco-Penedo, I. How Is the Association of Teat-End Severe Hyperkeratosis on Udder Health and Dairy Cow Behavior? Rev. Med. Vet. 2018, 169, 30–37. [Google Scholar]
- Cardozo, L.L.; Thaler Neto, A.; Souza, G.N.; Picinin, L.C.A.; Felipus, N.C.; Reche, N.L.M.; Schmidt, F.A.; Werncke, D.; Simon, E.E. Risk Factors for the Occurrence of New and Chronic Cases of Subclinical Mastitis in Dairy Herds in Southern Brazil. J. Dairy Sci. 2015, 98, 7675–7685. [Google Scholar] [CrossRef] [PubMed]
- Guarín, J.F.; Ruegg, P.L. Short Communication: Pre- and Postmilking Anatomical Characteristics of Teats and Their Associations with Risk of Clinical Mastitis in Dairy Cows. J. Dairy Sci. 2016, 99, 8323–8329. [Google Scholar] [CrossRef] [PubMed]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine Mastitis: Prevalence, Risk Factors and Isolation of Staphylococcus Aureus in Dairy Herds at Hawassa Milk Shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef]
- Guarín, J.F.; Paixão, M.G.; Ruegg, P.L. Association of Anatomical Characteristics of Teats with Quarter-Level Somatic Cell Count. J. Dairy Sci. 2017, 100, 643–652. [Google Scholar] [CrossRef]
- Neijenhuis, F.; Barkema, H.W.; Hogeveen, H.; Noordhuizen, J.P.T.M. Relationship between Teat-End Callosity and Occurrence of Clinical Mastitis. J. Dairy Sci. 2001, 84, 2664–2672. [Google Scholar] [CrossRef] [PubMed]
- Pantoja, J.C.F.; Correia, L.B.N.; Rossi, R.S.; Latosinski, G.S. Association between Teat-End Hyperkeratosis and Mastitis in Dairy Cows: A Systematic Review. J. Dairy Sci. 2020, 103, 1843–1855. [Google Scholar] [CrossRef]
- Mein, G.A. The Role of the Milking Machine in Mastitis Control. Vet. Clin. North Am.—Food Anim. Pract. 2012, 28, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Vlkova, H.; Babak, V.; Vrtkova, I.; Cervinkova, D.; Marosevic, D.; Moravkova, M.; Jaglic, Z. Epidemiology of Intramammary Infections with Staphylococcus Aureus and Mastitis Streptococci in a Dairy Cattle Herd with a History of Recurrent Clinical Mastitis. Pol. J. Vet. Sci. 2017, 20, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Shittu, A.; Abdullahi, J.; Jibril, A.; Mohammed, A.A.; Fasina, F.O. Sub-Clinical Mastitis and Associated Risk Factors on Lactating Cows in the Savannah Region of Nigeria. BMC Vet. Res. 2012, 8, 134. [Google Scholar] [CrossRef]
- Abrahmsén, M.; Persson, Y.; Kanyima, B.M.; Båge, R. Prevalence of Subclinical Mastitis in Dairy Farms in Urban and Peri-Urban Areas of Kampala, Uganda. Trop. Anim. Health Prod. 2014, 46, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Rowe, S.; Tranter, W.; Laven, R. Longitudinal Study of Herd Udder Hygiene and Its Association with Clinical Mastitis in Pasture-Based Dairy Cows. J. Dairy Sci. 2021, 104, 6051–6060. [Google Scholar] [CrossRef]
- Schreiner, D.A.; Ruegg, P.L. Relationship between Udder and Leg Hygiene Scores and Subclinical Mastitis. J. Dairy Sci. 2003, 86, 3460–3465. [Google Scholar] [CrossRef]
- Köster, G.; Tenhagen, B.A.; Scheibe, N.; Heuwieser, W. Factors Associated with High Milk Test Day Somatic Cell Counts in Large Dairy Herds in Brandenburg. II. Milking Practices. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2006, 53, 209–214. [Google Scholar] [CrossRef]
- Zucali, M.; Bava, L.; Tamburini, A.; Brasca, M.; Vanoni, L.; Sandrucci, A. Effects of Season, Milking Routine and Cow Cleanliness on Bacterial and Somatic Cell Counts of Bulk Tank Milk. J. Dairy Res. 2011, 78, 436–441. [Google Scholar] [CrossRef]
- Sandrucci, A.; Bava, L.; Zucali, M.; Tamburini, A. Management Factors and Cow Traits Influencing Milk Somatic Cell Counts and Teat Hyperkeratosis during Different Seasons. Rev. Bras. Zootec. 2014, 43, 505–511. [Google Scholar] [CrossRef]
- Huijps, K.; Hogeveen, H.; Lam, T.J.G.M.; Oude Lansink, A.G.J.M. Costs and Efficacy of Management Measures to Improve Udder Health on Dutch Dairy Farms. J. Dairy Sci. 2010, 93, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Vissio, C.; Bouman, M.; Larriestra, A.J. Milking Machine and Udder Health Management Factors Associated with Bulk Milk Somatic Cell Count in Uruguayan Herds. Prev. Vet. Med. 2018, 150, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.T.; O’Sullivan, K.; Berry, D.P.; More, S.J.; Meaney, W.J.; O’Callaghan, E.J.; O’Brien, B. Farm Management Factors Associated with Bulk Tank Somatic Cell Count in Irish Dairy Herds. Ir. Vet. J. 2009, 62, 45–51. [Google Scholar] [CrossRef]
- Cortinhas, C.S.; Botaro, B.G.; de Macedo, S.N.; dos Santos, M.V. Herd Characteristics and Management Practices Associated with Bulk Tank Milk Quality of Dairy Herds in Southeastern Brazil. Trop. Anim. Health Prod. 2018, 50, 1605–1610. [Google Scholar] [CrossRef]
- Halasa, T.; Nielen, M.; van Werven, T.; Hogeveen, H. A Simulation Model to Calculate Costs and Benefits of Dry Period Interventions in Dairy Cattle. Livest. Sci. 2010, 129, 80–87. [Google Scholar] [CrossRef]
- Schewe, R.L.; Kayitsinga, J.; Contreras, G.A.; Odom, C.; Coats, W.A.; Durst, P.; Hovingh, E.P.; Martinez, R.O.; Mobley, R.; Moore, S.; et al. Herd Management and Social Variables Associated with Bulk Tank Somatic Cell Count in Dairy Herds in the Eastern United States. J. Dairy Sci. 2015, 98, 7650–7665. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Asthana, R.K.; Kumar, S.; Kumar, S.; Ghosh, S.; Upadhaya, B. Use of Dry Cow Therapy for Control of Mastitis in Dairy Animals. J. Krishi Vigyan 2020, 8, 303. [Google Scholar] [CrossRef]
- Stevens, M.; Piepers, S.; De Vliegher, S. The Effect of Mastitis Management Input and Implementation of Mastitis Management on Udder Health, Milk Quality, and Antimicrobial Consumption in Dairy Herds. J. Dairy Sci. 2019, 102, 2401–2415. [Google Scholar] [CrossRef]
- Ingle, H.D.; Rice, C.A.; Black, R.A.; Childers, S.Z.; Eberhart, N.L.; Prado, M.E.; Krawczel, P.D. Effect of Switch Trimming on Udder and Teat Hygiene of Dairy Cows. J. Appl. Anim. Welf. Sci. 2018, 21, 239–243. [Google Scholar] [CrossRef]
- Nickerson, S.C.; Oliver, S.P. REVIEW: How Well Have United States Dairy Producers Adopted Mastitis-Control Technologies for Reducing Herd Somatic Cell Counts Improving Milk Quality? Prof. Anim. Sci. 2014, 30, 115–124. [Google Scholar] [CrossRef]
- Salovuo, H.; Ronkainen, P.; Heino, A.; Suokannas, A.; Ryhänen, E.L. Introduction of Automatic Milking System in Finland: Effect on Milk Quality. Agric. Food Sci. 2005, 14, 346–353. [Google Scholar] [CrossRef]
- Kolenda, M.; Piwczyński, D.; Brzozowski, M.; Sitkowska, B.; Wójcik, P. Comparison of Yield, Composition and Quality of Milk of Polish Holstein-Friesian Cows in Conventional and Automatic Milking Systems. Ann. Anim. Sci. 2021, 21, 709–720. [Google Scholar] [CrossRef]
- Ivemeyer, S.; Simantke, C.; Ebinghaus, A.; Poulsen, P.H.; Sorensen, J.T.; Rousing, T.; Palme, R.; Knierim, U. Herd-Level Associations between Human–Animal Relationship, Management, Fecal Cortisol Metabolites, and Udder Health of Organic Dairy Cows. J. Dairy Sci. 2018, 101, 7361–7374. [Google Scholar] [CrossRef] [PubMed]
- Winnicki, S.; Jugowar, J.; Aerts, J.; Sobek, Z. The Effect of Milking Systems on the Auantity and Quality of Cow Milk. J. Res. Appl. Agric. 2017, 62, 193–196. [Google Scholar]
- Toušová, R.; Ducháček, J.; Stádník, L.; Ptáček, M.; Beran, J. Porovnání Produkce a Kvality Mléka v Dojírně a Systému AMS. J. Cent. Eur. Agric. 2014, 15, 100–114. [Google Scholar] [CrossRef]
- Qadri, H.; Shah, A.H.; Alkhanani, M.; Almilaibary, A.; Mir, M.A. Immunotherapies against Human Bacterial and Fungal Infectious Diseases: A Review. Front. Med. 2023, 10, 1135541. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Saleem Bhat, S.; Omonijo, A.F.; Ganai, A.N.; Ibeagha-Awemu, M.E.; Mudasir Ahmad, S. Immunotherapy in Mastitis: State of Knowledge, Research Gaps and Way Forward. Vet. Q. 2024, 44, 1–23. [Google Scholar] [CrossRef]
- Neefjes, J.; Jongsma, M.L.M.; Paul, P.; Bakke, O. Towards a Systems Understanding of MHC Class i and MHC Class II Antigen Presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef]
- Rainard, P.; Gilbert, F.B.; Germon, P.; Foucras, G. Invited Review: A Critical Appraisal of Mastitis Vaccines for Dairy Cows. J. Dairy Sci. 2021, 104, 10427–10448. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Dubovi, E.J.; Gröhn, Y.T.; Brunner, M.A.; Hertl, J.A. Response to Modified Live and Killed Multivalent Viral Vaccine in Regularly Vaccinated, Fresh Dairy Cows. Vet. Ther. 2000, 1, 49–58. [Google Scholar] [PubMed]
- Herry, V.; Gitton, C.; Tabouret, G.; Répérant, M.; Forge, L.; Tasca, C.; Gilbert, F.B.; Guitton, E.; Barc, C.; Staub, C.; et al. Local Immunization Impacts the Response of Dairy Cows to Escherichia Coli Mastitis. Sci. Rep. 2017, 7, 3441. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yuan, L.; Wang, T.; Cao, L.; Liu, F.; Song, J.; Zhang, Y. Construction of the WaaF Subunit and DNA Vaccine Against Escherichia Coli in Cow Mastitis and Preliminary Study on Their Immunogenicity. Front. Vet. Sci. 2022, 9, 877685. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Hu, C.; Gong, R.; Chen, Y.; Ren, N.; Xiao, G.; Xie, Q.; Zhang, M.; Liu, Q.; Guo, A.; et al. Evaluation of a Novel Chimeric B Cell Epitope-Based Vaccine against Mastitis Induced by Either Streptococcus Agalactiae or Staphylococcus Aureus in Mice. Clin. Vaccine Immunol. 2011, 18, 893–900. [Google Scholar] [CrossRef][Green Version]
- Kiku, Y.; Ozawa, T.; Takahashi, H.; Kushibiki, S.; Inumaru, S.; Shingu, H.; Nagasawa, Y.; Watanabe, A.; Hata, E.; Hayashi, T. Effect of Intramammary Infusion of Recombinant Bovine GM-CSF and IL-8 on CMT Score, Somatic Cell Count, and Milk Mononuclear Cell Populations in Holstein Cows with Staphylococcus Aureus Subclinical Mastitis. Vet. Res. Commun. 2017, 41, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Kiku, Y.; Mizuno, M.; Inumaru, S.; Kushibiki, S.; Shingu, H.; Matsubara, T.; Takahashi, H.; Hayashi, T. Effect of Intramammary Infusion of RbGM-CSF on SCC and Expression of Polymorphonuclear Neutrophil Adhesion Molecules in Subclinical Mastitis Cows. Vet. Res. Commun. 2012, 36, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.B.; Albright, B.N.; Caswell, J.L. Effect of Interleukin-8 and Granulocyte Colony-Stimulating Factor on Priming and Activation of Bovine Neutrophils. Infect. Immun. 2003, 71, 1643–1649. [Google Scholar] [CrossRef]
- Daley, M.J.; Furda, G.; Dougherty, R.; Coyle, P.A.; Williams, T.J.; Johnston, P. Potentiation of Antibiotic Therapy for Bovine Mastitis by Recombinant Bovine Interleukin-2. J. Dairy Sci. 1992, 75, 3330–3338. [Google Scholar] [CrossRef]
- Daley, M.J.; Coyle, P.A.; Williams, T.J.; Furda, G.; Dougherty, R.; Hayes, P.W. Staphylococcus Aureus Mastitis: Pathogenesis and Treatment with Bovine Interleukin-1β and Interleukin-2. J. Dairy Sci. 1991, 74, 4413–4424. [Google Scholar] [CrossRef]
- Quiroga, G.H.; Sordillo, L.M.; Adkinson, R.W.; Nickerson, S.C. Cytologic Responses of Staphylococcus Aureus-Infected Mammary Glands of Heifers to Interferon Gamma and Interleukin-2 Treatment. Am. J. Vet. Res. 1993, 54, 1894–1900. [Google Scholar] [CrossRef] [PubMed]
- Zecconi, A.; Piccinini, R.; Fiorina, S.; Cabrini, L.; Daprà, V.; Amadori, M. Evaluation of Interleukin-2 Treatment for Prevention of Intramammary Infections in Cows after Calving. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Yagi, Y.; Shiono, H.; Yokomizo, Y.; Inumaru, S. Effects of Intramammary Infusions of Interleukin-8 on Milk Protein Composition and Induction of Acute-Phase Protein in Cows during Mammary Involution. Can. J. Vet. Res. 2008, 72, 291–296. [Google Scholar] [PubMed]
- Watanabe, A.; Hirota, J.; Shimizu, S.; Inumaru, S.; Kimura, K. Single Intramammary Infusion of Recombinant Bovine Interleukin-8 at Dry-off Induces the Prolonged Secretion of Leukocyte Elastase, Inflammatory Lactoferrin-Derived Peptides, and Interleukin-8 in Dairy Cows. Vet. Med. Int. 2012, 2012, 172072. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Yagi, Y.; Shiono, H.; Yokomizo, Y. Effect of Intramammary Infusion of Tumour Necrosis Factor-α on Milk Protein Composition and Induction of Acute-Phase Protein in the Lactating Cow. J. Vet. Med. Ser. B 2000, 47, 653–662. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Babiuk, L.A. Controlling Acute Escherichia Coli Mastitis during the Periparturient Period with Recombinant Bovine Interferon Gamma. Vet. Microbiol. 1991, 28, 189–198. [Google Scholar] [CrossRef]
- Solis-Castillo, L.A.; Garcia-Romo, G.S.; Diaz-Rodriguez, A.; Reyes-Hernandez, D.; Tellez-Rivera, E.; Rosales-Garcia, V.H.; Mendez-Cruz, A.R.; Jimenez-Flores, J.R.; Villafana-Vazquez, V.H.; Pedroza-Gonzalez, A. Tumor-Infiltrating Regulatory T Cells, CD8/Treg Ratio, and Cancer Stem Cells Are Correlated with Lymph Node Metastasis in Patients with Early Breast Cancer. Breast Cancer 2020, 27, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F. Molecular and Cellular Pathways of Immunoglobulin G Activity In Vivo. ISRN Immunol. 2014, 2014, 524081. [Google Scholar] [CrossRef]
- Wang, L.H.; Li, X.Y.; Jin, L.J.; You, J.S.; Zhou, Y.; Li, S.Y.; Xu, Y.P. Characterization of Chicken Egg Yolk Immunoglobulins (IgYs) Specific for the Most Prevalent Capsular Serotypes of Mastitis-Causing Staphylococcus Aureus. Vet. Microbiol. 2011, 149, 415–421. [Google Scholar] [CrossRef]
- Zhen, Y.H.; Jin, L.J.; Guo, J.; Li, X.Y.; Li, Z.; Fang, R.; Xu, Y.P. Characterization of Specific Egg Yolk Immunoglobulin (IgY) against Mastitis-Causing Staphylococcus Aureus. J. Appl. Microbiol. 2008, 105, 1529–1535. [Google Scholar] [CrossRef]
- Zhen, Y.H.; Jin, L.J.; Li, X.Y.; Guo, J.; Li, Z.; Zhang, B.J.; Fang, R.; Xu, Y.P. Efficacy of Specific Egg Yolk Immunoglobulin (IgY) to Bovine Mastitis Caused by Staphylococcus Aureus. Vet. Microbiol. 2009, 133, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.A.; Kerro-Dego, O.; Prado, M.E.; Headrick, S.I.; Lewis, M.J.; Siebert, L.J.; Pighetti, G.M.; Oliver, S.P. Protective Effect of Anti-SUAM Antibodies on Streptococcus Uberis Mastitis. Vet. Res. 2015, 46, 133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Tian, X.; Wang, Q.; Hao, J.; Jiang, J.; Wang, H. Regulation Mechanisms and Maintenance Strategies of Stemness in Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2024, 20, 455–483. [Google Scholar] [CrossRef] [PubMed]
- Yusoff, M.F.; Higashi, Y. Mesenchymal Stem/Stromal Cells for Therapeutic Angiogenesis. Cells 2023, 12, 2162. [Google Scholar] [CrossRef] [PubMed]
- Peralta, O.A.; Carrasco, C.; Vieytes, C.; Tamayo, M.J.; Muñoz, I.; Sepulveda, S.; Tadich, T.; Duchens, M.; Melendez, P.; Mella, A.; et al. Safety and Efficacy of a Mesenchymal Stem Cell Intramammary Therapy in Dairy Cows with Experimentally Induced Staphylococcus Aureus Clinical Mastitis. Sci. Rep. 2020, 10, 2843. [Google Scholar] [CrossRef] [PubMed]
- Saenz-de-Juano, M.D.; Silvestrelli, G.; Bauersachs, S.; Ulbrich, S.E. Determining Extracellular Vesicles Properties and MiRNA Cargo Variability in Bovine Milk from Healthy Cows and Cows Undergoing Subclinical Mastitis. BMC Genom. 2022, 23, 189. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Tamma, M.; Pathigadapa, U.; Reddanna, P.; Yenuganti, V.R. Drug Loading and Functional Efficacy of Cow, Buffalo, and Goat Milk-Derived Exosomes: A Comparative Study. Mol. Pharm. 2022, 19, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.; Atalla, H.; Karrow, N.; Mallard, B.A. The Bioactivity of Colostrum and Milk Exosomes of High, Average, and Low Immune Responder Cows on Human Intestinal Epithelial Cells. J. Dairy Sci. 2021, 104, 2499–2510. [Google Scholar] [CrossRef] [PubMed]
- Somiya, M.; Yoshioka, Y.; Ochiya, T. Biocompatibility of Highly Purified Bovine Milk-Derived Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1440132. [Google Scholar] [CrossRef]
- Zempleni, J.; Sukreet, S.; Zhou, F.; Wu, D.; Mutai, E. Milk-Derived Exosomes and Metabolic Regulation. Annu. Rev. Anim. Biosci. 2019, 7, 245–262. [Google Scholar] [CrossRef]
- Del Pozo-Acebo, L.; de las Hazas, M.C.L.; Tomé-Carneiro, J.; Gil-Cabrerizo, P.; San-Cristobal, R.; Busto, R.; García-Ruiz, A.; Dávalos, A. Bovine Milk-Derived Exosomes as a Drug Delivery Vehicle for Mirna-Based Therapy. Int. J. Mol. Sci. 2021, 22, 1105. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Sun, X.; Huang, M.; Ma, Q.; Du, L.; Cui, Y. Enhanced Neuroprotective Effects of Epicatechin Gallate Encapsulated by Bovine Milk-Derived Exosomes against Parkinson’s Disease through Antiapoptosis and Antimitophagy. J. Agric. Food Chem. 2021, 69, 5134–5143. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Shang, F.; Chen, X.; Ni, J.; Yu, L.; Zhang, M.; Sun, D.; Xue, T. The Anti-Biofilm Effect of Silvernanoparticle-Decorated Quercetin Nanoparticles on a Multi-Drug Resistant Escherichia Coli Strain Isolated from a Dairy Cow with Mastitis. PeerJ 2018, 2018, 5711. [Google Scholar] [CrossRef] [PubMed]
- Castelani, L.; Arcaro, J.R.P.; Braga, J.E.P.; Bosso, A.S.; Moura, Q.; Esposito, F.; Sauter, I.P.; Cortez, M.; Lincopan, N. Short Communication: Activity of Nisin, Lipid Bilayer Fragments and Cationic Nisin-Lipid Nanoparticles against Multidrug-Resistant Staphylococcus Spp. Isolated from Bovine Mastitis. J. Dairy Sci. 2019, 102, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Kalińska, A.; Jaworski, S.; Wierzbicki, M.; Gołębiewski, M. Silver and Copper Nanoparticles—An Alternative in Future Mastitis Treatment and Prevention? Int. J. Mol. Sci. 2019, 20, 1672. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro Machado, G.T.; Veleirinho, M.B.; Mazzarino, L.; Machado Filho, L.C.P.; Maraschin, M.; Cerri, R.L.A.; Kuhnen, S. Development of Propolis Nanoparticles for the Treatment of Bovine Mastitis: In Vitro Studies on Antimicrobial and Cytotoxic Activities. Can. J. Anim. Sci. 2019, 99, 713–723. [Google Scholar] [CrossRef]
- Orellano, M.S.; Isaac, P.; Breser, M.L.; Bohl, L.P.; Conesa, A.; Falcone, R.D.; Porporatto, C. Chitosan Nanoparticles Enhance the Antibacterial Activity of the Native Polymer against Bovine Mastitis Pathogens. Carbohydr. Polym. 2019, 213, 1–9. [Google Scholar] [CrossRef]
- Zhu, L.; Cao, X.; Xu, Q.; Su, J.; Li, X.; Zhou, W. Evaluation of the Antibacterial Activity of Tilmicosin-SLN against Streptococcus Agalactiae: In Vitro and in Vivo Studies. Int. J. Nanomed. 2018, 13, 4747–4755. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).