Abstract
Rabbit meat production faces challenges due to the prevalence of gastrointestinal diseases in rabbits, exacerbated by restrictions on antibiotic use in European animal production. Marine macroalgae, rich in bioactive compounds such as soluble polysaccharides, represent promising solutions to this problem. However, research on the effects of macroalgae and the underlying mechanisms in rabbits is limited, especially in commercial settings. This study aimed to evaluate the impact of Saccharina latissima (dehydrated) and Ulva lactuca (dehydrated and hydrolyzed extract) on rabbit on growth performance and gut health in a commercial farm context. A total of 96 litters (8 rabbits/litter) of crossbred rabbits weaned at 33 days of age were randomly assigned to 4 experimental groups (control, Saccharina latissima dehydrated, Ulva lactuca dehydrated and Ulva lactuca hydrolyzed extract; 24 replicates/treatment) and monitored from weaning to slaughter at 61 days of age. The key indicators of gut health were assessed 14 days post-weaning by counting coccidia, isolating specific microflora and examining histological samples. Additionally, the relevant intestinal markers (microbiome composition, mucin content and gene expression related to immune response and tight junction proteins) were determined in order to elucidate the potential mechanisms involved. The inclusion of macroalgae in the diet did not influence growth performance of the animals. S. latissima had a positive effect in reducing coccidia counts (p = 0.10) and improving mucosal morphology (p < 0.001), which can possibly be attributed to modulation of the microbiota and improved mucosal functionality. Ulva lactuca had a favorable effect on gut tight junction proteins (p < 0.001), enhancing intestinal barrier function. These findings suggest the potential of macroalgae to modify the intestinal microbiome by reducing the presence of inflammatory bacteria. Further research is warranted to elucidate the mechanisms involved and optimize macroalgae supplementation in rabbit nutrition for enhanced gut health.
1. Introduction
Rabbits raised for meat production suffer from a high incidence of gastrointestinal diseases that cause significant mortalities in rabbits, especially in the post-weaning phase [1]. During this critical period, bacterial infections and coccidiosis are significant challenges that can lead to severe digestive disturbances, increased morbidity and higher mortality rates, necessitating the use of antibiotics for effective management [2]. The stress of weaning, combined with the susceptibility of young rabbits to pathogenic microorganisms, underscores the critical need for antibiotic interventions to maintain health and productivity. Controlling these diseases has become more difficult due to the recent restrictions on the use of preventive antibiotics in animal production in Europe [3]. The industry, therefore, urgently needs innovative alternatives to help enhance rabbit gut health and to provide protection against pathogens. The first line of action is to ensure biosecurity and health management; once these are optimized, it is important to formulate adequate, balanced diets.
Marine macroalgae are promising ingredients for animal feed as they are rich in numerous bioactive compounds, especially soluble polysaccharides, which have already been demonstrated to have some benefits on gut health in some livestock species [4,5]. Some macroalgae polysaccharides, e.g., laminarin and fucoidan, which are present at high concentrations in species such as Saccharina latissima and Laminaria spp., have been widely studied in pigs. When purified extracts of these polysaccharides were supplemented separately, laminarin at 300 mg/kg complete feed and fucoidan at 240–360 mg/kg, they enhanced the performance and gut health of piglets in the post-weaning phase [6,7]. These types of macroalgal extracts have been shown to exert some types of action on the gastrointestinal tract [5], such as improvement of nutrient digestibility, stimulation of digestive function, support for immune system function, stimulation of the establishment of beneficial gut microbes and reduction of pathogen loads. In rabbits, the previous studies have shown that the addition of some macroalgae polysaccharides from Laminaria spp. (extract combined with plant extracts) or dehydrated Ulva lactuca improved growth performance, feed conversion and feed digestibility [8,9]. However, there is still a gap in understanding the potential benefits of macroalgae on rabbit gut health and their mechanisms of action, especially within commercial farm systems.
This study aimed to evaluate the impact of incorporating three different macroalgae products into rabbit diets on both growth performance and gut health in a commercial farm setting. By focusing on gut health, this research explores whether these macroalgae can improve the gut microbiota composition and overall digestive health in rabbits, providing insights that differ from the previous studies by examining both the methodological approach and potential health benefits. Throughout the trial, the growth and mortality parameters were monitored to gauge the response of the rabbits to the experimental diets. Key indicators of gut health, including coccidia counts, specific microflora populations and histological features, were assessed 14 days post-weaning to explore the potential benefits of including macroalgae in the diets. Additionally, an in-depth analysis of various intestinal markers, such as microbiome composition, mucin content and gene expression related to immune response and tight junction proteins, was conducted to elucidate the potential mechanisms underlying the observed effects of macroalgae on gut health.
2. Materials and Methods
2.1. Experimental Trial
All experimental procedures complied with Spanish regulations for the care and use of animals in research (Spanish Royal Decree 53/2013; BOE, 2013). The Bioethics Committee of the Universidad Politécnica de Madrid, Spain, approved these procedures (protocol code 2021-002, approval date: 24 February 2021).
The macroalgae products were selected according to the results of in vitro characterization [10] and the previous results obtained in an experimental farm set-up [11]. Two macroalgae were tested: Saccharina latissima (sugar kelp, SL treatment) and Ulva spp. (sea lettuce) were both used in dehydrated form (U treatment), and Ulva spp. was also used as a hydrolyzed aqueous extract (EU treatment). The products were elaborated from discards produced in industrial processing and provided by Porto-Muíños S.L. (Cerceda, A Coruña, Spain). To prepare the dehydrated samples, the macroalgae were oven-dried (<40 °C, 4–5 days), ground and milled (Komodin K-160 P, Lleal, Granollers, Spain) to 1 mm particle size. The hydrolyzed aqueous extract was prepared using an adaptation of the enzymatic method described by Hardouin and coworkers [12]. Briefly, Ulva spp. were combined with water at a 1:10 ratio, along with 4% Celluclast enzyme (Novozymes, Bagsvaerd, Denmark). This mixture was incubated at 50 °C for 6 h, after which the enzyme activity was halted by raising the temperature to 90 °C for 15 min. Ulvans, the polysaccharides of interest, were then precipitated using ethanol at a 1:4 ratio, and the resulting solution was filtered. The supernatant was discarded, and the remaining solid was dried and ground. Details on the chemical composition of the macroalgae products examined in this study are available elsewhere [10].
Four diets were tested: a control diet and diets elaborated with each of the three macroalgae products. The control diet was formulated in general agreement with the nutritional standards for growing rabbits [13], and its composition is shown in Table 1. The diets to be tested were obtained after the addition of macroalgae to the control diet (Table 1) at the following doses: SL at 0.25%, U at 1% and EU at 0.04%. The chemical composition of each experimental diet is shown in Table 2. The doses of inclusion of the macroalgae products were low and different depending on the species. In the case of SL, due to its high iodine concentration, the inclusion level was adjusted to avoid exceeding the maximum limit of iodine allowed by EU legislation in complete feed (Regulation EC 1459/2005). The EU was used at a lower dose than the U as it contained more of the polysaccharides of interest (ulvans).

Table 1.
Composition of the control diet (%).
Table 2.
Chemical composition of experimental diets (% as fed).
A total of 96 litters (8 rabbits/litter) of crossbred mixed-sex rabbits (New Zealand White X Californian) weaned at 33 d of age were randomly assigned to each experimental group. Each group was composed of 192 rabbits (24 litters/treatment), which were fed ad libitum with the diets between weaning (33 days of age, 905 ± 15.7 g) and slaughter (61 days of age, 2016 ± 20.8 g). The rabbits were housed collectively (8 rabbits per cage) in flat desk cages and had ad libitum access to water. Feeding troughs were shared by two litters in each experimental unit (12 experimental units, 2 litters/experimental unit per treatment for feed intake and feed conversion ratio).
An outbreak of an enteric disease of unidentified etiology occurred 10 days post-weaning, and the rabbits were treated with streptomycin for 4 days, as required on commercial farms.
2.2. Performance Parameters
Litter weight, feed intake and mortality were determined in rabbits in all experimental groups (n = 24 or 12), at age 33, 54 and 61 days, in order to calculate the average daily gain (ADG), average daily feed intake (ADFI) and feed conversion ratio (FCR). Feed intake was not recorded on day 61 for reasons beyond our control. Feed intake and feed conversion rate were calculated for every two cages as the rabbits shared feeding troughs (n = 12).
2.3. Sample Collection
At 47 days of age (14 days post-weaning), 8 rabbits from each group were randomly selected and slaughtered by head concussion and subsequent bleeding. The slaughter took place between 6 and 8 pm. Several samples were collected from each animal for the different types of analysis.
In order to prevent contamination, intestinal swabs were first collected from the proximal colon and ileocecal valve for Escherichia coli isolation. The intestinal swab specimens were transported to the laboratory in Amies medium and refrigerated until processing. For counting coccidia oocysts, feces was collected from the rectum and refrigerated until processing. For microbiome and Clostridium determination, the samples from the colon content were collected and frozen until processing. For crude mucin determination, intact samples of 20 cm of distal ileum digesta were collected and frozen. For gene expression analysis, 2 cm segments of the distal part of the cecal appendix were collected, cleaned with saline solution (NaCl 0.9%)—without touching the mucosa—and cut longitudinally. The samples were then scraped to obtain approximately 50 mg of mucosa, which was placed in vials containing 1 mL of RNA preserving solution and frozen at −80 °C [14]. For the morphometric study, samples of the jejunum of length 3 cm and of the ileocecal valve were collected and fixed in 10% buffered formalin for 24 h.
2.4. Histopathological Study and Morphometric Measures
Formalin-fixed samples of the jejunum and ileocecal valve were cut transversely in 0.5 cm pieces, which were routinely processed and placed in paraffin wax in individual cassettes. Histological sections of thickness 3 µm were cut from each sample, dewaxed, hydrated in a graded ethanol–water series and stained with hematoxylin-eosin and/or periodic acid–Schiff. The samples were examined in a light microscope (Olympus BX51, Olympus Iberia, Barcelona, Spain), and images for the histopathological study were captured with a camera attached to the microscope (Olympus EP50, Olympus Iberia, Barcelona, Spain). The morphometric measurements were made using ImageJ software (Version 1.54d). The length of villi and crypt depth were measured in 15 intact villi of the jejunum of each animal [15].
2.5. Coccidia Counts
The coccidia (Eimeria spp.) in feces were counted using the flotation technique with a saturated saline solution [16]. Briefly, samples of 3 g of feces were weighed in a plastic jar and 42 mL of water was added. Glass pearls were added to disaggregate the feces by shaking until homogenization. The contents of the jar were filtered, first through a 4 mm diameter mesh and then through a 150 µm diameter mesh. Two test tubes were filled with the filtrate and centrifuged at 2000 rpm for 10 min so that all Eimeria spp. oocysts were deposited at the bottom of the test tube. The supernatant was carefully removed, and the remaining sediment was homogenized by the addition of saturated saline solution to make up the volume of the test tube (15 mL). Two cells of the McMaster counting chamber were filled with the mixture, with the aid of a Pasteur pipette, and the solution was left for 2–3 min so that all oocysts floated to the top. The oocysts were then counted in a light microscope at 10X, in both cells of the chamber. The number of oocysts per gram of feces (opg) was calculated using the following equation:
2.6. E. coli Counts, EPEC and Antimicrobial Susceptibility Testing
Three approaches were used with E. coli as an indicator: E. coli counts, eae gene detection and antimicrobial susceptibility testing (AST) to determine multidrug resistance (MDR). Briefly, both swabs (colon and ileocecal content) were suspended in 9 mL of buffer peptone water (BPW; ApplyChem Panreac, Barcelona, Spain). One mL of the homogenate was inoculated on a 3M Petrifilm™ Select E. coli Count (SEC) plate, which was examined after incubation (24 h/44 °C) to determine the E. coli counts, following the manufacturer’s instructions. In addition, plates of three different selective agar media were each inoculated with 10 µL of the homogenate and incubated at 37 °C for 18–24 h: MacConkey lactose agar (LMAC, Oxoid, Thermo Fisher Scientific, Waltham, MA, USA), CHROMID® ESBL agar (bioMérieux, Inc., Hazelwood, MO, USA) and CHROMID® CARBA agar (bioMérieux, Inc., Hazelwood, MO, USA). The confluent growth on the LMAC plates was subjected to polymerase chain reaction (PCR) to detect the presence of the eae gene associated with EPEC. The positive confluents were further investigated to recover individual colonies, which were each individually analyzed by PCR. Finally, all isolates were confirmed as E. coli by PCR amplification of the β-d-glucuronidase-encoding gene (uidA) [17] stored in nutrient agar broth at 0.75% (Difco) at room temperature for further characterization. The PCR primers/conditions and methodology followed are described in detail elsewhere [18]. The minimal inhibitory concentrations (MICs) for the E. coli isolates were measured using the VITEK® 2 system (bioMérieux, Inc., Hazelwood, MO, USA) for conventional antimicrobial susceptibility testing (AST) with AST-GN96 test kit cards, according to the manufacturer’s instructions. The results of the antimicrobial susceptibility testing were evaluated based on the guidelines provided by the European Committee on AST (EUCAST 2023). Multidrug resistance (MDR) was defined according to the criteria proposed by Magiorakos et al. [19], as those showing acquired non-susceptibility to at least one agent in three or more antimicrobial categories, as previously described [19,20].
2.7. Determination of Clostridium Perfringens and Clostridioides Difficile by qPCR
The presence of Clostridium perfringens and Clostridioides difficile in fecal samples was determined by qPCR. The primers used to determine C. difficile were TPI Forward 5′CGTCATTAGCATCTTCAGCAGTTG 3′ and TPI Reverse 5′ GATGATTTAGCAAAAGTAGTTGTTGCA 3′ [21] and for C. perfringens were ClostP Forward 5′ TCTTACACGATGTATCAGAGGGT 3′ and ClostP Reverse 5′ ACATGTAGTCATCTGTTCCAGCA 3′ developed in this study by targeting the plc gene. Pure DNA isolated from C. perfringens CECT 376T and C. difficile CECT 531 T was used in the qPCR method. Serial dilutions of pure DNA were used for quantification by curve analysis. The qPCR reactions were composed of 5 μL of Power SYBR® Green PCR Mix (Applied BiosystemsTM, ThermoFisher Scientific, Waltham, MA, USA), 800 nM of each primer, 2 μL of DNA sample and made up to a total volume of 10 μL with water. The reaction was carried out in a QuantStudioTM 12k Flex real time PCR system (Applied BiosystemsTM, ThermoFisher Scientific, Waltham, MA, USA). The qPCR cycle included denaturalization for 10 min at 95 °C followed by 40 cycles of denaturalization at 95 °C for 15 s and annealing and extension at 60 °C for 1 min. This was followed by a melt curve analysis that included 60 °C for 1 min, followed by a temperature increase of 0.05 °C/s until reaching 95 °C.
2.8. Microbiome Analysis
Bacterial DNA was extracted from the colonic samples using the DNA Realpure Spin Food-Stool Kit® (Real, Durviz S.L., Valencia, Spain), following the manufacturer’s guidelines for fecal samples. The extracted DNA was quantified with a Qubit™4 fluorometer (Invitrogen, Thermo Fisher Scientific, Carlsbad, CA, USA) and the DNA HS Assay Kit (Invitrogen, Thermo Fisher Scientific, Eugene, OR, USA), then stored at −20 °C until it was needed for further analysis.
For the 16S rRNA amplicon sequencing, 2 μL of DNA from each sample was utilized to construct the libraries. The hypervariable regions of the 16S rRNA gene were amplified using two primer sets, v2-4–8 and v3-6, 7–9. Library preparation was carried out with the Ion 16STM Metagenomics and Ion XpressTM Plus Fragment Library Kits (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA). The PCR products were pooled with barcodes to create libraries, and the Ion XpressTM Barcode Adapters Kit (Life Technologies) was used for this purpose. The libraries were quantified with the Ion Universal Library Quantitation Kit (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA). Aliquots of 10 pM from each library were pooled and loaded onto an Ion OneTouch™ 2 System (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA), which performs template preparation and enrichment automatically. Template-positive ion sphere particles were then enriched using Dynabeads™ MyOne™ Streptavidin C1 magnetic beads (Invitrogen, Carlsbad, CA, USA) in an Ion One Touch ES instrument. Finally, the samples were loaded onto an Ion 520TM chip (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) in an Ion GeneStudioTM S5 System sequencer, using the Ion 520™ and Ion 530™ loading reagents from the OT2-Kit (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA).
The raw sequencing reads were obtained from the Torrent Suite software (v.5.12.2.) as fastq files for 16S rRNA amplicon sequencing. The fastq files were processed with QIIME 2 software v. 2022.11 [22]. To produce amplicon sequence variants (ASVs), the DADA2 method was used for quality filtration (Q score > 30), trimming, denoising and dereplication. Samples in which the total abundance of taxa was <10 (summed across all samples) were excluded from the analysis. The samples were rarefied to a sequencing depth of 20,000 reads. Taxonomy was assigned to ASVs by using a pre-trained naïve Bayes taxonomic classifier against the Greengenes 13_8 99% operational taxonomic unit (OTU) reference sequences.
2.9. Crude Mucin Determination
The ethanol precipitation method was used for crude mucin determination, as described by Leterme et al. [23] and Romero et al. [24], and using pectinase (Sigma P2401) to remove soluble fiber, as described by Abad et al. [25]. One gram of ileal content from each rabbit was used for the procedure.
2.10. Quantitative PCR Analysis of Gene Expression
The intestinal samples were homogenized in 1 mL of NZYol reagent (Nzytech, Lisboa, Portugal) using a TissueLyser II (Qiagen, Hilden, Germany) homogenizer with two 3 mm zirconia beads per sample for two cycles of 1 min at 30 Hz, and 1 min resting in between. After the total RNA extraction using NZYol reagent contents (Nzytech, Lisboa, Portugal), the sample was treated with DNase to remove the genomic DNA. The RNA integrity and quantity were assessed using a Bioanalyzer (Bonsai Technologies, Madrid, Spain) and a NanoDrop® ND-1000 spectrophotometer (NanoDrop® Technologies Inc., Wilmington, DL, USA), respectively. The RNA obtained was then stored at −80 °C for future analysis.
Quantitative PCR (Q-PCR) was performed to estimate the level of expression of genes related to gut immunity (IL-6, IL-10, TNF- α) and tight junction proteins (CLDN-1, OCLN, JAM-2, ZO-1), with glyceraldehyde-3-phosphate dehydrogenase (gapdh) as the housekeeping gene. The total RNA was reverse transcribed into cDNA with the HiScript II 1st strand cDNA Synthesis kit (NeoBiotech, Seoul, Republic of Korea) and qPCR was conducted in a MX3005P Stratagene using AceQ Universal SYBR qPCR Master Mix (NeoBiotech, Seoul, Republic of Korea). All of the genes and their primers are listed in Table 3. All reactions were run in triplicate. The relative expression of gene mRNA was determined by using the 2−Ct method. GAPDH was used to normalize expression of the targeted genes, and the control group was used as the calibration group.
Table 3.
List of genes, accession numbers, primer sequences and product sizes.
2.11. Statistical Analysis
The effect of including the macroalgae products in the rabbit diet on the growth and villous heigh, crypt depth, ileal mucin concentration and gene expression were analyzed by one-way ANOVA, with diet as the main effect. The initial mean weight of the litter was included as a covariate for growth traits. The residues of the model for each trait were analyzed to confirm they were normally distributed (Shapiro–Wilk test), and the homoscedasticity was evaluated from the graph of residue distribution and confirmed with Levene’s test. The variables met the homoscedasticity and normality requirements. Mortality and number of rabbits with growth, overgrowth or enteropathogenic E. coli were analyzed using a logistic model (considering a binomial distribution) with diet as the main effect, and the results were transformed from the logit scale.
Both coccidia and E. coli counts were analyzed using a Poisson regression, with diet as the main effect, and the results were transformed from the logarithmic scale. The control group value and the average values of the other treatments were compared by ANOVA and a post hoc Tukey test. A significance level of α = 0.05 was applied, and it was considered a trend when 0.05 ≤ α ≤ 0.15.
STAMP software (v 2.1.3) for the “Statistical Analysis of Taxonomic and Functional Profiles” [29] was used to determine statistical differences (p < 0.05) in the ASVs obtained for each test group, by applying a Kruskal–Wallis H test and a post hoc Tukey–Kramer test.
3. Results
3.1. Growth Performance and Mortality
Growth was similar in the groups of rabbits fed the macroalgae-supplemented diets and the controls, and no significant differences were observed when variables were compared across groups (Table 4). Nevertheless, the comparison between the control group and the average of all macroalgae treatments revealed some differences. Between 33 and 54 days of age, the average FCR of rabbits fed with all macroalgae diets tended to improve relative to the control group (2.62 vs. 2.49, p = 0.065). Between 54 and 61 days of age, ADG was lower in the average of the three experimental groups than in the control group (47.3 vs. 44.2, p = 0.038). Nevertheless, the final weight at 61 days of age was very similar in all groups. There were no significant differences in mortality among groups.
Table 4.
Effect of macroalgae treatments on growth performance. Mortality rates include estimated means transformed from the logit scale or untransformed with the original standard error.
3.2. Effects of Macroalgae on Gut Health Parameters
3.2.1. Populations of Selected Bacterial Species
No discrepancies were observed between the results of colon and ileocecal swabs obtained from the same animal with regard to E. coli counts, the presence of EPEC or MDR. Thus, for 20 of 40 rabbits, growth of E. coli was not observed on the 3M Petrifilm™ SEC plates (<10 cfu/g) or on ML plates. Additionally, the growth on the 3M Petrifilm™ SEC plates from the intestinal content of one rabbit yielded a negative result for E. coli identification. The study of susceptibility included the eae positive isolates and one representative E. coli from ML plates for a total of 19 animals. As a result, 12 rabbits were identified as carriers of MDR E. coli. No colonies were recovered from the CHROMID®ESB or CHROMID®CARBA culture media used for screening extended-spectrum beta-lactamases (ESBL) and carbapenemase-producing E. coli, respectively.
E. coli was detected in samples from rabbits in all groups (Table 5). However, growth of the bacterium was highly variable, with abnormal E. coli counts (<10 cfu/g or >108) for ≥75% of the samples within all the diet groups (SL, U, EU). Thus, no colonies were detected in SEC plates for some samples (62.5, 37.5 and 25% in SL, U and EU diets, respectively), while an overgrowth of the bacterium was observed in others (100, 87.5 and 87.5 in SL, U and EU diets, respectively). The eae gene associated with EPEC was determined by PCR in four animals, and neither of the two Clostridium species studied were detected in any of the animals. There were no significant differences among groups for these variables.
Table 5.
Effect of macroalgae treatments on microbial populations (counts and percentage of animals in each group in which each variable was detected. Estimated means transformed from the logarithmic or logit scale or untransformed with the original standard error) in colon samples from rabbits of age 47 d.
3.2.2. Coccidia
Most rabbits had mixed infections with Eimeria spp., in some cases reaching remarkably high levels. No significant differences were observed among experimental groups, although the values tended to be higher in the EU group than in the SL group (Table 6, p = 0.10). The coccidia counts were lowest in the SL group in almost all rabbits, while the highest values corresponded to the EU group, with counts of more than 20,000 opg reached in many samples.
Table 6.
Effect of macroalgae treatments on the recount of coccidia in feces of rabbits at 47 d of age (transformed from the logarithmic scale or untransformed with the original standard error).
3.2.3. Histological Examination, Morphometric Study and Crude Mucin Content in Ileum
Along with the coccidia detected in the rabbit feces, Eimeria spp. were found in some samples from different locations, mostly concentrated in the distal half of the small intestine (jejunum, ileum) and the caecum. Different stages of the life cycle of the species (schizonts, gametocytes and oocysts) were found invading the enterocytes (Figure 1). In the areas affected by coccidia, destruction of enterocytes and villous atrophy were observed, and a variable number of leukocytes (predominantly heterophils) infiltrated the lamina propria. The sections affected by coccidia appeared to be more susceptible to proliferation of bacteria and invasion of damaged mucosa, as the apical part of the villi was eroded and had bacteria adhering to their surface (effacing-attaching lesion) or even penetrating the brush border of the enterocytes and appearing in the cytoplasm.
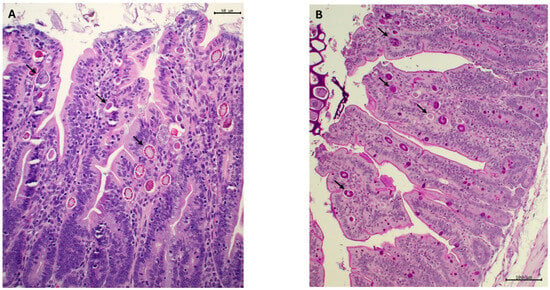
Figure 1.
Sections of ileal mucosa of rabbits at 47 d of age, stained with hematoxylin-eosin (A) and periodic acid–Schiff, PAS (B). (A) The epithelium of the villi has a scalloped appearance. The apical edge is eroded, and remnants of partially digested cellular and food debris appear in the lumen. The epithelial lining shows multiple developmental stages of coccidia (arrows), which are intensely marked by PAS staining ((B), arrows). Bars: 50 µm (A) and 100 µm (B).
The effacing-attaching lesion associated with bacterial proliferation were observed in many of the samples (Figure 2), mainly in the jejunum, ileum, caecum and colon. Large numbers of bacteria were observed beside or attached to enterocytes, and in some cases, they colonized the surface of the epithelium. In many cases, bacteria had invaded enterocytes and colonized deeper locations of the mucosa, causing a large loss in villi area. In the affected areas, the mucosal architecture was highly altered, with destruction of enterocytes and effacement of the brush border. The lamina propria was infiltrated with leukocytes, predominantly heterophils and lymphocytes. Hypertrophy of caliciform cells was also observed in some sections.
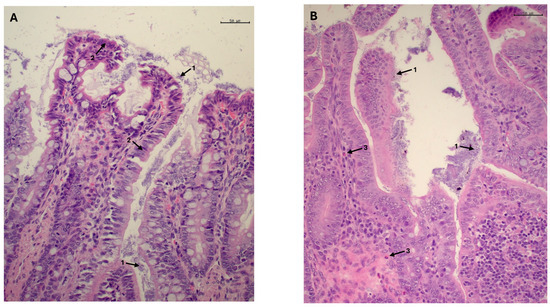
Figure 2.
Mucosa of the ileocecal valve of rabbits at age 47 days. Numerous coccobacilli are observed adhering to the epithelial surface of the enterocytes, both at the apical portion of the villi and in deeper regions (arrow 1). In some areas, the epithelial cells remain intact; however, in others, the enterocyte exhibit a cuboidal, swollen or deformed appearance (arrow 2), resulting in the epithelial border having a scalloped appearance (A). In these regions, the lamina propria exhibits a mixed population of heterophils, lymphocytes and plasma cells ((B), arrow 3). Bars: 50 µm (A,B).
Significant differences between experimental groups were observed in relation to the morphometric measures of the jejunal mucosa (Table 7). In the rabbits in the SL group, villi height (p < 0.001) and crypt depth (p < 0.001) were both greater than in the other algae groups, and the control group showed an intermediate value between the SL and both EU and U groups. Villi height and crypt depth were lowest in the EU group. The treatments did not affect the VH:CD ratio (p = 0.55).
Table 7.
Effect of macroalgae treatments on villi height and crypt depth (µm) of jejunal mucosa, and ileal crude mucin concentration in rabbits at 47 d of age. Different letters indicate statistically significant differences.
No differences (p = 0.67) between experimental groups were observed regarding crude mucin concentration (Table 7).
3.2.4. Microbiome
The colon microbiota was dominated by Firmicutes (60%) and Bacteroidetes (25%) with no difference between treatment groups. The results obtained from the analysis of 16S RNA amplicon sequencing showed significant differences for Cyanobacteria (p = 0.04) and TM7 (Saccharibacteria) (p = 0.007) at the phylum level (Figure 3A,B). For both phyla, the relative abundance was lower in the macroalgae treatments than in the control. Cyanobacteria occurred at lowest levels in the EU and U groups, whereas the lowest levels of TM7 were found in the SL group. Regarding family level, there were some significant differences (Figure 3C–E) in the families p_Cyanobacteria; c_4C0d-2; o_YS2; f_ (p = 0.04), p_TM7; c_TM7-3; o__CW040; f__F16 (p = 0.007) and p_Bacteroidetes; c_Flavobacteriia; o_Flavobacteriales; and f_Cryomorphaceae (p = 0.04). Within the latter family, significant differences (p = 0.008) were also found at the genus level for Fluviicula (Figure 3F), which were higher in the microbiota of rabbits in the U and EU groups than in the control group.
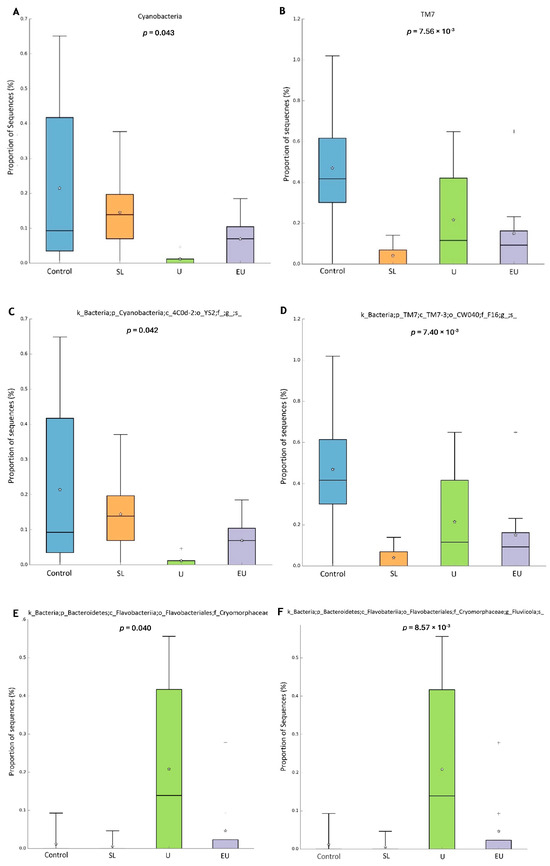
Figure 3.
Significant differences in colon microbiota between experimental groups of rabbits at 47 days of age. (A) Phylum Cyanobacteria. (B) Phylum TM7. (C) Family YS2. (D) Family TM7-F16. (E) Family Cryomorphaceae. (F) Genus Fluviicula. SL: dehydrated S. latissima. U: dehydrated Ulva spp. EU: hydrolyzed aqueous extract of Ulva lactuca.
3.2.5. Immune-Related and Tight Junction Protein Gene Expression in Cecal Appendix
Regarding interleukins (IL-6 and IL-10) and TNF-α, there were no significant differences (p > 0.05) among experimental groups (Figure 4).
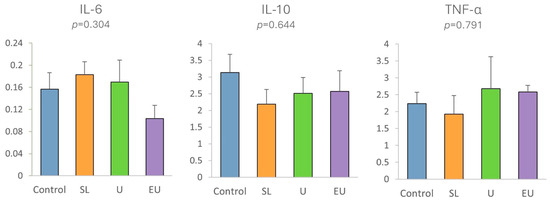
Figure 4.
Gene expression in cecal appendix mucosa of rabbits at 47 days of age, of interleukins (IL-6, IL-10) and TNF-α in experimental diets. SL: dehydrated S. latissima. U: dehydrated Ulva spp. EU: extract of Ulva lactuca.
In the case of tight junction proteins (Figure 5), significant differences were only observed in the case of claudins-1 (p < 0.001), as expression was lowest in the SL group and highest in the U group. Although not statistically significant, the expression of ZO-1 genes tended to be higher (p = 0.11) in all macroalgae-supplemented groups than in the control group, especially in the U and EU groups.
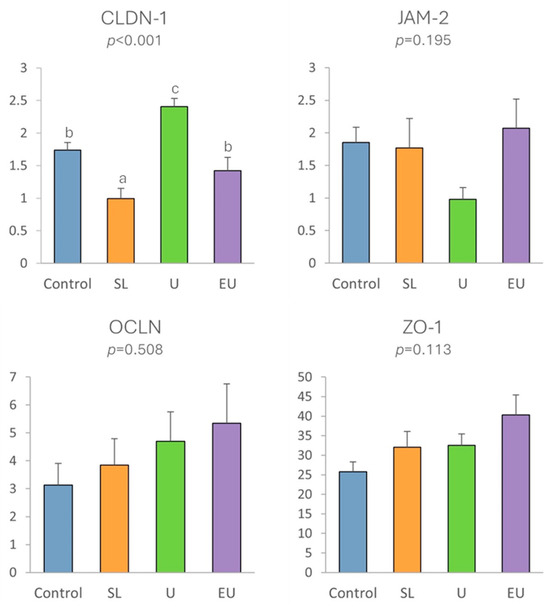
Figure 5.
Expression in cecal appendix mucosa of tight junction proteins in relation to experimental diets. SL: dehydrated S. latissima. U: dehydrated Ulva spp. EU: hydrolyzed aqueous extract of Ulva lactuca. Different letters indicate statistically significant differences.
4. Discussion
The study findings indicate that the inclusion of Saccharina latissima (dehydrated) and Ulva lactuca (dehydrated or aqueous extract) in the rabbit diets yielded favorable results. As a result, at the end of the production cycle, the final weights of the rabbits were similar to those of the control group. Additionally, the slightly enhanced conversion ratio suggests improved utilization of the feed. These results may be related to the improvement in digestibility, as observed for the same macroalgae when included at 1.025% in rabbit diets [11]. Similar results were reported for rabbits fed a diet supplemented with 0.3% Laminaria spp. and plant polyphenols [9], as this diet improved both ADG and FCR because it reduced feed intake but the rabbits reached a higher final weight. The addition of 1% Ulva spp. to the rabbit diet improved ADG and FC [8]. In post-weaning piglets, the inclusion of laminarin at 0.03% in diets also improved ADG and G:F ratio [7,15,30], as well as energy and protein digestibility [7,15]. The improved digestibility may be linked to a better functionality of the intestinal mucosa due to the enhanced nutrient transport [15] and the mucosal morphology of the duodenum [31].
As often occurs in commercial rabbit husbandry, an outbreak of enteric disease emerged during the post-weaning phase in the animals in the present study. Most rabbits were naturally infected with Eimeria spp., with high numbers of oocysts per gram of feces in some (>10,000 opg is considered pathological [32]). The inclusion of SL in the diet appeared to lower the incidence of Eimeria spp. relative to the EU group, while values were intermediate in the control group. The presence of coccidia in rabbits post-weaning can be used as a marker of health and immune status. In some cases of acute coccidiosis, this parasite can act as a primary pathogen causing clinical disease, especially in the post-weaning phase, which is a critical period as rabbits are particularly sensitive to digestive disorders [33,34]. Nevertheless, proliferation of the epithelial coccidia in the intestinal mucosa often produces sub-clinical infections, which are linked to poorer growth and a higher risk of infection with other pathogens. In these cases, coccidia cause gastrointestinal dysbiosis, which can weaken rabbits during stressful periods such as weaning.
This imbalance is often accompanied by the appearance of opportunistic pathogens, which can lead to more severe infections, such as colibacillosis and clostridia. E. coli is a usual member of the rabbit digestive microbiota, but stress or other pathogens like coccidia may trigger enhanced growth in the gut, in some cases, resulting in death [35]. In fact, colibacillosis is considered the most important enteric disease in rabbits, and it is more common in the early weaning period than in the suckling period and may also occur in adults, probably associated with stress at weaning and change in diet leading to an increase in cecal pH [1,35,36]. At the same time, when these bacterial processes appear, coccidia also propagate and complicate the entire process [37,38]. In this case, rabbits appeared to suffer concomitant infections involving coccidia and E. coli, although the presence of other pathogens cannot be ruled out. Both processes altered the mucosal architecture and protection against other pathogens, which could lead to a multifactorial dysbiosis that is more difficult to control and diagnose. In this regard, the rabbits exhibited abnormal levels of E. coli, considering the usual low E. coli counts in rabbit intestine, with numerous samples either showing no growth or displaying an overgrowth exceeding normal levels. These results can be explained by the fact that rabbits were treated with antibiotics, which may have led to a sharp reduction in bacterial abundance or to dysbiosis. In any case, bacterial counts were high in many samples, indicating the existence of a digestive pathology and dysbiosis, which can be at least partly linked to E. coli proliferation. The effacing-attaching lesions observed in the histological examination were also compatible with overgrowth of this bacterium.
By contrast, the Clostridium species studied were not found in colon samples, and therefore these particular species may not have been involved in the enteric disease observed in the trial. Nevertheless, other Clostridium species (such as C. spiroforme) can affect rabbits, and isolation of the species is not always possible, even in rabbits with clinical symptoms compatible with Clostridium [1]. The results obtained do not allow correct comparison between experimental groups regarding these gut health variables. The previous studies in piglets have observed a reduction in colon populations of E. coli when laminarin was included in the feed [6,31,39]. Nevertheless, the macroalgae products studied in this trial did not directly inhibit E. coli growth when tested in vitro [10], and also, no influence was observed in the variation in the E. coli populations in vivo [5].
Rabbits fed the SL diet exhibited improvements in the morphometric indices of gut health relative to the control group, specifically, a statistically significant increase in crypt depth and numerical increase in villi height. Conversely, this positive effect was not observed in rabbits fed the diets supplemented with U or EU, as the values of the measures were lower than in the controls, particularly in the villi height in the EU group. Similar results to those observed for the SL diet were obtained when piglets in the post-weaning phase were fed laminarin and/or fucoidan, which increased villi height and VFA concentration [31], or Ecklonia cava, which increased villi height in the ileum [39]. Our findings can also be compared with those of other research carried out with these macroalgae in rabbits. For example, S. latissima was highly fermented in vitro by cecal microbiota [10], and it also improved the digestibility of dry matter and crude protein [11], which may be linked to the improved status and altered morphology of intestinal mucosa. Altogether, these results may indicate that the addition of S. latissima to the rabbit diet enhanced the functionality of the intestinal mucosa, which would protect the gut mucosa against some pathogens like coccidia [4], though changes in the intestinal microbiota profile or in the end products of microbial fermentation may occur, as observed in vitro [10]. These results are consistent with the previous findings and suggest that some polysaccharides in S. latissima—such as laminarin and fucoidan—exert a positive effect on gut health, as already suggested [4,5,6].
The intestinal microbiome plays a pivotal role in maintaining intestinal health, and although the research is at an early stage, modulation of the microbiome through diet has been suggested to be important to reduce the incidence of digestive disorders in livestock [40]. The results of the gut microbiome in the animals from our study should be interpreted with caution, as all groups (including the control) had to receive an antibiotic due to intestinal dysbiosis.
The available information on the intestinal microbiome in rabbits is quite limited; however, our results revealed a characteristic composition consistent with previous descriptions by other authors [41,42]. When analyzing the impact of macroalgae supplementation on the colon microbiota, notable findings emerged, especially concerning the TM7 and cyanobacteria populations. TM7 is a subgroup of Gram-positive uncultivable bacteria associated with oral inflammation [43]. The TM7 bacterial division was originally described in natural environmental habitats [43,44,45,46], but it is also found in the feces of, e.g., mice. In humans, TM7 has been detected in subgingival plaque, and studies showed an association of the TM7 bacterial division with inflammatory mucosal diseases [43,46]. The TM7 division of bacteria has been suggested to play an important role in the early stages of inflammatory mucosal processes, probably by modifying growth conditions for competing bacteria [47]. The inclusion of macroalgae in rabbit feed could, therefore, reduce the appearance of inflammatory processes at the gut level, as the TM7 division was less abundant in all macroalgae-supplemented groups, especially in SL and EU, than in the control group.
On the other hand, gut cyanobacteria research is scarce [48]. The studies carried out to date regarding the presence of cyanobacteria in gut microbiota indicate significant differences in abundance between healthy control and various disease groups, implying that the presence of these organisms may be related to neurodevelopment, neurodegeneration and obesity and also to gastrointestinal, respiratory and human eye diseases [49,50,51]. The available studies provide preliminary insights into the association between these microorganisms and certain diseases, but the evidence is only correlative. It remains to be elucidated whether gut cyanobacteria can contribute to any disease or whether changes in abundance occur as a result of the disorder. Further research aiming to identify gut cyanobacteria species, culture them in vitro and characterize them at molecular, biochemical and physiological levels is encouraged in order to clarify the role of these microorganisms in health and disease development [48]. In the case of Cryomorphaceae, we did not find any studies on rabbits. The few relevant human studies indicate that the abundance of the genus Flavobacterium is associated with individuals of normal weight, whereas populations were less abundant in obese individuals [52]. More studies are necessary to clarify whether an increase in the abundance of this genus in rabbits receiving U or UE supplements can improve health status.
Regulation of the immune response, encompassing the production of mucins, interleukins, tight junction proteins and other mediators like pro and anti-inflammatory cytokines, emerges as another critical aspect, alongside the intestinal microbiota, in preserving intestinal health [53]. These factors play important roles in maintaining the integrity of the intestinal barrier, modulating inflammation and orchestrating the local immune response, thereby substantially contributing to the balance and homeostasis of the gastrointestinal tract. In the present study, the inclusion of macroalgae did not affect ileal mucin concentration, which provides protection against pathogens by trapping them and creating a barrier that hinders their entry into enterocytes; it also did not significantly influence the expression of the pro-inflammatory (IL-6 and TNF-α) or anti-inflammatory (IL-10) cytokines in the cecal appendix.
However, a significant effect on the expression of tight junction proteins was observed. The expression of claudin-1 was highest in the rabbits in group U, while that of zonula occludens tended to be higher in both the U and EU groups than in the control group. These tight junction proteins play a crucial role in regulating the permeability of the lateral unions between enterocytes, and higher levels of expression indicate a stronger seal and lower permeability of the mucosa, resulting in a strengthened mucosal barrier against pathogens and toxins [26,28,54,55]. In humans, it has been demonstrated that the presence of some dietary components, as well as some commensal and probiotic strains, leads to an increase in tight junction protein expression at the cell boundaries and in some cases prevents or reverses the adverse effects of pathogens [54]. In our study, the higher level of expression of these tight junction proteins in rabbits fed Ulva lactuca may indicate a potential mechanism of action of this macroalgae, even though the improvement in the intestinal barrier was insufficient to maintain gut health during infection, as the worst results regarding coccidia counts and morphometric measures of jejunal mucosa were obtained in the rabbits in these groups. To our knowledge, this potential mechanism of action of Ulva lactuca has not previously been assessed in rabbits, but similar results have been observed in weanling piglets fed with 0.1–0.5% of an extract derived from another green macroalgae—Blidingia spp.—which increased the expression of CLDN-1, OCLN and ZO-1 [56].
Our study has a limitation: the animals experienced digestive issues that necessitated antibiotic treatment. This treatment could have influenced the composition of the intestinal microbiota and the markers of gut health. However, such occurrences are very common during the weaning period in commercial rabbit farms in Spain and worldwide, and consequently, our study reflects the real-world conditions under which algae supplementation would be used. Despite these challenges, our results are intriguing as they highlight the potential of macroalgae to modulate intestinal flora and improve gut health. Moving forward, further in vivo studies with a reduced incidence of digestive diseases, without antibiotic interference, and potentially exploring combinations of macroalgae in diets are essential to elucidate their mechanisms of action and their full potential for enhancing intestinal health in rabbits.
5. Conclusions
In conclusion, the present study, conducted in a commercial rabbit farm system, demonstrates the potential of macroalgae to improve the intestinal health of rabbits. Inclusion of Saccharina latissima in the rabbit diet had a positive effect on reducing coccidia counts and improving intestinal mucosal morphology, and Ulva lactuca revealed positive effects on the expression of gut tight junction proteins, associated with improved intestinal barrier function. Both effects have been previously described in other farmed species. Further in vivo studies conducted with a minor incidence of digestive diseases, in the absence of antibiotic treatment, and possibly combining both macroalgae in the diets are necessary to elucidate the mechanisms of action and their potential for improving intestinal health in rabbits.
Author Contributions
Conceptualization, J.G., M.M., M.L.-A. and A.P.L.; methodology, S.A.-S., J.G., A.P.L., A.C.-C., A.M. (Azucena Mora), A.L., R.P. and M.L.-A.; formal analysis, S.A.-S., J.G. and M.L.-A.; investigation, S.A-S., J.G., A.P.L., A.C.-C., A.M. (Azucena Mora), A.L., R.P., M.M. and M.L.-A.; resources, J.G., A.M. (Antonio Muíños), E.C. and M.L.-A.; data curation, S.A.-S., J.G., A.P.L., A.C.-C., A.M. (Azucena Mora), A.L., R.P. and M.L.-A.; writing—original draft preparation, S.A.-S., J.G., A.P.L., A.C.-C., M.M. and M.L.-A.; writing—review and editing, S.A.-S., J.G., A.P.L., A.C.-C., A.M. (Azucena Mora), A.L., R.P., M.M. and M.L.-A.; supervision, J.G., A.P.L. and M.L.-A.; project administration, J.G., A.M. (Antonio Muíños), E.C. and M.L.-A.; funding acquisition, J.G., A.M. (Antonio Muíños), E.C. and M.L.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was conducted as part of the TIRAC innovation project, which received 80% of its funding from the European Agricultural Fund for Rural Development (EAFRD) of the European Union and 20% from the Ministry of Agriculture, Fisheries and Food, under the National Rural Development Program 2014–2020. The General Directorate for Rural Development, Innovation and Agrifood Training (DGDRIFA) administers this support. The project had a budget of EUR 492,580.38, with a total grant amount of EUR 485,043.58. The funding reference is 2020-PN216 (O00000226e2000044671). Sabela Al-Soufi, Marta Miranda and Marta López-Alonso are affiliated with the “Grupo de Potencial Crecimiento,” which is funded by GAIN (Axencia Galega de Innovación) under grant number ED431B 2023/008. Additionally, this research was supported by Project PID2022-143041OB-100, funded by MICIU/AEI/10.13039/501100011033 and ERDF/EU, and grant ID ED431C 2021/11.
Institutional Review Board Statement
All experimental procedures adhered to the Spanish regulations for the welfare and use of animals in research (Spanish Royal Decree 53/2013; BOE, 2013). These procedures received approval from the Bioethics Committee at the Universidad Politécnica de Madrid, Spain (protocol code 2021-002, approved on 24 February 2021).
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors are grateful for the use of RIAIDT-USC analytical facilities and technical support from Nerea Bermúdez and Carlos Fernández.
Conflicts of Interest
Author Antonio Muíños was employed by the company Porto Muíños S.L. Author Eugenio Cegarra was employed by the company De Heus Nutrición Animal. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Solans, L.; Arnal, J.L.; Sanz, C.; Benito, A.; Chacón, G.; Alzuguren, O.; Fernández, A.B. Rabbit Enteropathies on Commercial Farms in the Iberian Peninsula: Etiological Agents Identified in 2018–2019. Animals 2019, 9, 1142. [Google Scholar] [CrossRef] [PubMed]
- El-Ashram, S.; Aboelhadid, S.M.; Abdel-Kafy, E.S.M.; Hashem, S.A.; Mahrous, L.N.; Farghly, E.M.; Kamel, A.A. Investigation of Pre- and Post-Weaning Mortalities in Rabbits Bred in Egypt, with Reference to Parasitic and Bacterial Causes. Animals 2020, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- European Union. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. Off. J. Eur. Union 2003, L268, 29–43. [Google Scholar]
- Evans, F.D.; Critchley, A.T. Seaweeds for Animal Production Use. J. Appl. Phycol. 2014, 26, 891–899. [Google Scholar] [CrossRef]
- Sweeney, T.; O’Doherty, J.V. Marine Macroalgal Extracts to Maintain Gut Homeostasis in the Weaning Piglet. Domest. Anim. Endocrinol. 2016, 56, S84–S89. [Google Scholar] [CrossRef]
- McDonnell, P.; Figat, S.; Odoherty, J.V. The Effect of Dietary Laminarin and Fucoidan in the Diet of the Weanling Piglet on Performance, Selected Faecal Microbial Populations and Volatile Fatty Acid Concentrations. Animal 2010, 4, 579–585. [Google Scholar] [CrossRef]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of Supplementing Varying Inclusion Levels of Laminarin and Fucoidan on Growth Performance, Digestibility of Diet Components, Selected Faecal Microbial Populations and Volatile Fatty Acid Concentrations in Weaned Pigs. Anim. Feed Sci. Technol. 2013, 183, 151–159. [Google Scholar] [CrossRef]
- El-banna, S.G.; Hassan, A.A.; Okab, A.B.; Koriem, A.A.; Ayoub, M.A. Effect of Feeding Diets Supplemented with Seaweed on Growth Performance and Some Blood Hematological and Biochemical Characteristics of Male Baladi Rabbits. In Proceedings of the 4th International Conference on Rabbit Production in Hot Climates, Sharm El-Shiekh, Egypt, 24–27 February 2005; Volume 382, pp. 373–382. [Google Scholar]
- Rossi, R.; Vizzarri, F.; Chiapparini, S.; Ratti, S.; Casamassima, D.; Palazzo, M.; Corino, C. Effects of Dietary Levels of Brown Seaweeds and Plant Polyphenols on Growth and Meat Quality Parameters in Growing Rabbit. Meat Sci. 2020, 161, 107987. [Google Scholar] [CrossRef] [PubMed]
- Al-Soufi, S.; Nicodemus, N.; Carro, M.D.; López-Alonso, M.; Miranda, M.; Muíños, A.; Cegarra, E.; Vázquez-Belda, B.; Domínguez, H.; Torres, M.D.; et al. Marine Macroalgae in Rabbit Nutrition: In Vitro Digestibility, Caecal Fermentability, and Microbial Inhibitory Activity of Seven Macroalgae Species from Galicia (NW Spain). Agriculture 2023, 13, 1995. [Google Scholar] [CrossRef]
- Alfonzo, R.; Pérez, E.; Al-Soufi, S.; de la Cruz, P.; Silva, V.; Martín, L.; del Pozo, R.; Rybicka, A.; Buján, M.; Domínguez, H.; et al. Efecto de la Inclusión de Productos de Algas Sobre la Digestibilidad Fecal y Otros Parámetros Digestivos en Gazapos en Crecimiento. In Proceedings of the XLVII Symposium de Cunicultura de ASESCU, León, Spain, 31 May–1 June 2023; pp. 77–81. [Google Scholar]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-Assisted Extraction (EAE) for the Production of Antiviral and Antioxidant Extracts from the Green Seaweed Ulva Armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- de Blas, C.; Mateos, G.G. Feed Formulation. In Nutrition of the Rabbit; CABI: Wallingford, UK, 2020. [Google Scholar]
- Ocasio-Vega, C.; Delgado, R.; Abad-Guamán, R.; Carabaño, R.; Carro, M.D.; Menoyo, D.; García, J. The Effect of Cellobiose on the Health Status of Growing Rabbits Depends on the Dietary Level of Soluble Fiber. J. Anim. Sci. 2018, 96, 1806–1817. [Google Scholar] [CrossRef] [PubMed]
- Heim, G.; Walsh, A.M.; Sweeney, T.; Doyle, D.N.; O’Shea, C.J.; Ryan, M.T.; O’Doherty, J.V. Effect of Seaweed-Derived Laminarin and Fucoidan and Zinc Oxide on Gut Morphology, Nutrient Transporters, Nutrient Digestibility, Growth Performance and Selected Microbial Populations in Weaned Pigs. Br. J. Nutr. 2014, 111, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- MAFF. Manual de Técnicas de Laboratorio Parasitolóxico Veterinario; Boletín Técnico N° 18; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1986; pp. 2–67. [Google Scholar]
- Gómez-Duarte, O.G.; Arzuza, O.; Urbina, D.; Bai, J.; Guerra, J.; Montes, O.; Puello, M.; Mendoza, K.; Castro, G.Y. Detection of Escherichia coli Enteropathogens by Multiplex Polymerase Chain Reaction from Children’s Diarrheal Stools in Two Caribbean-Colombian Cities. Foodborne Pathog. Dis. 2010, 7, 199–206. [Google Scholar] [CrossRef]
- Díaz-Jiménez, D.; García-Meniño, I.; Herrera, A.; Lestón, L.; Mora, A. Microbiological Risk Assessment of Turkey and Chicken Meat for Consumer: Significant Differences Regarding Multidrug Resistance, Mcr or Presence of Hybrid AEPEC/ExPEC Pathotypes of E. coli. Food Control 2021, 123, 107713. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- García, V.; García-Meniño, I.; Gómez, V.; Jiménez-Orellana, M.; Méndez, A.; Aguarón, A.; Roca, E.; Mora, A. Mobile Colistin Resistance (MCR), Extended-Spectrum Beta-Lactamase (ESBL) and Multidrug Resistance Monitoring in Escherichia coli (Commensal and Pathogenic) in Pig Farming: Need of Harmonized Guidelines and Clinical Breakpoints. Front. Microbiol. 2022, 13, 1042612. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Z.; Shi, Q.; Huang, S.; Yu, T.; Zhang, L.; Yang, H. Multiplex PCR Method for Simultaneous Detection of Five Pathogenic Bacteria Closely Related to Foodborne Diseases. 3 Biotech 2021, 11, 219. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Leterme, P.; Froidmont, E.; Rossi, F.; Théwis, A. The High Water-Holding Capacity of Pea Inner Fibers Affects the Ileal Flow of Endogenous Amino Acids in Pigs. J. Agric. Food Chem. 1998, 46, 1927–1934. [Google Scholar] [CrossRef]
- Romero, C.; Nicodemus, N.; Rodríguez, J.D.; García, A.I.; de Blas, C.D. Effect of Type of Grinding of Barley and Dehydrated Alfalfa on Performance, Digestion, and Crude Mucin Ileal Concentration in Growing Rabbits. J. Anim. Sci. 2011, 89, 2472–2484. [Google Scholar] [CrossRef][Green Version]
- Abad, R.; Ibáñez, M.A.; Carabaño, R.; García, J. Quantification of Soluble Fibre in Feedstuffs for Rabbits and Evaluation of the Interference between the Determinations of Soluble Fibre and Intestinal Mucin. Anim. Feed Sci. Technol. 2013, 182, 61–70. [Google Scholar] [CrossRef]
- Li, S.; Liu, T.; Wang, K.; Li, C.; Wu, F.; Yang, X.; Zhao, M.; Chen, B.; Chen, X. The Ratios of Dietary Non-Fibrous Carbohydrate (NFC) to Neutral Detergent Fiber (NDF) Influence Intestinal Immunity of Rabbits by Regulating Gut Microbiota Composition and Metabolites. Front. Microbiol. 2023, 14, 1146787. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Menoyo, D.; Abad-Guamán, R.; Nicodemus, N.; Carabaño, R.; García, J. Effect of Dietary Soluble Fibre Level and N-6/n-3 Fatty Acid Ratio on Digestion and Health in Growing Rabbits. Anim. Feed Sci. Technol. 2019, 255, 114222. [Google Scholar] [CrossRef]
- Khalid, A.R.; Yasoob, T.B.; Zhang, Z.; Zhu, X.; Hang, S. Dietary Moringa Oleifera Leaf Powder Improves Jejunal Permeability and Digestive Function by Modulating the Microbiota Composition and Mucosal Immunity in Heat Stressed Rabbits. Environ. Sci. Pollut. Res. 2022, 29, 80952–80967. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Heim, G.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of Maternal Dietary Supplementation of Laminarin and Fucoidan, Independently or in Combination, on Pig Growth Performance and Aspects of Intestinal Health. Anim. Feed Sci. Technol. 2015, 204, 28–41. [Google Scholar] [CrossRef]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of Dietary Laminarin and Fucoidan on Selected Microbiota, Intestinal Morphology and Immune Status of the Newly Weaned Pig. Br. J. Nutr. 2013, 110, 1630–1638. [Google Scholar] [CrossRef]
- Coudert, P.; Jobert, J.L.; Larour, G.; Guittet, M. Relation Entre l’ Entéropathie Épizootique Du Lapin (EEL) et l’infestation Par Les Coccidies: Enquête Épidémiologique. In Proceedings of the 10émes Journées de la Recherche Cunicole, Paris, France, 19 November 2003; pp. 239–242. [Google Scholar]
- Fortun-Lamothe, L.; Boullier, S. A Review on the Interactions between Gut Microflora and Digestive Mucosal Immunity. Possible Ways to Improve the Health of Rabbits. Livest. Sci. 2007, 107, 1–18. [Google Scholar] [CrossRef]
- Gidenne, T.; Fortun-Lamothe, L. Feeding Strategy for Young Rabbits around Weaning: A Review of Digestive Capacity and Nutritional Needs. Anim. Sci. 2002, 75, 169–184. [Google Scholar] [CrossRef]
- Milon, A.; Oswald, E.; De Rycke, J. Rabbit EPEC: A Model for the Study of Enteropathogenic Escherichia coli. Vet. Res. 1999, 30, 203–219. [Google Scholar]
- Blanco, J.E.; Blanco, M.; Blanco, J.; Mora, A.; Balaguer, L.; Cuervo, L.; Balsalobre, C.; Muñoa, F. Prevalence and Characteristics of Enteropathogenic Escherichia coli with the Eae Gene in Diarrhoeic Rabbits. Microbiol. Immunol. 1997, 41, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Maxie, M.G. Chapter 1: Alimentary System. In Jubb, Kennedy & Palmer´s Pathology of Domestic Animals; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015; Volume 2, pp. 60–242. [Google Scholar]
- Percy, D.H.; Barthold, S.W. Chapter 6. Rabbit. In Pathology of Laboratory Rodents and Rabbits; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 253–307. [Google Scholar]
- Choi, Y.; Hosseindoust, A.; Goel, A.; Lee, S.; Jha, P.K.; Kwon, I.K.; Chae, B.J. Effects of Ecklonia Cava as Fucoidan-Rich Algae on Growth Performance, Nutrient Digestibility, Intestinal Morphology and Caecal Microflora in Weanling Pigs. Asian-Australas. J. Anim. Sci. 2017, 30, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, S.; Yan, C. Gut Microbiota Implications for Health and Welfare in Farm Animals: A Review. Animals 2022, 12, 93. [Google Scholar] [CrossRef]
- Hu, X.; Wang, F.; Yang, S.; Yuan, X.; Yang, T.; Zhou, Y.; Li, Y. Rabbit Microbiota across the Whole Body Revealed by 16S RRNA Gene Amplicon Sequencing. BMC Microbiol. 2021, 21, 312. [Google Scholar] [CrossRef]
- Combes, S.; Michelland, R.J.; Monteils, V.; Cauquil, L.; Soulié, V.; Tran, N.U.; Gidenne, T.; Fortun-Lamothe, L. Postnatal Development of the Rabbit Caecal Microbiota Composition and Activity. FEMS Microbiol. Ecol. 2011, 77, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Brinig, M.M.; Lepp, P.W.; Ouverney, C.C.; Armitage, G.C.; Relman, D.A. Prevalence of Bacteria of Division TM7 in Human Subgingival Plaque and Their Association with Disease. Appl. Environ. Microbiol. 2003, 69, 1687–1694. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Tyson, G.W.; Webb, R.I.; Wagner, A.M.; Blackall, L.L. Investigation of Candidate Division TM7, a Recently Recognized Major Lineage of the Domain Bacteria, with No Known Pure-Culture Representatives. Appl. Environ. Microbiol. 2001, 67, 411–419. [Google Scholar] [CrossRef]
- Kumar, P.S.; Griffen, A.L.; Barton, J.A.; Paster, B.J.; Moeschberger, M.L.; Leys, E.J. New Bacterial Species Associated with Chronic Periodontitis. J. Dent. Res. 2003, 82, 338–344. [Google Scholar] [CrossRef]
- Ouverney, C.C.; Armitage, G.C.; Relman, D.A. Single-Cell Enumeration of an Uncultivated TM7 Subgroup in the Human Subgingival Crevice. Appl. Environ. Microbiol. 2003, 69, 6294–6298. [Google Scholar] [CrossRef]
- Kuehbacher, T.; Rehman, A.; Lepage, P.; Hellmig, S.; Fölsch, U.R.; Schreiber, S.; Ott, S.J. Intestinal TM7 Bacterial Phylogenies in Active Inflammatory Bowel Disease. J. Med. Microbiol. 2008, 57, 1569–1576. [Google Scholar] [CrossRef]
- Hu, C.; Rzymski, P. Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life 2022, 12, 476. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse Taxa of Cyanobacteria Produce-N-Methylamino-L-Alanine, a Neurotoxic Amino Acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Sukhchuluun, G.; Bo, T.B.; Chi, Q.S.; Yang, J.J.; Chen, B.; Zhang, L.; Wang, D.H. Huddling Remodels Gut Microbiota to Reduce Energy Requirements in a Small Mammal Species during Cold Exposure. Microbiome 2018, 6, 103. [Google Scholar] [CrossRef]
- Shi, T.T.; Xin, Z.; Hua, L.; Wang, H.; Zhao, R.X.; Yang, Y.L.; Xie, R.R.; Liu, H.Y.; Yang, J.K. Comparative Assessment of Gut Microbial Composition and Function in Patients with Graves’ Disease and Graves’ Orbitopathy. J. Endocrinol. Investig. 2021, 44, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Palmas, V.; Pisanu, S.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Loviselli, A.; Manzin, A.; et al. Gut Microbiota Markers Associated with Obesity and Overweight in Italian Adults. Sci. Rep. 2021, 11, 5532. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L. Enteric Immunity: Happy Gut, Healthy Animal. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 1–18. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of Tight Junction Permeability by Intestinal Bacteria and Dietary Components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as Nutritional and Functional Food Sources: Revisiting Our Understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, B.; Xue, H.; Liu, C.; Song, W. Blidingia sp. Extracts Improve Intestinal Health and Reduce Diarrhoea in Weanling Piglets. J. Anim. Physiol. Anim. Nutr. 2023, 107, 1198–1205. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).