Abstract
This study aimed to determine the optimal growth stage and additives for producing high quality millet silage through two experiments. Experiment 1: Whole-plant millet from the same field and under uniform management was harvested at the heading, sizing, milking, dough, and full-maturity stages. Then, it was chopped into 2–3 cm segments, vacuum-sealed in plastic bags without any further treatment, stored at 20 °C, and opened after 60 days. The results indicated that the dough stage had the highest water-soluble carbohydrate (WSC) and crude protein (CP) contents. The lactic acid (LA) and acetic acid (AA) contents during the dough and maturity stages were significantly higher than other stages, with the lowest pH observed during the dough stage. Experiment 2: The whole-plant millet was harvested at the dough stage. It was then chopped into 2–3 cm segments using a forage chopper, mixed thoroughly, and subjected to different treatments—inoculation with 106 CFU/g FM of Lactiplantibacillus plantarum (LP), adding of 1% FM sucrose (S), and a combination of Lactiplantibacillus plantarum and sucrose (MIX)—with a control group (CK) receiving an equivalent amount of water. The MIX treatment significantly enhanced the WSC content compared to other treatments (p < 0.05), and both the LP and MIX treatments showed superior LA and AA contents and lactic acid bacteria counts. These additives significantly altered the bacterial community, shifting dominance from Proteobacteria in the CK and raw materials to Firmicutes. Klebsiella dominated the CK group but was significantly reduced in the additive treatments, where Lentilactobacillus became the dominant genus. Therefore, we recommend harvesting millet at the dough stage and adding a mixture of Lactiplantibacillus plantarum and sugar to improve fermentation quality and aerobic stability.
1. Introduction
Millet (Setaria italica) is one of the world’s most important and oldest domesticated crops. Noted for its low water requirement among all cereal crops and its adaptability to grow well in poor soils [1], it is also a short-season crop. Millet is rich in crude protein and fiber, making it an excellent feedstock [2]. However, its high moisture content can make storage challenging, and silage is a preferred method for preserving such feedstock. Silage involves chopping fresh plants, compacting them to reduce oxygen, and allowing anaerobic fermentation to convert soluble carbohydrates into organic acids, reducing pH and inhibiting the growth of spoilage organisms, thereby preserving nutrients [3].
Due to its soft texture, good nutritional value, and palatability, whole-plant millet has high potential for animal feed. Research by Costa and others has shown that intercropping millet with Paiaguas palisadegrass in different pasture systems enhances the dry matter content of the silage [4]. Amer and others suggest that, compared to corn silage, millet silage contains higher CP and ADF levels. Replacing 67% of diet silage with millet silage in dairy cows’ feed did not affect dry matter and crude protein intake, milk production, or lactose concentrations, but did increase milk fat concentration, thereby providing more energy-corrected milk [5].
Achieving high-quality whole-plant millet silage is complex due to the influence of multiple factors. Santos et al. [6] explored the effect of the growth period on the nutritional quality of Pennisetum glaucum (L.) R. silage and found that the growth period of pearl millet had a significant effect on the nutritional quality of silage, among which the DM content increased with the extension of growth period, while CP and ADF showed a decreasing trend. Hill et al. [7] demonstrated that adding silage inoculants can improve the fermentation characteristics and dry matter (DM) content of millet silage, also providing additional energy when fed to cattle.
Previous studies have shown that the nutritional composition of whole-plant millet, such as cellulose, carbohydrates, protein, moisture content, and the number of naturally adhering lactic acid bacteria, varies at different growth stages [6]. These factors are crucial in influencing the fermentation quality of silage. However, whole-plant millet has low water-soluble carbohydrate (WSC) content before ensiling, which may not meet the substrate requirements for high-quality silage fermentation without any treatments [8]. Research indicates that Lactiplantibacillus plantarum has excellent fermentation capabilities, promoting rapid fermentation of silage, lowering pH levels, inhibiting the growth of harmful microorganisms, and helping to improve silage quality [9]. Muck et al. [10] noted that silage microbial inoculants can significantly alter various aspects of silage fermentation, such as pH, lactic acid (LA) content, acetic acid (AA) content, DM loss, and the digestibility of DM and fiber. Similarly, Rinne, M et al. [11] suggest that the addition of LAB to silage accelerates LA fermentation compared to untreated material. Wang, X et al. [12] emphasized that LAB is a key factor in improving the fermentation quality and nutritional value of silage.
Therefore, we hypothesize that the nutritional composition and the quantity and types of adhering microorganisms vary across the important growth stages of millet. The addition of the widely recognized Lactiplantibacillus plantarum and soluble sugars can regulate the microbial diversity of millet silage, thereby controlling the fermentation quality. Based on this, our experiment focuses on whole-plant millet, studying the effects of different growth stages and the addition of Lactiplantibacillus plantarum and soluble sugars on the fermentation quality, aerobic stability, and bacterial communities of millet. The aim is to explore how various growth stages and additives influence the nutritional components, fermentation quality, aerobic stability, and bacterial dynamics of the silage. The goal is to provide a theoretical foundation for the development of whole-plant millet silage formulations and their application in feed.
2. Materials and Methods
2.1. Raw Material and Silage Preparation
Experiment 1: Whole-plant millet from the same field and under uniform management was harvested at the heading, sizing, milking, dough, and full-maturity stages. It was then chopped into 2–3 cm segments using a forage chopper, mixed thoroughly, and packed into silage bags weighing 300 g each without any further treatment. These bags were vacuum-sealed, with four replicates per treatment, and stored at room temperature (20 °C) for 60 days.
Experiment 2: A randomized block design was employed to evaluate the effects of different treatments on whole-plant millet silage harvested at the dough stage. The factors in this design are the treatments, which included inoculation with 106 CFU/g FM of Lactiplantibacillus plantarum (LP group), adding 1% FM sucrose (S group), and a combination of Lactiplantibacillus plantarum (106 CFU/g FM) and sucrose (1% FM) (MIX group), with a control group (CK) receiving an equivalent amount of water. The additive used was a homofermentative bacterium, classified as a biological additive. We dissolved the bacterial powder and sugar in water and added them in liquid form. Each treatment was randomly assigned within each block, with four replicates per treatment. The chopped millet segments were mixed thoroughly and packed into silage bags weighing 300 g each. The silage was stored in the laboratory, with the temperature maintained at around 20 °C throughout the entire storage period. At silo opening and after 7 days of aerobic exposure, the samples were analyzed to evaluate the impact of the various additives on the nutritional components, fermentation quality, aerobic stability, and bacterial communities of the whole-plant millet silage.
2.2. Silage Extract Preparation
After opening the silo, the silage was thoroughly mixed. A 20 g sample was then placed in a 200 mL conical flask, to which 180 mL of distilled water was added. The mixture was blended for 1 min. The resultant filtrate was passed through double-layer cheesecloth and subsequently through a No. 12 filter paper (Fushun city Minsheng filter paper factory, Fushun, China). The collected filtrate was used for pH analysis, microbial count, and water-soluble carbohydrate (WSC) content analysis. For organic acid analysis, the filtrate was further filtered through a 0.22 µm filter and stored at −20 °C.
2.3. Nutritional Component Analysis
An amount of 100 g of silage sample was dried in an oven (Shanghai YiHeng Scientific, Shanghai, China) at 65 °C for 48 h to determine the DM content. The dried samples were ground using a grinder (FS-6D; Shenzhen Fuchi Machinery Equipment Co., Ltd., Shenzhen, China), and samples were collected for nutritional component analysis. CP content was determined via the Kjeldahl method using an automatic Kjeldahl apparatus (KT8000, Foss Co., Ltd., Hillerød, Denmark). NDF and ADF contents were determined using an ANKOM A2000 fiber analyzer (Ankom Technology, Macedon, NY, USA). WSC content was measured using the anthrone–sulfuric acid method [13].
2.4. Fermentation Quality Analysis
The pH was measured using a pH meter (PB-10 Sartorius, Gottingen, Germany). Organic acids were analyzed using a high-performance liquid chromatograph (SHI-MADZE-10A, chromatographic column: Shodex Rspak KC-811 S-DVB gel Column 30 mm × 8 mm, detector: SPD-M10AVP, mobile phase: 3 mmol/L perchloric acid, flow rate: 0.6 mL/min; column temperature 50 °C, detection wavelength 210 nm, sample volume 5 µL).
2.5. Aerobic Stability Analysis
After opening the silo, 130 g of the sample was transferred into a 500 mL polyethylene bottle and compacted. A thermometer probe (model AZ88598, Hengxin Technology Co., Ltd., Shenzhen, China) was inserted at the center of the sample without touching the bottle. We then placed paper scraps in an empty bottle and inserted the thermometer probe without it touching the sides to record the temperature, which was considered the room temperature. The temperature of the sample was recorded every 15 min. If the sample’s temperature exceeded the ambient temperature by more than 2 °C, indicating the onset of spoilage, the time of this occurrence was noted.
2.6. Microbial Count
Microbial counts were assessed using the plate count method. Various media supplied by Guangzhou HuanKai Microbial Sci. & Tech. Co., Ltd., Guangzhou, China, were employed for counting different microbes: lactic acid bacteria were cultured on De Man Rogosa Sharpe medium, yeast on potato dextrose agar (PDA), and general bacteria on nutrient agar (NA). These cultures were incubated at 37 °C for 48 h prior to counting.
2.7. Bacterial Community Analysis
Following the method of Mu et al. [14], DNA was extracted using an EZNA® Stool DNA Kit. The primers 341F (5′-CCTACGGGNGGCWGCAG-3′) and 805R (5′-GACTACHVGGGTATCTAATCC-3′) were used for polymerase chain reaction (PCR) to amplify the V3-V4 region of the bacterial 16S rDNA gene. PCR products were purified using AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA), quantified using Qubit (Invitrogen, Waltham, MA, USA) [15], and sequenced on the Illumina NovaSeq PE250 platform. The raw data from the sequencer were assembled using FLASH (v1.2.8), low-quality sequences were removed using fqtrim (v0.94), and chimeras were filtered using Vsearch (v2.3.4). Bacterial community diversity was calculated using QIIME2 (2019.7). Additionally, sequence annotation was performed using BLAST (v2.16.0); the comparison databases were SILVA and NT-16S. Related charts were drawn using R-3.4.4.
2.8. Data Processing
Experimental data were preprocessed using Excel 2016. The effects of growth stage and additives on nutritional component, fermentation quality and microbial count and diversity were analyzed with one-way ANOVA. Tukey’s multiple comparison method was used to analyze the difference between the least squares means of each treatment group. The results are presented as mean ± standard deviation (Mean ± SEM).
3. Results
3.1. Effect of Different Growth Stages on Nutritional Components of Whole-Plant Millet Silage
As the growth stage progressed, the DM content gradually increased, with the milk and dough stages around 30% and 35%, respectively, reaching the highest at the full-maturity stage (40%). CP content had a decreasing trend before increasing, with a significant increase at the dough stage compared to the milking and sizing stages (p < 0.05). The WSC content during the sizing and dough stages was significantly higher than in the other growth stages (p < 0.05), with no significant difference between them. At the full-maturity stage, the WSC content significantly decreased compared to other stages (p < 0.05). Throughout maturation, the NDF content exhibited wave-like fluctuations, generally decreasing and reaching its lowest at full maturity of 54.2%. There were no significant differences in the ADF content between growth stages (Table 1).

Table 1.
Effects of different growth stages on nutrients of whole-plant millet silage.
The number of lactic acid bacteria (LAB) was lowest during the milk stage and highest at full-maturity stage (p < 0.05). Yeast counts were lowest in the milk stage and highest in the sizing stage. The general bacteria numbers were highest in the milk stage, significantly higher than in other stages (p < 0.05), and notably lower in the dough and full-maturity stages compared to the other stages (p < 0.05) (Table 2).
Table 2.
Effects of different growth stages on microbial counts of whole-plant millet silage.
The highest pH value was observed in the full-maturity stage (4.20), with the lowest in the milk stage (4.03), with significant differences (p < 0.05). Additionally, the lactic acid (LA) content during the heading, dough, and full-maturity stages was significantly higher than during the milking and dough stages (p < 0.05), although no significant differences were observed among the heading, dough, and full-maturity stages. As the growth stages progressed, the acetic acid (AA) content exhibited an increasing trend, leading to a decreasing trend in the LA/AA ratio. Specifically, the AA content at the dough stage was significantly higher than at the milk stage (p < 0.05). No propionic acid (PA) and butyric acid (BA) were detected in the silage at any growth stage (Table 3).
Table 3.
Effect of different growth stages on fermentation quality of whole-plant millet silage.
3.2. Effect of Different Additives on the Quality of Whole-Plant Millet Silage
The DM content in the S group was the highest at 38.5%, significantly exceeding that of the CK group (p < 0.05). At silo opening, the ADF content in the S group was significantly lower than in the other groups (p < 0.05), and the NDF content was also lower than in the other groups, although not significantly. After the aerobic stability test, the NDF and ADF contents in all treated groups were significantly lower than those in the CK group (p < 0.05). There were no significant differences in CP content among the treatment groups. The WSC content was higher in all additive groups compared to the CK group, with the MIX group displaying the highest level, significantly outperforming other treatment groups (p < 0.05) (Table 4).
Table 4.
Effects of different additives on nutrients at silo opening and after aerobic stability test of whole-plant millet silage.
LAB counts in all treatment groups were significantly greater than in the CK group, and the counts of general bacteria were significantly lower than in the CK group (p < 0.05). The general bacteria counts in the LP and S groups were notably lower than in the MIX group, while the yeast count in the MIX group was significantly higher than in the CK group (p < 0.05) (Table 5).
Table 5.
Effects of different additives on microbial counts at silo opening and after aerobic stability test of whole-plant millet silage.
3.3. Effect of Different Additives on the Aerobic Stability of Whole-Plant Millet Silage
During 7 days of aerobic exposure, there was no significant temperature increase in any of the experimental groups, with aerobic stability exceeding 168 h. However, post-aerobic stability testing revealed an increase in pH values across all groups, although all additive groups remained significantly lower than the CK group (p < 0.05). Additionally, the LA and AA contents in all treatment groups, except the S group, decreased compared to their levels upon opening, with the most notable decrease observed in the LP group (p < 0.01) (Table 6).
Table 6.
Effect of different additives on fermentation quality of whole-grain silage before and after aerobic stabilization experiment.
3.4. Effect of Different Additives on the Bacterial Communities of Whole-Plant Millet Silage
All treatment groups achieved a Good’s coverage index of 1.00. The Chao1, Shannon, and Simpson indices decreased for all treatments, with the additive groups showing significantly lower Shannon diversity compared to the CK group (p < 0.05). The MIX group had the lowest Shannon and Simpson indices, followed by the LP group. After the aerobic stability test, all groups except CK showed an increase in these diversity indices (Table 7).
Table 7.
Effects of different additives on α diversity before and after aerobic stability test of whole-plant millet silage.
PCA analysis revealed that the microbial communities in the raw materials were distinct from those in the silage treatments, indicating significant changes in microbial composition due to the ensiling process. Notably, the CK and MIX groups demonstrated closely related microbial communities both before and after the aerobic stability test, suggesting that these treatments may have similar impacts on microbial dynamics in the silage (Figure 1).
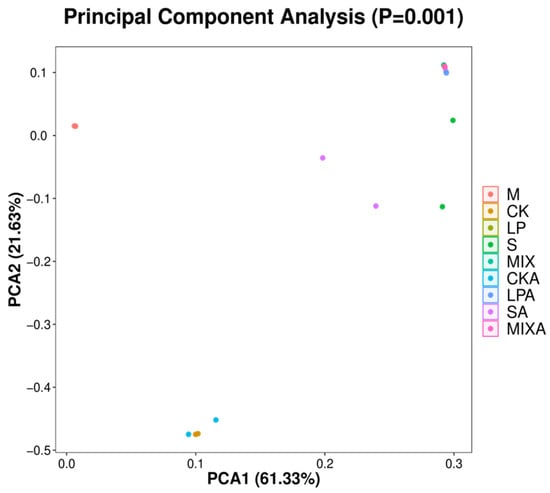
Figure 1.
Principal component analysis (PCA) of bacterial communities in whole-plant millet raw material and silage. Note: M: raw material; CK, LP, S, and MIX represent control, Lactiplantibacillus plantarum, sucrose, and a mixture of Lactiplantibacillus plantarum and sucrose treatments, respectively, at silo opening. Corresponding CKA, LPA, SA, and MIXA denote the groups after the aerobic stability test.
At the phylum level, Proteobacteria and Firmicutes were predominant across all samples. In the raw materials and the CK group, Proteobacteria were dominant, with relative abundances of 91.6% and 88.6% respectively, while Firmicutes were less prevalent at 5.16% and 11.29%. Conversely, in the LP, S, and MIX groups, the abundance of Firmicutes significantly increased to 94.64%, 63.63%, and 95.26%, respectively, effectively suppressing the dominant Proteobacteria observed in the CK group. This shift highlights the effectiveness of the additives in altering the microbial landscape of the silage (Figure 2 and Figure 3a).
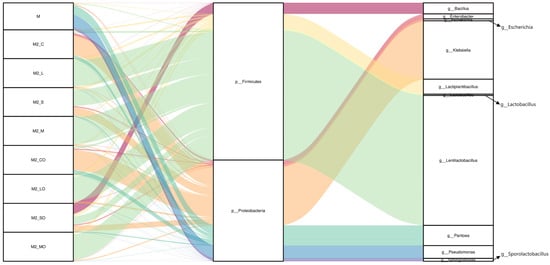
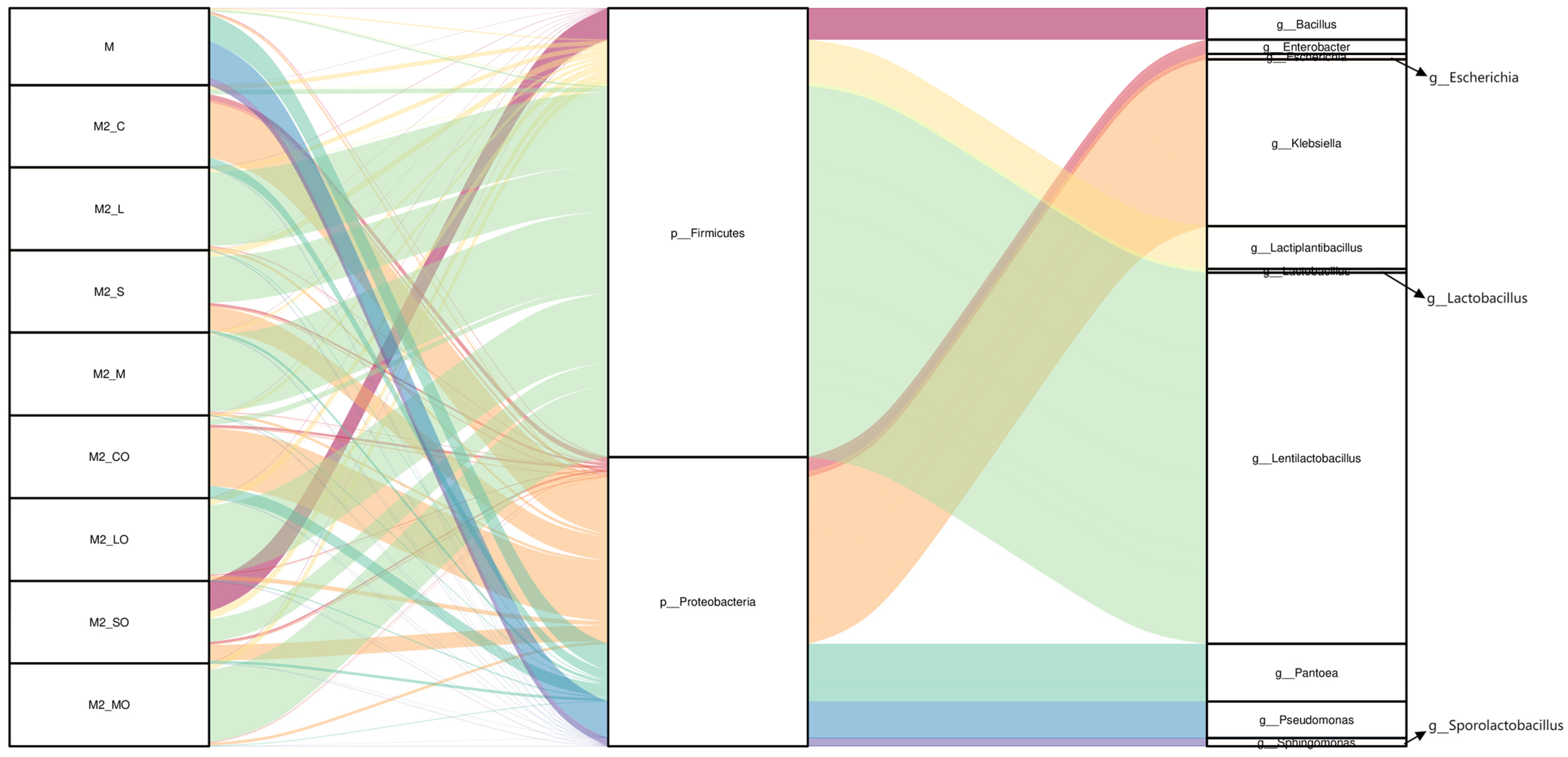
Figure 2.
Relative abundance of bacterial communities in whole-plant millet raw material and silage. Note: M: raw material; M2_C, M2_L, M2_S, and M2_M represent control, Lactiplantibacillus plantarum, sucrose, and a mixture of Lactiplantibacillus plantarum and sucrose treatments, respectively, at silo opening. M2_CO, M2_LO, M2_SO, and M2_MO denote the groups post-aerobic stabilization. CK, LP, S, and MIX represent control, Lactiplantibacillus plantarum, sucrose, and a mixture of Lactiplantibacillus plantarum and sucrose treatments, respectively. The corresponding CKA, LPA, SA, and MIXA denote the groups after the aerobic stability test.
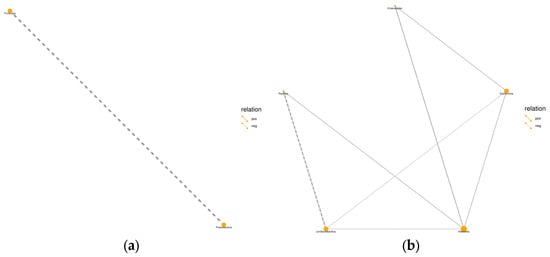
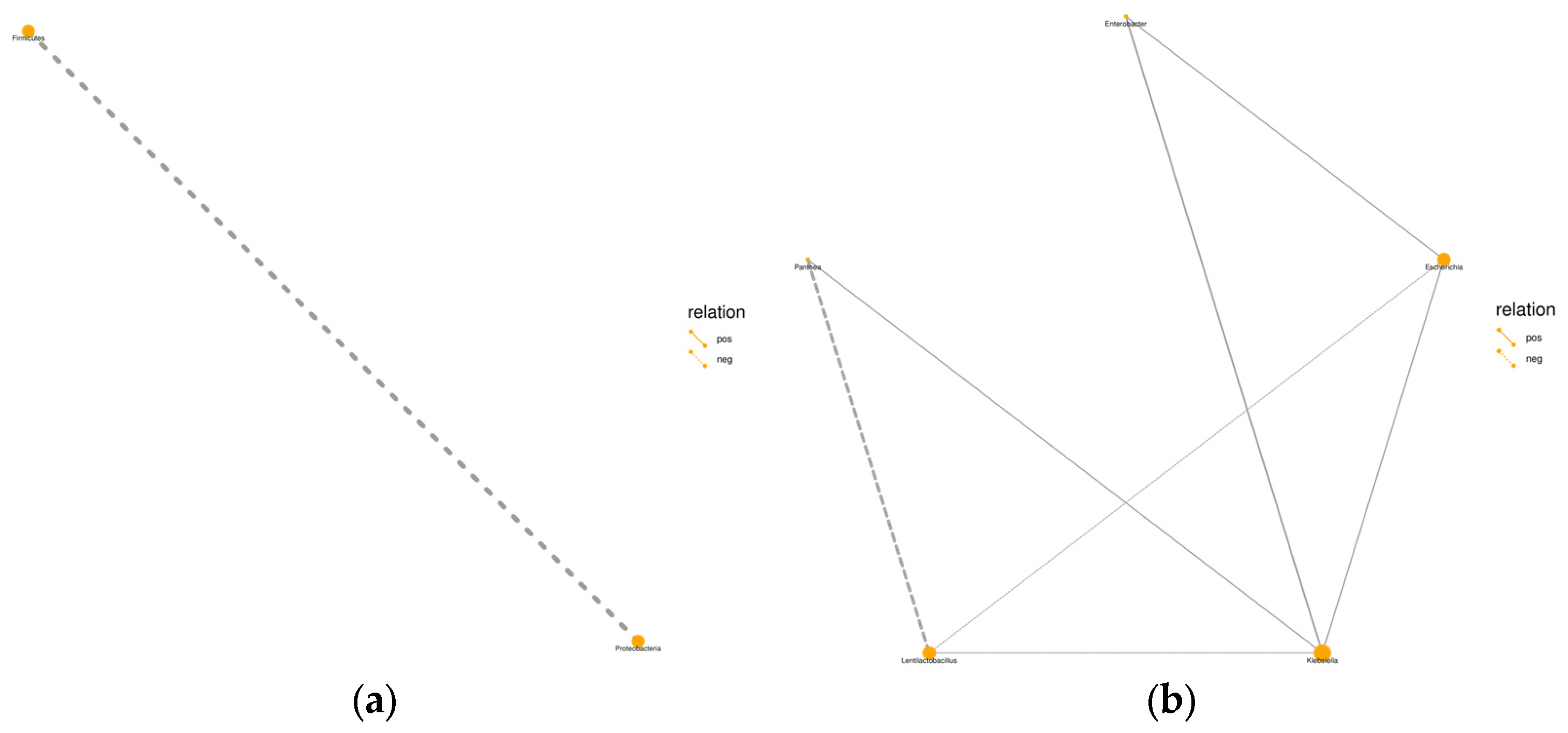
Figure 3.
Relationship between bacterial communities in whole-plant millet silage. (a) Phylum level, (b) genus level. Note: pos: positive impact; neg: negative impact.
At the genus level, the raw material was dominated by Pseudomonas, Pantoea, and Sphingomonas. However, post-ensiling, the CK group’s microbial composition shifted to being dominated by Klebsiella, Pantoea, and Enterobacter. The additive treatments significantly altered these distributions, with Lentilactobacillus becoming the dominant genus in the LP, S, and MIX groups, showing a marked ability to suppress undesirable genera such as the Klebsiella and Pantoea found in the CK group (Figure 2 and Figure 3b). Additionally, Pantoea was detected in all treatment groups, with reduced relative abundances compared to the raw material (CK: 11.79%, LP: 0.84%, S: 3.40%, MIX: 0.86%). Notably, after the aerobic stability test, Bacillus in the S group increased dramatically (from 0.04% at silo opening to 36.6%), dominating the microbial community (Figure 2).
4. Discussion
4.1. Effect of Different Growth Stages on Nutritional Components of Whole-Plant Millet Silage
The degree of maturity at harvest significantly influences the yield, nutritional content, digestibility, and ensiling potential of crops [16]. As the growth stage advances, changes in some nutritional values of silage are mostly associated with the degree of seed development and vary according to the different types and varieties of forage crops. Among these, the DM content is a crucial factor affecting silage quality. In this experiment, as the growth stage progressed, the DM content in all treatment groups gradually increased. Khan et al. studied the effects of the pre-heading, heading, and milk stages on the quality of millet silage and found that the DM content increased with the maturity of the millet, which is consistent with our findings [17]. By the time of full maturity, the DM content reached about 40%, as the water content in the plants decreased and sugars in the seeds were converted into starch, thereby increasing the DM content in the seed portion and consequently the whole plant [18,19]. A high DM content can complicate the compaction process during ensiling, leaving more air, which promotes the growth of yeasts and molds, thereby increasing the risk of “secondary fermentation” during feeding. Conversely, a low DM content, indicative of high moisture, can lead to excessive propagation of clostridia, which convert lactic acid into butyric acid, degrading silage quality. This process also triggers effluent loss, resulting in the depletion of WSCs and proteins and ultimately reducing intake by livestock such as cattle and sheep.
Hassanat et al. studied millet at the vegetative and heading stages and found that the cell wall components of the plant significantly increased with the extended growth period. The maximum values for NDF, ADF, cellulose, and lignin were found in the stems, followed by the whole plant and then the leaves. In contrast, hemicellulose, cutin, and silica content were higher in the leaves of the plant [9]. Additionally, KILIÇALP et al. studied oat silage at different growth stages and showed that the NDF content increases as the growth stage advances [20]. This difference may be attributed to the varying proportions of stems, leaves, and grains in different varieties, with the ADF and NDF contents being lower in leaves and grains compared to stems [21]. In this study, during the maturation process of forage millet, the NDF content tends to decrease. It is highest at the heading stage, remains relatively stable during the milk and dough stages, and significantly decreases by the mature stage. This trend may be due to the continuous deposition of starch in the grains during the filling process, which leads to a relative decrease in the cellulose content.
As the whole-plant millet matures, the CP content initially decreases until the filling stage, then increases, with the CP content at the dough stage being significantly higher than at the filling and milk stages. Jorge et al. found that the CP content decreases as the growth stage progresses, which is consistent with our results, which showed a decline in the CP content from the heading to the milk ripening stages [22]. This pattern is echoed in studies by Kurniawan [23], which observed a continuous accumulation of CP in whole-plant millet silage from the pre-milk to dough stages. Ferraretto [24] suggests that an increase in starch content within the silage reduces the proportions of both NDF and CP in the whole plant, explaining why the NDF and CP contents are lower during and after the dough stage compared to the heading stage. However, Yin reports a gradual decrease in CP content from the filling to the dough stage, which may be attributed to varietal differences. This complex interaction between growth stages and CP content highlights the dynamic nature of plant biochemistry in response to developmental changes.
WSC is a digestible part of the feed that is directly proportional to the feed’s digestibility and nutritional value. In this study, the WSC content gradually increased from the heading to the dough stages, reaching its peak at the dough stage and then is rapidly decreased by full maturity. This pattern is similar to Chu’s [25] findings on the impact of irrigation on WSC content and grain yield in different wheat varieties, where the WSC content continuously increased from the jointing to flowering stages and then rapidly decreased during the grain-filling stage. The accumulation of WSCs is closely linked to photosynthesis, which accumulates WSCs through photosynthesis as the plant matures. In the later stages of growth, as light intensity decreases, the rate of photosynthesis drops, leading to a reduction in soluble sugar accumulation [26,27]. Additionally, during the late growth stages, leaf senescence occurs, during which chlorophyll and other cellular components break down, releasing nitrogen and carbohydrates [28] and further contributing to the decline in WSC content as the growth period concludes.
In this study, the numbers of LAB and the contents of LA and AA are significantly higher at the dough stage compared to the milk stage, while the number of general bacteria is significantly lower. LAB efficiently convert WSCs into organic acids such as LA and AA under anaerobic conditions, which helps lower the pH and inhibits the growth of harmful microbes. Lactic acid, in particular, is the most effective organic acid in reducing pH in the silage environment, being 10–12 times more acidic than other organic acids [29]. Additionally, as the growth stage progresses, the number of general bacteria initially increase and then decrease, peaking during the milk stage. These general bacteria compete with LAB for fermentation substrates, leading to energy losses in the silage [30]. Consequently, selecting a growth stage with appropriate DM content is crucial to control moisture and air in the silage, thus reducing the number of general bacteria and enhancing silage quality. The LA content typically increases with the WSC content. However, this correlation was not observed in Experiment 1. This discrepancy could be due to several factors. Variations in the types and activities of the microorganisms present can affect the fermentation process. Different strains of lactic acid bacteria may have varying efficiencies in converting WSCs to lactic acid. Temperature, pH, and moisture content can influence microbial activity and fermentation efficiency. Additionally, other nutrients essential for microbial growth and activity may be limited, affecting the overall fermentation process and the production of LA.
The LA/AA ratio is commonly used as a qualitative indicator of fermentation and the aerobic stability of silage. A higher LA/AA ratio often indicates a more efficient fermentation process, as lactic acid is a stronger acid and more effective at lowering pH, which preserves the silage. A balanced or lower LA/AA ratio can improve the aerobic stability of silage. While lactic acid is important for initial preservation, AA plays a crucial role in inhibiting the growth of spoilage organisms when the silage is exposed to air. It is generally recommended that the concentration of AA should be around 3% DM [31]. Ahtoh et al. found that at low levels of AA (below 4.1% DM), nutritional losses can be highly variable [32]. Dnaiel et al. reported that adding 5% DM of AA to the daily diet of dairy cows resulted in lower feed intake within 4 h of the morning feeding, but no differences in intake were observed over the entire day. Additionally, the average milk yield and milk quality throughout the study period did not differ from the control group [33]. H. Danner et al. indicated that the AA content required for successful silage preservation can be very high. Silage with AA concentrations exceeding 5% DM remained stable for over 100 h, whereas untreated silage with an AA concentration of 1.28% DM remained stable for only 35 h [34]. Therefore, an appropriate amount of acetic acid can help prevent spoilage during feed-out. Additionally, both acids influence the palatability of the silage. Excessive AA might reduce palatability and intake by animals due to its strong vinegar-like smell and taste. However, moderate levels can be beneficial for stability without compromising intake.
4.2. Effect of Different Additives on the Quality of Whole-Plant Millet Silage
At silo open, the pH levels in the LP, S, and MIX additive treatment groups were significantly lower than in the CK group, indicating that the additives positively influenced the preservation of the silage, leading to enhanced fermentation quality [26]. Moreover, the MIX group exhibited superior characteristics, with a pH value of 3.96, an LA content of 6.59% DM and a better LAB count and LA/AA ratio compared to the other two additive groups. In the LP group, homolactic fermentation LAB rapidly produced a significant amount of LA using the WSCs available in the raw materials at the beginning of fermentation, thereby lowering the pH in the silage environment. Adding sucrose provided a fermentation substrate beneficial for LAB proliferation, facilitating further acid production and pH reduction [35]. The S group had the highest DM content (38.5%), which was significantly higher than the CK group, which aligns with similar findings reported by Kang [36]. The addition of sucrose enhanced the early production of LA during ensiling, rapidly reducing the silage pH and thereby inhibiting harmful microbial activity and minimizing nutrient losses [37]. Furthermore, sucrose itself contributes to a high DM content [38]; the MIX group had a lower DM content compared to the S group, likely due to the additional use of LP which consumes nutrients, thus reducing its DM content.
The additive groups also showed lower NDF and ADF contents than the CK group, similar to the findings by You [39], potentially due to the presence of fibrolytic enzyme-producing microbes that reduce fiber content [40]. Lentilactobbacillus buchneri has been shown to produce ferulic acid esterase—a part of the hemicellulose-degrading enzyme system that breaks ester bonds between hemicelluloses and lignin—enhancing cellulose utilization [41]. Research by Bao [42] and others indicates that LAB can produce a significant amount of cellulase during fermentation, reducing the fiber content in silage and enhancing the quality. Moreover, the organic acids produced during ensiling can hydrolyze digestible parts of the cell walls, further reducing fiber content [43]. Additionally, Zhang et al. found that Lactiplantibacillus plantarum also has the ability to degrade cellulose [44].
4.3. Effect of Different Additives on the VFA Contents and Aerobic Stability of Whole-Plant Millet Silage
During the 7-day (168 h) monitoring of aerobic stability, it was observed that the temperature of all groups did not exceed room temperature by more than 2 °C. However, the contents of LA and AA in all treatment groups (except for group S) decreased after the aerobic stability test and the pH levels increased, though they remained significantly lower than those in the CK group. Upon aerobic exposure, air penetrates the silage, promoting the rapid growth and reproduction of yeasts and general bacteria, which utilize the remaining sugars and LA as fermentation substrates [45,46]. The AA produced by heterofermentative LAB metabolism in the silage has antifungal activity, which can enhance the aerobic stability of the silage [47,48]. Additionally, the experiment was conducted in winter, when the indoor temperature was relatively low (around 20 °C), which may have influenced the aerobic stability results.
Studies have found that the LA content in Experiment 2 is lower than in Experiment 1. This may be due to Experiment 2 being conducted in the second year based on the results of Experiment 1. Owing to differences in the climate and other environmental factors, the millet grown in different years may have varying growth conditions and durations, leading to differences in the composition and quality of silage materials. In Experiment 1, the WSC content of the dough stage raw material was 92.7 ± 10.4 g/kg DM, while in Experiment 2, it was 21.3 ± 3.96 g/kg DM. This difference in WSC content is likely the main reason for the variation in lactic acid content between the two experiments. The higher acetic acid content in the additive group is related to the significantly higher presence of Lentilactobacillus in the microbial composition compared to the control group. Studies have shown that Lentilactobacillus, such as Lentilactobacillus buchneri, can produce more AA [45]. Therefore, even though we added Lactiplantibacillus plantarum, the AA content in the additive group was significantly higher than in the control group.
Yeasts can convert organic matter into ethanol, which is then converted to acetic acid by acetic acid bacteria [49]. Under the stimulation of acetic acid, yeast cells can harden their cell walls to limit the entry of acetates, thereby increasing their tolerance to acetic acid [50]. This might explain why both the yeast content and AA (acetic acid) levels are relatively high in our experiment. Additionally, Cai et al. found that adding Lactobacillus plantarum to silage improved fermentation quality but did not inhibit the growth of yeasts in the silage [51]. This is consistent with the findings of our study.
Lactic acid bacteria are capable of generating amylase, which hydrolyzes starch into dextrin and ultimately into glucose. They utilize glucose as a carbon source to generate pyruvate through glycolysis and subsequently produce lactic acid under the action of lactic dehydrogenase [52]. Menglei Xia et al. demonstrated that acetic acid bacteria and lactic acid bacteria were negatively correlated during the fermentation process and that the metabolism of acetic acid bacteria inhibited the growth and metabolism of lactic acid bacteria [53]. Based on this determination, the relatively high concentration of acetic acid in Experiment 2 of this study might inhibit the activity of lactic acid bacteria and decrease the efficiency of lactic acid conversion, thereby retaining more WSCs.
4.4. Effect of Different Additives on the Bacterial Communities of Whole-Plant Millet Silage
The Goods_coverage index reached 1.00 in all groups, suggesting that the sequencing depth was sufficient to reliably analyze the bacterial communities. Post-ensiling, a decrease in the Chao1, Shannon, and Simpson indices across all treatment groups indicated a reduction in microbial diversity. This trend aligns with findings by Xin et al. [54], which attribute the reduction in diversity mainly to the acidic anaerobic environment of the silage inhibiting the growth of some microbes. Similarly, Wayne Polley et al. [55] reported that the microbial diversity tends to decrease when beneficial bacteria are abundant. In contrast, studies by Wang [56] and Ni [57] on soy and moringa leaf silage observed an increase in bacterial diversity post-ensiling, potentially influenced by the specific plant materials used. PCA analysis showed that the bacterial communities in the raw material were distinctly separated from those in the silage across all treatment groups, highlighting differences in microbial composition due to the ensiling process.
Microbial abundance analysis showed that Proteobacteria and Firmicutes dominated both the raw material and all treatment groups. Specifically, Proteobacteria accounted for 88.6% and 91.6% in the CK and raw material groups, respectively, while Firmicutes had relative abundances of 5.16% and 11.3%. These phyla likely flourished due to the low pH and anaerobic conditions favorable for their growth in silage. The use of additives significantly increased the relative abundance of Firmicutes, suggesting that additives can effectively alter the silage bacterial community. This observation aligns with the findings of Yuan [58] and Wang [59] and is possibly due to the increase in the Lentilactobacillus and Lactiplantibacillus genera in the additive groups, both of which belong to the Firmicutes phylum. During the silage fermentation process, Lentilactobacillus and Lactiplantibacillus were the dominant genera in the additive groups, differing from previous research results [60]. Both genera belong to the LAB group and played a dominant role during the silage fermentation process, initiating LA fermentation and improving fermentation quality. Klebsiella, a common pathogen in silage, can cause DM loss and produce carbon dioxide, also leading to mastitis and reduced milk production [61]; Enterobacter competes with LAB for sugars, degrades proteins, ferments AA, lowers the quality of silage, and produces toxic compounds [57,62]. Compared to the CK group, the relative abundances of Klebsiella and Enterobacter genera decreased in all three additive groups, with the most significant reductions observed in the LP and MIX groups. This decrease supports the findings of Zhang [63] and Jiang [64], highlighting the effectiveness of these additives in suppressing undesirable bacteria in silage.
Additives can inhibit the growth of harmful microbes in silage, with LP showing a superior effect compared to adding sucrose alone. The reduction in Enterobacter in silage reflects a comprehensive change across various factors, including the availability of nutrients and water, the efficiency of converting nutrients to fermentation products, the activity of LAB, pH levels, and temperature—all contributing to more favorable conditions within the silage environment [65]. Post-ensiling, the relative abundance of the Pantoea genus in all treatment groups decreased compared to the raw material. This is consistent with the findings of Cheng [66]. The role of the Pantoea genus in silage remains ambiguous. Research, such as that by Ogunade [67] and Zhang [68], suggests that the Pantoea genus is negatively correlated with ammonia nitrogen content, reducing ammonia nitrogen content and helping to preserve proteins. Conversely, Li [69] and You [39] consider the Pantoea genus as an undesirable microbe in silage as it competes with LAB for nutrients. Given these conflicting roles, further research is necessary to clarify the function of microbes like Pantoea in silage, potentially paving the way for their development as microbial inoculants.
5. Conclusions
Harvesting whole-plant millet at the dough stage and applying a combined additive of Lactiplantibacillus plantarum and sucrose can improve fermentation quality and aerobic stability by optimizing microbial composition.
Author Contributions
N.Z. and X.H.: Both authors contributed equally to the original drafting of the manuscript, methodology formulation, conducting investigations, performing formal analysis, and curating data. M.Y.: Contributed to reviewing and curating data and conducting investigations. C.L.: Responsible for experimental design and manuscript review. C.W.: Acquired funding, curated data, and contributed to editing. H.H.: Involved in editing, supervision, funding acquisition, data curation, and conceptualization of the research. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “the earmarked fund for China Agriculture Research System, CARS-38”.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lu, H.; Zhang, J.; Liu, K.B.; Wu, N.; Li, Y.; Zhou, K.; Ye, M.; Zhang, T.; Zhang, H.; Yang, X.; et al. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago. Proc. Natl. Acad. Sci. USA 2009, 106, 7367–7372. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.M.; Sebola, N.A.; Mabelebele, M. The nutritional use of millet grain for food and feed: A review. Agric. Food Secur. 2021, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Hao, X.; Li, Y.; Zhang, Q.; Wang, C.; Han, H. Microbial Communities and Metabolites of Whole Crop Corn Silage Inoculated with Lentilactobacillus plantarum and Lentilactobacillus buchneri. Processes 2022, 10, 2369. [Google Scholar] [CrossRef]
- Costa, R.; Costa, K.; Souza, W.; Epifânio, P.; Santos, C.; Silva, J.; Oliveira, S. Production and quality of silages pearl millet and paiaguas palisadegrass in monocropping and intercropping in different forage systems. Biosci. J. 2018, 34, 957–967. [Google Scholar] [CrossRef]
- Amer, S.; Mustafa, A.F. Short communication: Effects of feeding pearl millet silage on milk production of lactating dairy cows. J. Dairy Sci. 2010, 93, 5921–5925. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.D.; Neves, A.L.A.; Pereira, L.G.R.; Sollenberger, L.E.; Muniz, E.N.; Souza, E.Y.B.; Sobral, A.J.S.; Costa, N.V.; Gonçalves, L.C. Performance, agronomic traits, ensilability and nutritive value of pearl millet cultivar harvested at different growth stages. J. Agric. Sci. 2020, 158, 225–232. [Google Scholar] [CrossRef]
- Hill, G.M.; Utley, P.R.; Gates, R.N.; Hanna, W.W.; Johnson, J.C., Jr. Pearl Millet Silage for Growing Beef Heifers and Steers. J. Prod. Agric. 1999, 12, 653–658. [Google Scholar] [CrossRef]
- Nussio, L.G.; Park, R.S.; Stronge, M.D. Silage Production from Tropical Forages; Wageningen Academic: Wageningen, The Netherlands, 2005. [Google Scholar]
- Hassanat, F.; Mustafa, A.F.; Seguin, P. Effects of inoculation on ensiling characteristics, chemical composition and aerobic stability of regular and brown midrib millet silages. Anim. Feed. Sci. Technol. 2007, 139, 125–140. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Rinne, M.; Franco, M.; Tapio, I.; Stefanski, T.; Bayat, A.; Mäntysaari, P. Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows. Agriculture 2022, 12, 266. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Xie, Y.; Zhang, Y.; Lin, Y.; Zheng, Y.; Yang, X.; Wang, N.; Ni, K.; Yang, F. Effect of Sucrose and Lactic Acid Bacteria Additives on Fermentation Quality, Chemical Composition and Protein Fractions of Two Typical Woody Forage Silages. Agriculture 2021, 11, 256. [Google Scholar] [CrossRef]
- Kasmaei, K.M.; Dicksved, J.; Spörndly, R.; Udén, P. Separating the effects of forage source and field microbiota on silage fermentation quality and aerobic stability. Grass Forage Sci. 2017, 72, 281–289. [Google Scholar] [CrossRef]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef]
- Fang, D.; Dong, Z.; Wang, D.; Li, B.; Shi, P.; Yan, J.; Zhuang, D.; Shao, T.; Wang, W.; Gu, M. Evaluating the fermentation quality and bacterial community of high-moisture whole-plant quinoa silage ensiled with different additives. J. Appl. Microbiol. 2022, 132, 3578–3589. [Google Scholar] [CrossRef] [PubMed]
- Buxton, D.R.; O’Kiely, P. Preharvest Plant Factors Affecting Ensiling. In Silage Science and Technology; Agronomy Monographs; Springer: Cham, Switzerland, 2003; pp. 199–250. [Google Scholar]
- Khan, S.H.; Azim, A.; Sarwar, M.; Khan, A.G. Effect of maturity on comparative nutritive value and fermentation characteristics of maize, sorghum and millet silage. Pak. J. Bot. 2011, 43, 2967–2970. [Google Scholar]
- Allen, M.; Coors, J.G.; Roth, G. Silage Science and Technology. Agronomy Monograph 42. Corn Silage 2003, 42, 547–608. [Google Scholar]
- Oskey, M.; Velasquez, C.; Peña, O.M.; Andrae, J.; Bridges, W.; Ferreira, G.; Aguerre, M.J. Yield, nutritional composition, and digestibility of conventional and brown midrib (BMR) pearl millet as affected by planting and harvesting dates and interseeded cowpea. Animals 2023, 13, 260. [Google Scholar] [CrossRef]
- Kiliçalp, N. Cultivar and Harvest Stage Effects on Nutritive Value of Whole Crop Oat (Avena sativa L.) Silages. J. Agric. Nat. 2022, 26, 437–449. [Google Scholar] [CrossRef]
- Hassanat, F.; Mustafa, A.F.; Seguin, P. Effect of the brown midrib trait and stage of development at harvest on cell wall composition and degradability of forage pearl millet leaves and stems. Can. J. Anim. Sci. 2007, 87, 421–429. [Google Scholar] [CrossRef]
- Morales, J.U.; Alatorre, J.A.H.; Nieto, C.A.R.; José, F.; Becerra, C. Forage production and nutritional content of silage from three varieties of pearl millet (Pennisetum glaucum) harvested at two maturity stages. J. Anim. Plant Sci. 2015, 27, 4161–4169. [Google Scholar]
- Kurniawan, W.; Napirah, A.; Isnaeni, P.; Bain, A. Nutrient qualities hydroponic corn green fodder (HCGF) of yellow-maize on different harvest time. IOP Conf. Ser. Earth Environ. Sci. 2024, 1341, 012062. [Google Scholar] [CrossRef]
- Ferraretto, L.; Shaver, R.; Luck, B. Silage review: Recent advances and future technologies for whole-plant and fractionated corn silage harvesting. J. Dairy Sci. 2018, 101, 3937–3951. [Google Scholar] [CrossRef]
- Chu, P.F.; Yu, Z.W.; Wang, D.; Zhang, Y.L.; Xu, Z.Z. Effects of irrigation stage and amount on winter wheat fructan accumulation and translocation after anthesis and water use efficiency. Ying Yong Sheng Tai Xue Bao 2009, 20, 2691–2698. [Google Scholar]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Afzal, S.; Chaudhary, N.; Singh, N.K. Role of Soluble Sugars in Metabolism and Sensing Under Abiotic Stress. Plant Growth Regul. Signal. Under Stress Cond. 2021, 14, 305–334. [Google Scholar]
- Webster, J. The Biochemistry of Silage; McDonald, P., Henderson, A.R., Heron, S.J.E., Marlow, B., Eds.; Chalcombe Publications: Southampton, UK, 1991; p. 340. ISBN 0-948617-225. [Google Scholar]
- Kung, L. Silage fermentation and additives. Arch. Latinoam. Prod. Anim. 2018, 26, 61–66. [Google Scholar]
- Gerlach, K.; Daniel, J.L.P.; Jobim, C.C.; Nussio, L.G. A data analysis on the effect of acetic acid on dry matter intake in dairy cattle. Anim. Feed. Sci. Technol. 2021, 272, 114782. [Google Scholar] [CrossRef]
- Maмaeв, A.; Ocипян, Б.; Koзлoвa, B. Role of acetic acid in the sustainability of silage from cereal grasses to adverse micloflore at air access. Adapt. Fodd. Prod. 2019, 2019, 48–57. [Google Scholar] [CrossRef]
- Daniel, J.L.P.; Amaral, R.C.; Sá Neto, A.; Cabezas-Garcia, E.H.; Bispo, A.W.; Zopollatto, M.; Cardoso, T.L.; Spoto, M.H.F.; Santos, F.A.P.; Nussio, L.G. Performance of dairy cows fed high levels of acetic acid or ethanol. J. Dairy Sci. 2013, 96, 398–406. [Google Scholar] [CrossRef]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Zi, X.; Liu, Y.; Chen, T.; Li, M.; Zhou, H.; Tang, J. Effects of Sucrose, Glucose and Molasses on Fermentation Quality and Bacterial Community of Stylo Silage. Fermentation 2022, 8, 191. [Google Scholar] [CrossRef]
- Kang, J.; Tang, S.; Zhong, R.; Tan, Z.; Wu, D. Alfalfa Silage Treated with Sucrose Has an Improved Feed Quality and More Beneficial Bacterial Communities. Front. Microbiol. 2021, 12, 670165. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Zhang, Z.X.; Shimojo, M.; Wang, T.; Masuda, Y. Comparison of Fermentation Characteristics of Italian Ryegrass (Lolium multiflorum Lam.) and Guineagrass (Panicum maximum Jacq.) during the Early Stage of Ensiling. Asian-Australas. J. Anim. Sci. 2005, 18, 1727–1734. [Google Scholar] [CrossRef]
- Rezaei, J.; Rouzbehan, Y.; Fazaeli, H. Nutritive value of fresh and ensiled amaranth (Amaranthus hypochondriacus) treated with different levels of molasses. Anim. Feed. Sci. Technol. 2009, 151, 153–160. [Google Scholar] [CrossRef]
- You, J.; Zhang, H.; Zhu, H.; Xue, Y.; Cai, Y.; Zhang, G. Microbial Community, Fermentation Quality, and In Vitro Degradability of Ensiling Caragana with Lactic Acid Bacteria and Rice Bran. Front. Microbiol. 2022, 13, 804429. [Google Scholar] [CrossRef]
- He, L.; Chen, N.; Lv, H.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef]
- Nsereko, V.L.; Smiley, B.K.; Rutherford, W.M.; Spielbauer, A.; Forrester, K.J.; Hettinger, G.H.; Harman, E.K.; Harman, B.R. Influence of inoculating forage with lactic acid bacterial strains that produce ferulate esterase on ensilage and ruminal degradation of fiber. Anim. Feed. Sci. Technol. 2008, 145, 122–135. [Google Scholar] [CrossRef]
- Bao, J.; Wang, L.; Yu, Z. Effects of Different Moisture Levels and Additives on the Ensiling Characteristics and In Vitro Digestibility of Stylosanthes Silage. Anim. Open Access J. 2022, 12, 1555. [Google Scholar] [CrossRef]
- Larsen, S.U.; Hjort-Gregersen, K.; Vazifehkhoran, A.H.; Triolo, J.M. Co-ensiling of straw with sugar beet leaves increases the methane yield from straw. Bioresour. Technol. 2017, 245 Pt A, 106–115. [Google Scholar] [CrossRef]
- Zhang, H.; Wen, B.; Liu, Y.; Du, G.; Wei, X.; Imam, K.M.S.U.; Zhou, H.; Fan, S.; Wang, F.; Wang, Y.; et al. A reverse catalytic triad Asp containing loop shaping a wide substrate binding pocket of a feruloyl esterase from Lactobacillus plantarum. Int. J. Biol. Macromol. 2021, 184, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Mugabe, W.; Shao, T.; Li, J.; Dong, Z.; Yuan, X. Effect of hexanoic acid, Lactobacillus plantarum and their combination on the aerobic stability of napier grass silage. J. Appl. Microbiol. 2020, 129, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.J.; Wang, J.; Guo, G.; Wen, A.Y.; Desta, S.T.; Shao, T. Effects of ethanol, molasses and Lactobacillus plantarum on fermentation characteristics and aerobic stability of total mixed ration silages. Grass Forage Sci. 2016, 71, 328–338. [Google Scholar] [CrossRef]
- Dong, Z.; Li, J.; Wang, S.; Dong, D.; Shao, T. Time of Day for Harvest Affects the Fermentation Parameters, Bacterial Community, and Metabolic Characteristics of Sorghum-Sudangrass Hybrid Silage. Msphere 2022, 7, e00168-22. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, E.; Shapiro, O.H.; Siboni, N.; Kushmaro, A. Advantage of Using Inosine at the 3′ Termini of 16S rRNA Gene Universal Primers for the Study of Microbial Diversity. Appl. Environ. Microbiol. 2006, 72, 6902–6906. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, D.; Niu, D.; Zhao, Y. Acetic acid production from food wastes using yeast and acetic acid bacteria micro-aerobic fermentation. Bioprocess Biosyst. Eng. 2015, 38, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Vitorino, M.V.; Godinho, C.P.; Bourbon-Melo, N.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Yeast adaptive response to acetic acid stress involves structural alterations and increased stiffness of the cell wall. Sci. Rep. 2021, 11, 12652. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics and aerobic deterioration of silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Xia, M.; Zhang, X.; Xiao, Y.; Sheng, Q.; Tu, L.; Chen, F.; Yan, Y.; Zheng, Y.; Wang, M. Interaction of acetic acid bacteria and lactic acid bacteria in multispecies solid-state fermentation of traditional Chinese cereal vinegar. Front. Microbiol. 2022, 13, 964855. [Google Scholar] [CrossRef]
- Xin, Y.; Chen, C.; Zhong, Y.; Bu, X.; Huang, S.; Tahir, M.; Du, Z.; Liu, W.; Yang, W.; Li, J.; et al. Effect of storage time on the silage quality and microbial community of mixed maize and faba bean in the Qinghai-Tibet Plateau. Front. Microbiol. 2022, 13, 1090401. [Google Scholar] [CrossRef]
- Wayne Polley, H.; Wilsey, B.J.; Derner, J.D. Dominant species constrain effects of species diversity on temporal variability in biomass production of tallgrass prairie. Oikos 2007, 116, 2044–2052. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.-Y.; Chen, X.-Y.; Zhang, Q. Effects of Wilting and Lactobacillus plantarum Addition on the Fermentation Quality and Microbial Community of Moringa oleifera Leaf Silage. Front. Microbiol. 2018, 9, 1817. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.-A.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Yuan, X.; Li, J.; Dong, Z.; Shao, T. The reconstitution mechanism of napier grass microiota during the ensiling of alfalfa and their contributions to fermentation quality of silage. Bioresour. Technol. 2020, 297, 122391. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, M.; Wu, S.; Zou, X.; Chen, X.; Ge, L.; Zhang, Q. Effects of Gallic Acid on Fermentation Parameters, Protein Fraction, and Bacterial Community of Whole Plant Soybean Silage. Front. Microbiol. 2021, 12, 662966. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, H.-J.; Yu, Z. Effects of sucrose, formic acid and lactic acid bacteria inoculant on quality, in vitro rumen digestibility and fermentability of drooping wild ryegrass (Elymus nutans Griseb.) silage. J. Anim. Feed. Sci. 2017, 26, 26–32. [Google Scholar] [CrossRef]
- Lv, H.; Pian, R.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of citric acid on fermentation characteristics and bacterial diversity of Amomum villosum silage. Bioresour. Technol. 2020, 307, 123290. [Google Scholar] [CrossRef]
- Muck, R.E. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef]
- Zhang, T.; Li, L.; Wang, X.; Zeng, Z.; Hu, Y.; Cui, Z.-J. Effects of Lactobacillus buchneri and Lactobacillus plantarum on fermentation, aerobic stability, bacteria diversity and ruminal degradability of alfalfa silage. World J. Microbiol. Biotechnol. 2009, 25, 965–971. [Google Scholar] [CrossRef]
- Jiang, F.-G.; Cheng, H.-J.; Liu, D.; Wei, C.; An, W.-J.; Wang, Y.-F.; Sun, H.-T.; Song, E.-L. Treatment of Whole-Plant Corn Silage with Lactic Acid Bacteria and Organic Acid Enhances Quality by Elevating Acid Content, Reducing pH, and Inhibiting Undesirable Microorganisms. Front. Microbiol. 2020, 11, 593088. [Google Scholar] [CrossRef] [PubMed]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.O.; Spoelstra, S.F. Microbiology of Ensiling. In Silage Science and Technology; American Society of Agronomy: Madison, WI, USA, 2003; pp. 31–93. [Google Scholar]
- Cheng, Q.; Chen, L.; Chen, Y.; Li, P.; Chen, C. Effects of LAB Inoculants on the Fermentation Quality, Chemical Composition, and Bacterial Community of Oat Silage on the Qinghai-Tibetan Plateau. Microorganisms 2022, 10, 787. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Jiang, Y.; Pech Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Wang, Z.; Bao, J.; Zhao, M.; Si, Q.; Sun, P.; Ge, G.; Jia, Y. Effects of Different Types of LAB on Dynamic Fermentation Quality and Microbial Community of Native Grass Silage during Anaerobic Fermentation and Aerobic Exposure. Microorganisms 2023, 11, 513. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yuan, Z.; Sun, Y.; Kong, X.; Dong, P.; Zhang, J. A reused method for molasses-processed wastewater: Effect on silage quality and anaerobic digestion performance of Pennisetum purpereum. Bioresour. Technol. 2017, 241, 1003–1011. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).